
Skeletal Muscle-Specific Bis Depletion Leads to Muscle Dysfunction and Early Death Accompanied by Impairment in Protein Quality Control
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
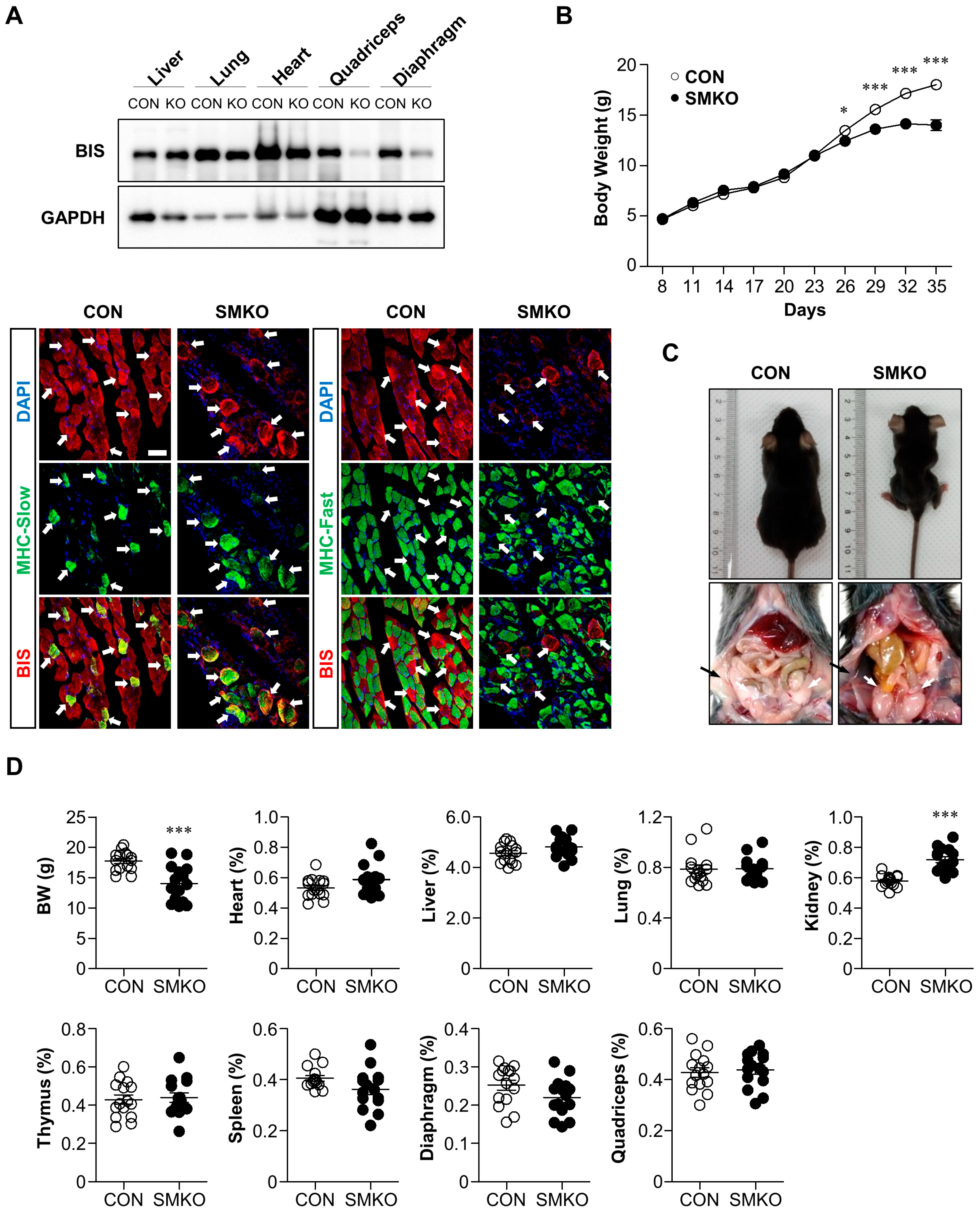
2.1. Bis-SMKO Mice Exhibited Growth Retardation and Early Lethality
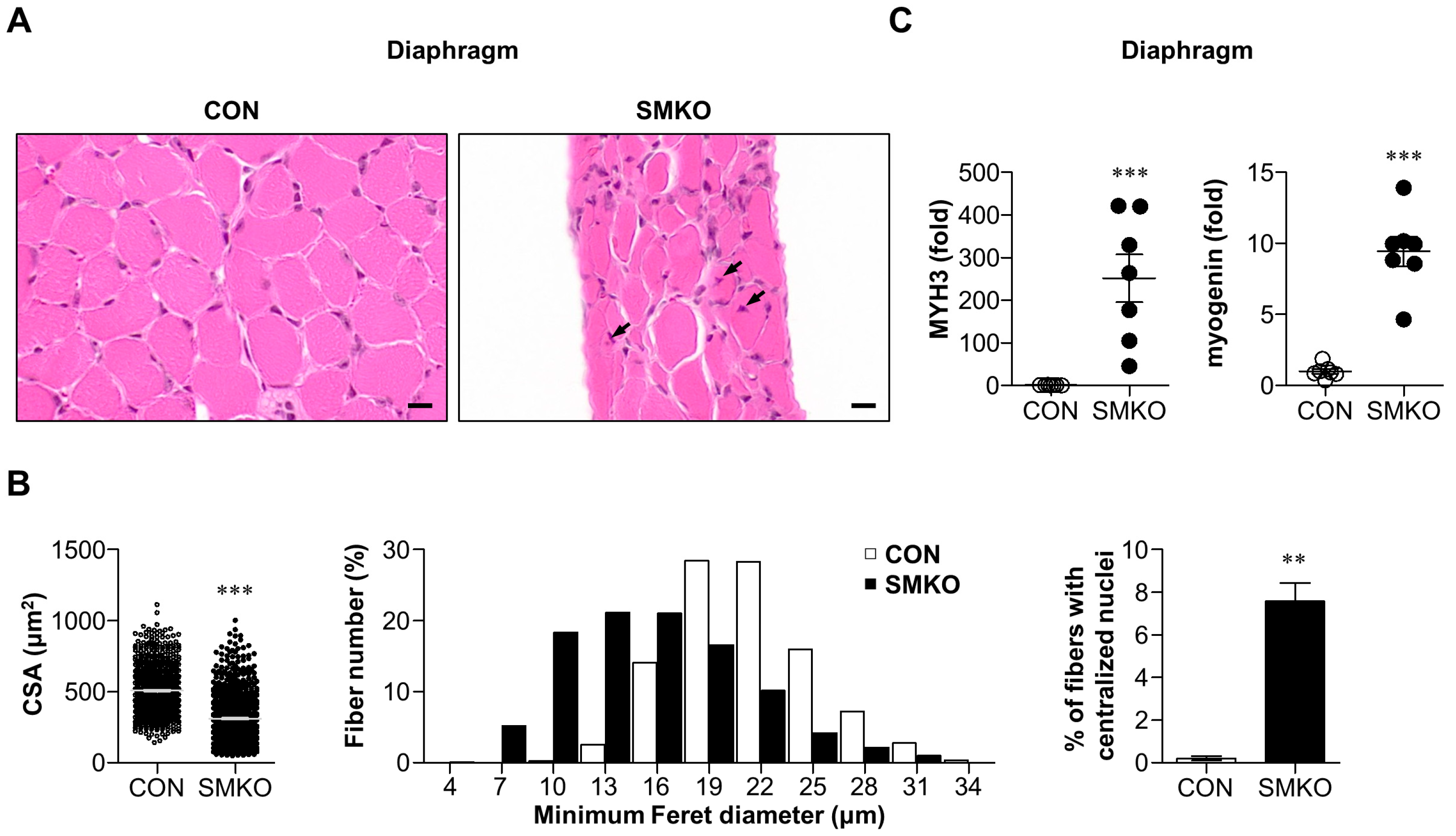
2.2. Muscle Degeneration and Regeneration Were Observed in Bis-SMKO Skeletal Muscles
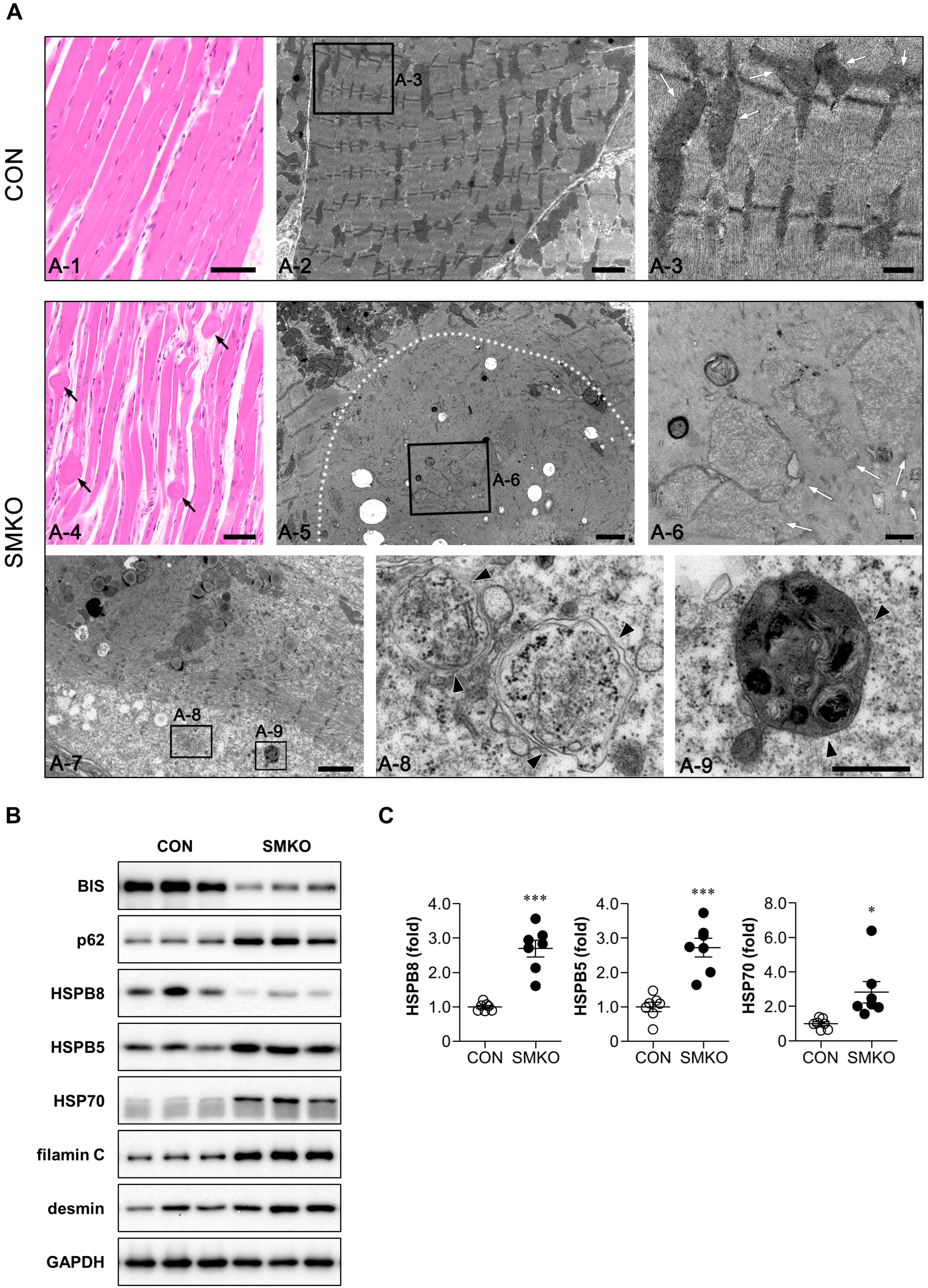
2.3. Accumulation of Heat Shock Proteins (HSPs) and z-Disk Proteins in Bis-SMKO Skeletal Muscles
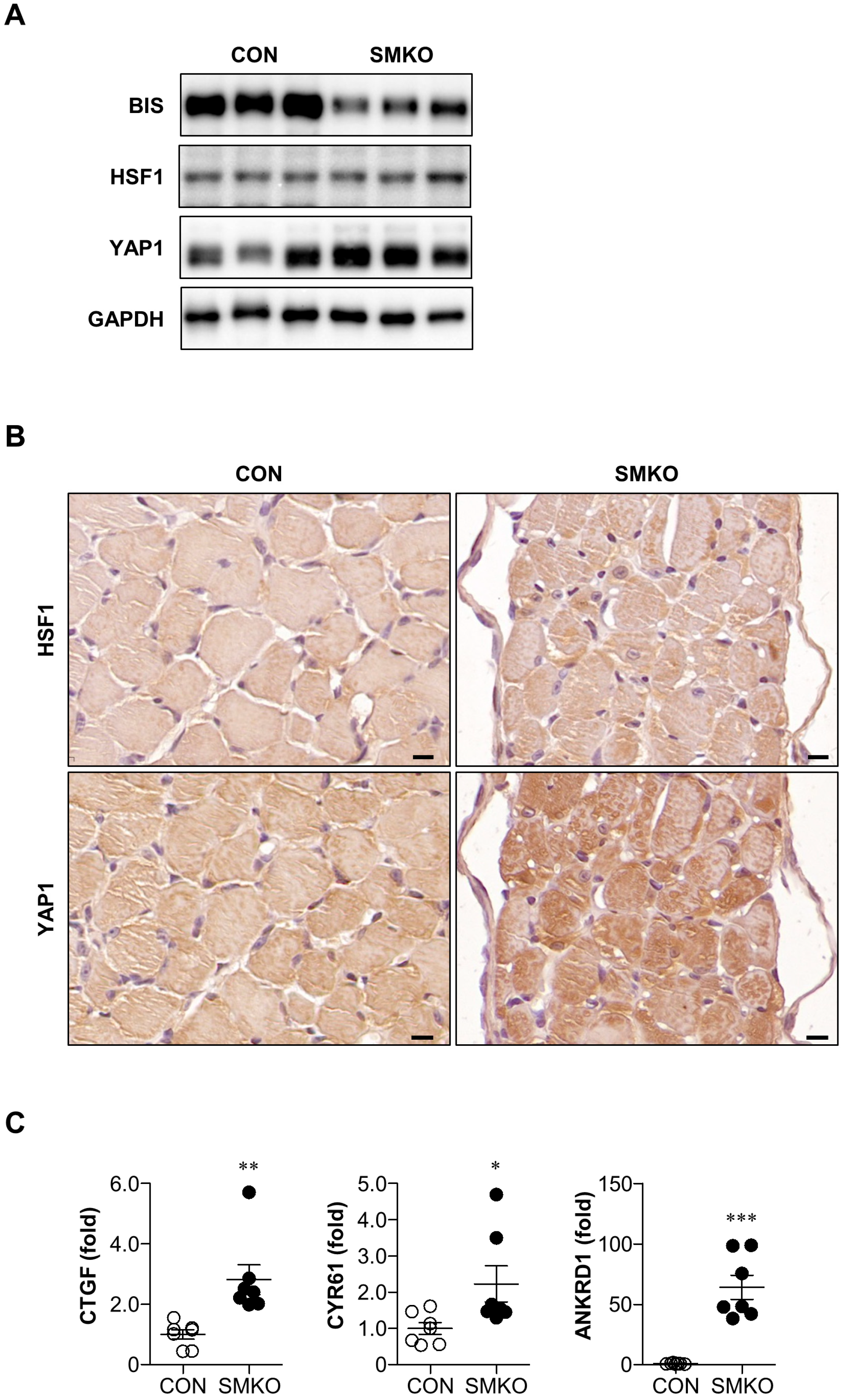
2.4. YAP1 Signaling Was Enhanced in Bis-SMKO Diaphragm
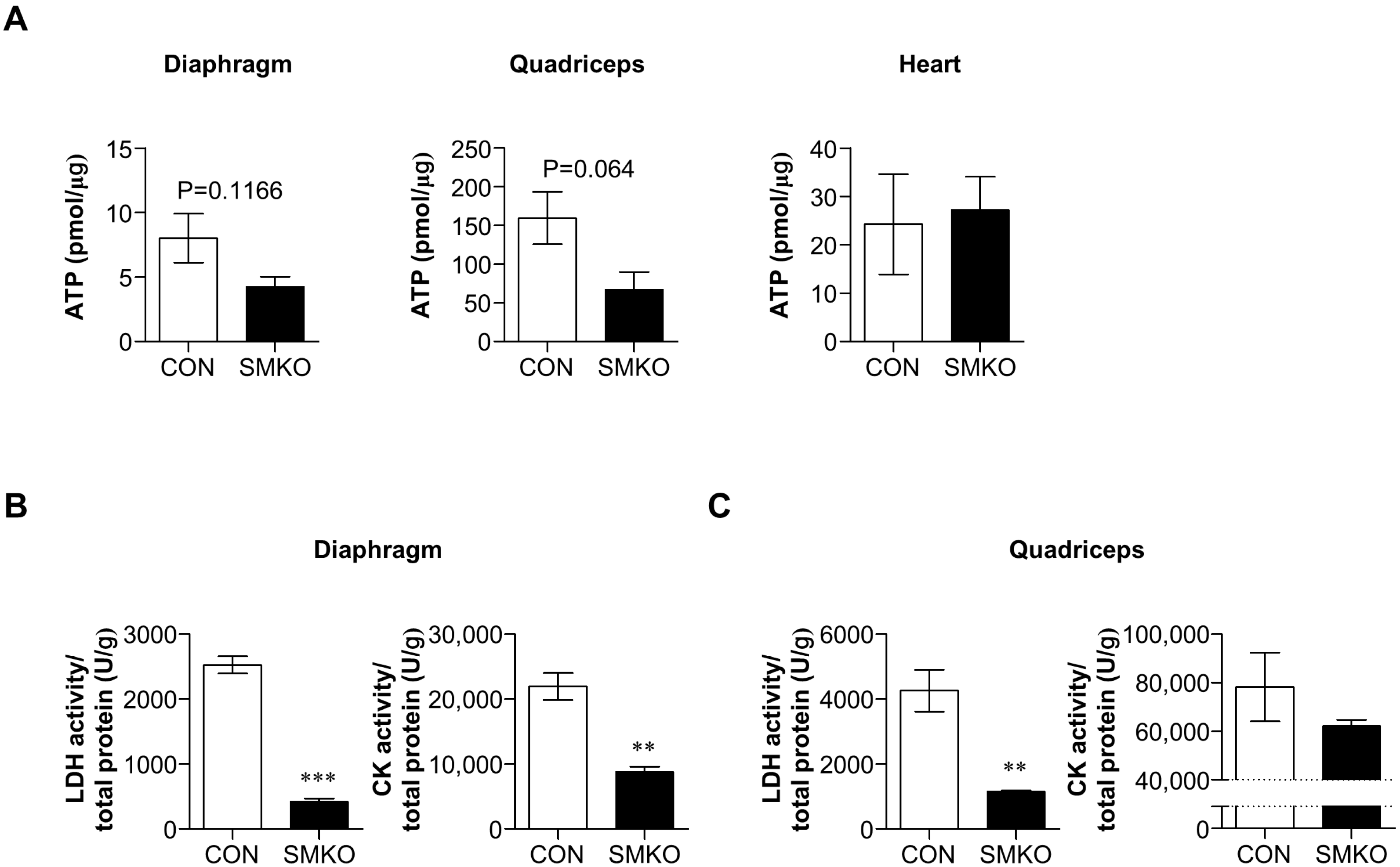
2.5. Bis-SMKO Skeletal Muscles Exhibited Defective Energy Metabolism
3. Discussion
4. Materials and Methods
4.1. Animals
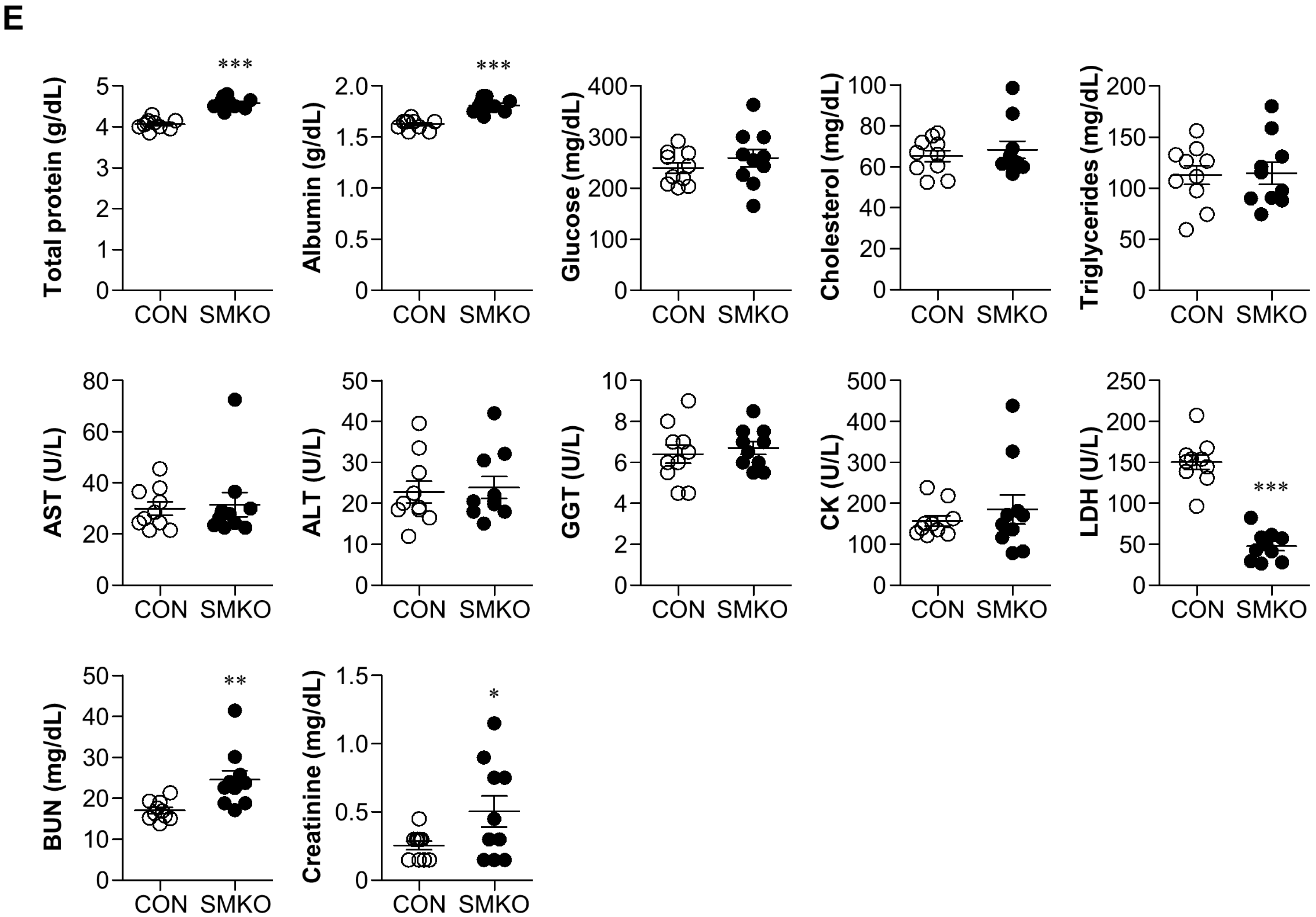
4.2. Biochemical Analysis
4.3. Electron Microscopy
4.4. Histological Analysis
4.5. Quantitative Real-Time Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
4.6. Western Blotting
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lee, J.H.; Takahashi, T.; Yasuhara, N.; Inazawa, J.; Kamada, S.; Tsujimoto, Y. Bis, a Bcl-2-binding protein that synergizes with Bcl-2 in preventing cell death. Oncogene 1999, 18, 6183–6190. [Google Scholar] [CrossRef] [PubMed]
