
Zooming into Gut Dysbiosis in Parkinson’s Disease: New Insights from Functional Mapping
 , , , , ,
, , , , , {kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
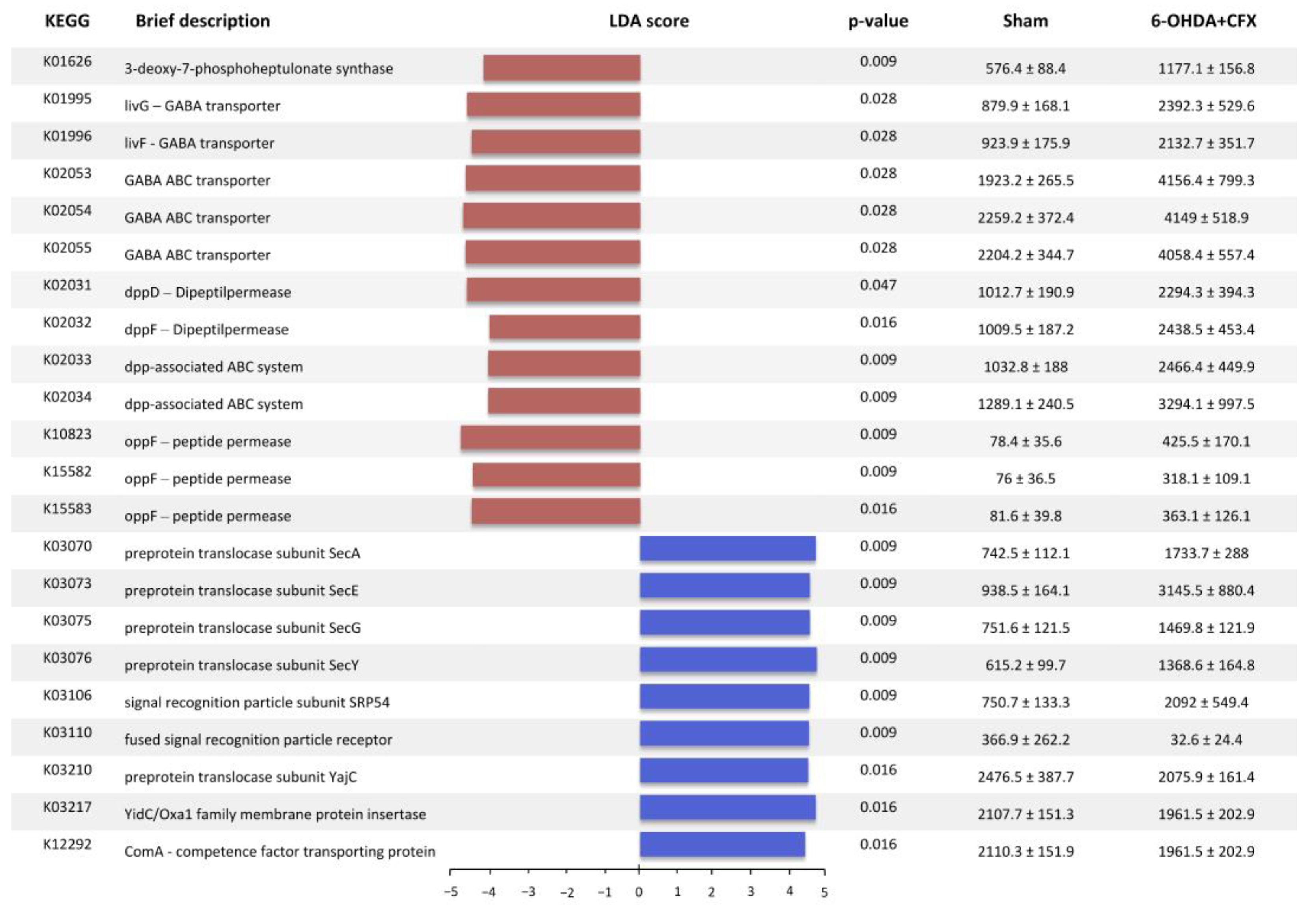
2.1. Mapping of Functions Related to SCFAs Metabolism and Acetate and Butyrate Fecal Level Quantification
2.2. Mapping of Functions Related to QS
3. Materials and Methods
3.1. Establishment of Dual-Hit Mouse Model
3.2. Genomic DNA Extraction, Illumina MiSeq Sequencing of Barcoded 16S rRNA Gene Amplicons, and Sequence Data Processing
3.3. In Silico Predicted Metagenomic Analysis Based on Microbiota Sequencing Data
3.4. Fecal SCFAs Extraction
3.5. Gas Chromatography–Mass Spectrometry (GC/MS) Analysis
3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Avagliano, C.; Coretti, L.; Lama, A.; Pirozzi, C.; De Caro, C.; De Biase, D.; Turco, L.; Mollica, M.P.; Paciello, O.; Calignano, A.; et al. Dual-Hit Model of Parkinson’s Disease: Impact of Dysbiosis on 6-Hydroxydopamine-Insulted Mice—Neuroprotective and Anti-Inflammatory Effects of Butyrate. Int. J. Mol. Sci. 2022, 23, 6367. [Google Scholar] [CrossRef]
- Lama, A.; Annunziata, C.; Coretti, L.; Pirozzi, C.; Di Guida, F.; Nitrato Izzo, A.; Cristiano, C.; Mollica, M.P.; Chiariotti, L.; Pelagalli, A.; et al. N-(1-Carbamoyl-2-Phenylethyl) Butyramide Reduces Antibiotic-Induced Intestinal Injury, Innate Immune Activation and Modulates Microbiota Composition. Sci. Rep. 2019, 9, 4832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, Z.; Yue, W.; Kined, C.; Wang, P.; Liu, R.; Liu, J.; Chen, X. Bacillus Licheniformis Reverses the Environmental Ceftriaxone Sodium-Induced Gut Microbial Dysbiosis and Intestinal Inflammation in Mice. Ecotoxicol. Environ. Saf. 2023, 257, 114890. [Google Scholar] [CrossRef] [PubMed]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2 for Prediction of Metagenome Functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef] [PubMed]
- Trompette, A.; Gollwitzer, E.S.; Yadava, K.; Sichelstiel, A.K.; Sprenger, N.; Ngom-Bru, C.; Blanchard, C.; Junt, T.; Nicod, L.P.; Harris, N.L.; et al. Gut Microbiota Metabolism of Dietary Fiber Influences Allergic Airway Disease and Hematopoiesis. Nat. Med. 2014, 20, 159–166. [Google Scholar] [CrossRef]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Feng, J.; Liu, C.; Wu, H.; Qiu, Z.; Ge, J.; Sun, S.; Hong, X.; Li, Y.; Wang, X.; et al. Machine Learning Aided Construction of the Quorum Sensing Communication Network for Human Gut Microbiota. Nat. Commun. 2022, 13, 3079. [Google Scholar] [CrossRef]
- Mukherjee, S.; Bassler, B.L. Bacterial Quorum Sensing in Complex and Dynamically Changing Environments. Nat. Rev. Microbiol. 2019, 17, 371–382. [Google Scholar] [CrossRef]
