
Fatty Acid Composition of Dry and Germinating Pollen of Gymnosperm and Angiosperm Plants



Abstract
:1. Introduction
2. Results
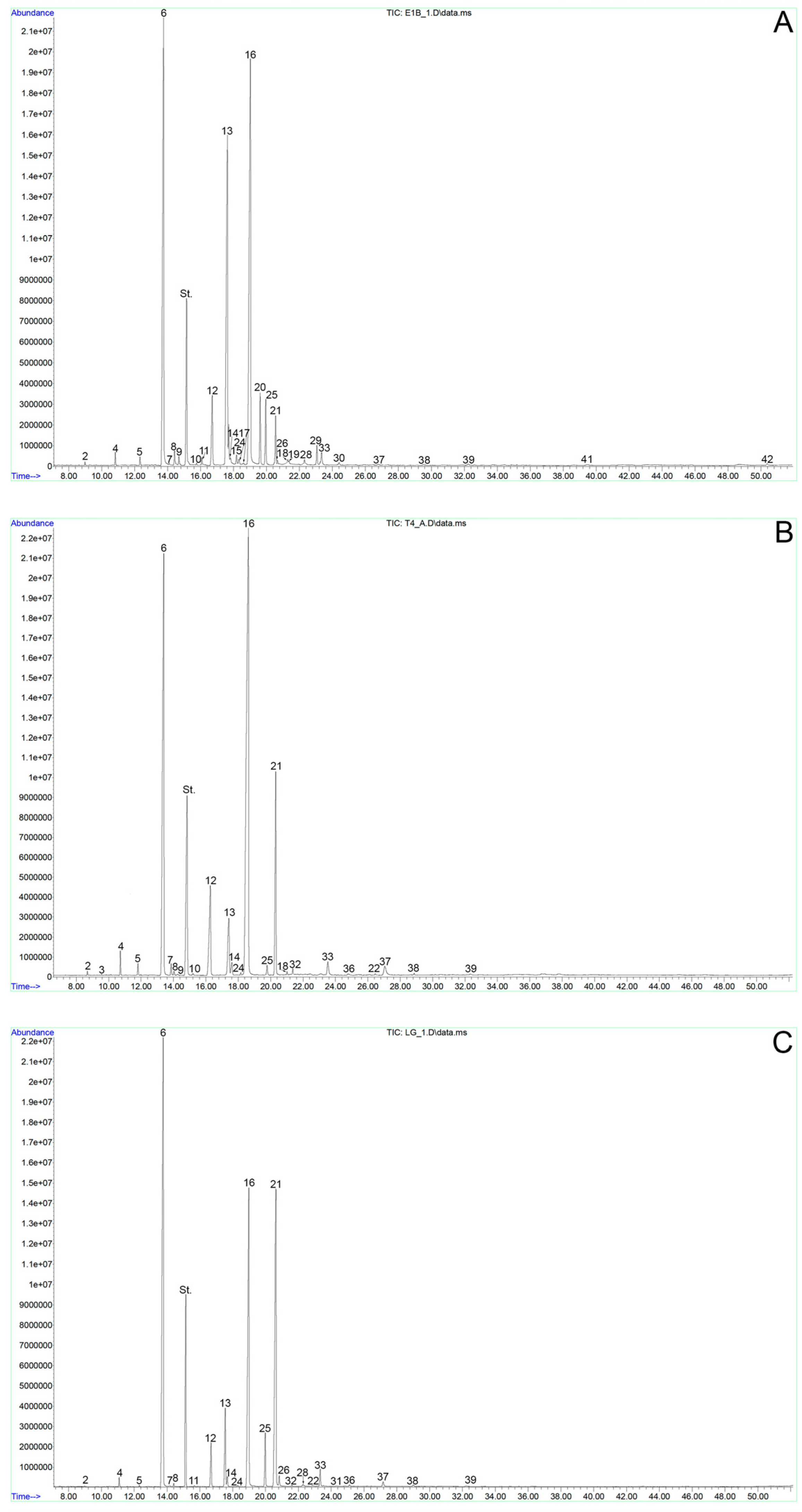
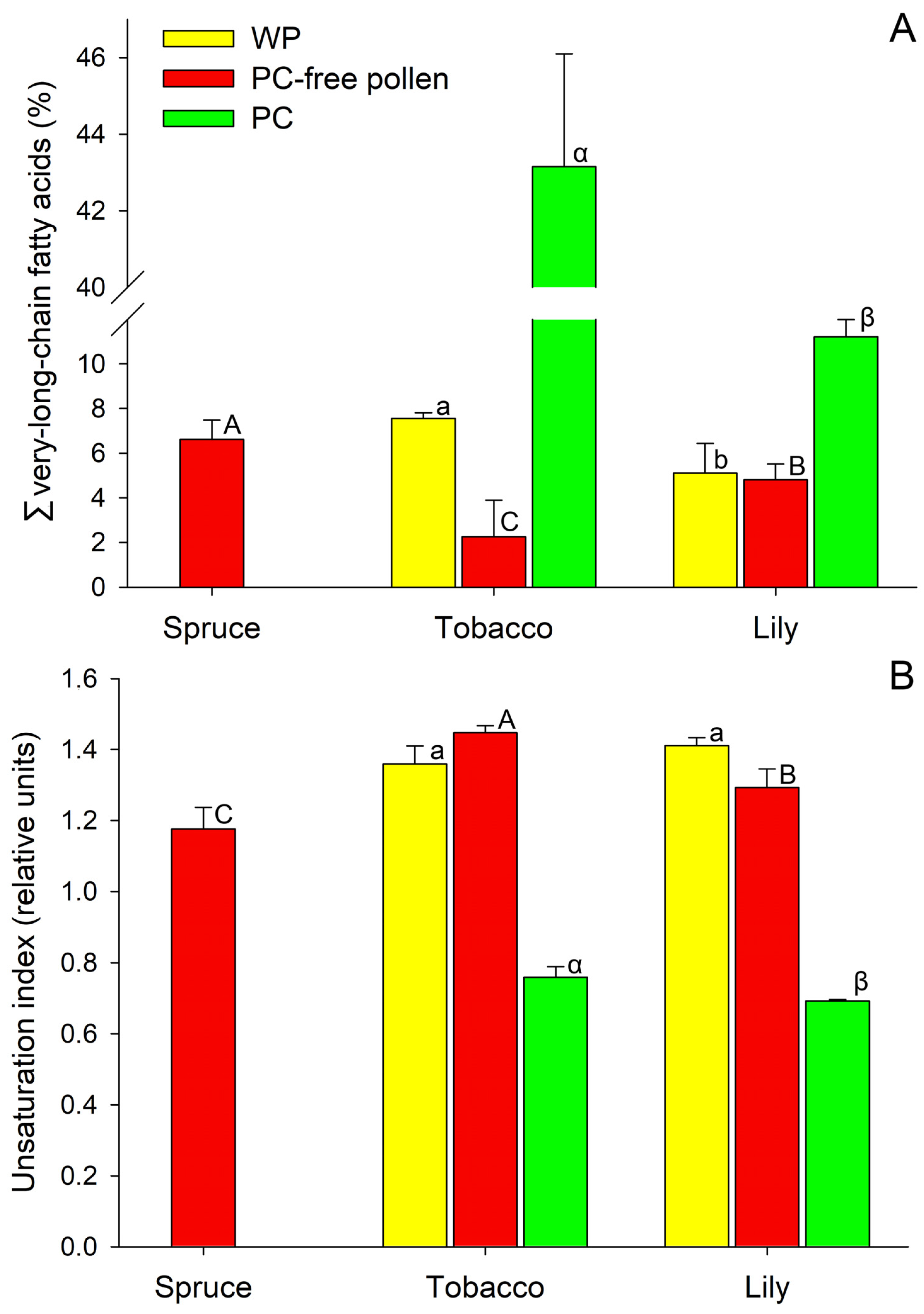
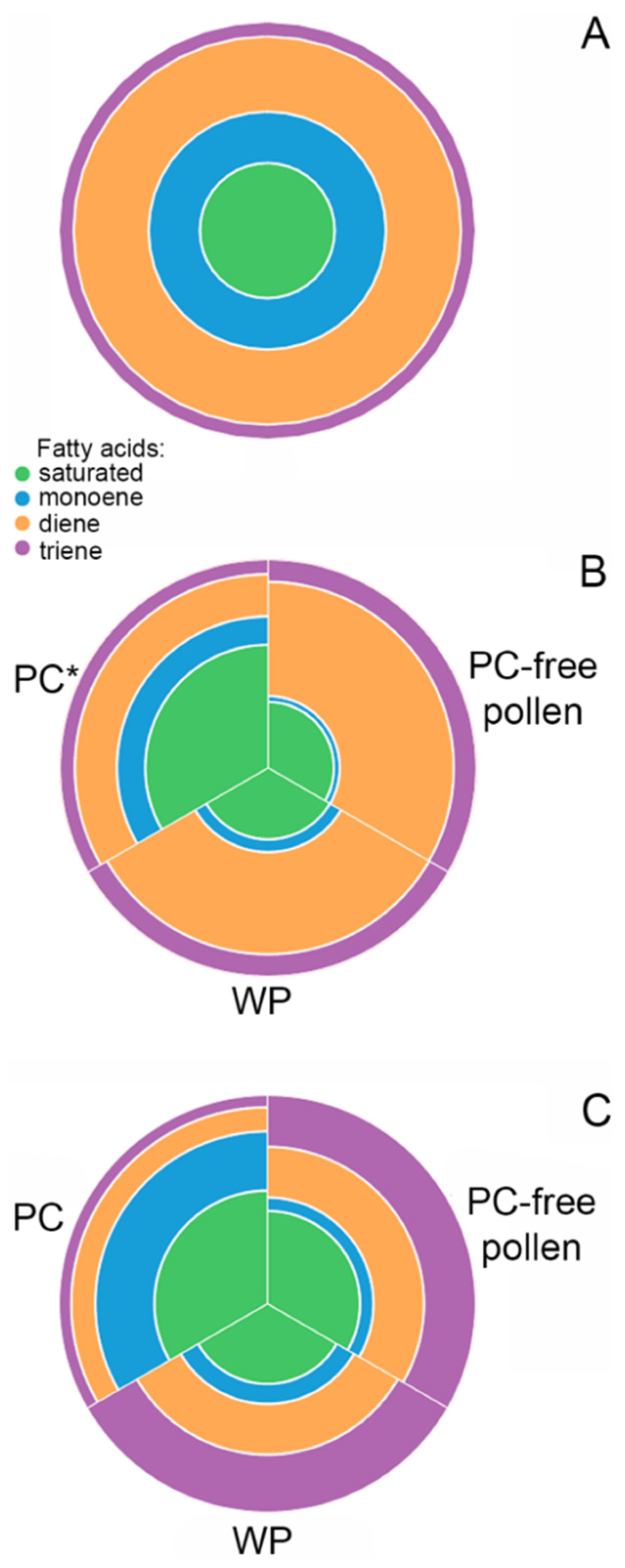
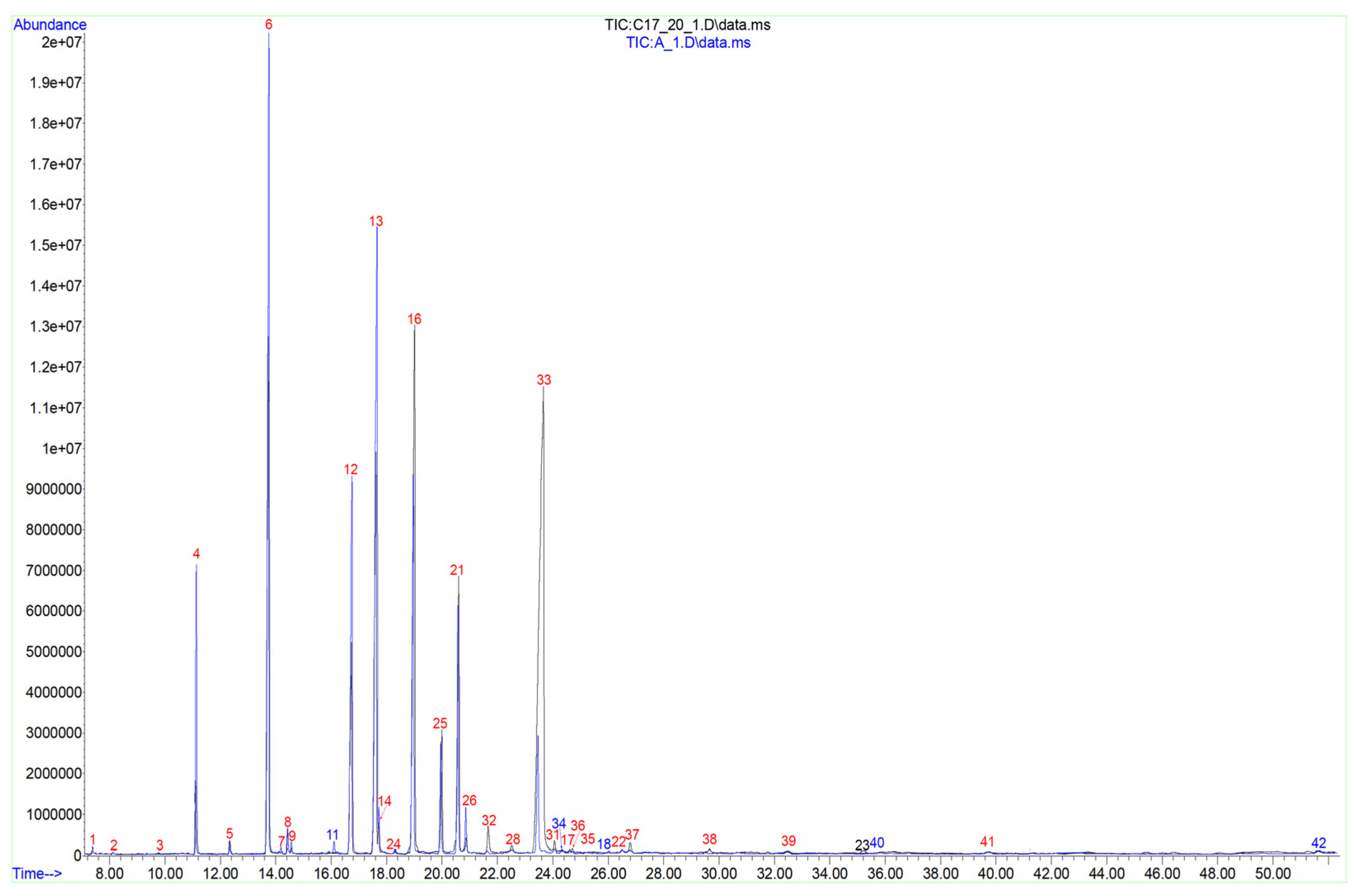
2.1. Fatty Acid Composition of Spruce Pollen
2.2. Fatty Acid Composition of Tobacco Pollen
2.3. Fatty Acid Composition of Lily Pollen
2.4. The Lipid Absolute Contents of Spruce, Tobacco and Lily Pollen
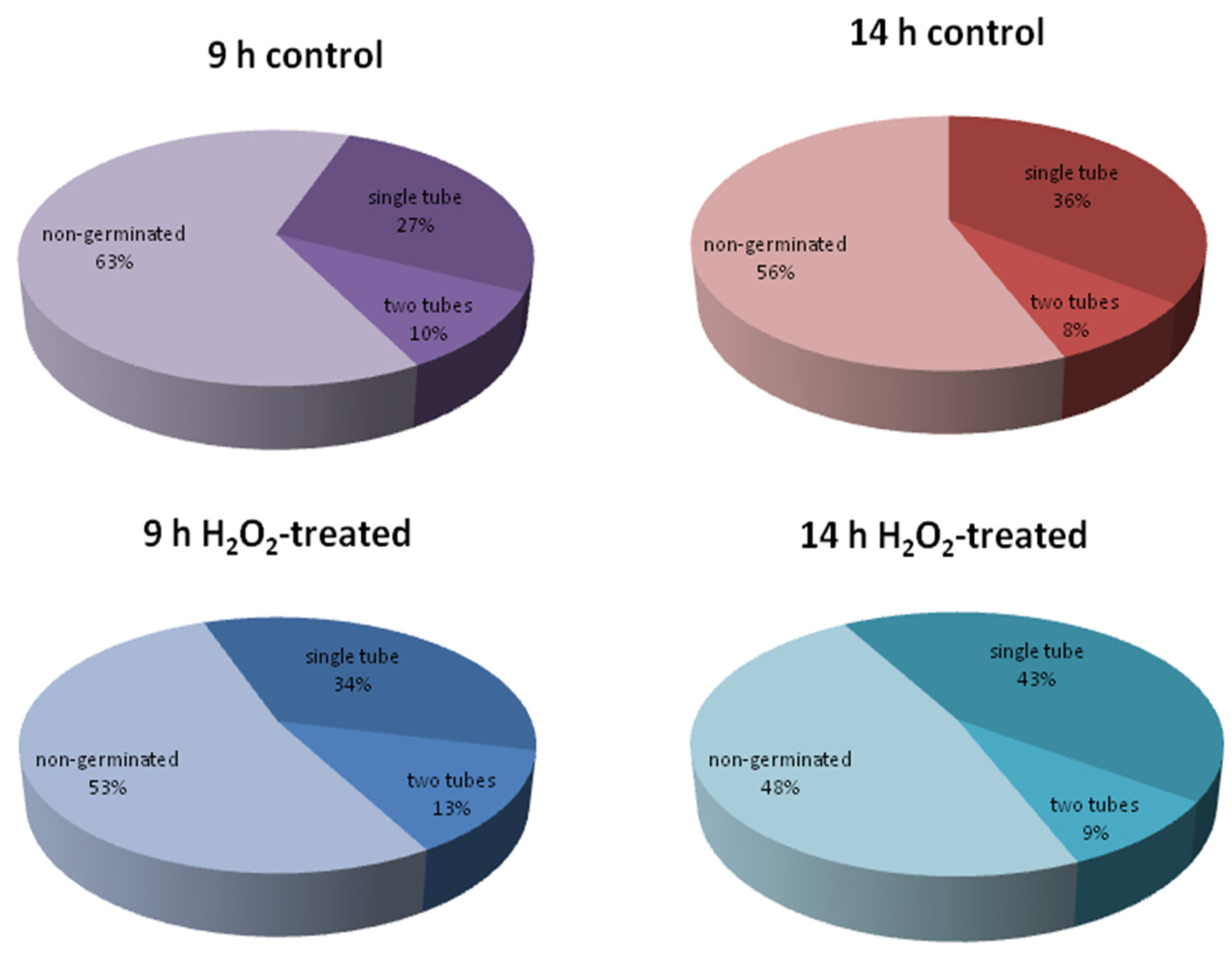
2.5. The Effect of Hydrogen Peroxide on Tobacco and Spruce Pollen Germination
2.6. Dynamics of the Fatty Acid Composition and Lipid Absolute Content of Spruce Pollen during Germination with or without H2O2
2.7. Dynamics of the Fatty Acid Composition and Lipid Absolute Content of Tobacco Pollen during Germination with or without H2O2
3. Discussion
4. Materials and Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Breygina, M.; Klimenko, E.; Schekaleva, O. Pollen germination and pollen tube growth in gymnosperms. Plants 2021, 10, 1301. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Teng, N.; Wu, X.; Wang, Y.; Tang, W.; Šamaj, J.; Baluška, F.; Lin, J. Disruption of actin filaments by latrunculin B affects cell wall construction in Picea meyeri pollen tube by disturbing vesicle trafficking. Plant Cell Physiol. 2007, 48, 19–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maksimov, N.; Evmenyeva, A.; Breygina, M.; Yermakov, I. The role of reactive oxygen species in pollen germination in Picea pungens (blue spruce). Plant Reprod. 2018, 18, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Lazzaro, M.D.; Marom, E.Y.; Reddy, A.S.N. Polarized cell growth, organelle motility, and cytoskeletal organization in conifer pollen tube tips are regulated by KCBP, the calmodulin-binding kinesin. Planta 2013, 238, 587–597. [Google Scholar] [CrossRef]
- Sowa, S.; Connor, K.F. Biochemical changes during pollen germination measured in vivo by infrared spectroscopy. Plant Sci. 1995, 105, 23–30. [Google Scholar] [CrossRef]
- Sidorov, R.A.; Kuznetsova, E.I.; Pchelkin, V.P.; Zhukov, A.V.; Gorshkova, E.N.; Tsydendambaev, V.D. Fatty acid composition of the pollen lipids of Cycas revoluta Thunb. Grasas Y Aceites 2016, 67, 5. [Google Scholar] [CrossRef] [Green Version]
- Muren, R.C.; Ching, T.M.; Ching, K.I.M.K. Metabolic Study of Douglas-fir Pollen Germination in vitro. Physiol. Plant 1979, 46, 287–292. [Google Scholar] [CrossRef]