- Takayama, S.; Xie, Z.; Reed, J.C. An evolutionarily conserved family of Hsp70/Hsc70 molecular chaperone regulators. J. Biol. Chem. 1999, 274, 781–786. [Google Scholar] [CrossRef] [PubMed]
- De Marco, M.; Basile, A.; Iorio, V.; Festa, M.; Falco, A.; Ranieri, B.; Pascale, M.; Sala, G.; Remondelli, P.; Capunzo, M.; et al. Role of BAG3 in cancer progression: A therapeutic opportunity. Semin. Cell Dev. Biol. 2018, 78, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Rapino, F.; Abhari, B.A.; Jung, M.; Fulda, S. NIK is required for NF-kappaB-mediated induction of BAG3 upon inhibition of constitutive protein degradation pathways. Cell Death Dis. 2015, 6, e1692. [Google Scholar] [CrossRef]
- Yang, D.; Zhou, J.; Wang, H.; Wang, Y.; Yang, G.; Zhang, Y. High expression of BAG3 predicts a poor prognosis in human medulloblastoma. Tumour Biol. 2016, 37, 13215–13224. [Google Scholar] [CrossRef]
- Shields, S.; Conroy, E.; O’Grady, T.; McGoldrick, A.; Connor, K.; Ward, M.P.; Useckaite, Z.; Dempsey, E.; Reilly, R.; Fan, Y.; et al. BAG3 promotes tumour cell proliferation by regulating EGFR signal transduction pathways in triple negative breast cancer. Oncotarget 2018, 9, 15673–15690. [Google Scholar] [CrossRef]
