
Evaluation of the Virulence Potential of Listeria monocytogenes through the Characterization of the Truncated Forms of Internalin A

 , , ,
, , ,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
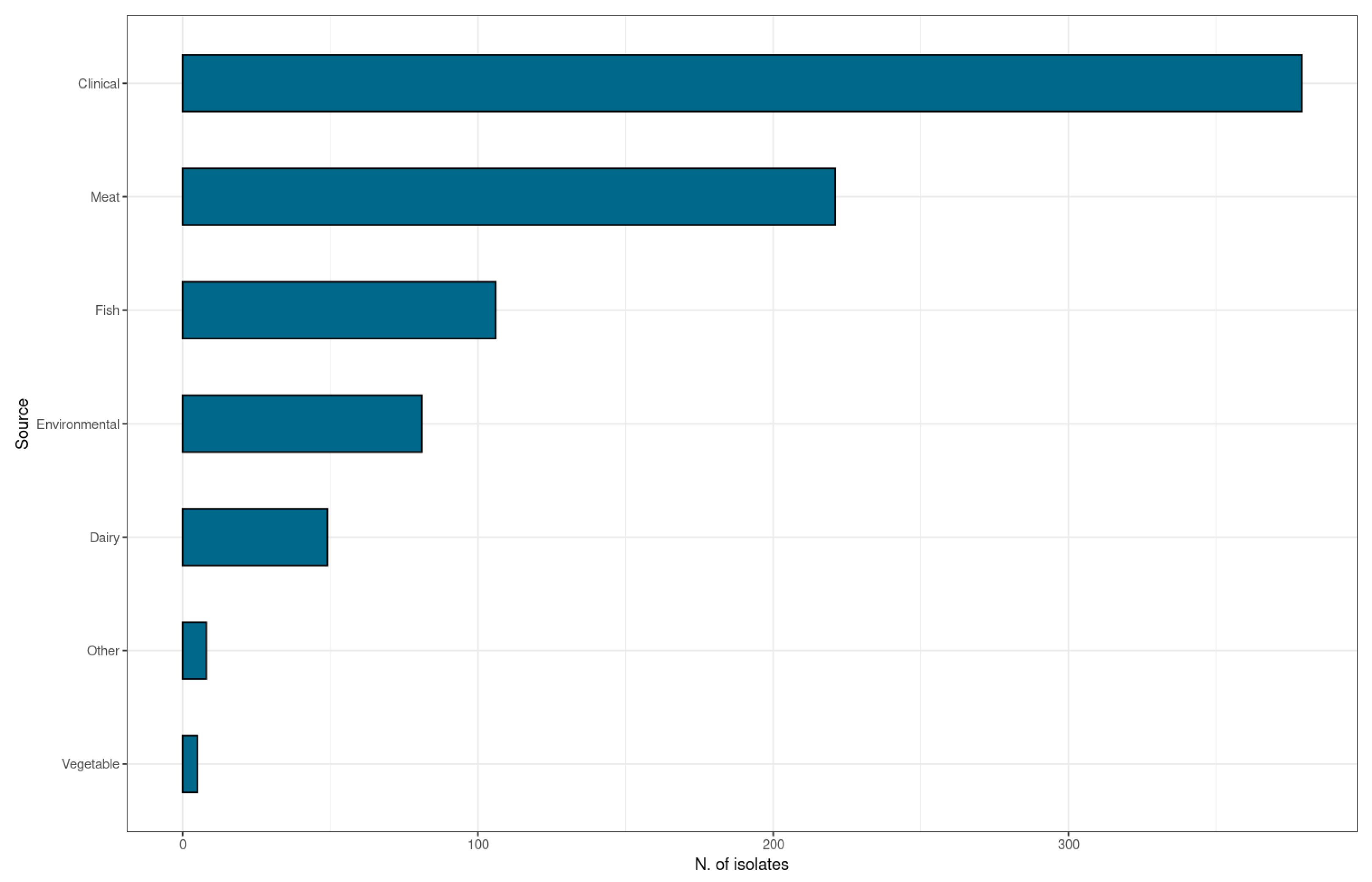
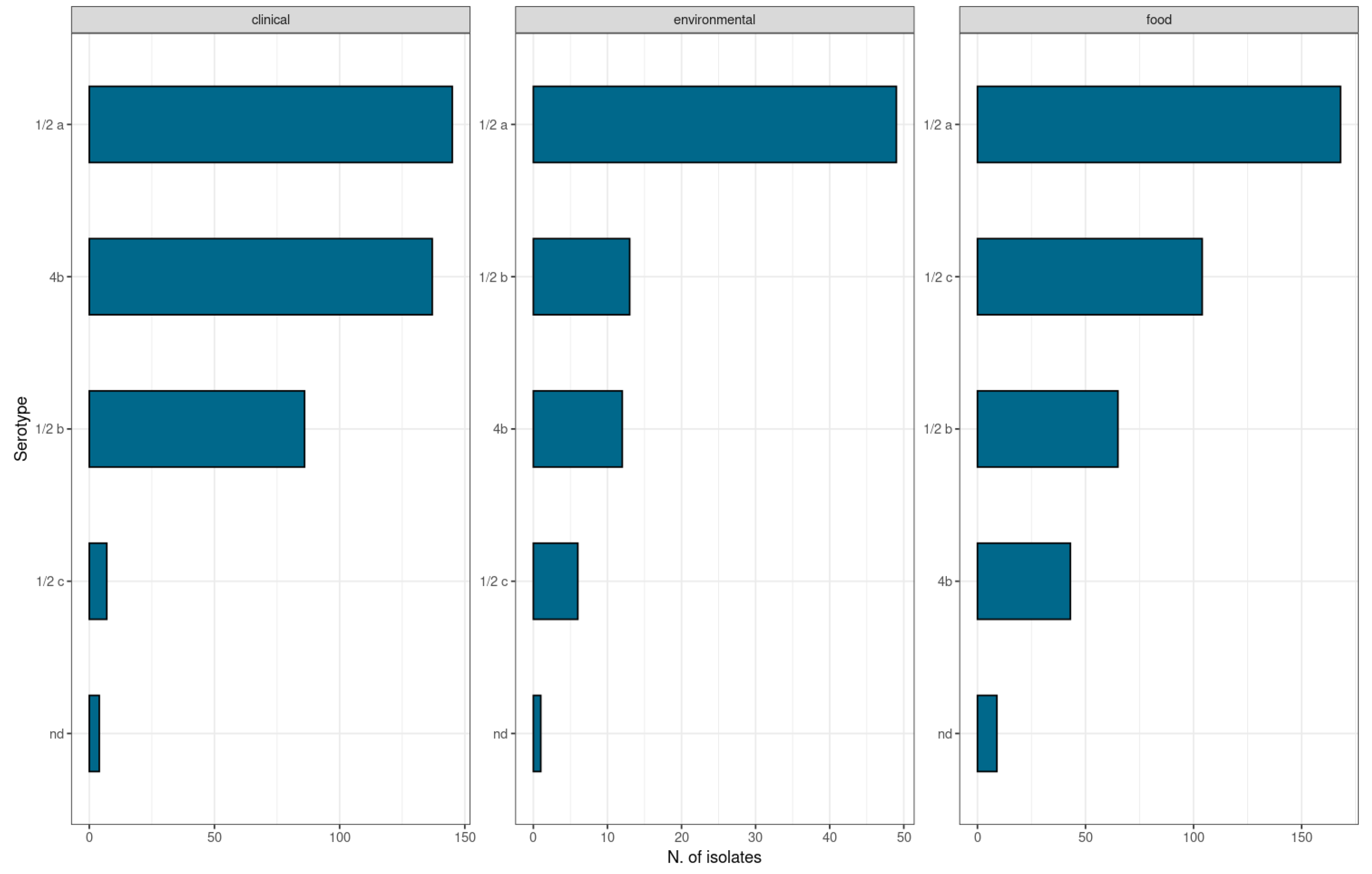
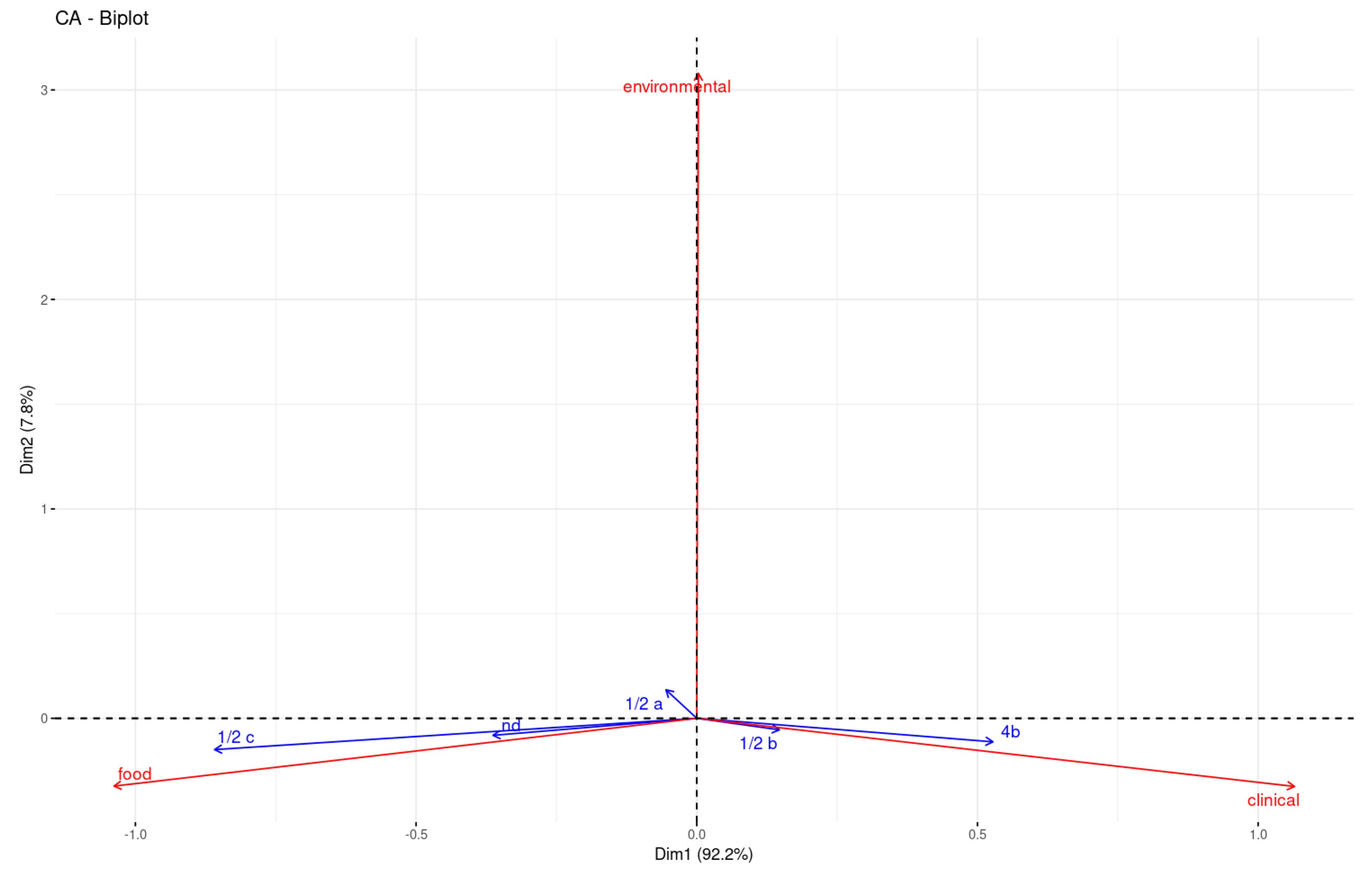
2.1. Isolates
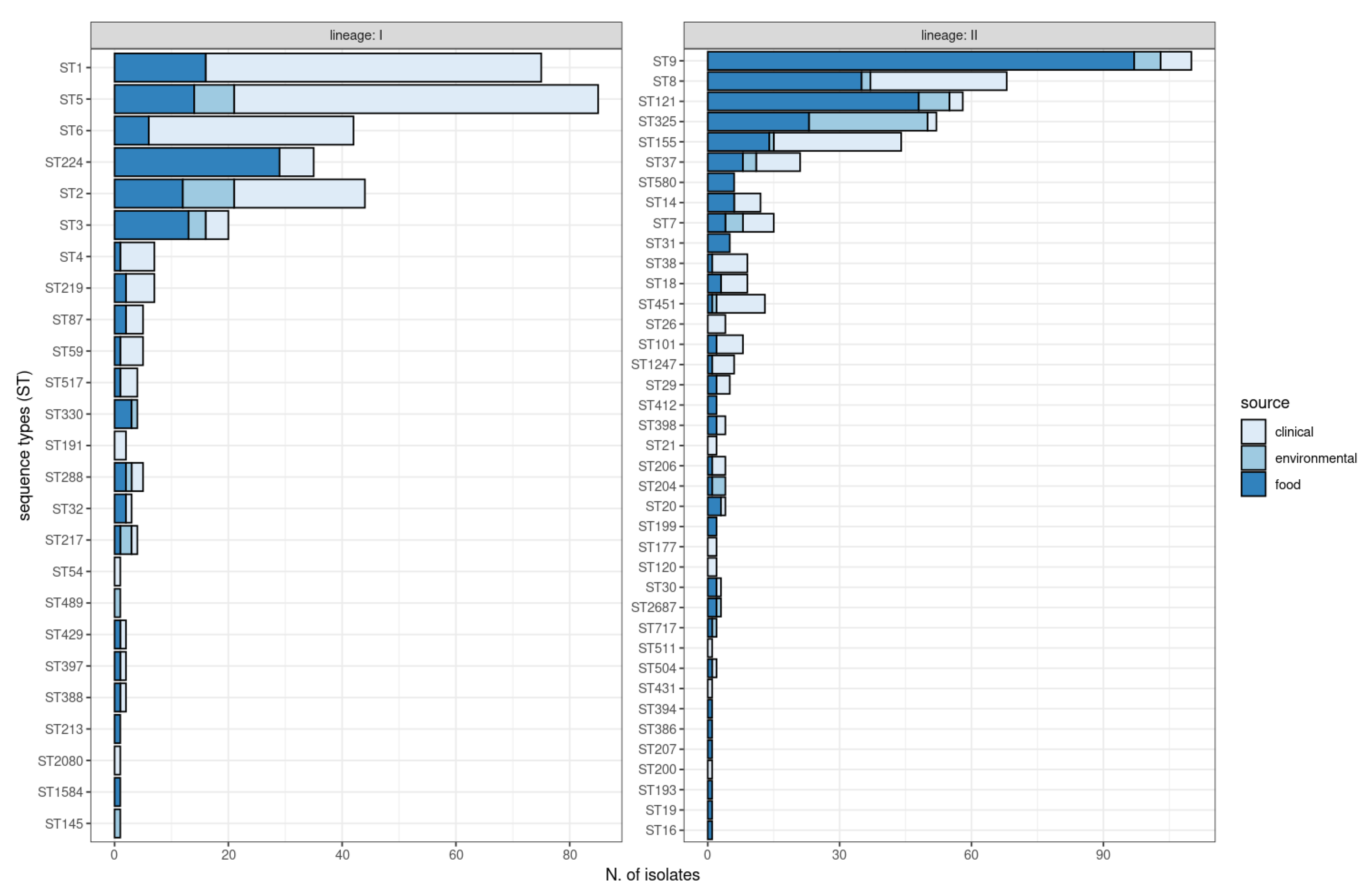
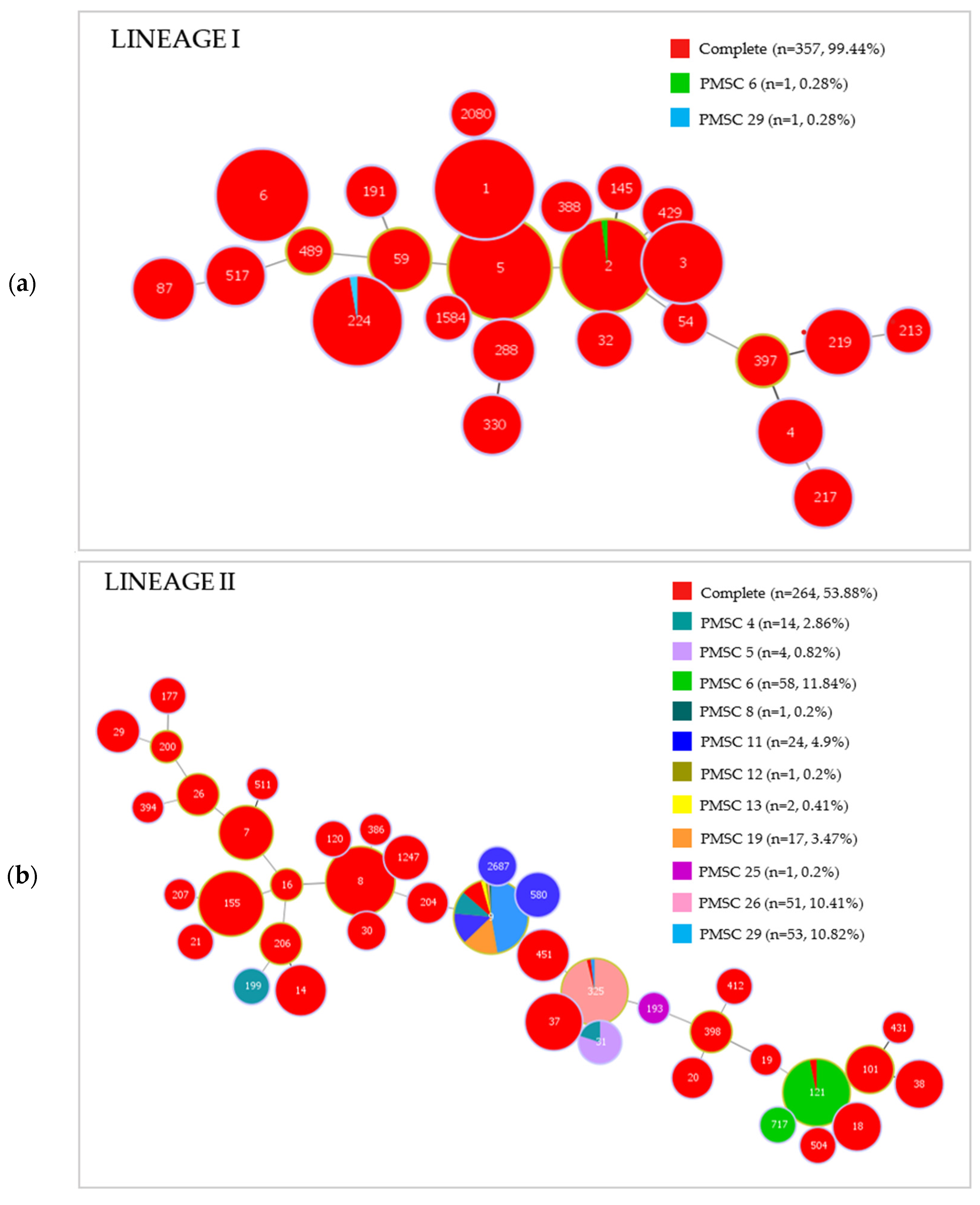
2.2. Multi-Locus Sequence Typing (MLST)
2.3. inlA Sequencing
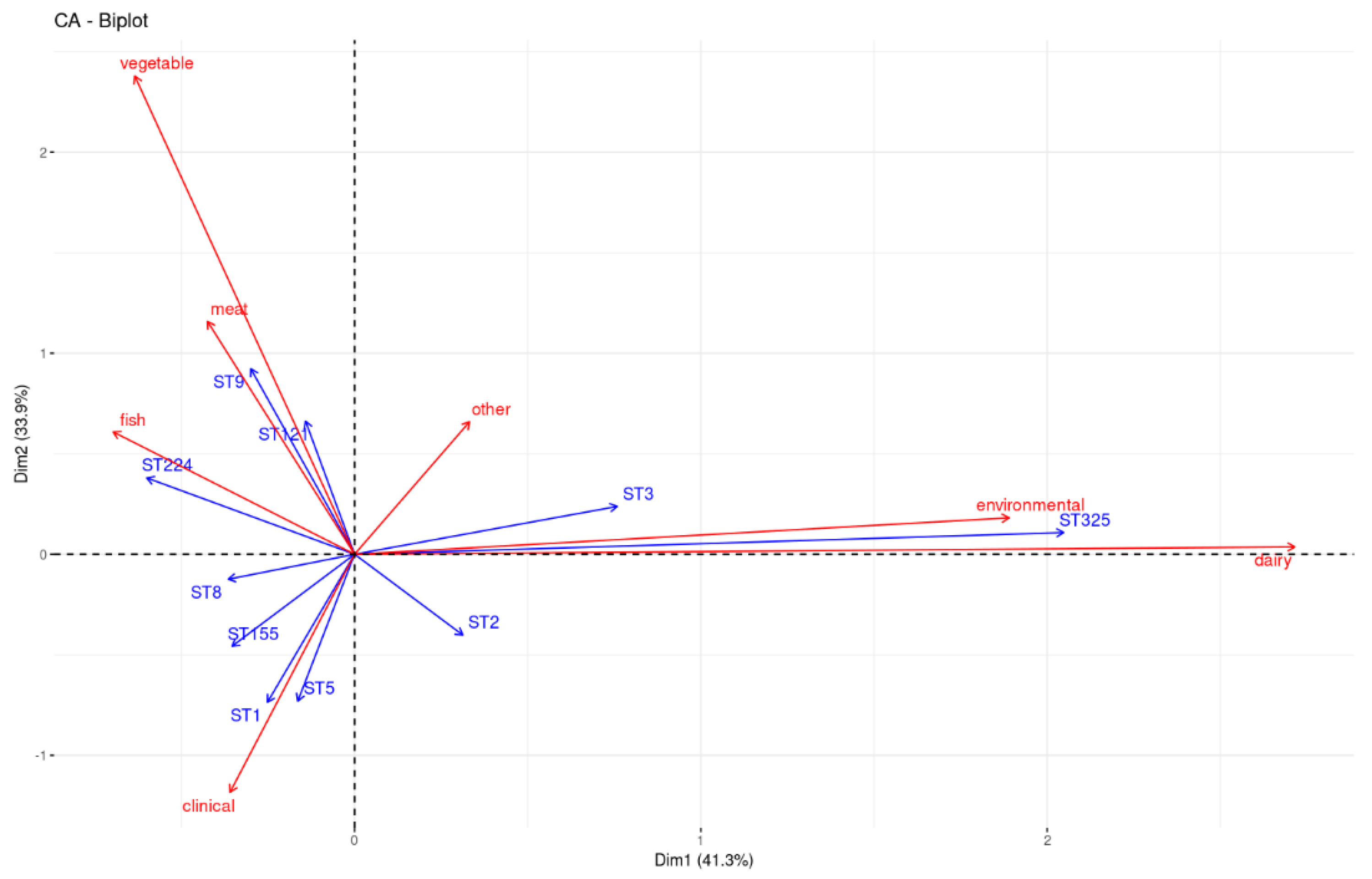
3. Discussion
4. Materials and Methods
4.1. L. Monocytogenes Isolates
4.2. Sanger Sequencing
