
Production of Mare Chorionic Girdle Organoids That Secrete Equine Chorionic Gonadotropin
 , , , ,
, , , ,
Abstract
:1. Introduction
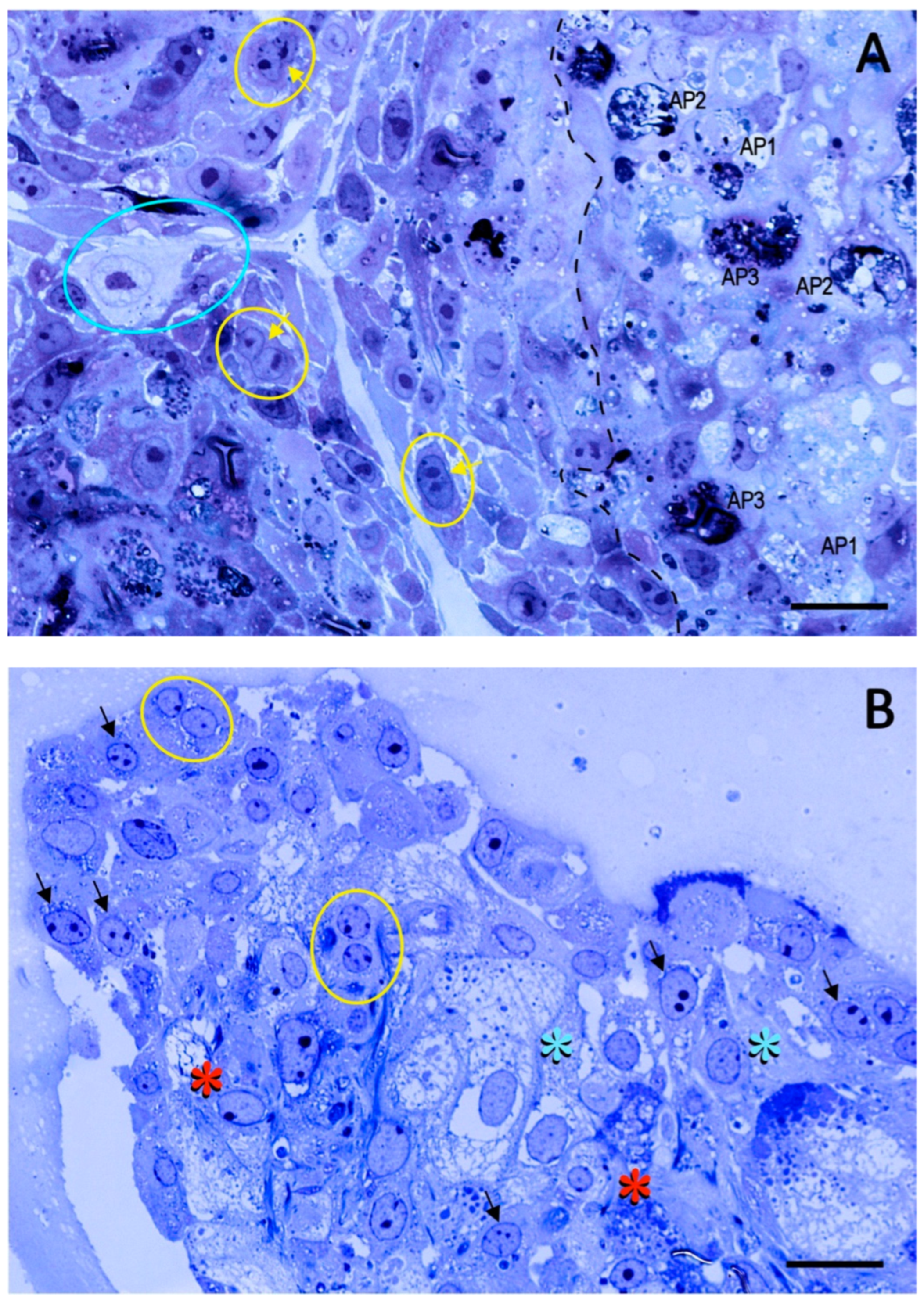
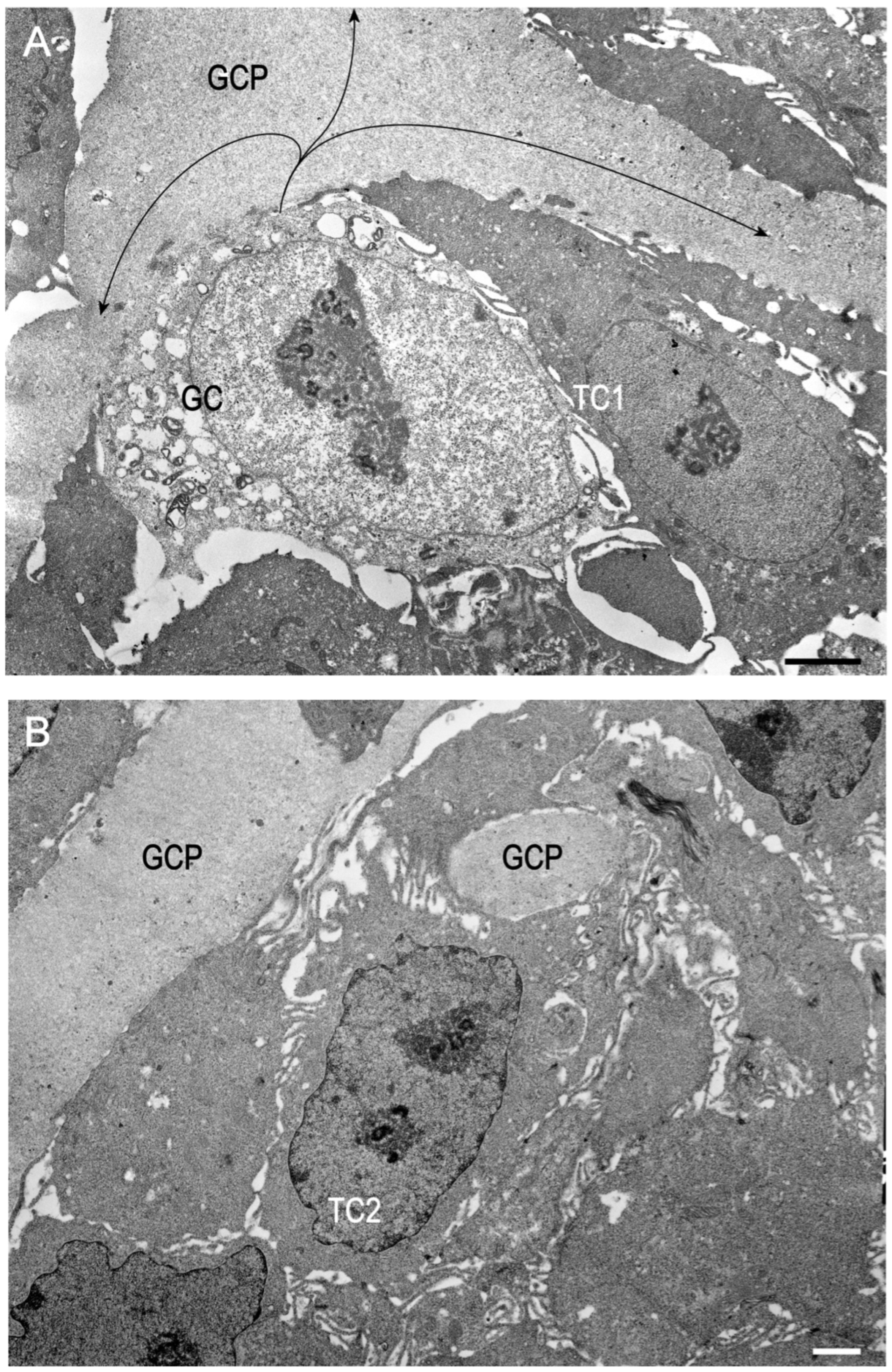