- Stanley, R.G. Pollen Chemistry and Tube Growth. In Pollen Development and Physiology; Elsevier: Amsterdam, The Netherlands, 1971; pp. 131–155. [Google Scholar]
- Shi, J.; Cui, M.; Yang, L.; Kim, Y.J.; Zhang, D. Genetic and Biochemical Mechanisms of Pollen Wall Development. Trends Plant Sci. 2015, 20, 741–753. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Xu, C.; Li-Beisson, Y.; Philippar, K. Fatty Acid and Lipid Transport in Plant Cells. Trends Plant Sci. 2016, 21, 145–158. [Google Scholar] [CrossRef] [Green Version]
- Piffanelli, P.; Ross, J.H.E.; Murphy, D.J. Biogenesis and function of the lipidic structures of pollen grains. Sex. Plant Reprod. 1998, 11, 65–80. [Google Scholar] [CrossRef]
- Pacini, E.; Hesse, M. Pollenkitt—Its composition, forms and functions. Flora Morphol. Distrib. Funct. Ecol. Plants 2005, 200, 399–415. [Google Scholar] [CrossRef]
- Carmichael, J.S.; Friedman, W.E. Double fertilization in Gnetum gnemon (Gnetaceae): Its bearing on the evolution of sexual reproduction within the gnetales and the anthophyte clade. Am. J. Bot. 1996, 83, 767–780. [Google Scholar] [CrossRef]
- Hesse, M. Pollenkitt is lacking in Gnetum gnemon (Gnetaceae). Plant Syst. Evol. 1980, 136, 41–46. [Google Scholar] [CrossRef]
- Ischebeck, T. Lipids in pollen—They are different. Biochim. Biophys. Acta—Mol. Cell Biol. Lipids 2016, 1861, 1315–1328. [Google Scholar] [CrossRef] [PubMed]
- Firon, N.; Nepi, M.; Pacini, E. Water status and associated processes mark critical stages in pollen development and functioning. Ann. Bot. 2012, 109, 1201–1213. [Google Scholar] [CrossRef] [Green Version]
- Manning, R. Fatty acids in pollen: A review of their importance for honey bees. Bee World 2001, 82, 60–75. [Google Scholar] [CrossRef]
- Manning, R.; Harvey, M. Fatty acids in honeybee-collected pollens from six endemic Western Australian eucalypts and the possible significance to the Western Australian beekeeping industry. Aust. J. Exp. Agric. 2002, 42, 217–223. [Google Scholar] [CrossRef]
- Preuss, D.; Lemieux, B.; Yen, G.; Davis, R.W. A conditional sterile mutation eliminates surface components from Arabidopsis pollen and disrupts cell signaling during fertilization. Genes Dev. 1993, 7, 974–985. [Google Scholar] [CrossRef] [Green Version]
- Jessen, D.; Olbrich, A.; Knüfer, J.; Krüger, A.; Hoppert, M.; Polle, A.; Fulda, M. Combined activity of LACS1 and LACS4 is required for proper pollen coat formation in Arabidopsis. Plant J. 2011, 68, 715–726. [Google Scholar] [CrossRef]