4.2.1. DNA Extraction from L. monocytogenes Isolates
4.2.2. Multi-Locus Sequence Typing (MLST)
4.2.3. inlA Sequencing
4.3. Whole-Genome Sequencing and In Silico Analysis
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vázquez-Boland, J.A.; Kuhn, M.; Berche, P.; Chakraborty, T.; Domínguez-Bernal, G.; Goebel, W.; González-Zorn, B.; Wehland, J.; Kreft, J. Listeria Pathogenesis and Molecular Virulence Determinants. Clin. Microbiol. Rev. 2001, 14, 584–640. [Google Scholar] [CrossRef] [Green Version]
- Maury, M.M.; Bracq-Dieye, H.; Huang, L.; Vales, G.; Lavina, M.; Thouvenot, P.; Disson, O.; Leclercq, A.; Brisse, S.; Lecuit, M. Hypervirulent Listeria monocytogenes Clones’ Adaption to Mammalian Gut Accounts for Their Association with Dairy Products. Nat. Commun. 2019, 10, 2488. [Google Scholar] [CrossRef] [Green Version]
- Dreyer, M.; Aguilar-Bultet, L.; Rupp, S.; Guldimann, C.; Stephan, R.; Schock, A.; Otter, A.; Schüpbach, G.; Brisse, S.; Lecuit, M.; et al. Listeria monocytogenes Sequence Type 1 Is Predominant in Ruminant Rhombencephalitis. Sci. Rep. 2016, 6, 36419. [Google Scholar] [CrossRef] [Green Version]
- Larsen, M.H.; Koch, A.G.; Ingmer, H. Listeria monocytogenes Efficiently Invades Caco-2 Cells After Low-Temperature Storage in Broth and on Deli Meat. Foodborne Pathog. Dis. 2010, 7, 1013–1018. [Google Scholar] [CrossRef]
- EFSA Panel on Biological Hazards (BIOHAZ); Ricci, A.; Allende, A.; Bolton, D.; Chemaly, M.; Davies, R.; Fernández Escámez, P.S.; Girones, R.; Herman, L.; Koutsoumanis, K.; et al. Listeria monocytogenes Contamination of Ready-to-eat Foods and the Risk for Human Health in the EU. EFS2 2018, 16, e05134. [Google Scholar] [CrossRef]
- CDC. Listeria (Listeriosis)|Listeria. Available online: https://www.cdc.gov/listeria/index.html (accessed on 13 September 2022).