- Zhang, J.; Zuo, K.; Fang, C.; Yin, X.; Liu, X.; Zhong, J.; Li, K.; Li, J.; Xu, L.; Yang, X. Altered Synthesis of Genes Associated with Short-Chain Fatty Acids in the Gut of Patients with Atrial Fibrillation. BMC Genom. 2021, 22, 634. [Google Scholar] [CrossRef]
- Oliphant, K.; Allen-Vercoe, E. Macronutrient Metabolism by the Human Gut Microbiome: Major Fermentation by-Products and Their Impact on Host Health. Microbiome 2019, 7, 91. [Google Scholar] [CrossRef]
- Mallet, D.; Goutaudier, R.; Barbier, E.L.; Carnicella, S.; Colca, J.R.; Fauvelle, F.; Boulet, S. Re-Routing Metabolism by the Mitochondrial Pyruvate Carrier Inhibitor MSDC-0160 Attenuates Neurodegeneration in a Rat Model of Parkinson’s Disease. Mol. Neurobiol. 2022, 59, 6170–6182. [Google Scholar] [CrossRef] [PubMed]
- Toomey, C.E.; Heywood, W.E.; Evans, J.R.; Lachica, J.; Pressey, S.N.; Foti, S.C.; Al Shahrani, M.; D’Sa, K.; Hargreaves, I.P.; Heales, S.; et al. Mitochondrial Dysfunction Is a Key Pathological Driver of Early Stage Parkinson’s. Acta Neuropathol. Commun. 2022, 10, 134. [Google Scholar] [CrossRef] [PubMed]
- Billingsley, K.J.; Barbosa, I.A.; Bandrés-Ciga, S.; Quinn, J.P.; Bubb, V.J.; Deshpande, C.; Botia, J.A.; Reynolds, R.H.; Zhang, D.; Simpson, M.A.; et al. Mitochondria Function Associated Genes Contribute to Parkinson’s Disease Risk and Later Age at Onset. NPJ Park. Dis. 2019, 5, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quansah, E.; Peelaerts, W.; Langston, J.W.; Simon, D.K.; Colca, J.; Brundin, P. Targeting Energy Metabolism via the Mitochondrial Pyruvate Carrier as a Novel Approach to Attenuate Neurodegeneration. Mol. Neurodegener. 2018, 13, 28. [Google Scholar] [CrossRef] [Green Version]
- Gray, L.R.; Tompkins, S.C.; Taylor, E.B. Regulation of Pyruvate Metabolism and Human Disease. Cell. Mol. Life Sci. 2014, 71, 2577–2604. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.S.; Santosh, W.; Kumar, S.; Christlet, H.T.T. Metabolic Profiling of Parkinson’s Disease: Evidence of Biomarker from Gene Expression Analysis and Rapid Neural Network Detection. J. Biomed. Sci. 2009, 16, 63. [Google Scholar] [CrossRef] [Green Version]
- Mallet, D.; Dufourd, T.; Decourt, M.; Carcenac, C.; Bossù, P.; Verlin, L.; Fernagut, P.-O.; Benoit-Marand, M.; Spalletta, G.; Barbier, E.L.; et al. A Metabolic Biomarker Predicts Parkinson’s Disease at the Early Stages in Patients and Animal Models. J. Clin. Investig. 2022, 132, e146400. [Google Scholar] [CrossRef]
- Paxinos, G.; Franklin, K.B.J. Paxinos and Franklin’s the Mouse Brain in Stereotaxic Coordinates, 5th ed.; Academic Press: Cambridge, MA, USA, 2019. [Google Scholar]