- Lee, J.J.; Lee, J.S.; Cui, M.N.; Yun, H.H.; Kim, H.Y.; Lee, S.H.; Lee, J.H. BIS targeting induces cellular senescence through the regulation of 14-3-3 zeta/STAT3/SKP2/p27 in glioblastoma cells. Cell Death Dis. 2014, 5, e1537. [Google Scholar] [CrossRef]
- Sturner, E.; Behl, C. The Role of the Multifunctional BAG3 Protein in Cellular Protein Quality Control and in Disease. Front. Mol. Neurosci. 2017, 10, 177. [Google Scholar] [CrossRef]
- Myers, V.D.; McClung, J.M.; Wang, J.; Tahrir, F.G.; Gupta, M.K.; Gordon, J.; Kontos, C.H.; Khalili, K.; Cheung, J.Y.; Feldman, A.M. The Multifunctional Protein BAG3: A Novel Therapeutic Target in Cardiovascular Disease. JACC Basic Transl. Sci. 2018, 3, 122–131. [Google Scholar] [CrossRef]
- Homma, S.; Iwasaki, M.; Shelton, G.D.; Engvall, E.; Reed, J.C.; Takayama, S. BAG3 deficiency results in fulminant myopathy and early lethality. Am. J. Pathol. 2006, 169, 761–773. [Google Scholar] [CrossRef]
- Youn, D.Y.; Lee, D.H.; Lim, M.H.; Yoon, J.S.; Lim, J.H.; Jung, S.E.; Yeum, C.E.; Park, C.W.; Youn, H.J.; Lee, J.S.; et al. Bis deficiency results in early lethality with metabolic deterioration and involution of spleen and thymus. Am. J. Physiol. Endocrinol. Metab. 2008, 295, E1349–E1357. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Koren, S.A.; Cvetojevic, G.; Girardi, P.; Johnson, G.V.W. The role of BAG3 in health and disease: A “Magic BAG of Tricks”. J. Cell. Biochem. 2022, 123, 4–21. [Google Scholar] [CrossRef] [PubMed]