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European Union One Health 2021 Zoonoses Report. EFS2 2022, 20, e07666. [Google Scholar] [CrossRef]
- Kurpas, M.; Osek, J.; Moura, A.; Leclercq, A.; Lecuit, M.; Wieczorek, K. Genomic Characterization of Listeria monocytogenes Isolated From Ready-to-Eat Meat and Meat Processing Environments in Poland. Front. Microbiol. 2020, 11, 1412. [Google Scholar] [CrossRef]
- Ragon, M.; Wirth, T.; Hollandt, F.; Lavenir, R.; Lecuit, M.; Le Monnier, A.; Brisse, S. A New Perspective on Listeria monocytogenes Evolution. PLoS Pathog. 2008, 4, e1000146. [Google Scholar] [CrossRef] [Green Version]
- Cruz, C.D.; Pitman, A.R.; Harrow, S.A.; Fletcher, G.C. Listeria monocytogenes Associated with New Zealand Seafood Production and Clinical Cases: Unique Sequence Types, Truncated InlA, and Attenuated Invasiveness. Appl. Environ. Microbiol. 2014, 80, 1489–1497. [Google Scholar] [CrossRef] [Green Version]
- Manuel, C.S.; Van Stelten, A.; Wiedmann, M.; Nightingale, K.K.; Orsi, R.H. Prevalence and Distribution of Listeria monocytogenes InlA Alleles Prone to Phase Variation and InlA Alleles with Premature Stop Codon Mutations among Human, Food, Animal, and Environmental Isolates. Appl. Environ. Microbiol. 2015, 81, 8339–8345. [Google Scholar] [CrossRef] [Green Version]
- Nadon, C.A.; Woodward, D.L.; Young, C.; Rodgers, F.G.; Wiedmann, M. Correlations between Molecular Subtyping and Serotyping of Listeria monocytogenes. J. Clin. Microbiol. 2001, 39, 2704–2707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maury, M.M.; Tsai, Y.-H.; Charlier, C.; Touchon, M.; Chenal-Francisque, V.; Leclercq, A.; Criscuolo, A.; Gaultier, C.; Roussel, S.; Brisabois, A.; et al. Uncovering Listeria monocytogenes Hypervirulence by Harnessing Its Biodiversity. Nat. Genet. 2016, 48, 308–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira da Silva, M.; Ferreira, V.; Magalhães, R.; Almeida, G.; Alves, A.; Teixeira, P. Detection of Premature Stop Codons Leading to Truncated Internalin A among Food and Clinical Strains of Listeria monocytogenes. Food Microbiol. 2017, 63, 6–11. [Google Scholar] [CrossRef]
- Wu, S.; Wu, Q.; Zhang, J.; Chen, M.; Guo, W. Analysis of Multilocus Sequence Typing and Virulence Characterization of Listeria monocytogenes Isolates from Chinese Retail Ready-to-Eat Food. Front. Microbiol. 2016, 7, 168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacquet, C.; Doumith, M.; Gordon, J.I.; Martin, P.M.V.; Cossart, P.; Lecuit, M. A Molecular Marker for Evaluating the Pathogenic Potential of Foodborne Listeria monocytogenes. J. Infect. Dis. 2004, 189, 2094–2100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamburro, M.; Ripabelli, G.; Fanelli, I.; Maria Grasso, G.; Lucia Sammarco, M. Typing of Listeria monocytogenes Strains Isolated in Italy by Inl A Gene Characterization and Evaluation of a New Cost-Effective Approach to Antisera Selection for Serotyping. J. Appl. Microbiol. 2010, 108, 1602–1611. [Google Scholar] [CrossRef] [PubMed]
- Gelbíčová, T.; Koláčková, I.; Pantu, R. A Novel Mutation Leading to a Premature Stop Codon in InlA of Listeria monocytogenes Isolated from Neonatal Listeriosis. New Microbiol. 2015, 38, 293–296. [Google Scholar] [PubMed]
- Moura, A.; Criscuolo, A.; Pouseele, H.; Maury, M.M.; Leclercq, A.; Tarr, C.; Björkman, J.T.; Dallman, T.; Reimer, A.; Enouf, V.; et al. Whole Genome-Based Population Biology and Epidemiological Surveillance of Listeria monocytogenes. Nat. Microbiol. 2017, 2, 16185. [Google Scholar] [CrossRef] [Green Version]
- Nightingale, K.K.; Windham, K.; Martin, K.E.; Yeung, M.; Wiedmann, M. Select Listeria monocytogenes Subtypes Commonly Found in Foods Carry Distinct Nonsense Mutations in InlA, Leading to Expression of Truncated and Secreted Internalin A, and Are Associated with a Reduced Invasion Phenotype for Human Intestinal Epithelial Cells. Appl. Environ. Microbiol. 2005, 71, 8764–8772. [Google Scholar] [CrossRef] [Green Version]
- Felício, M.T.S.; Hogg, T.; Gibbs, P.; Teixeira, P.; Wiedmann, M. Recurrent and Sporadic Listeria monocytogenes Contamination in Alheiras Represents Considerable Diversity, Including Virulence-Attenuated Isolates. Appl. Environ. Microbiol. 2007, 73, 3887–3895. [Google Scholar] [CrossRef] [Green Version]