- Chichiriccò, G.; Pacini, E.; Lanza, B. Pollenkitt of some monocotyledons: Lipid composition and implications for pollen germination. Plant Biol. 2019, 21, 920–926. [Google Scholar] [CrossRef]
- Zhang, M.; Fan, J.; Taylor, D.C.; Ohlrogge, J.B. DGAT1 and PDAT1 acyltransferases have overlapping functions in Arabidopsis triacylglycerol biosynthesis and are essential for normal pollen and seed development. Plant Cell 2009, 21, 3885–3901. [Google Scholar] [CrossRef] [Green Version]
- Mellema, S.; Eichenberger, W.; Rawyler, A.; Suter, M.; Tadege, M.; Kuhlemeier, C. The ethanolic fermentation pathway supports respiration and lipid biosynthesis in tobacco pollen. Plant J. 2002, 30, 329–336. [Google Scholar] [CrossRef] [Green Version]
- Ischebeck, T.; Valledor, L.; Lyon, D.; Gingl, S.; Nagler, M.; Meijón, M.; Egelhofer, V.; Weckwerth, W. Comprehensive cell-specific protein analysis in early and late pollen development from diploid microsporocytes to pollen tube growth. Mol. Cell. Proteom. 2014, 13, 295–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krawczyk, H.E.; Rotsch, A.H.; Herrfurth, C.; Scholz, P.; Shomroni, O.; Salinas-Riester, G.; Feussner, I.; Ischebeck, T. Heat stress leads to rapid lipid remodeling and transcriptional adaptations in Nicotiana tabacum pollen tubes. Plant Physiol. 2022, 189, 490–515. [Google Scholar] [CrossRef] [PubMed]
- Serrano, N.; Pejchar, P.; Soukupová, H.; Hubálek, M.; Potocký, M. Comprehensive analysis of glycerolipid dynamics during tobacco pollen germination and pollen tube growth. Front. Plant Sci. 2022, 13, 1028311. [Google Scholar] [CrossRef] [PubMed]
- Sklar, L.A.; Hudson, B.S.; Simoni, R.D. Conjugated polyene fatty acids as membrane probes: Preliminary characterization. Proc. Natl. Acad. Sci. USA 1975, 72, 1649–1653. [Google Scholar] [CrossRef] [Green Version]
- Hiscock, S.J.; Bright, J.; McInnis, S.M.; Desikan, R.; Hancock, J.T. Signaling on the stigma. Potential new roles for ROS and NO in plant cell signaling. Plant Signal. Behav. 2007, 2, 23–24. [Google Scholar] [CrossRef] [Green Version]
- Breygina, M.; Klimenko, E.; Shilov, E.; Podolyan, A.; Mamaeva, A.; Zgoda, V.; Fesenko, I. Hydrogen peroxide in tobacco stigma exudate affects pollen proteome and membrane potential in pollen tubes. Plant Biol. 2021, 23, 592–602. [Google Scholar] [CrossRef]
- Breygina, M.; Klimenko, E.; Podolyan, A.; Voronkov, A. Dynamics of pollen activation and the role of H+-ATPase in pollen germination in blue spruce (Picea pungens). Plants 2020, 9, 1760. [Google Scholar] [CrossRef]