- Arndt, V.; Dick, N.; Tawo, R.; Dreiseidler, M.; Wenzel, D.; Hesse, M.; Furst, D.O.; Saftig, P.; Saint, R.; Fleischmann, B.K.; et al. Chaperone-assisted selective autophagy is essential for muscle maintenance. Curr. Biol. 2010, 20, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Hishiya, A.; Kitazawa, T.; Takayama, S. BAG3 and Hsc70 interact with actin capping protein CapZ to maintain myofibrillar integrity under mechanical stress. Circ. Res. 2010, 107, 1220–1231. [Google Scholar] [CrossRef]
- Gunay, K.A.; Silver, J.S.; Chang, T.L.; Bednarski, O.J.; Bannister, K.L.; Rogowski, C.J.; Olwin, B.B.; Anseth, K.S. Myoblast mechanotransduction and myotube morphology is dependent on BAG3 regulation of YAP and TAZ. Biomaterials 2021, 277, 121097. [Google Scholar] [CrossRef]
- Feldman, A.M.; Gordon, J.; Wang, J.; Song, J.; Zhang, X.Q.; Myers, V.D.; Tilley, D.G.; Gao, E.; Hoffman, N.E.; Tomar, D.; et al. BAG3 regulates contractility and Ca(2+) homeostasis in adult mouse ventricular myocytes. J. Mol. Cell. Cardiol. 2016, 92, 10–20. [Google Scholar] [CrossRef]
- Fang, X.; Bogomolovas, J.; Wu, T.; Zhang, W.; Liu, C.; Veevers, J.; Stroud, M.J.; Zhang, Z.; Ma, X.; Mu, Y.; et al. Loss-of-function mutations in co-chaperone BAG3 destabilize small HSPs and cause cardiomyopathy. J. Clin. Investig. 2017, 127, 3189–3200. [Google Scholar] [CrossRef]
