
Pharmacological Analysis of GABAA Receptor and Sigma1R Chaperone Interaction: Research Report I―Investigation of the Anxiolytic, Anticonvulsant and Hypnotic Effects of Allosteric GABAA Receptors’ Ligands
 , , , , and
, , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
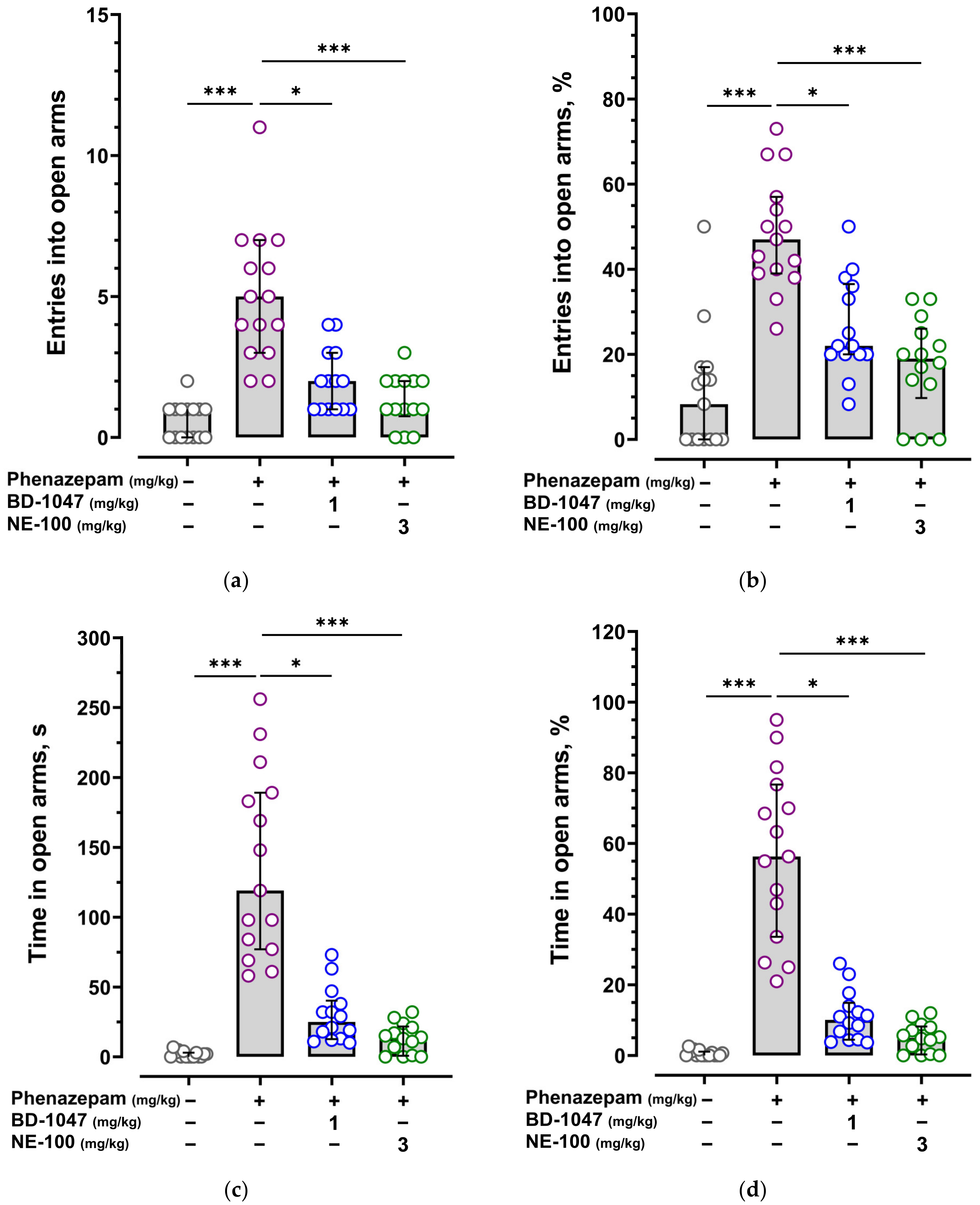
2.1. Anxiolytic Effect of Benzodiazepines
2.2. Anticonvulsant Effect of Benzodiazepines
2.3. Hypnotic Effect of Pentobarbital
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Experimental Animals
4.3. Ethical Approval
4.4. In Vivo Study
4.4.1. In Vivo Experimental Design
- (1)
- Control group: administration of vehicle 1 and vehicle 2 (n = 10);
- (2)
- Three groups: administration of BD-1047 (1, 10, and 20 mg/kg) and vehicle 2 (n = 9–11);
- (3)
- Three groups: administration of BD-1047 (1, 10, and 20 mg/kg) and diazepam 1 mg/kg (n = 5–8);
- (4)
- Administration of vehicle 1 and diazepam 1 mg/kg (n = 8).
- (1)
- Control group: administration of vehicle 1 and vehicle 2 (n = 12);
- (2)
- Two groups: administration of PRE-084 (5 and 20 mg/kg) and vehicle 2 (n = 10–12);
- (3)
- Two groups: administration of PRE-084 (1, 10, and 20 mg/kg) and diazepam 1 mg/kg (n = 9–10);
- (4)
- Administration of vehicle 1 and diazepam 1 mg/kg (n = 9).
- (1)
- Control group: administration of saline (n = 10);
- (2)
- Two groups: administration of BD-1047 (1 and 10 mg/kg), (n = 10);
- (3)
- Two groups, administration of PRE-084 (1 and 5 mg/kg), (n = 9–10);
- (4)
- Administration of diazepam 1 mg/kg (n = 10).