- Breygina, M.; Maksimov, N.; Polevova, S.; Evmenyeva, A. Bipolar pollen germination in blue spruce (Picea pungens). Protoplasma 2019, 256, 941–949. [Google Scholar] [CrossRef]
- Mongrand, S.; Badoc, A.; Patouille, B.; Lacomblez, C.; Chavent, M.; Cassagne, C.; Bessoule, J.-J. Taxonomy of gymnospermae: Multivariate analyses of leaf fatty acid composition. Phytochemistry 2001, 58, 101–115. [Google Scholar] [CrossRef]
- Takagi, T.; Itabashi, Y. Cis-5-Olefinic Unusual Fatty Acids in Seed Lipids of Gymnospermae and Their Distribution in Triacylglycerols. Lipids 1982, 17, 716–723. [Google Scholar] [CrossRef]
- Ching, T.M.; Ching, K.K. Fatty acids in pollen of some coniferous species. Science 1962, 138, 890–891. [Google Scholar] [CrossRef]
- Erdyneeva, S.A.; Shiretorova, V.G.; Tykheev, Z.A.; Radnaeva, L.D. Fatty-Acid Composition of Pollen from Pinus sylvestris, P. sibirica, and P. pumila. Chem. Nat. Compd. 2021, 57, 741–742. [Google Scholar] [CrossRef]
- Andrikopoulos, N.K.; Siafaka-Kapadai, A.; Demopoulos, C.A.; Kapoulas, V.M. Lipids of Pinus halepensis pollen. Phytochemistry 1985, 24, 2953–2957. [Google Scholar] [CrossRef]
- Chung, H.K.; Lee, K.Y. A study of the chemical constituents of gymnosperm pollens. Seoul J. Med. 1977, 18, 125–136. [Google Scholar]
- Mei, S.; Yang, C.; Song, X.; Wang, T.; Wang, Y.; Geng, Y.; Wang, J.; Su, S. Analysis of the effect of sterilization and storage on the quality of Pinus yunnanensis pollen based on untargeted metabonomics. J. Food Process. Preserv. 2021, 45, e16033. [Google Scholar] [CrossRef]
- Wolff, R.L.; Comps, B.; Marpeau, A.M.; Deluc, L.G. Taxonomy of Pinus species based on the seed oil fatty acid compositions. Trees—Struct. Funct. 1997, 12, 113–118. [Google Scholar] [CrossRef]
- Song, L.; Wen, S.; Ye, Q.; Lou, H.; Gao, Y.; Bajpai, V.K.; Carpena, M.; Prieto, M.-A.; Simal-Gandara, J.; Xiao, J.; et al. Advances on delta 5-unsaturated-polymethylene-interrupted fatty acids: Resources, biosynthesis, and benefits. Crit. Rev. Food Sci. Nutr. 2023, 63, 767–789. [Google Scholar] [CrossRef]
- Yamaoka, Y.; Shin, S.; Choi, B.Y.; Kim, H.; Jang, S.; Kajikawa, M.; Yamano, T.; Kong, F.; Légeret, B.; Fukuzawa, H.; et al. The bZIP1 Transcription Factor Regulates Lipid Remodeling and Contributes to ER Stress Management in Chlamydomonas reinhardtii. Plant Cell 2019, 31, 1127–1140. [Google Scholar] [CrossRef] [Green Version]