- Yun, H.H.; Jung, S.Y.; Park, B.W.; Ko, J.S.; Yoo, K.; Yeo, J.; Kim, H.L.; Park, H.J.; Youn, H.J.; Lee, J.H. An Adult Mouse Model of Dilated Cardiomyopathy Caused by Inducible Cardiac-Specific Bis Deletion. Int. J. Mol. Sci. 2021, 22, 1343. [Google Scholar] [CrossRef]
- Jung, S.Y.; Yun, H.H.; Lim, J.H.; Lee, D.H.; Seo, S.B.; Baek, J.Y.; Lee, J.; Yoo, K.; Kim, H.; Kim, H.L.; et al. Hepatocyte-specific deletion of Bis causes senescence in the liver without deteriorating hepatic function. Biochem. Biophys. Res. Commun. 2022, 619, 42–48. [Google Scholar] [CrossRef]
- Mourkioti, F.; Slonimsky, E.; Huth, M.; Berno, V.; Rosenthal, N. Analysis of CRE-mediated recombination driven by myosin light chain 1/3 regulatory elements in embryonic and adult skeletal muscle: A tool to study fiber specification. Genesis 2008, 46, 424–430. [Google Scholar] [CrossRef]
- Forcina, L.; Cosentino, M.; Musaro, A. Mechanisms Regulating Muscle Regeneration: Insights into the Interrelated and Time-Dependent Phases of Tissue Healing. Cells 2020, 9, 1297. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Kuo, C.F.; Sir, D.; Wang, L.; Govindarajan, S.; Petrovic, L.M.; Ou, J.H. Autophagy inhibits oxidative stress and tumor suppressors to exert its dual effect on hepatocarcinogenesis. Cell Death Differ. 2015, 22, 1025–1034. [Google Scholar] [CrossRef] [PubMed]