4.4.2. Elevated Plus Maze Test
4.4.3. Pentylenetetrazole-Induced Seizures
4.4.4. Barbiturate-Induced Sleep
4.4.5. Statistical Analysis
4.5. In Vitro Radioligand Binding Assay
4.5.1. Membrane Preparation
4.5.2. Radioligand Binding Procedure
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| (+)-SKF 10.047 | (1S,9S,13S)-1,13-dimethyl-10-prop-2-enyl-10-azatricyclo [7.3.1.02,7]trideca-2(7),3,5-trien-4-ol |
| BD-1047 | N’-[2-(3,4-dichlorophenyl)ethyl]-N,N,N’-trimethylethane-1,2-diamine |
| BiP | Endoplasmic reticulum chaperone BiP; GRP78 |
| CB1R | Cannabinoid receptors type 1 |
| E1R | (4R,5S)-2-(5-methyl-2-oxo-4-phenyl-pyrrolidin-1-yl)-acetamide |
| EPM | Elevated plus maze test |
| ER | Endoplasmic reticulum |
| GABA | γ-aminobutyric acid |
| GABAAR | GABAA receptor |
| GluN | NMDA Receptor Subunit |
| IRE1 | Serine/threonine-protein kinase/endoribonuclease IRE1 |
| MT1 receptor | Melatonin receptor, type 1 |
| MT3 receptor | Melatonin receptor, type 3 |
| NE-100 | N-[2-[4-methoxy-3-(2-phenylethoxy)phenyl]ethyl]-N-propylpropan-1-amine |
| NMDARs | The N-methyl-D-aspartate receptors |
| NQO2 | Ribosyldihydronicotinamide dehydrogenase [quinone]; NRH: quinone reductase 2; QR2 |
| OF | Open-field test |
| PAMs | Positive allosteric modulators |
| PKC | Protein kinase C |
| PRE-084 | 2-morpholin-4-ylethyl 1-phenylcyclohexane-1-carboxylate |
| PTZ | Pentylenetetrazole |
| Scn1a | Rodent sodium channel protein type 1 subunit alpha gene |
| scn1a | Zebrafish sodium channel protein type 1 subunit alpha gene |
| Sigma1R | Sigma nonopioid intracellular receptor 1; Sigma1R chaperon |
| Sigmar1 | Rodent Sigma1R gene |
| SKF83959 | 9-chloro-3-methyl-5-(3-methylphenyl)-1,2,4,5-tetrahydro-3-benzazepine-7,8-diol;hydrobromide |
| SOMCL-668 | 3-methyl-1-phenyl-2,3,4,5-tetrahydro-1H-benzo[d]azepin-7-ol |
References
- Voronin, M.V.; Vakhitova, Y.V.; Tsypysheva, I.P.; Tsypyshev, D.O.; Rybina, I.V.; Kurbanov, R.D.; Abramova, E.V.; Seredenin, S.B. Involvement of Chaperone Sigma1R in the Anxiolytic Effect of Fabomotizole. Int. J. Mol. Sci. 2021, 22, 5455. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.J.; Hibbs, R.E. Direct Structural Insights into GABA(A) Receptor Pharmacology. Trends Biochem. Sci. 2021, 46, 502–517. [Google Scholar] [CrossRef] [PubMed]
- Deutsch, S.I.; Rosse, R.B.; Mastropaolo, J. Environmental stress-induced functional modification of the central benzodiazepine binding site. Clin. Neuropharmacol. 1994, 17, 205–228. [Google Scholar] [CrossRef] [PubMed]
- Seredenin, S.B.; Blednov, I.A. A pharmacogenetic approach to the design of new selective, anxiolytic drugs. In Proceedings of the Second International Golden Ring Conference on Biological Basis of Individual Sensitivity to Psychotropic Drugs, Moscow, Russia, 22–26 May 1993; pp. 25–38. [Google Scholar]
- Seredenin, S.B.; Voronina, T.A.; Neznamov, G.G.; Blednov Iu, A.; Badyshtov, B.A.; Viglinskaia, I.V.; Kozlovskaia, M.M.; Kolotilinskaia, N.V.; Iarkova, M.A.; Savel’ev, V.L.; et al. The pharmacogenetic concept of the anxio-selective effect. Vestn. Ross Akad. Med. Nauk. 1998, 11, 3–9. [Google Scholar]
- Chekina, K.S.; Yarkova, M.A.; Seredenin, S.B. Benzodiazepine reception in C57BL/6 and BALB/c mice depending on the type of stress factor. Bull. Eksp.Biol. Med. 2009, 148, 606–608. [Google Scholar] [CrossRef]
- Yarkova, M.A.; Seredenin, S.B. Temporal characteristics of stress-induced decrease in benzodiazepine reception in C57BL/6 and BALB/c mice. Bull. Exp. Biol. Med. 2014, 157, 762–764. [Google Scholar] [CrossRef]
- Iarkova, M.A. Analysis of the binding capacity of the benzodiazepine site of gabaa receptor in mice C57BL/6 and BALB/C pretreated with anxiolytics. Eksp. Klin. Farmakol. 2011, 74, 3–7. [Google Scholar]
- Seredenin, S.B.; Iarkova, M.A.; Voronin, M.V. 3H-Diazepam binding in the brain of intact animals with various reactivity to emotional stress. Eksp. Klin. Farmakol. 2001, 64, 63–65. [Google Scholar]
- Nutt, D.J.; Malizia, A.L. New insights into the role of the GABA(A)-benzodiazepine receptor in psychiatric disorder. Br. J. Psychiatry J. Ment. Sci. 2001, 179, 390–396. [Google Scholar] [CrossRef]
- Goldschen-Ohm, M.P. Benzodiazepine Modulation of GABA(A) Receptors: A Mechanistic Perspective. Biomolecules 2022, 12, 1784. [Google Scholar] [CrossRef]
- Seredenin, S.B.; Antipova, T.A.; Voronin, M.V.; Kurchashova, S.Y.; Kuimov, A.N. Interaction of afobazole with sigma1-receptors. Bull. Exp. Biol. Med. 2009, 148, 42–44. [Google Scholar] [CrossRef] [PubMed]
- Neznamov, G.G.; Siuniakov, S.A.; Chumakov, D.V.; Bochkarev, V.K.; Seredenin, S.B. Clinical study of the selective anxiolytic agent afobazol. Eksp. Klin. Farmakol. 2001, 64, 15–19. [Google Scholar]
- Syunyakov, T.S.; Neznamov, G.G. Evaluation of the therapeutic efficacy and safety of the selective anxiolytic afobazole in generalized anxiety disorder and adjustment disorders: Results of a multicenter randomized comparative study of diazepam. Ter. Arkh. 2016, 88, 73–86. [Google Scholar] [CrossRef] [PubMed]
- Seredenin, S.B.; Voronin, M.V. Neuroreceptor mechanisms of the afobazole effect. Eksp. Klin. Farmakol. 2009, 72, 3–11. [Google Scholar]
- Seredenin, S.B.; Viglinskaia, A.O.; Mozhaeva, T.; Kolyvanov, G.B.; Litvin, A.A.; Avdiunina, N.I.; Savel’ev, V.L.; Zgerdev, V.P. Afobazole metabolism in rats. Eksp. Klin. Farmakol. 2008, 71, 50–52. [Google Scholar] [PubMed]