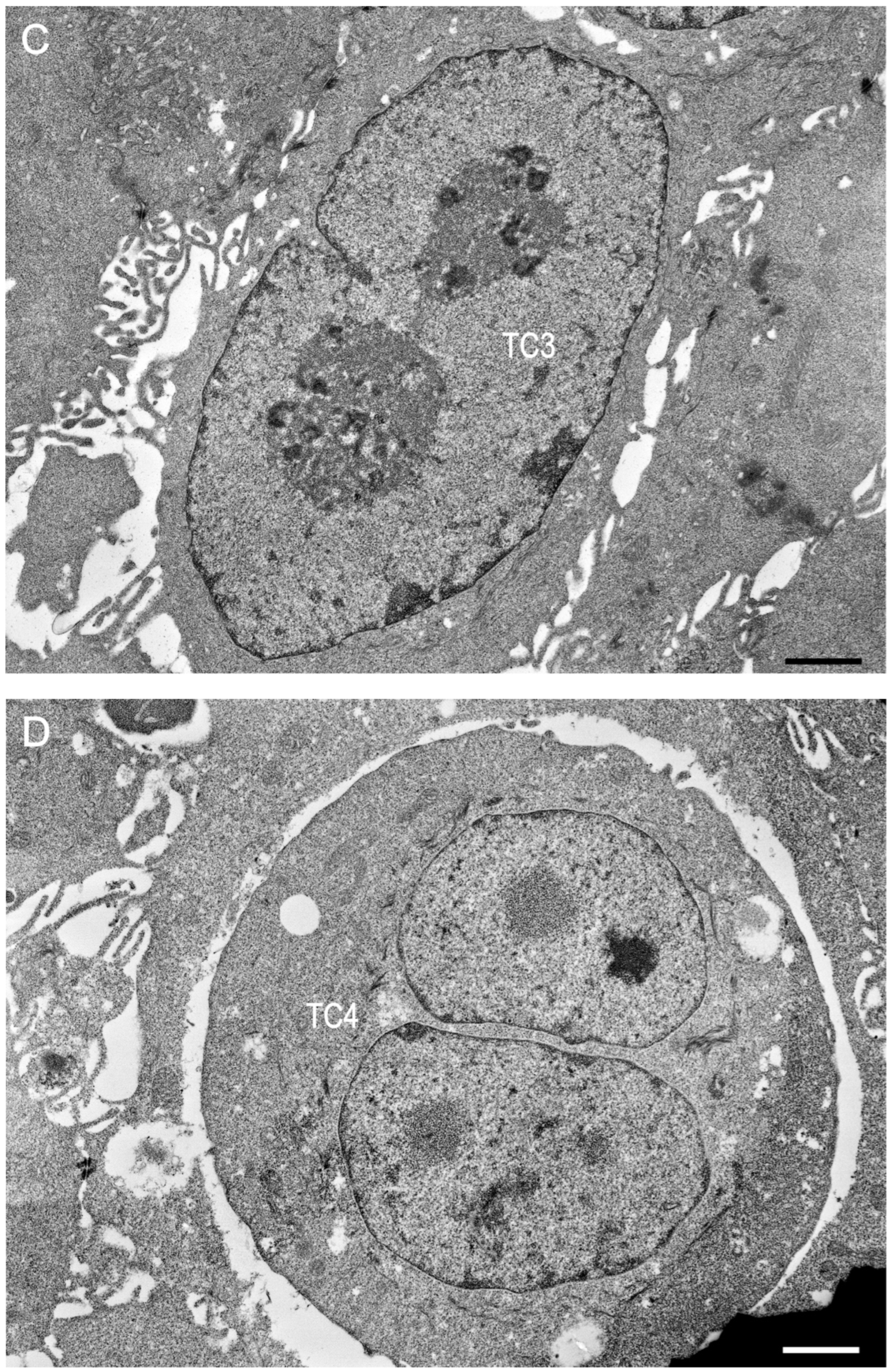
2. Results
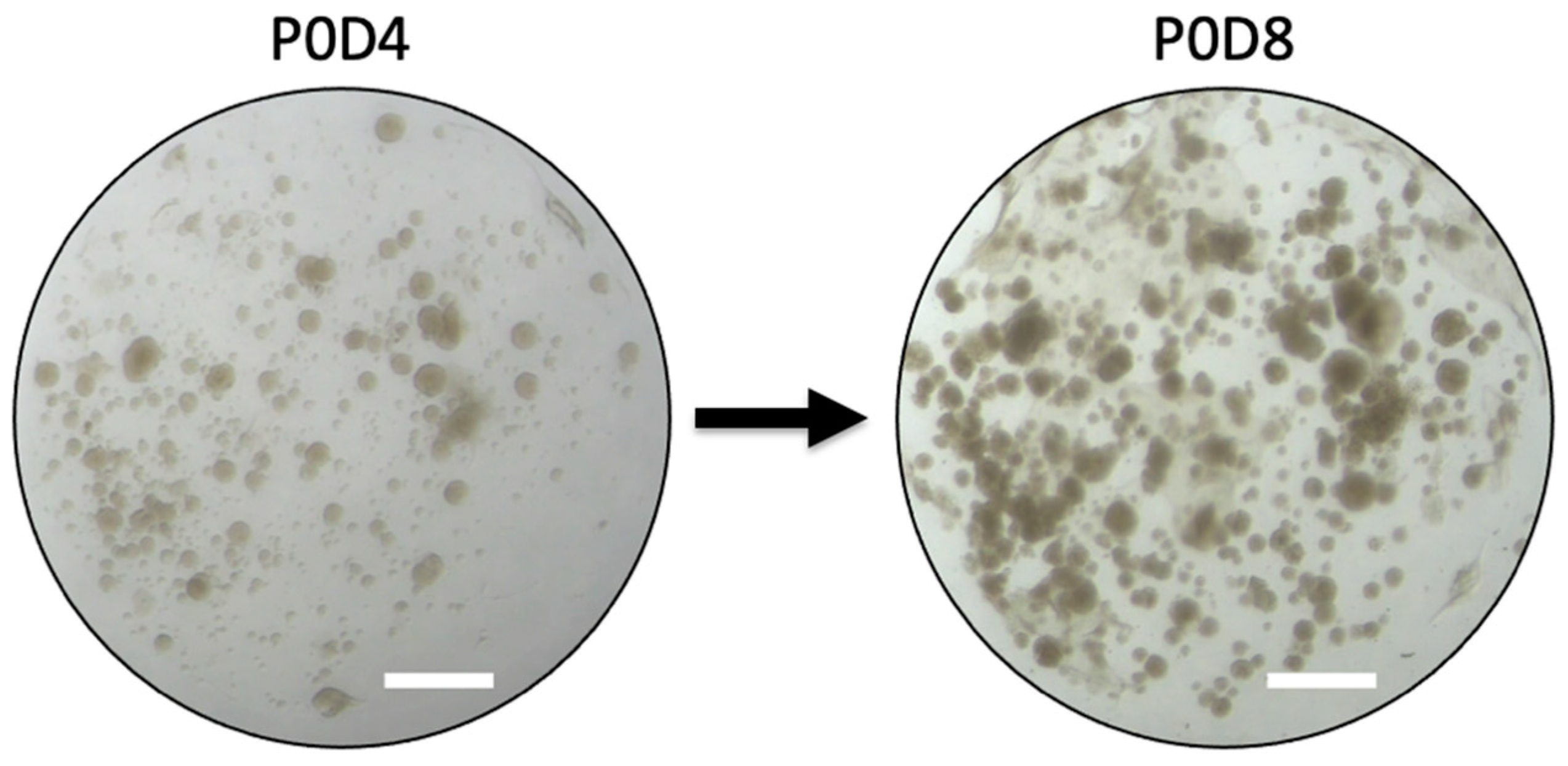
2.1. Preliminary Data