- Van Stelten, A.; Nightingale, K.K. Development and Implementation of a Multiplex Single-Nucleotide Polymorphism Genotyping Assay for Detection of Virulence-Attenuating Mutations in the Listeria monocytogenes Virulence-Associated Gene InlA. Appl. Environ. Microbiol. 2008, 74, 7365–7375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olier, M.; Pierre, F.; Rousseaux, S.; Lemaître, J.-P.; Rousset, A.; Piveteau, P.; Guzzo, J. Expression of Truncated Internalin A Is Involved in Impaired Internalization of Some Listeria monocytogenes Isolates Carried Asymptomatically by Humans. IAI 2003, 71, 1217–1224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rousseaux, S.; Olier, M.; Lemaître, J.P.; Piveteau, P.; Guzzo, J. Use of PCR-Restriction Fragment Length Polymorphism of InlA for Rapid Screening of Listeria monocytogenes Strains Deficient in the Ability to Invade Caco-2 Cells. Appl. Environ. Microbiol. 2004, 70, 2180–2185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jonquières, R.; Bierne, H.; Mengaud, J.; Cossart, P. The InlA Gene of Listeria monocytogenes LO28 Harbors a Nonsense Mutation Resulting in Release of Internalin. Infect. Immun. 1998, 66, 3420–3422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Handa-Miya, S.; Kimura, B.; Takahashi, H.; Sato, M.; Ishikawa, T.; Igarashi, K.; Fujii, T. Nonsense-Mutated InlA and PrfA Not Widely Distributed in Listeria monocytogenes Isolates from Ready-to-Eat Seafood Products in Japan. Int. J. Food Microbiol. 2007, 117, 312–318. [Google Scholar] [CrossRef]
- Van Stelten, A.; Simpson, J.M.; Ward, T.J.; Nightingale, K.K. Revelation by Single-Nucleotide Polymorphism Genotyping That Mutations Leading to a Premature Stop Codon in InlA Are Common among Listeria monocytogenes Isolates from Ready-To-Eat Foods but Not Human Listeriosis Cases. Appl. Environ. Microbiol. 2010, 76, 2783–2790. [Google Scholar] [CrossRef] [Green Version]
- Tsai, Y.-H.; Moura, A.; Gu, Z.-Q.; Chang, J.-H.; Liao, Y.-S.; Teng, R.-H.; Tseng, K.-Y.; Chang, D.-L.; Liu, W.-R.; Huang, Y.-T.; et al. Genomic Surveillance of Listeria monocytogenes in Taiwan, 2014 to 2019. Microbiol. Spectr. 2022, 10, e0182522. [Google Scholar] [CrossRef]
- Ji, S.; Song, Z.; Luo, L.; Wang, Y.; Li, L.; Mao, P.; Ye, C.; Wang, Y. Whole-Genome Sequencing Reveals Genomic Characterization of Listeria monocytogenes from Food in China. Front. Microbiol. 2022, 13, 1049843. [Google Scholar] [CrossRef]
- Knudsen, G.M.; Nielsen, J.B.; Marvig, R.L.; Ng, Y.; Worning, P.; Westh, H.; Gram, L. Genome-Wide-Analyses of Listeria monocytogenes from Food-Processing Plants Reveal Clonal Diversity and Date the Emergence of Persisting Sequence Types. Environ. Microbiol. Rep. 2017, 9, 428–440. [Google Scholar] [CrossRef] [Green Version]
- Datta, A.R.; Burall, L.S. Serotype to Genotype: The Changing Landscape of Listeriosis Outbreak Investigations. Food Microbiol. 2018, 75, 18–27. [Google Scholar] [CrossRef]
- Ulloa, S.; Arata, L.; Alarcón, P.; Araya, P.; Hormazábal, J.C.; Fernández, J. Caracterización genética de cepas de Listeria monocytogenes aisladas durante los años 2007-2014 en Chile. Rev. Chil. Infectol. 2019, 36, 585–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fagerlund, A.; Langsrud, S.; Møretrø, T. In-Depth Longitudinal Study of Listeria monocytogenes ST9 Isolates from the Meat Processing Industry: Resolving Diversity and Transmission Patterns Using Whole-Genome Sequencing. Appl. Environ. Microbiol. 2020, 86, e00579-20. [Google Scholar] [CrossRef] [PubMed]
- Guidi, F.; Orsini, M.; Chiaverini, A.; Torresi, M.; Centorame, P.; Acciari, V.A.; Salini, R.; Palombo, B.; Brandi, G.; Amagliani, G.; et al. Hypo- and Hyper-Virulent Listeria monocytogenes Clones Persisting in Two Different Food Processing Plants of Central Italy. Microorganisms 2021, 9, 376. [Google Scholar] [CrossRef] [PubMed]
- Disson, O.; Moura, A.; Lecuit, M. Making Sense of the Biodiversity and Virulence of Listeria monocytogenes. Trends Microbiol. 2021, 29, 811–822. [Google Scholar] [CrossRef]
- Jennison, A.V.; Masson, J.J.; Fang, N.-X.; Graham, R.M.; Bradbury, M.I.; Fegan, N.; Gobius, K.S.; Graham, T.M.; Guglielmino, C.J.; Brown, J.L.; et al. Analysis of the Listeria monocytogenes Population Structure among Isolates from 1931 to 2015 in Australia. Front. Microbiol. 2017, 8, 603. [Google Scholar] [CrossRef] [Green Version]
- Bustamante, F.; Maury-Sintjago, E.; Leal, F.C.; Acuña, S.; Aguirre, J.; Troncoso, M.; Figueroa, G.; Parra-Flores, J. Presence of Listeria monocytogenes in Ready-to-Eat Artisanal Chilean Foods. Microorganisms 2020, 8, 1669. [Google Scholar] [CrossRef]
- Shen, J.; Rump, L.; Zhang, Y.; Chen, Y.; Wang, X.; Meng, J. Molecular Subtyping and Virulence Gene Analysis of Listeria monocytogenes Isolates from Food. Food Microbiol. 2013, 35, 58–64. [Google Scholar] [CrossRef]
- Su, X.; Cao, G.; Zhang, J.; Pan, H.; Zhang, D.; Kuang, D.; Yang, X.; Xu, X.; Shi, X.; Meng, J. Characterization of Internalin Genes in Listeria monocytogenes from Food and Humans, and Their Association with the Invasion of Caco-2 Cells. Gut Pathog. 2019, 11, 30. [Google Scholar] [CrossRef]