- Mozhaeva, T.Y.; Yarkova, M.A.; Lezina, V.P.; Seredenin, S.B. Synthesis and pharmacological activity of the main metabolite of afobazole and its analogs. Pharm. Chem. J. 2011, 45, 147. [Google Scholar] [CrossRef]
- Kilb, W. When Are Depolarizing GABAergic Responses Excitatory? Front. Mol. Neurosci. 2021, 14, 747835. [Google Scholar] [CrossRef]
- Ghit, A.; Assal, D.; Al-Shami, A.S.; Hussein, D.E.E. GABA(A) receptors: Structure, function, pharmacology, and related disorders. J. Genet, Eng. Biotechnol. 2021, 19, 123. [Google Scholar] [CrossRef]
- Sallard, E.; Letourneur, D.; Legendre, P. Electrophysiology of ionotropic GABA receptors. Cell Mol. Life Sci. 2021, 78, 5341–5370. [Google Scholar] [CrossRef]
- Kasaragod, V.B.; Mortensen, M.; Hardwick, S.W.; Wahid, A.A.; Dorovykh, V.; Chirgadze, D.Y.; Smart, T.G.; Miller, P.S. Mechanisms of inhibition and activation of extrasynaptic alphabeta GABA(A) receptors. Nature 2022, 602, 529–533. [Google Scholar] [CrossRef]
- Hortnagl, H.; Tasan, R.O.; Wieselthaler, A.; Kirchmair, E.; Sieghart, W.; Sperk, G. Patterns of mRNA and protein expression for 12 GABAA receptor subunits in the mouse brain. Neuroscience 2013, 236, 345–372. [Google Scholar] [CrossRef] [PubMed]
- Sequeira, A.; Shen, K.; Gottlieb, A.; Limon, A. Human brain transcriptome analysis finds region- and subject-specific expression signatures of GABA(A)R subunits. Commun. Biol. 2019, 2, 153. [Google Scholar] [CrossRef] [PubMed]
- Calcaterra, N.E.; Barrow, J.C. Classics in chemical neuroscience: Diazepam (valium). ACS Chem. Neurosci. 2014, 5, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Masiulis, S.; Desai, R.; Uchanski, T.; Serna Martin, I.; Laverty, D.; Karia, D.; Malinauskas, T.; Zivanov, J.; Pardon, E.; Kotecha, A.; et al. GABA(A) receptor signalling mechanisms revealed by structural pharmacology. Nature 2019, 565, 454–459. [Google Scholar] [CrossRef]
- Olsen, R.W. GABA(A) receptor: Positive and negative allosteric modulators. Neuropharmacology 2018, 136, 10–22. [Google Scholar] [CrossRef]
- Ramanjaneyulu, R.; Ticku, M.K. Interactions of pentamethylenetetrazole and tetrazole analogues with the picrotoxinin site of the benzodiazepine-GABA receptor-ionophore complex. Eur. J. Pharmacol. 1984, 98, 337–345. [Google Scholar] [CrossRef]
- Huang, R.Q.; Bell-Horner, C.L.; Dibas, M.I.; Covey, D.F.; Drewe, J.A.; Dillon, G.H. Pentylenetetrazole-induced inhibition of recombinant gamma-aminobutyric acid type A (GABA(A)) receptors: Mechanism and site of action. J. Pharmacol. Exp. Ther. 2001, 298, 986–995. [Google Scholar]
- Fisher, R.S. Animal models of the epilepsies. Brain Research. Brain Res. Rev. 1989, 14, 245–278. [Google Scholar] [CrossRef]
- Schmidt, H.R.; Betz, R.M.; Dror, R.O.; Kruse, A.C. Structural basis for sigma1 receptor ligand recognition. Nat. Struct. Mol. Biol. 2018, 25, 981–987. [Google Scholar] [CrossRef]
- Schmidt, H.R.; Zheng, S.; Gurpinar, E.; Koehl, A.; Manglik, A.; Kruse, A.C. Crystal structure of the human sigma1 receptor. Nature 2016, 532, 527–530. [Google Scholar] [CrossRef]
- Ortega-Roldan, J.L.; Ossa, F.; Schnell, J.R. Characterization of the human sigma-1 receptor chaperone domain structure and binding immunoglobulin protein (BiP) interactions. J. Biol. Chem. 2013, 288, 21448–21457. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Xiao, Y.; Ji, Y.; Sun, Z.; Zhou, X. An open-like conformation of the sigma-1 receptor reveals its ligand entry pathway. Nat. Commun. 2022, 13, 1267. [Google Scholar] [CrossRef]
- Aishwarya, R.; Abdullah, C.S.; Morshed, M.; Remex, N.S.; Bhuiyan, M.S. Sigmar1’s Molecular, Cellular, and Biological Functions in Regulating Cellular Pathophysiology. Front. Physiol. 2021, 12, 705575. [Google Scholar] [CrossRef] [PubMed]
- Ossa, F.; Schnell, J.R.; Ortega-Roldan, J.L. A Review of the Human Sigma-1 Receptor Structure. Adv. Exp. Med. Biol. 2017, 964, 15–29. [Google Scholar] [CrossRef] [PubMed]
- Laurini, E.; Marson, D.; Fermeglia, M.; Pricl, S. 3D Homology Model of Sigma1 Receptor. Handb. Exp. Pharmacol. 2017, 244, 27–50. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Guo, Q.; Fang, L.P.; Yao, H.; Scheller, A.; Kirchhoff, F.; Huang, W. Specific detection and deletion of the sigma-1 receptor widely expressed in neurons and glial cells in vivo. J. Neurochem. 2022. [Google Scholar] [CrossRef]
- Hayashi, T.; Su, T.P. Sigma-1 receptor chaperones at the ER-mitochondrion interface regulate Ca(2+) signaling and cell survival. Cell 2007, 131, 596–610. [Google Scholar] [CrossRef]
- Mori, T.; Hayashi, T.; Hayashi, E.; Su, T.P. Sigma-1 receptor chaperone at the ER-mitochondrion interface mediates the mitochondrion-ER-nucleus signaling for cellular survival. PLoS ONE 2013, 8, e76941. [Google Scholar] [CrossRef]
- Sharma, N.; Patel, C.; Shenkman, M.; Kessel, A.; Ben-Tal, N.; Lederkremer, G.Z. The Sigma-1 receptor is an ER-localized type II membrane protein. J. Biol. Chem. 2021, 297, 101299. [Google Scholar] [CrossRef]
- Bhattacharyya, R.; Black, S.E.; Lotlikar, M.S.; Fenn, R.H.; Jorfi, M.; Kovacs, D.M.; Tanzi, R.E. Axonal generation of amyloid-beta from palmitoylated APP in mitochondria-associated endoplasmic reticulum membranes. Cell Rep. 2021, 35, 109134. [Google Scholar] [CrossRef]
- Zhemkov, V.; Ditlev, J.A.; Lee, W.R.; Wilson, M.; Liou, J.; Rosen, M.K.; Bezprozvanny, I. The role of sigma 1 receptor in organization of endoplasmic reticulum signaling microdomains. eLife 2021, 10, e65192. [Google Scholar] [CrossRef] [PubMed]