- Paz Ramos, A.; Lague, P.; Lamoureux, G.; Lafleur, M. Effect of saturated very long-chain fatty acids on the organization of lipid membranes: A study combining 2H NMR spectroscopy and molecular dynamics simulations. J. Phys. Chem. B 2016, 120, 6951–6960. [Google Scholar] [CrossRef]
- Williams, C.G. Conifer Reproductive Biology; Springer: Dordrecht, The Netherlands, 2009; ISBN 978-1-4020-9601-3. [Google Scholar]
- Williams, J.H. Novelties of the flowering plant pollen tube underlie diversification of a key life history stage. Pnas 2008, 105, 11259–11263. [Google Scholar] [CrossRef] [Green Version]
- Rothnie, N.E.; Palmer, M.V.; Burke, D.G.; Sang, J.P.; Hilliard, E.P.; Salisbury, P.A.; Evans, D.E.; Knox, R.B.; Williams, E.G. Comparative analysis of fatty acids in pollen and seed of rapeseed. Phytochemistry 1987, 26, 1895–1897. [Google Scholar] [CrossRef]
- Piffanelli, P.; Ross, J.H.E.; Murphy, D.J. Intra-and extracellular lipid composition and associated gene expression patterns during pollen development in Brassica napus. Plant J. 1997, 11, 549–562. [Google Scholar] [CrossRef]
- Mansy, S.S.; Szostak, J.W. Thermostability of model protocell membranes. Proc. Natl. Acad. Sci. USA 2008, 105, 13351–13355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michaelian, K.; Rodríguez, O. Prebiotic fatty acid vesicles through photochemical dissipative structuring. Rev. Cuba. Química 2019, 31, 354–370. [Google Scholar]
- Fan, Y.; Fang, Y.; Ma, L. The self-crosslinked ufasome of conjugated linoleic acid: Investigation of morphology, bilayer membrane and stability. Colloids Surf. B Biointerfaces 2014, 123, 8–14. [Google Scholar] [CrossRef]
- Verma, P.; Kaur, K.; Wanchoo, R.K.; Toor, A.P. Esterification of acetic acid to methyl acetate using activated TiO2 under UV light irradiation at ambient temperature. J. Photochem. Photobiol. A Chem. 2017, 336, 170–175. [Google Scholar] [CrossRef]
- Barrantes, F.J. Fluorescence sensors for imaging membrane lipid domains and cholesterol. In Current Topics in Membranes; Elsevier: Amsterdam, The Netherlands, 2021; Volume 88, pp. 257–314. ISBN 9780323911146. [Google Scholar]
- Kuypers, F.A.; van den Berg, J.J.M.; Schalkwijk, C.; Roelofsen, B.; Kamp, J.A.F.O. den Parinaric acid as a sensitive fluorescent probe for the determination of lipid peroxidation. Biochim. Biophys. Acta (BBA)/Lipids Lipid Metab. 1987, 921, 266–274. [Google Scholar] [CrossRef] [Green Version]
- Lundberg, W. Alkali-isomerization reactions of unsaturated fatty acids. Am. J. Clin. Nutr. 1958, 6, 592–593. [Google Scholar] [CrossRef] [PubMed]