- Van Stelten, A.; Roberts, A.R.; Manuel, C.S.; Nightingale, K.K. Listeria monocytogenes Isolates Carrying Virulence-Attenuating Mutations in Internalin A Are Commonly Isolated from Ready-to-Eat Food Processing Plant and Retail Environments. J. Food Prot. 2016, 79, 1733–1740. [Google Scholar] [CrossRef]
- Filipello, V.; Mughini-Gras, L.; Gallina, S.; Vitale, N.; Mannelli, A.; Pontello, M.; Decastelli, L.; Allard, M.W.; Brown, E.W.; Lomonaco, S. Attribution of Listeria monocytogenes Human Infections to Food and Animal Sources in Northern Italy. Food Microbiol. 2020, 89, 103433. [Google Scholar] [CrossRef]
- Ortiz, S.; López-Alonso, V.; Rodríguez, P.; Martínez-Suárez, J.V. The Connection between Persistent, Disinfectant-Resistant Listeria monocytogenes Strains from Two Geographically Separate Iberian Pork Processing Plants: Evidence from Comparative Genome Analysis. Appl. Environ. Microbiol. 2016, 82, 308–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiavano, G.F.; Ateba, C.N.; Petruzzelli, A.; Mele, V.; Amagliani, G.; Guidi, F.; De Santi, M.; Pomilio, F.; Blasi, G.; Gattuso, A.; et al. Whole-Genome Sequencing Characterization of Virulence Profiles of Listeria monocytogenes Food and Human Isolates and In Vitro Adhesion/Invasion Assessment. Microorganisms 2022, 10, 62. [Google Scholar] [CrossRef] [PubMed]
- Magagna, G.; Filipello, V.; Spinelli, E.; Gori, M.; Tanzi, E.; Finazzi, G. Rare Internalin A Premature Stop Codon on Listeria monocytogenes on Italian PDO Cheeses Isolates. Eur. J. Public Health 2021, 31, ckab165.635. [Google Scholar] [CrossRef]
- Medeiros, M.; de Castro, V.H.L.; Mota, A.L.A.d.A.; Pereira, M.G.; De Martinis, E.C.P.; Perecmanis, S.; Santana, A.P. Assessment of Internalin A Gene Sequences and Cell Adhesion and Invasion Capacity of Listeria monocytogenes Strains Isolated from Foods of Animal and Related Origins. Foodborne Pathog. Dis. 2021, 18, 243–252. [Google Scholar] [CrossRef]
- Dellafiora, L.; Filipello, V.; Dall’Asta, C.; Finazzi, G.; Galaverna, G.; Losio, M.N. A Structural Study on the Listeria monocytogenes Internalin A—Human E-Cadherin Interaction: A Molecular Tool to Investigate the Effects of Missense Mutations. Toxins 2020, 12, 60. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Francisco, A.P.; Vaz, C.; Monteiro, P.T.; Melo-Cristino, J.; Ramirez, M.; Carriço, J.A. PHYLOViZ: Phylogenetic Inference and Data Visualization for Sequence Based Typing Methods. BMC Bioinform. 2012, 13, 87. [Google Scholar] [CrossRef] [Green Version]
- Magagna, G.; Finazzi, G.; Filipello, V. Newly Designed Primers for the Sequencing of the InlA Gene of Lineage I and II Listeria monocytogenes Isolates. Int. J. Mol. Sci. 2022, 23, 14106. [Google Scholar] [CrossRef]
- Babraham Bioinformatics—FastQC A Quality Control Tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 25 January 2022).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- BBMap. Available online: https://sourceforge.net/projects/bbmap/ (accessed on 11 February 2022).
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality Assessment Tool for Genome Assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joensen, K.G.; Scheutz, F.; Lund, O.; Hasman, H.; Kaas, R.S.; Nielsen, E.M.; Aarestrup, F.M. Real-Time Whole-Genome Sequencing for Routine Typing, Surveillance, and Outbreak Detection of Verotoxigenic Escherichia Coli. J. Clin. Microbiol. 2014, 52, 1501–1510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and Applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C.J. Open-Access Bacterial Population Genomics: BIGSdb Software, the PubMLST.Org Website and Their Applications. Wellcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- R Core Team; European Environment Agency. 2020. Available online: https://www.eea.europa.eu/data-and-maps/indicators/oxygen-consuming-substances-in-rivers/r-development-core-team-2006 (accessed on 10 October 2022).
- Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses, R Package Version 1.0.7; 2020. Available online: https://CRAN.R-project.org/package=factoextra (accessed on 10 October 2022).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PMSC Mutation Type | Nucleotide Position of Mutation | Length of Truncated InlA (aa) | Lineage | References |
|---|---|---|---|---|
| 1 | 1818 (T→A) | 605 | I | Nightingale et al., 2005 [20] |
| 2 | 1966 (C→T) | 655 | I | Nightingale et al., 2005 [20] |
| 3 | 2100 (C→G) | 699 | II | Nightingale et al., 2005 [20] |
| 4 | 12 (deletion A) | 8 | II | Felício et al., 2007 [21] |
| 5 | 565 (C→T) | 188 | II | Van Stelten and Nightingale, 2008 [22] |
| 6 | 1474 (C→T) | 491 | II | Olier et al., 2003 [23] |
| 7 | 1684 (C→T) | 561 | Van Stelten and Nightingale, 2008 [22] | |
| 8 | 1380 (G→A) | 459 | II | Rousseaux et al., 2004 [24] |