- Su, T.P.; Su, T.C.; Nakamura, Y.; Tsai, S.Y. The Sigma-1 Receptor as a Pluripotent Modulator in Living Systems. Trends Pharmacol. Sci. 2016, 37, 262–278. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, H.R.; Kruse, A.C. The Molecular Function of sigma Receptors: Past, Present, and Future. Trends Pharmacol. Sci. 2019, 40, 636–654. [Google Scholar] [CrossRef]
- Salaciak, K.; Pytka, K. Revisiting the sigma-1 receptor as a biological target to treat affective and cognitive disorders. Neurosci. Biobehav. Rev. 2021, 132, 1114–1136. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Veeranan-Karmegam, R.; Baker, F.C.; Mysona, B.A.; Bagchi, P.; Liu, Y.; Smith, S.B.; Gonsalvez, G.B.; Bollinger, K.E. Defining the Ligand-dependent Interactome of the Sigma 1 Receptor. bioRxiv 2022. [Google Scholar] [CrossRef]
- Chevallier, N.; Keller, E.; Maurice, T. Behavioural phenotyping of knockout mice for the sigma-1 (sigma(1)) chaperone protein revealed gender-related anxiety, depressive-like and memory alterations. J. Psychopharmacol. 2011, 25, 960–975. [Google Scholar] [CrossRef]
- Kamei, H.; Noda, Y.; Kameyama, T.; Nabeshima, T. Role of (+)-SKF-10,047-sensitive sub-population of sigma 1 receptors in amelioration of conditioned fear stress in rats: Association with mesolimbic dopaminergic systems. Eur. J. Pharmacol. 1997, 319, 165–172. [Google Scholar] [CrossRef]
- Kamei, H.; Kameyama, T.; Nabeshima, T. SKF-10,047 reverses stress-induced motor suppression: Interaction with dopaminergic system. Eur. J. Pharmacol. 1994, 260, 39–46. [Google Scholar] [CrossRef]
- Kamei, H.; Kameyama, T.; Nabeshima, T. (+)-SKF-10,047 and dextromethorphan ameliorate conditioned fear stress through the activation of phenytoin-regulated sigma 1 sites. Eur. J. Pharmacol. 1996, 299, 21–28. [Google Scholar] [CrossRef]
- Kamei, H.; Kameyama, T.; Nabeshima, T. (+)-SKF-10,047 and dextromethorphan ameliorate conditioned fear stress via dopaminergic systems linked to phenytoin-regulated sigma 1 sites. Eur. J. Pharmacol. 1996, 309, 149–158. [Google Scholar] [CrossRef]
- Noda, Y.; Kamei, H.; Kamei, Y.; Nagai, T.; Nishida, M.; Nabeshima, T. Neurosteroids ameliorate conditioned fear stress: An association with sigma receptors. Neuropsychopharmacology 2000, 23, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Vavers, E.; Zvejniece, B.; Stelfa, G.; Svalbe, B.; Vilks, K.; Kupats, E.; Mezapuke, R.; Lauberte, L.; Dambrova, M.; Zvejniece, L. Genetic inactivation of the sigma-1 chaperone protein results in decreased expression of the R2 subunit of the GABA-B receptor and increased susceptibility to seizures. Neurobiol. Dis. 2021, 150, 105244. [Google Scholar] [CrossRef] [PubMed]
- Johnston, G.A. Advantages of an antagonist: Bicuculline and other GABA antagonists. Br. J. Pharmacol. 2013, 169, 328–336. [Google Scholar] [CrossRef]
- Zhang, B.; Wang, L.; Chen, T.; Hong, J.; Sha, S.; Wang, J.; Xiao, H.; Chen, L. Sigma-1 receptor deficiency reduces GABAergic inhibition in the basolateral amygdala leading to LTD impairment and depressive-like behaviors. Neuropharmacology 2017, 116, 387–398. [Google Scholar] [CrossRef]
- Vavers, E.; Svalbe, B.; Lauberte, L.; Stonans, I.; Misane, I.; Dambrova, M.; Zvejniece, L. The activity of selective sigma-1 receptor ligands in seizure models in vivo. Behav. Brain Res. 2017, 328, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Pozdnyakova, N.; Krisanova, N.; Dudarenko, M.; Vavers, E.; Zvejniece, L.; Dambrova, M.; Borisova, T. Inhibition of sigma-1 receptors substantially modulates GABA and glutamate transport in presynaptic nerve terminals. Exp. Neurol. 2020, 333, 113434. [Google Scholar] [CrossRef]
- Muller, W.E.; Siebert, B.; Holoubek, G.; Gentsch, C. Neuropharmacology of the anxiolytic drug opipramol, a sigma site ligand. Pharmacopsychiatry 2004, 37, S189–S197. [Google Scholar] [CrossRef]
- Keshavarz, M.; Yekzaman, B. Amelioration of Pentylenetetrazole-Induced Seizures by Modulators of Sigma, N-Methyl-D-Aspartate, and Ryanodine Receptors in Mice. Iran J. Med. Sci. 2018, 43, 195–201. [Google Scholar]
- Thurgur, C.; Church, J. The anticonvulsant actions of sigma receptor ligands in the Mg2+-free model of epileptiform activity in rat hippocampal slices. Br. J. Pharmacol. 1998, 124, 917–929. [Google Scholar] [CrossRef]
- Ishima, T.; Fujita, Y.; Hashimoto, K. Interaction of new antidepressants with sigma-1 receptor chaperones and their potentiation of neurite outgrowth in PC12 cells. Eur. J. Pharmacol. 2014, 727, 167–173. [Google Scholar] [CrossRef]
- Voronin, M.V.; Vakhitova, Y.V.; Seredenin, S.B. Chaperone Sigma1R and Antidepressant Effect. Int. J. Mol. Sci. 2020, 21, 7088. [Google Scholar] [CrossRef] [PubMed]
- Olsen, R.W. Picrotoxin-like channel blockers of GABAA receptors. Proc. Natl. Acad. Sci. USA 2006, 103, 6081–6082. [Google Scholar] [CrossRef]
- Hasebe, S.; Ago, Y.; Watabe, Y.; Oka, S.; Hiramatsu, N.; Tanaka, T.; Umehara, C.; Hashimoto, H.; Takuma, K.; Matsuda, T. Anti-anhedonic effect of selective serotonin reuptake inhibitors with affinity for sigma-1 receptors in picrotoxin-treated mice. Br. J. Pharmacol. 2017, 174, 314–327. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Chen, Y.; Zhao, R.; Wang, G.; Friedman, E.; Zhang, A.; Zhen, X. Allosteric modulation of sigma-1 receptors elicits anti-seizure activities. Br. J. Pharmacol. 2015, 172, 4052–4065. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.C.; Bing, G.; Jhoo, W.K.; Kim, W.K.; Shin, E.J.; Im, D.H.; Kang, K.S.; Ko, K.H. Metabolism to dextrorphan is not essential for dextromethorphan’s anticonvulsant activity against kainate in mice. Life Sci. 2003, 72, 769–783. [Google Scholar] [CrossRef] [PubMed]