- Shayanmehr, M.-R.; Elhamirad, A.-H.; Armin, M. Different Conditions for Alkaline Isomerization and the Production of Conjugated Linoleic Acid (CLA) from Linoleic Acid. J. Biochem. Tech. 2018, 2, 1–8. [Google Scholar]
- Svetashev, V.I.; Imbs, A.B. Isomerization of octadecapentaenoic acid (18:5n-3) in algal lipid samples under derivatization for GC and GC-MS analysis. J. Phycol. 2014, 50, 322–327. [Google Scholar] [CrossRef]
- Pierson, E.S.; Lichtscheidl, I.K.; Derksen, J.A.N. Structure and Behaviour of Organelles in Living Pollen Tubes of Lilium longiflorum. J. Exp. Bot. 1990, 41, 1461–1468. [Google Scholar] [CrossRef]
- Rosen, W.G.; Gawlik, S.R.; Dashek, W.V.; Siegesmund, K.A. Fine structure and cytochemistry of lilium pollen tubes. Am. J. Bot. 1964, 51, 61–71. [Google Scholar] [CrossRef]
- Cresti, M.; Keijzer, C.J.; Tiezzi, A.; Ciampolini, F.; Focardi, S. Stigma of Nicotiana: Ultrastructural and biochemical studies. Am. J. Bot. 1986, 73, 1713–1722. [Google Scholar] [CrossRef]
- Nitsch, J.P. Deux espaces photoperiodiques de jours courts: Plumbago indica L. et P. zeyelanica. Bull. Bot. Fr. 1965, 112, 517–522. [Google Scholar] [CrossRef] [Green Version]
- Voronkov, A.S.; Ivanova, T.V. Fatty Acids Composition of the Epiphytic Ferns, Platycerium bifurcatum and Asplenium nidus, and the Terrestrial Fern, Asplenium trichomanes. Am. Fern J. 2021, 111, 117–128. [Google Scholar] [CrossRef]
- Voronkov, A.; Ivanova, T. Significance of Lipid Fatty Acid Composition for Resistance to Winter Conditions in Asplenium scolopendrium. Biology 2022, 11, 507. [Google Scholar] [CrossRef]
- Voronkov, A.S.; Ivanova, T.V.; Kuznetsova, E.I.; Kumachova, T.K. Adaptations of Malus domestica Borkh. (Rosaceae) Fruits Grown at Different Altitudes. Russ. J. Plant Physiol. 2019, 66, 922–931. [Google Scholar] [CrossRef]
- Ivanova, T.V.; Voronkov, A.S.; Kuznetsova, E.I.; Kumachova, T.K.; Zhirov, V.K.; Tsydendambaev, V.D. Lipid Fatty Acids from the Pericarp of Cydonia oblonga Mill. and Mespilus germanica L. are Involved in Plant Adaptation to Altitudinal Zonality. Dokl. Biochem. Biophys. 2019, 486, 229–233. [Google Scholar] [CrossRef]
- Lyons, J.M.; Wheaton, T.A.; Pratt, H.K. Relationship between the Physical Nature of Mitochondrial Membranes and Chilling Sensitivity in Plants. Plant Physiol. 1964, 39, 262–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | FA | Spruce Pollen | Tobacco | Lily | ||||
|---|---|---|---|---|---|---|---|---|
| WP | PC-Free Pollen | PC | WP | PC-Free Pollen | PC | |||