- Avagliano, C.; Russo, R.; De Caro, C.; Cristiano, C.; La Rana, G.; Piegari, G.; Paciello, O.; Citraro, R.; Russo, E.; De Sarro, G.; et al. Palmitoylethanolamide Protects Mice against 6-OHDA-Induced Neurotoxicity and Endoplasmic Reticulum Stress: In Vivo and in Vitro Evidence. Pharmacol. Res. 2016, 113, 276–289. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a Chimera-Checked 16S RRNA Gene Database and Workbench Compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langille, M.G.I.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Vega Thurber, R.L.; Knight, R.; et al. Predictive Functional Profiling of Microbial Communities Using 16S RRNA Marker Gene Sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef] [PubMed]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic Biomarker Discovery and Explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamas, A.; Regal, P.; Vázquez, B.; Cepeda, A.; Franco, C.M. Short Chain Fatty Acids Commonly Produced by Gut Microbiota Influence Salmonella Enterica Motility, Biofilm Formation, and Gene Expression. Antibiotics 2019, 8, 265. [Google Scholar] [CrossRef] [Green Version]
- Amrutha, B.; Sundar, K.; Shetty, P.H. Effect of Organic Acids on Biofilm Formation and Quorum Signaling of Pathogens from Fresh Fruits and Vegetables. Microb. Pathog. 2017, 111, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; O’Neill, A.M.; Williams, M.R.; Cau, L.; Nakatsuji, T.; Horswill, A.R.; Gallo, R.L. Short Chain Fatty Acids Produced by Cutibacterium Acnes Inhibit Biofilm Formation by Staphylococcus Epidermidis. Sci. Rep. 2020, 10, 21237. [Google Scholar] [CrossRef]
- Meng, F.; Zhao, H.; Nie, T.; Lu, F.; Zhang, C.; Lu, Y.; Lu, Z. Acetate Activates Lactobacillus Bacteriocin Synthesis by Controlling Quorum Sensing. Appl. Environ. Microbiol. 2021, 87, e00720-21. [Google Scholar] [CrossRef]
- Hawver, L.A.; Giulietti, J.M.; Baleja, J.D.; Ng, W.-L. Quorum Sensing Coordinates Cooperative Expression of Pyruvate Metabolism Genes to Maintain a Sustainable Environment for Population Stability. mBio 2016, 7, e01863-16. [Google Scholar] [CrossRef] [Green Version]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Turco, L.; Opallo, N.; Buommino, E.; De Caro, C.; Pirozzi, C.; Mattace Raso, G.; Lembo, F.; Coretti, L. Zooming into Gut Dysbiosis in Parkinson’s Disease: New Insights from Functional Mapping. Int. J. Mol. Sci. 2023, 24, 9777. https://doi.org/10.3390/ijms24119777
Turco L, Opallo N, Buommino E, De Caro C, Pirozzi C, Mattace Raso G, Lembo F, Coretti L. Zooming into Gut Dysbiosis in Parkinson’s Disease: New Insights from Functional Mapping. International Journal of Molecular Sciences. 2023; 24(11):9777. https://doi.org/10.3390/ijms24119777
Chicago/Turabian StyleTurco, Luigia, Nicola Opallo, Elisabetta Buommino, Carmen De Caro, Claudio Pirozzi, Giuseppina Mattace Raso, Francesca Lembo, and Lorena Coretti. 2023. "Zooming into Gut Dysbiosis in Parkinson’s Disease: New Insights from Functional Mapping" International Journal of Molecular Sciences 24, no. 11: 9777. https://doi.org/10.3390/ijms24119777