- Martin, P.; Reeder, T.; Sourbron, J.; de Witte, P.A.M.; Gammaitoni, A.R.; Galer, B.S. An Emerging Role for Sigma-1 Receptors in the Treatment of Developmental and Epileptic Encephalopathies. Int. J. Mol. Sci. 2021, 22, 8416. [Google Scholar] [CrossRef]
- Sourbron, J.; Smolders, I.; de Witte, P.; Lagae, L. Pharmacological Analysis of the Anti-epileptic Mechanisms of Fenfluramine in scn1a Mutant Zebrafish. Front. Pharmacol. 2017, 8, 191. [Google Scholar] [CrossRef]
- Brunklaus, A.; Zuberi, S.M. Dravet syndrome—From epileptic encephalopathy to channelopathy. Epilepsia 2014, 55, 979–984. [Google Scholar] [CrossRef]
- Sachidanandan, D.; Reddy, H.P.; Mani, A.; Hyde, G.J.; Bera, A.K. The Neuropeptide Orexin-A Inhibits the GABA(A) Receptor by PKC and Ca(2+)/CaMKII-Dependent Phosphorylation of Its beta(1) Subunit. J. Mol. Neurosci. 2017, 61, 459–467. [Google Scholar] [CrossRef]
- Barr, J.L.; Zhao, P.; Brailoiu, G.C.; Brailoiu, E. Choline-Sigma-1R as an Additional Mechanism for Potentiation of Orexin by Cocaine. Int. J. Mol. Sci. 2021, 22, 5160. [Google Scholar] [CrossRef]
- Hafycz, J.M.; Strus, E.; Naidoo, N. Reducing ER stress with chaperone therapy reverses sleep fragmentation and cognitive decline in aged mice. Aging Cell 2022, 21, e13598. [Google Scholar] [CrossRef] [PubMed]
- Tobler, I.; Kopp, C.; Deboer, T.; Rudolph, U. Diazepam-induced changes in sleep: Role of the alpha 1 GABA(A) receptor subtype. Proc. Natl. Acad. Sci. USA 2001, 98, 6464–6469. [Google Scholar] [CrossRef] [PubMed]
- Greenfield, L.J., Jr. Molecular mechanisms of antiseizure drug activity at GABAA receptors. Seizure 2013, 22, 589–600. [Google Scholar] [CrossRef] [PubMed]
- Engin, E.; Benham, R.S.; Rudolph, U. An Emerging Circuit Pharmacology of GABA(A) Receptors. Trends Pharmacol. Sci. 2018, 39, 710–732. [Google Scholar] [CrossRef]
- Akk, G.; Bracamontes, J.; Steinbach, J.H. Activation of GABA(A) receptors containing the alpha4 subunit by GABA and pentobarbital. J. Physiol. 2004, 556, 387–399. [Google Scholar] [CrossRef]
- Feng, H.J.; Macdonald, R.L. Barbiturates require the N terminus and first transmembrane domain of the delta subunit for enhancement of alpha1beta3delta GABAA receptor currents. J. Biol. Chem. 2010, 285, 23614–23621. [Google Scholar] [CrossRef]
- Brickley, S.G.; Mody, I. Extrasynaptic GABA(A) receptors: Their function in the CNS and implications for disease. Neuron 2012, 73, 23–34. [Google Scholar] [CrossRef]
- Wafford, K.A. The Pharmacology of Extrasynaptic GABAA Receptors. In Extrasynaptic GABAA Receptors; Errington, A.C., Di Giovanni, G., Crunelli, V., Eds.; Springer: New York, NY, USA, 2014; pp. 51–74. [Google Scholar] [CrossRef]
- Fritschy, J.-M.; Panzanelli, P. Extrasynaptic GABAA Receptors: Subunit Composition, Distribution, and Regulation. In Extrasynaptic GABAA Receptors; Errington, A.C., Di Giovanni, G., Crunelli, V., Eds.; Springer: New York, NY, USA, 2014; pp. 15–32. [Google Scholar] [CrossRef]
- Hayashi, T.; Su, T.P. Intracellular dynamics of sigma-1 receptors (sigma(1) binding sites) in NG108-15 cells. J. Pharmacol. Exp. Ther. 2003, 306, 726–733. [Google Scholar] [CrossRef]
- Kopanchuk, S.; Vavers, E.; Veiksina, S.; Ligi, K.; Zvejniece, L.; Dambrova, M.; Rinken, A. Intracellular dynamics of the Sigma-1 receptor observed with super-resolution imaging microscopy. PLoS ONE 2022, 17, e0268563. [Google Scholar] [CrossRef]
- Qin, Y.; Xu, W.; Li, K.; Luo, Q.; Chen, X.; Wang, Y.; Chen, L.; Sha, S. Repeated inhibition of sigma-1 receptor suppresses GABAA receptor expression and long-term depression in the nucleus accumbens leading to depressive-like behaviors. Front. Mol. Neurosci. 2022, 15, 959224. [Google Scholar] [CrossRef]
- Sanchez-Blazquez, P.; Rodriguez-Munoz, M.; Herrero-Labrador, R.; Burgueno, J.; Zamanillo, D.; Garzon, J. The calcium-sensitive Sigma-1 receptor prevents cannabinoids from provoking glutamate NMDA receptor hypofunction: Implications in antinociception and psychotic diseases. Int. J. Neuropsychopharmacol. Off. Sci. J. Coll. Int. Neuropsychopharmacol. (CINP) 2014, 17, 1943–1955. [Google Scholar] [CrossRef] [PubMed]
- Haller, J.; Varga, B.; Ledent, C.; Freund, T.F. CB1 cannabinoid receptors mediate anxiolytic effects: Convergent genetic and pharmacological evidence with CB1-specific agents. Behav. Pharmacol. 2004, 15, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Munoz, M.; Onetti, Y.; Cortes-Montero, E.; Garzon, J.; Sanchez-Blazquez, P. Cannabidiol enhances morphine antinociception, diminishes NMDA-mediated seizures and reduces stroke damage via the sigma 1 receptor. Mol. Brain 2018, 11, 51. [Google Scholar] [CrossRef] [PubMed]
- Balasuriya, D.; Stewart, A.P.; Edwardson, J.M. The sigma-1 receptor interacts directly with GluN1 but not GluN2A in the GluN1/GluN2A NMDA receptor. J. Neurosci. 2013, 33, 18219–18224. [Google Scholar] [CrossRef]
- Pabba, M.; Wong, A.Y.; Ahlskog, N.; Hristova, E.; Biscaro, D.; Nassrallah, W.; Ngsee, J.K.; Snyder, M.; Beique, J.C.; Bergeron, R. NMDA receptors are upregulated and trafficked to the plasma membrane after sigma-1 receptor activation in the rat hippocampus. J. Neurosci. 2014, 34, 11325–11338. [Google Scholar] [CrossRef]