- Carra, S.; Seguin, S.J.; Lambert, H.; Landry, J. HspB8 chaperone activity toward poly(Q)-containing proteins depends on its association with Bag3, a stimulator of macroautophagy. J. Biol. Chem. 2008, 283, 1437–1444. [Google Scholar] [CrossRef]
- Luo, M.; Meng, Z.; Moroishi, T.; Lin, K.C.; Shen, G.; Mo, F.; Shao, B.; Wei, X.; Zhang, P.; Wei, Y.; et al. Heat stress activates YAP/TAZ to induce the heat shock transcriptome. Nat. Cell. Biol. 2020, 22, 1447–1459. [Google Scholar] [CrossRef]
- Wu, W.; Sun, X.; Shi, X.; Lai, L.; Wang, C.; Xie, M.; Qin, G.; Qiu, H. Hsp22 Deficiency Induces Age-Dependent Cardiac Dilation and Dysfunction by Impairing Autophagy, Metabolism, and Oxidative Response. Antioxidants 2021, 10, 1550. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Wang, J.M.; Yan, J.; Zhang, D.L.; Liu, B.Q.; Jiang, J.Y.; Li, C.; Li, S.; Meng, X.N.; Wang, H.Q. BAG3 promotes autophagy and glutaminolysis via stabilizing glutaminase. Cell Death Dis. 2019, 10, 284. [Google Scholar] [CrossRef] [PubMed]
- An, M.X.; Li, S.; Yao, H.B.; Li, C.; Wang, J.M.; Sun, J.; Li, X.Y.; Meng, X.N.; Wang, H.Q. BAG3 directly stabilizes Hexokinase 2 mRNA and promotes aerobic glycolysis in pancreatic cancer cells. J. Cell Biol. 2017, 216, 4091–4105. [Google Scholar] [CrossRef] [PubMed]