| 1 | 10:0 | - | 0.01 ± 0.01 | - | 0.08 ± 0.02β | trace # | - | 0.19 ± 0.01α |
| 2 | 12:0 | 0.10 ± 0.01A | 0.03 ± 0.01b | 0.06 ± 0.02B | 0.29 ± 0.04β | 0.22 ± 0.08a | 0.04 ± 0.01B | 2.97 ± 0.04α |
| 3 | 13:0 | - | 0.01 ± 0.01 | 0.01 ± 0.01 | 0.02 ± 0.01α | trace | - | 0.03 ± 0.01α |
| 4 | 14:0 | 0.53 ± 0.06A | 0.49 ± 0.05b | 0.42 ± 0.07A | 1.22 ± 0.14β | 0.75 ± 0.08a | 0.44 ± 0.08A | 5.56 ± 0.27α |
| 5 | 15:0 | 0.30 ± 0.03A | 0.36 ± 0.05a | 0.29 ± 0.06A | 0.26 ± 0.02α | 0.10 ± 0.01b | 0.08 ± 0.01B | 0.28 ± 0.01α |
| 6 | 16:0 | 23.12 ± 1.57C | 24.91 ± 0.10b | 26.93 ± 0.76B | 11.65 ± 1.12β | 30.25 ± 1.09a | 37.35 ± 3.00A | 23.68 ± 1.01α |
| 7 | 7–16:1 | 0.57 ± 0.05A | 0.53 ± 0.14a | 0.42 ± 0.06B | 0.50 ± 0.06α | 0.25 ± 0.01b | 0.17 ± 0.05C | 0.55 ± 0.01α |
| 8 | 9–16:1 | 0.23 ± 0.01A | 0.14 ± 0.01b | 0.12 ± 0.01C | 0.13 ± 0.03β | 0.23 ± 0.01a | 0.19 ± 0.01B | 0.25 ± 0.02α |
| 9 | 14–16:1 | 0.44 ± 0.04A | 0.07 ± 0.02a | 0.02 ± 0.01B | 0.12 ± 0.01α | 0.01 ± 0.01b | - | 0.04 ± 0.01β |
| 10 | 7,10–16:2 | 0.01 ± 0.01A | trace | 0.03 ± 0.01A | - | - | - | - |
| 11 | 10–17:1 | 0.09 ± 0.02A | - | - | - | 0.02 ± 0.01 | 0.01 ± 0.01B | 0.07 ± 0.03 |
| 12 | 18:0 | 3.48 ± 0.26A | 1.75 ± 0.23b | 1.68 ± 0.17C | 4.82 ± 0.07β | 2.89 ± 0.37a | 2.61 ± 0.14B | 11.98 ± 0.90α |
| 13 | 9–18:1 | 21.47 ± 0.94A | 4.32 ± 0.83b | 1.36 ± 0.09C | 11.82 ± 0.91β | 7.68 ± 0.53a | 4.75 ± 0.26B | 25.11 ± 1.18α |
| 14 | 11–18:1 | 1.21 ± 0.28A | 0.83 ± 0.03a | 0.86 ± 0.03B | 0.61 ± 0.09β | 0.68 ± 0.19a | 0.65 ± 0.05C | 0.91 ± 0.11α |
| 15 | 5,9–18:2 | 0.33 ± 0.10 | - | - | - | - | - | - |
| 16 | 9,12–18:2 | 33.79 ± 3.74B | 47.47 ± 1.80a | 54.10 ± 0.50A | 17.46 ± 1.54α | 23.97 ± 0.68b | 24.04 ± 1.44C | 10.99 ± 0.07β |
| 17 | 10,13–18:2 | 0.05 ± 0.01 | 0.07 ± 0.02a | - | 0.79 ± 0.10 | 0.04 ± 0.04a | - | trace |
| 18 | 10,12–18:2 | 0.58 ± 0.09A | 0.60 ± 0.75 | 0.36 ± 0.06A | - | trace | - | 0.13 ± 0.01 |
| 19 | 9,11–18:2 | 0.73 ± 0.09 | - | - | - | - | - | - |
| 20 | 5,9,12–18:3 | 3.56 ± 0.49 | - | - | - | - | - | |
| 21 | 9,12,15–18:3 | 2.53 ± 0.28C | 10.68 ± 1.64b | 10.97 ± 0.51B | 6.92 ± 0.39α | 27.69 ± 0.80a | 24.80 ± 0.84A | 5.79 ± 0.42α |
| 22 | 9,11,13–18:3 | - | trace | 0.02 ± 0.02A | 0.08 ± 0.01α | trace | 0.02 ± 0.01A | 0.13 ± 0.05α |
| 23 | 9,11,13,15–18:4 | - | trace | - | 0.02 ± 0.01 | - | - | - |
| 24 | 19:0 | 0.26 ± 0.16A | 0.13 ± 0.01a | 0.10 ± 0.03A | 0.06 ± 0.01β | 0.07 ± 0.01b | 0.04 ± 0.02B | 0.13 ± 0.02α |
| 25 | 20:0 | 2.71 ± 0.15A | 0.92 ± 0.21b | 0.30 ± 0.05B | 2.70 ± 0.14α | 2.40 ± 0.41a | 2.57 ± 0.34A | 2.83 ± 0.17α |