- Bina, P.; Rezvanfard, M.; Ahmadi, S.; Zarrindast, M.R. Anxiolytic-Like Effects and Increase in Locomotor Activity Induced by Infusions of NMDA into the Ventral Hippocampus in Rat: Interaction with GABAergic System. Basic Clin. Neurosci. 2014, 5, 267–276. [Google Scholar]
- Boyce-Rustay, J.M.; Holmes, A. Genetic inactivation of the NMDA receptor NR2A subunit has anxiolytic- and antidepressant-like effects in mice. Neuropsychopharmacology 2006, 31, 2405–2414. [Google Scholar] [CrossRef]
- Chen, S.; Xu, D.; Fan, L.; Fang, Z.; Wang, X.; Li, M. Roles of N-Methyl-D-Aspartate Receptors (NMDARs) in Epilepsy. Front. Mol. Neurosci. 2021, 14, 797253. [Google Scholar] [CrossRef]
- Poleszak, E.; Socala, K.; Szopa, A.; Wrobel, A.; Szewczyk, B.; Kasperek, R.; Blicharska, E.; Nowak, G.; Wlaz, P. Involvement of NMDA receptor complex in the anxiolytic-like effects of chlordiazepoxide in mice. J. Neural. Transm. 2011, 118, 857–864. [Google Scholar] [CrossRef]
- Garcia-Gutierrez, M.S.; Manzanares, J. The cannabinoid CB1 receptor is involved in the anxiolytic, sedative and amnesic actions of benzodiazepines. J. Psychopharmacol. 2010, 24, 757–765. [Google Scholar] [CrossRef]
- Fujimoto, M.; Hayashi, T.; Urfer, R.; Mita, S.; Su, T.P. Sigma-1 receptor chaperones regulate the secretion of brain-derived neurotrophic factor. Synapse 2012, 66, 630–639. [Google Scholar] [CrossRef] [PubMed]
- Ha, Y.; Dun, Y.; Thangaraju, M.; Duplantier, J.; Dong, Z.; Liu, K.; Ganapathy, V.; Smith, S.B. Sigma receptor 1 modulates endoplasmic reticulum stress in retinal neurons. Investig. Ophthalmol. Vis. Sci. 2011, 52, 527–540. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Munoz, M.; Cortes-Montero, E.; Garzon-Nino, J.; Sanchez-Blazquez, P. The ALS-related sigma1R E102Q Mutant Eludes Ligand Control and Exhibits Anomalous Response to Calcium. Int. J. Mol. Sci. 2020, 21, 7339. [Google Scholar] [CrossRef] [PubMed]
- Kopp, M.C.; Larburu, N.; Durairaj, V.; Adams, C.J.; Ali, M.M.U. UPR proteins IRE1 and PERK switch BiP from chaperone to ER stress sensor. Nat. Struct. Mol. Biol. 2019, 26, 1053–1062. [Google Scholar] [CrossRef] [PubMed]
- Rosen, D.A.; Seki, S.M.; Fernandez-Castaneda, A.; Beiter, R.M.; Eccles, J.D.; Woodfolk, J.A.; Gaultier, A. Modulation of the sigma-1 receptor-IRE1 pathway is beneficial in preclinical models of inflammation and sepsis. Sci. Transl. Med. 2019, 11, eaau5266. [Google Scholar] [CrossRef]
- Connolly, C.N.; Krishek, B.J.; McDonald, B.J.; Smart, T.G.; Moss, S.J. Assembly and cell surface expression of heteromeric and homomeric gamma-aminobutyric acid type A receptors. J. Biol. Chem. 1996, 271, 89–96. [Google Scholar] [CrossRef]
- Whittsette, A.L.; Wang, Y.J.; Mu, T.W. The endoplasmic reticulum membrane complex promotes proteostasis of GABAA receptors. iScience 2022, 25, 104754. [Google Scholar] [CrossRef]
- Yuan, B.; Hatchett-Walker, C.; Long, P.; Xu, Z.; Stephenson, F.A.; Haider, S.; Jovanovic, J.N. Control of cell surface expression of GABA(A) receptors by a conserved region at the end of the N-terminal extracellular domain of receptor subunits. J. Biol. Chem. 2022, 298, 102590. [Google Scholar] [CrossRef]
- Wang, Y.J.; Di, X.J.; Mu, T.W. Quantitative interactome proteomics identifies a proteostasis network for GABAA receptors. J. Biol. Chem. 2022, 298, 102423. [Google Scholar] [CrossRef]
- Di, X.J.; Han, D.Y.; Wang, Y.J.; Chance, M.R.; Mu, T.W. SAHA enhances Proteostasis of epilepsy-associated alpha1(A322D)beta2gamma2 GABA(A) receptors. Chem. Biol. 2013, 20, 1456–1468. [Google Scholar] [CrossRef]
- Fu, Y.L.; Han, D.Y.; Wang, Y.J.; Di, X.J.; Yu, H.B.; Mu, T.W. Remodeling the endoplasmic reticulum proteostasis network restores proteostasis of pathogenic GABAA receptors. PLoS ONE 2018, 13, e0207948. [Google Scholar] [CrossRef] [PubMed]
- Di, X.J.; Wang, Y.J.; Cotter, E.; Wang, M.; Whittsette, A.L.; Han, D.Y.; Sangwung, P.; Brown, R.; Lynch, J.W.; Keramidas, A.; et al. Proteostasis Regulators Restore Function of Epilepsy-Associated GABAA Receptors. Cell Chem. Biol. 2021, 28, 46–59.e47. [Google Scholar] [CrossRef] [PubMed]
- Bown, C.D.; Wang, J.F.; Chen, B.; Young, L.T. Regulation of ER stress proteins by valproate: Therapeutic implications. Bipolar Disord. 2002, 4, 145–151. [Google Scholar] [CrossRef]
- Winterer, G. Valproate and GABAergic system effects. Neuropsychopharmacology 2003, 28, 2050–2051. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, G. The mechanisms of action of valproate in neuropsychiatric disorders: Can we see the forest for the trees? Cell Mol. Life Sci. 2007, 64, 2090–2103. [Google Scholar] [CrossRef]
- Monnet, F.P. Sigma-1 receptor as regulator of neuronal intracellular Ca2+: Clinical and therapeutic relevance. Biol. Cell Under Auspices Eur. Cell Biol. Organ. 2005, 97, 873–883. [Google Scholar] [CrossRef] [PubMed]
- Krebs, J.; Agellon, L.B.; Michalak, M. Ca(2+) homeostasis and endoplasmic reticulum (ER) stress: An integrated view of calcium signaling. Biochem. Biophys. Res. Commun. 2015, 460, 114–121. [Google Scholar] [CrossRef]
- Voronin, M.V.; Abramova, E.V.; Verbovaya, E.R.; Vakhitova, Y.V.; Seredenin, S.B. Chaperone-Dependent Mechanisms as a Pharmacological Target for Neuroprotection. Int. J. Mol. Sci. 2023, 24, 823. [Google Scholar] [CrossRef]