| 9 | 1540 (deletion G) | 518 | Rousseaux et al., 2004 [24] | |
| 10 | 1961 (insertion T) | 676 | Rousseaux et al., 2004 [24] | |
| 11 | 2054 (G→A) | 684 | II | Rousseaux et al., 2004 [24] |
| 12 | 1637 (deletion A) | 576 | Jonquières et al., 1998 [25] | |
| 13 | 1579 (A→T) | 526 | Handa-Miya et al., 2007 [26] | |
| 14 | 1615 (C→T) | 538 | II | Ragon et al., 2008 [9] |
| 15 | 229 (C→T) | 76 | II | Van Stelten et al., 2010 [27] |
| 19 | 976 (G→T) | 325 | II | Gelbíčová et al., 2015 [18] |
| 20 | 288 (C→A) | 95 | I | Moura et al., 2016 [19] |
| 21 | 806 (T→A) | 268 | I | Moura et al., 2016 [19] |
| 22 | 1756 (C→T) | 585 | I | Moura et al., 2016 [19] |
| 23 | 1939 (A→T) | 646 | I | Moura et al., 2016 [19] |
| 24 | 13 (C→T) | 4 | II | Moura et al., 2016 [19] |
| 25 | 12 (deletion A) | 25 | II | Moura et al., 2016 [19] |
| 26 | 277 (G→T) | 92 | II | Moura et al., 2016 [19] |
| 27 | 576 (insertion T) | 194 | II | Moura et al., 2016 [19] |
| 28 | 736–738 (CCA→TAG) | 245 | II | Moura et al., 2016 [19] |
| 29 | 1635 (deletion A) | 576 | II | Moura et al., 2016 [19] |
| 30 | 1741 (C→T) | 580 | II | Moura et al., 2016 [19] |
| 31 | 2208 (deletion A) | 753 | I | Kurpas et al., 2020 [8] |
| 32 | 1041 (C→A) | 346 | I | Tsai et al., 2022 [28] |
| 33 | 937 (deletion C) | 312 | Ji et al., 2023 [29] |
| ST 1 | Lineage | Internalin A | |
|---|---|---|---|
| Complete | PMSC 2 | ||
| ST2 | I | 97.7% (n = 43) | 2.3% (n = 1) |
| ST9 | II | 9.1% (n = 10) | 90.9% (n = 100) |
| ST31 | II | 0 | 100% (n = 5) |
| ST121 | II | 3.4% (n = 2) | 96.6% (n = 56) |
| ST193 | II | 0 | 100% (n = 1) |
| ST199 | II | 0 | 100% (n = 2) |
| ST224 | I | 97.1% (n = 34) | 2.9% (n = 1) |
| ST325 | II | 1.9% (n = 1) | 98.1% (n = 51) |
| ST580 | II | 0 | 100% (n = 6) |
| ST717 | II | 0 | 100% (n = 2) |
| ST2687 | II | 0 | 100% (n = 3) |
| Patient | Age (Years) | Underlying Conditions | Outcome | Site of Infection | ST 1 | CC 2 | PMSC 3 Type |
|---|---|---|---|---|---|---|---|
| #1 | 99 | Immunosuppressive drugs | Unknown | Blood | 2 | 2 | 6 |
| #2 | 73 | Previous heart attack | Unknown | Liquor | 9 | 9 | 19 |
| #3 | 61 | Unknown | Unknown | Blood | 9 | 9 | 29 |
| #4 | 74 | Unknown | Unknown | Blood | 9 | 9 | 29 |
| #5 | 81 | Unknown | Unknown | Blood | 9 | 9 | 29 |
| #6 | 87 | Liver failure | Favorable | Peritoneal fluid | 9 | 9 | 29 |
| #7 | 67 | Cancer | Favorable | Blood | 9 | 9 | 19 |
| #8 | 38 | Unknown | Unknown | Blood | 121 | 121 | 6 |
| #9 | 71 | Cancer | Favorable | Blood | 121 | 121 | 6 |
| #10 | 89 | Immunosuppressive therapy | Unknown | Blood | 121 | 121 | 6 |
| #11 | 71 | Cancer | Lethal | Blood | 325 | 31 | 26 |
| Gene | Sequence |
|---|---|
| abcZ | abcZoF: GTTTTCCCAGTCACGACGTTGTATCGCTGCTGCCACTTTTATCCA abcZoR: TTGTGAGCGGATAACAATTTCTCAAGGTCGCCGTTTAGAG |
| bglA | bglAoF: GTTTTCCCAGTCACGACGTTGTAGCCGACTTTTTATGGGGTGGAG bglAoR: TTGTGAGCGGATAACAATTTCCGATTAAATACGGTGCGGACATA |
| cat | catoF: GTTTTCCCAGTCACGACGTTGTAATTGGCGCATTTTGATAGAGA catoR: TTGTGAGCGGATAACAATTTCAGATTGACGATTCCTGCTTTTG |
| dapE | dapEoF: GTTTTCCCAGTCACGACGTTGTACGACTAATGGGCATGAAGAACAAG dapEoR: TTGTGAGCGGATAACAATTTCATCGAACTATGGGCATTTTTACC |
| dat | datoF: GTTTTCCCAGTCACGACGTTGTAGAAAGAGAAGATGCCACAGTTGA datoR: TTGTGAGCGGATAACAATTTCTGCGTCCATAATACACCATCTTT |
| ldh | ldhoF: GTTTTCCCAGTCACGACGTTGTAGTATGATTGACATAGATAAAGA ldhoR: TTGTGAGCGGATAACAATTTCTATAAATGTCGTTCATACCAT |
| lhkA | lhkAoF: GTTTTCCCAGTCACGACGTTGTAAGAATGCCAACGACGAAACC lhkAoR: TTGTGAGCGGATAACAATTTCTGGGAAACATCAGCAATAAAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magagna, G.; Gori, M.; Russini, V.; De Angelis, V.; Spinelli, E.; Filipello, V.; Tranquillo, V.M.; De Marchis, M.L.; Bossù, T.; Fappani, C.; et al. Evaluation of the Virulence Potential of Listeria monocytogenes through the Characterization of the Truncated Forms of Internalin A. Int. J. Mol. Sci. 2023, 24, 10141. https://doi.org/10.3390/ijms241210141
Magagna G, Gori M, Russini V, De Angelis V, Spinelli E, Filipello V, Tranquillo VM, De Marchis ML, Bossù T, Fappani C, et al. Evaluation of the Virulence Potential of Listeria monocytogenes through the Characterization of the Truncated Forms of Internalin A. International Journal of Molecular Sciences. 2023; 24(12):10141. https://doi.org/10.3390/ijms241210141
Chicago/Turabian StyleMagagna, Giulia, Maria Gori, Valeria Russini, Veronica De Angelis, Elisa Spinelli, Virginia Filipello, Vito Massimo Tranquillo, Maria Laura De Marchis, Teresa Bossù, Clara Fappani, and et al. 2023. "Evaluation of the Virulence Potential of Listeria monocytogenes through the Characterization of the Truncated Forms of Internalin A" International Journal of Molecular Sciences 24, no. 12: 10141. https://doi.org/10.3390/ijms241210141