| 26 | 11–20:1 | 0.48 ± 0.05B | 0.25 ± 0.15b | - | 0.38 ± 0.01β | 0.71 ± 0.02a | 0.62 ± 0.01A | 1.19 ± 0.05α |
| 28 | 11,14–20:2 | 0.36 ± 0.06A | 0.37 ± 0.03a | - | 1.11 ± 0.14α | 0.15 ± 0.02b | 0.15 ± 0.03B | 0.25 ± 0.08β |
| 29 | 5,11,14–20:3 | 1.05 ± 0.14 | - | - | - | - | - | - |
| 30 | 8,11,14–20:3 | 0.01 ± 0.01 | - | - | - | - | - | - |
| 31 | 11,14,17–20:3 | - | 0.12 ± 0.09a | - | 0.35 ± 0.02α | 0.03 ± 0.01a | 0.02 ± 0.01 | 0.08 ± 0.02β |
| 32 | 21:0 | - | trace | 0.05 ± 0.03A | 0.77 ± 0.09α | 0.10 ± 0.05 | 0.09 ± 0.03A | 0.18 ± 0.03β |
| 33 | 22:0 | 0.86 ± 0.14A | 3.77 ± 0.73a | 0.87 ± 0.09A | 29.77 ± 1.81α | 0.87 ± 0.31b | 0.89 ± 0.19A | 3.71 ± 0.16β |
| 34 | 13–22:1 | - | - | - | - | trace | - | 0.14 ± 0.01 |
| 35 | 13,16–22:2 | - | 0.20 ± 0.05 | - | 0.75 ± 0.08α | trace | - | 0.11 ± 0.01β |
| 36 | 23:0 | - | 0.16 ± 0.01a | 0.08 ± 0.03A | 1.40 ± 0.16α | 0.14 ± 0.01a | 0.06 ± 0.01A | 0.47 ± 0.08β |
| 37 | 24:0 | 0.76 ± 0.12A | 1.59 ± 0.40a | 0.93 ± 0.08A | 5.29 ± 0.55α | 0.42 ± 0.11b | 0.35 ± 0.07B | 1.34 ± 0.07β |
| 38 | 25:0 | 0.07 ± 0.03A | 0.09 ± 0.06a | 0.01 ± 0.01B | 0.13 ± 0.01α | 0.05 ± 0.05a | 0.02 ± 0.01B | 0.09 ± 0.01β |
| 39 | 26:0 | 0.16 ± 0.03A | 0.13 ± 0.07a | 0.01 ± 0.01C | 0.47 ± 0.08α | 0.08 ± 0.06a | 0.04 ± 0.01B | 0.54 ± 0.04α |
| 40 | 27:0 | - | - | - | - | trace | - | 0.01 ± 0.01 |
| 41 | 28:0 | 0.15 ± 0.12 | 0.01 ± 0.01a | - | 0.03 ± 0.01β | 0.14 ± 0.24a | - | 0.23 ± 0.05α |
| 42 | 30:0 | 0.01 ± 0.01 | - | - | - | 0.02 ± 0.01 | - | 0.04 ± 0.01 |
| FAs absolute content, µmol/g | 151.11 ± 5.52B | 160.85 ± 13.72b | 136.59 ± 11.60B | 19.88 ± 1.75β | 580.67 ± 79.11a | 430.68 ± 76.10A | 28.24 ± 3.65α | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Breygina, M.; Voronkov, A.; Ivanova, T.; Babushkina, K. Fatty Acid Composition of Dry and Germinating Pollen of Gymnosperm and Angiosperm Plants. Int. J. Mol. Sci. 2023, 24, 9717. https://doi.org/10.3390/ijms24119717
Breygina M, Voronkov A, Ivanova T, Babushkina K. Fatty Acid Composition of Dry and Germinating Pollen of Gymnosperm and Angiosperm Plants. International Journal of Molecular Sciences. 2023; 24(11):9717. https://doi.org/10.3390/ijms24119717
Chicago/Turabian StyleBreygina, Maria, Alexander Voronkov, Tatiana Ivanova, and Ksenia Babushkina. 2023. "Fatty Acid Composition of Dry and Germinating Pollen of Gymnosperm and Angiosperm Plants" International Journal of Molecular Sciences 24, no. 11: 9717. https://doi.org/10.3390/ijms24119717