- Morin-Surun, M.P.; Collin, T.; Denavit-Saubie, M.; Baulieu, E.E.; Monnet, F.P. Intracellular sigma1 receptor modulates phospholipase C and protein kinase C activities in the brainstem. Proc. Natl. Acad. Sci. USA 1999, 96, 8196–8199. [Google Scholar] [CrossRef]
- Leidenheimer, N.J.; Whiting, P.J.; Harris, R.A. Activation of calcium-phospholipid-dependent protein kinase enhances benzodiazepine and barbiturate potentiation of the GABAA receptor. J. Neurochem. 1993, 60, 1972–1975. [Google Scholar] [CrossRef]
- Di, T.; Zhang, S.; Hong, J.; Zhang, T.; Chen, L. Hyperactivity of Hypothalamic-Pituitary-Adrenal Axis Due to Dysfunction of the Hypothalamic Glucocorticoid Receptor in Sigma-1 Receptor Knockout Mice. Front. Mol. Neurosci. 2017, 10, 287. [Google Scholar] [CrossRef] [PubMed]
- De Koninck, Y.; Mody, I. The effects of raising intracellular calcium on synaptic GABAA receptor-channels. Neuropharmacology 1996, 35, 1365–1374. [Google Scholar] [CrossRef] [PubMed]
- Taleb, O.; Trouslard, J.; Demeneix, B.A.; Feltz, P.; Bossu, J.L.; Dupont, J.L.; Feltz, A. Spontaneous and GABA-evoked chloride channels on pituitary intermediate lobe cells and their internal Ca requirements. Pflug. Arch. 1987, 409, 620–631. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.G.; Choi, S.R.; Kang, D.W.; Kim, J.; Park, J.B.; Lee, J.H.; Kim, H.W. Sigma-1 receptor increases intracellular calcium in cultured astrocytes and contributes to mechanical allodynia in a model of neuropathic pain. Brain Res. Bull. 2022, 178, 69–81. [Google Scholar] [CrossRef]
- Hayashi, T.; Su, T.P. Regulating ankyrin dynamics: Roles of sigma-1 receptors. Proc. Natl. Acad. Sci. USA 2001, 98, 491–496. [Google Scholar] [CrossRef]
- Hong, W.; Nuwayhid, S.J.; Werling, L.L. Modulation of bradykinin-induced calcium changes in SH-SY5Y cells by neurosteroids and sigma receptor ligands via a shared mechanism. Synapse 2004, 54, 102–110. [Google Scholar]
- Tchedre, K.T.; Huang, R.Q.; Dibas, A.; Krishnamoorthy, R.R.; Dillon, G.H.; Yorio, T. Sigma-1 receptor regulation of voltage-gated calcium channels involves a direct interaction. Investig. Ophthalmol. Vis. Sci. 2008, 49, 4993–5002. [Google Scholar] [CrossRef]
- Gravielle, M.C. Regulation of GABA(A) Receptors Induced by the Activation of L-Type Voltage-Gated Calcium Channels. Membranes 2021, 11, 486. [Google Scholar] [CrossRef]
- Blazon, M.; LaCarubba, B.; Bunda, A.; Czepiel, N.; Mallat, S.; Londrigan, L.; Andrade, A. N-type calcium channels control GABAergic transmission in brain areas related to fear and anxiety. OBM Neurobiol. 2021, 5. [Google Scholar] [CrossRef]
- Hayashi, T.; Su, T.P. Sigma-1 receptors (sigma(1) binding sites) form raft-like microdomains and target lipid droplets on the endoplasmic reticulum: Roles in endoplasmic reticulum lipid compartmentalization and export. J. Pharmacol. Exp. Ther. 2003, 306, 718–725. [Google Scholar] [CrossRef]
- Takebayashi, M.; Hayashi, T.; Su, T.P. Sigma-1 receptors potentiate epidermal growth factor signaling towards neuritogenesis in PC12 cells: Potential relation to lipid raft reconstitution. Synapse 2004, 53, 90–103. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Su, T.P. Sigma-1 receptors at galactosylceramide-enriched lipid microdomains regulate oligodendrocyte differentiation. Proc. Natl. Acad. Sci. USA 2004, 101, 14949–14954. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Su, T.P. The potential role of sigma-1 receptors in lipid transport and lipid raft reconstitution in the brain: Implication for drug abuse. Life Sci. 2005, 77, 1612–1624. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Su, T.P. Cholesterol at the endoplasmic reticulum: Roles of the sigma-1 receptor chaperone and implications thereof in human diseases. Subcell Biochem. 2010, 51, 381–398. [Google Scholar] [CrossRef] [PubMed]
- Lingwood, D.; Simons, K. Lipid rafts as a membrane-organizing principle. Science 2010, 327, 46–50. [Google Scholar] [CrossRef]
- Sooksawate, T.; Simmonds, M.A. Effects of membrane cholesterol on the sensitivity of the GABA(A) receptor to GABA in acutely dissociated rat hippocampal neurones. Neuropharmacology 2001, 40, 178–184. [Google Scholar] [CrossRef]
- Sogaard, R.; Werge, T.M.; Bertelsen, C.; Lundbye, C.; Madsen, K.L.; Nielsen, C.H.; Lundbaek, J.A. GABA(A) receptor function is regulated by lipid bilayer elasticity. Biochemistry 2006, 45, 13118–13129. [Google Scholar] [CrossRef]
- Nothdurfter, C.; Tanasic, S.; Di Benedetto, B.; Uhr, M.; Wagner, E.M.; Gilling, K.E.; Parsons, C.G.; Rein, T.; Holsboer, F.; Rupprecht, R.; et al. Lipid raft integrity affects GABAA receptor, but not NMDA receptor modulation by psychopharmacological compounds. Int. J. Neuropsychopharmacol. Off. Sci. J. Coll. Int. Neuropsychopharmacol. (CINP) 2013, 16, 1361–1371. [Google Scholar] [CrossRef]
- Sierra-Valdez, F.J.; Ruiz-Suarez, J.C.; Delint-Ramirez, I. Pentobarbital modifies the lipid raft-protein interaction: A first clue about the anesthesia mechanism on NMDA and GABAA receptors. Biochim. Biophys. Acta 2016, 1858, 2603–2610. [Google Scholar] [CrossRef]
- GOST 33215-2014; Guidelines for Accommodation and Care of Animals. Environment, Housing and Management. In Interstate Counsil for Standartization, Metrology and Certification. Standartinform: Moscow, Russia, 2016. Available online: http://protect.gost.ru/document.aspx?control=7&id=202494 (accessed on 28 May 2023).