- Kimura, K.; Ooms, A.; Graf-Riesen, K.; Kuppusamy, M.; Unger, A.; Schuld, J.; Daerr, J.; Lother, A.; Geisen, C.; Hein, L.; et al. Overexpression of human BAG3(P209L) in mice causes restrictive cardiomyopathy. Nat. Commun. 2021, 12, 3575. [Google Scholar] [CrossRef]
- Baldan, S.; Meriin, A.B.; Yaglom, J.; Alexandrov, I.; Varelas, X.; Xiao, Z.J.; Sherman, M.Y. The Hsp70-Bag3 complex modulates the phosphorylation and nuclear translocation of Hippo pathway protein Yap. J. Cell Sci. 2021, 134, jcs259107. [Google Scholar] [CrossRef]
- Iyer, S.R.; Shah, S.B.; Ward, C.W.; Stains, J.P.; Spangenburg, E.E.; Folker, E.S.; Lovering, R.M. Differential YAP nuclear signaling in healthy and dystrophic skeletal muscle. Am. J. Physiol. Cell Physiol. 2019, 317, C48–C57. [Google Scholar] [CrossRef]
- Watt, K.I.; Turner, B.J.; Hagg, A.; Zhang, X.; Davey, J.R.; Qian, H.; Beyer, C.; Winbanks, C.E.; Harvey, K.F.; Gregorevic, P. The Hippo pathway effector YAP is a critical regulator of skeletal muscle fibre size. Nat. Commun. 2015, 6, 6048. [Google Scholar] [CrossRef] [PubMed]
- Bouhy, D.; Juneja, M.; Katona, I.; Holmgren, A.; Asselbergh, B.; De Winter, V.; Hochepied, T.; Goossens, S.; Haigh, J.J.; Libert, C.; et al. A knock-in/knock-out mouse model of HSPB8-associated distal hereditary motor neuropathy and myopathy reveals toxic gain-of-function of mutant Hspb8. Acta Neuropathol. 2018, 135, 131–148. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, S.-Y.; Riew, T.-R.; Yun, H.H.; Lim, J.H.; Hwang, J.-W.; Jung, S.W.; Kim, H.L.; Lee, J.-S.; Lee, M.-Y.; Lee, J.-H. Skeletal Muscle-Specific Bis Depletion Leads to Muscle Dysfunction and Early Death Accompanied by Impairment in Protein Quality Control. Int. J. Mol. Sci. 2023, 24, 9635. https://doi.org/10.3390/ijms24119635
Jung S-Y, Riew T-R, Yun HH, Lim JH, Hwang J-W, Jung SW, Kim HL, Lee J-S, Lee M-Y, Lee J-H. Skeletal Muscle-Specific Bis Depletion Leads to Muscle Dysfunction and Early Death Accompanied by Impairment in Protein Quality Control. International Journal of Molecular Sciences. 2023; 24(11):9635. https://doi.org/10.3390/ijms24119635
Chicago/Turabian StyleJung, Soon-Young, Tae-Ryong Riew, Hye Hyeon Yun, Ji Hee Lim, Ji-Won Hwang, Sung Won Jung, Hong Lim Kim, Jae-Seon Lee, Mun-Yong Lee, and Jeong-Hwa Lee. 2023. "Skeletal Muscle-Specific Bis Depletion Leads to Muscle Dysfunction and Early Death Accompanied by Impairment in Protein Quality Control" International Journal of Molecular Sciences 24, no. 11: 9635. https://doi.org/10.3390/ijms24119635