- National Research Council (U.S.); Committee for the Update of the Guide for the Care and Use of Laboratory Animals; Institute for Laboratory Animal Research (U.S.); National Academies Press (U.S.). Guide for the Care and Use of Laboratory Animals, 8th ed.; National Academies Press: Washington, DC, USA, 2011; p. 220. [Google Scholar]
- Hubrecht, R.C.; Carter, E. The 3Rs and Humane Experimental Technique: Implementing Change. Animals 2019, 9, 754. [Google Scholar] [CrossRef]
- Bastami, Z.; Soltanpour, S.; Panahi-Azar, V.; Jouyban, A. Solubility of clonazepam and diazepam in binary and ternary mixtures of polyethylene glycols 400 or 600, propylene glycol and water at 298.2K—Experimental data and modeling. J. Serb. Chem. Soc. 2014, 79, 445–456. [Google Scholar] [CrossRef]
- Polyethylene glycol [MAK Value Documentation, 1998]. In The MAK-Collection for Occupational Health and Safety; Wiley: Hoboken, NJ, USA, 2012; pp. 248–270. [CrossRef]
- Huang, L.-F.; Dong, J. Formulation Strategies and Practice Used for Drug Candidates with Water-Insoluble Properties for Toxicology, Biology, and Pharmacology Studies in Discovery Support. In Water-Insoluble Drug Formulation, 2nd ed.; Liu, R., Ed.; CRC Press: Boca Raton, FL, USA, 2008; pp. 113–132. [Google Scholar] [CrossRef]
- Griebel, G.; Belzung, C.; Perrault, G.; Sanger, D.J. Differences in anxiety-related behaviours and in sensitivity to diazepam in inbred and outbred strains of mice. Psychopharmacology 2000, 148, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Lepicard, E.M.; Joubert, C.; Hagneau, I.; Perez-Diaz, F.; Chapouthier, G. Differences in anxiety-related behavior and response to diazepam in BALB/cByJ and C57BL/6J strains of mice. Pharmacol. Biochem. Behav. 2000, 67, 739–748. [Google Scholar] [CrossRef] [PubMed]
- Seredenin, S.B.; Nadorova, A.V.; Kolik, L.G.; Yarkova, M.A. Effects of phenazepam on the behavior of C57BL/6 and BALB/c mice in the open field test after naloxone pretreatment. Bull. Exp. Biol. Med. 2013, 155, 346–349. [Google Scholar] [CrossRef]
- Seo, S.Y.; Kim, S.P.; Bang, S.K.; Kang, S.Y.; Cho, S.J.; Choi, K.H.; Ryu, Y. The effect of acupuncture stimulation on alleviating emotional changes due to acute alcohol administration and the possibility of sigma(1) receptor involvement. Integr. Med. Res. 2021, 10, 100497. [Google Scholar] [CrossRef]
- Maurice, T.; Casalino, M.; Lacroix, M.; Romieu, P. Involvement of the sigma 1 receptor in the motivational effects of ethanol in mice. Pharmacol. Biochem. Behav. 2003, 74, 869–876. [Google Scholar] [CrossRef]
- Malik, M.; Rangel-Barajas, C.; Mach, R.H.; Luedtke, R.R. The effect of the sigma-1 receptor selective compound LS-1-137 on the DOI-induced head twitch response in mice. Pharmacol. Biochem. Behav. 2016, 148, 136–144. [Google Scholar] [CrossRef]
- Griesmaier, E.; Posod, A.; Gross, M.; Neubauer, V.; Wegleiter, K.; Hermann, M.; Urbanek, M.; Keller, M.; Kiechl-Kohlendorfer, U. Neuroprotective effects of the sigma-1 receptor ligand PRE-084 against excitotoxic perinatal brain injury in newborn mice. Exp. Neurol. 2012, 237, 388–395. [Google Scholar] [CrossRef]
- Hedrich, H.J. The Laboratory Mouse, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2012; pp. 731–732. [Google Scholar]
- Raskovic, A.; Horvat, O.; Jakovljevic, V.; Sabo, J.; Vasic, R. Interaction of alcoholic extracts of hops with pentobarbital and diazepam in mice. Eur. J. Drug Metab. Pharmacokinet. 2007, 32, 45–49. [Google Scholar] [CrossRef]
- File, S.E. Factors controlling measures of anxiety and responses to novelty in the mouse. Behav. Brain Res. 2001, 125, 151–157. [Google Scholar] [CrossRef]
- Mandhane, S.N.; Aavula, K.; Rajamannar, T. Timed pentylenetetrazol infusion test: A comparative analysis with s.c.PTZ and MES models of anticonvulsant screening in mice. Seizure 2007, 16, 636–644. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.H.; Chan, M.H. Developmental lead exposure differentially alters the susceptibility to chemoconvulsants in rats. Toxicology 2002, 173, 249–257. [Google Scholar] [CrossRef]
- Mesdaghinia, A.; Yazdanpanah, H.; Seddighi, M.; Banafshe, H.R.; Heydari, A. Effect of short-term lead exposure on PTZ-induced seizure threshold in mice. Toxicol. Lett. 2010, 199, 6–9. [Google Scholar] [CrossRef] [PubMed]
- Koutroumanidou, E.; Kimbaris, A.; Kortsaris, A.; Bezirtzoglou, E.; Polissiou, M.; Charalabopoulos, K.; Pagonopoulou, O. Increased seizure latency and decreased severity of pentylenetetrazol-induced seizures in mice after essential oil administration. Epilepsy Res. Treat. 2013, 2013, 532657. [Google Scholar] [CrossRef] [PubMed]
- Van Erum, J.; Van Dam, D.; De Deyn, P.P. PTZ-induced seizures in mice require a revised Racine scale. Epilepsy Behav. EB 2019, 95, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Dela Pena, I.J.I.; Kim, H.J.; de la Pena, J.B.; Kim, M.; Botanas, C.J.; You, K.Y.; Woo, T.; Lee, Y.S.; Jung, J.C.; Kim, K.M.; et al. A tryptic hydrolysate from bovine milk alphas1-casein enhances pentobarbital-induced sleep in mice via the GABAA receptor. Behav. Brain Res. 2016, 313, 184–190. [Google Scholar] [CrossRef]
- Matsumoto, K.; Ojima, K.; Watanabe, H. Neurosteroidal modulation of social isolation-induced decrease in pentobarbital sleep in mice. Brain Res. 1996, 708, 1–6. [Google Scholar] [CrossRef]
- Ojima, K.; Matsumoto, K.; Tohda, M.; Watanabe, H. Hyperactivity of central noradrenergic and CRF systems is involved in social isolation-induced decrease in pentobarbital sleep. Brain Res. 1995, 684, 87–94. [Google Scholar] [CrossRef]
- Ghorbani, A.; Rakhshandeh, H.; Sadeghnia, H.R. Potentiating Effects of Lactuca sativa on Pentobarbital-Induced Sleep. Iran J. Pharm. Res. 2013, 12, 401–406. [Google Scholar]
- Egashira, N.; Nogami, A.; Iwasaki, K.; Ishibashi, A.; Uchida, N.; Takasaki, K.; Mishima, K.; Nishimura, R.; Oishi, R.; Fujiwara, M. Yokukansan Enhances Pentobarbital-Induced Sleep in Socially Isolated Mice: Possible Involvement of GABA(A)—Benzodiazepine Receptor Complex. J. Pharmacol. Sci. 2011, 116, 316–320. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Voronin, M.V.; Shangin, S.V.; Litvinova, S.A.; Abramova, E.V.; Kurbanov, R.D.; Rybina, I.V.; Vakhitova, Y.V.; Seredenin, S.B. Pharmacological Analysis of GABAA Receptor and Sigma1R Chaperone Interaction: Research Report I―Investigation of the Anxiolytic, Anticonvulsant and Hypnotic Effects of Allosteric GABAA Receptors’ Ligands. Int. J. Mol. Sci. 2023, 24, 9580. https://doi.org/10.3390/ijms24119580
Voronin MV, Shangin SV, Litvinova SA, Abramova EV, Kurbanov RD, Rybina IV, Vakhitova YV, Seredenin SB. Pharmacological Analysis of GABAA Receptor and Sigma1R Chaperone Interaction: Research Report I―Investigation of the Anxiolytic, Anticonvulsant and Hypnotic Effects of Allosteric GABAA Receptors’ Ligands. International Journal of Molecular Sciences. 2023; 24(11):9580. https://doi.org/10.3390/ijms24119580
Chicago/Turabian StyleVoronin, Mikhail V., Stanislav V. Shangin, Svetlana A. Litvinova, Elena V. Abramova, Rustam D. Kurbanov, Inna V. Rybina, Yulia V. Vakhitova, and Sergei B. Seredenin. 2023. "Pharmacological Analysis of GABAA Receptor and Sigma1R Chaperone Interaction: Research Report I―Investigation of the Anxiolytic, Anticonvulsant and Hypnotic Effects of Allosteric GABAA Receptors’ Ligands" International Journal of Molecular Sciences 24, no. 11: 9580. https://doi.org/10.3390/ijms24119580