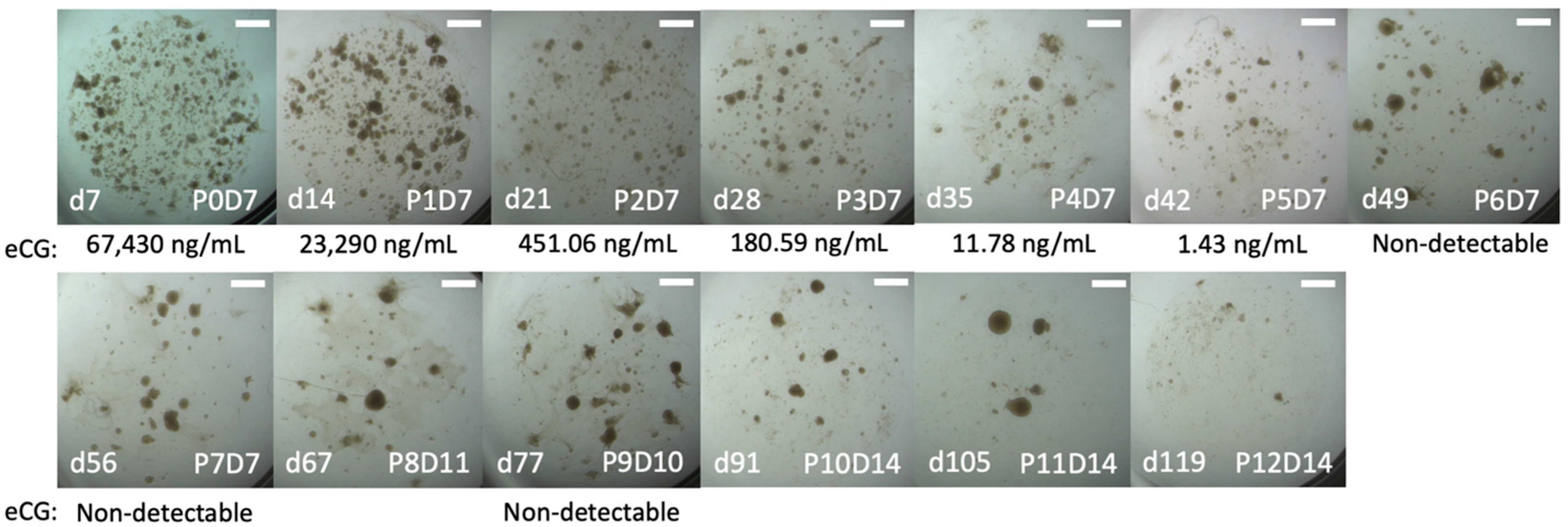
2.2. Trial 1
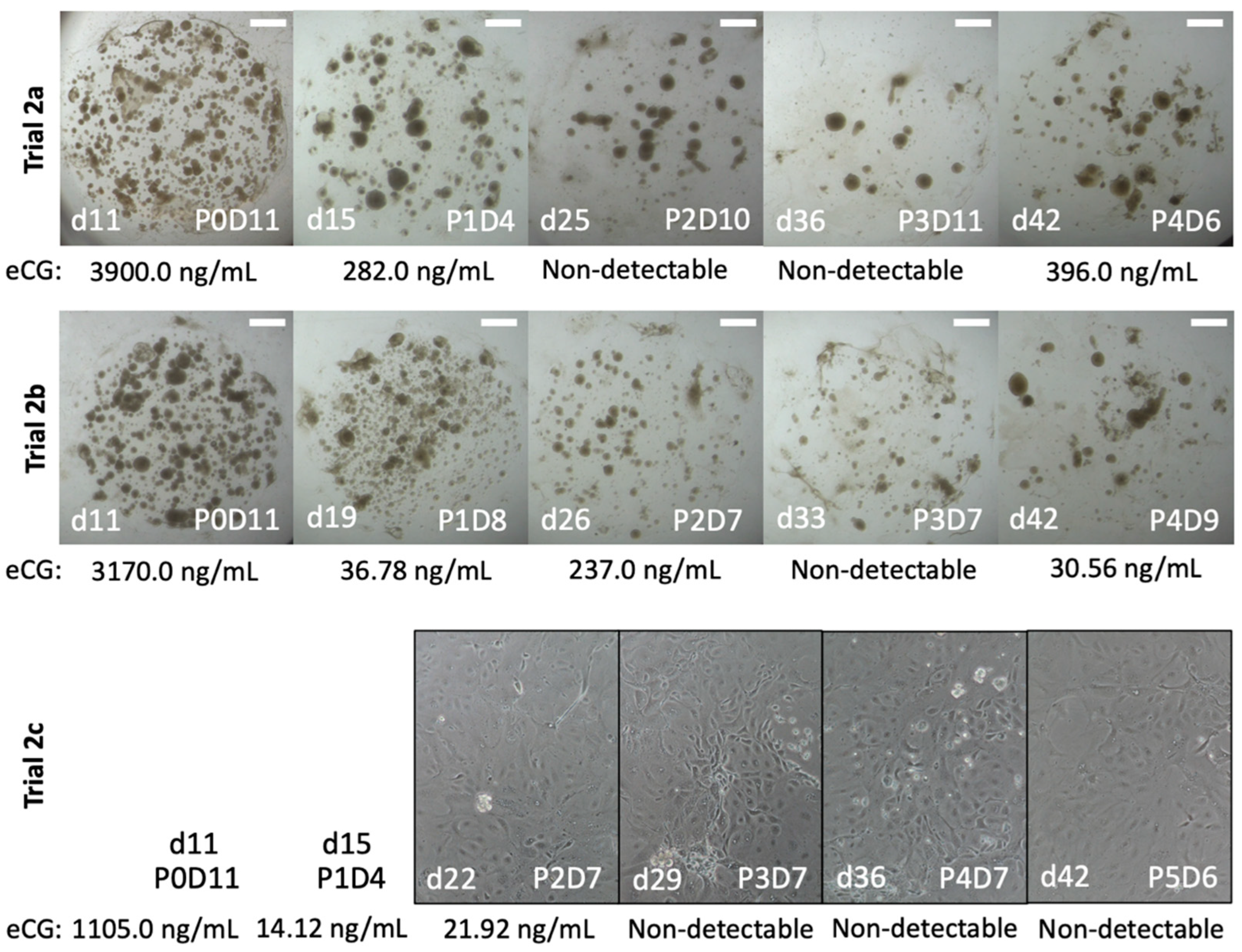
2.3. Trial 2
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Experimental Design
4.3. Organoid Culture
4.4. Monolayer Cell Culture
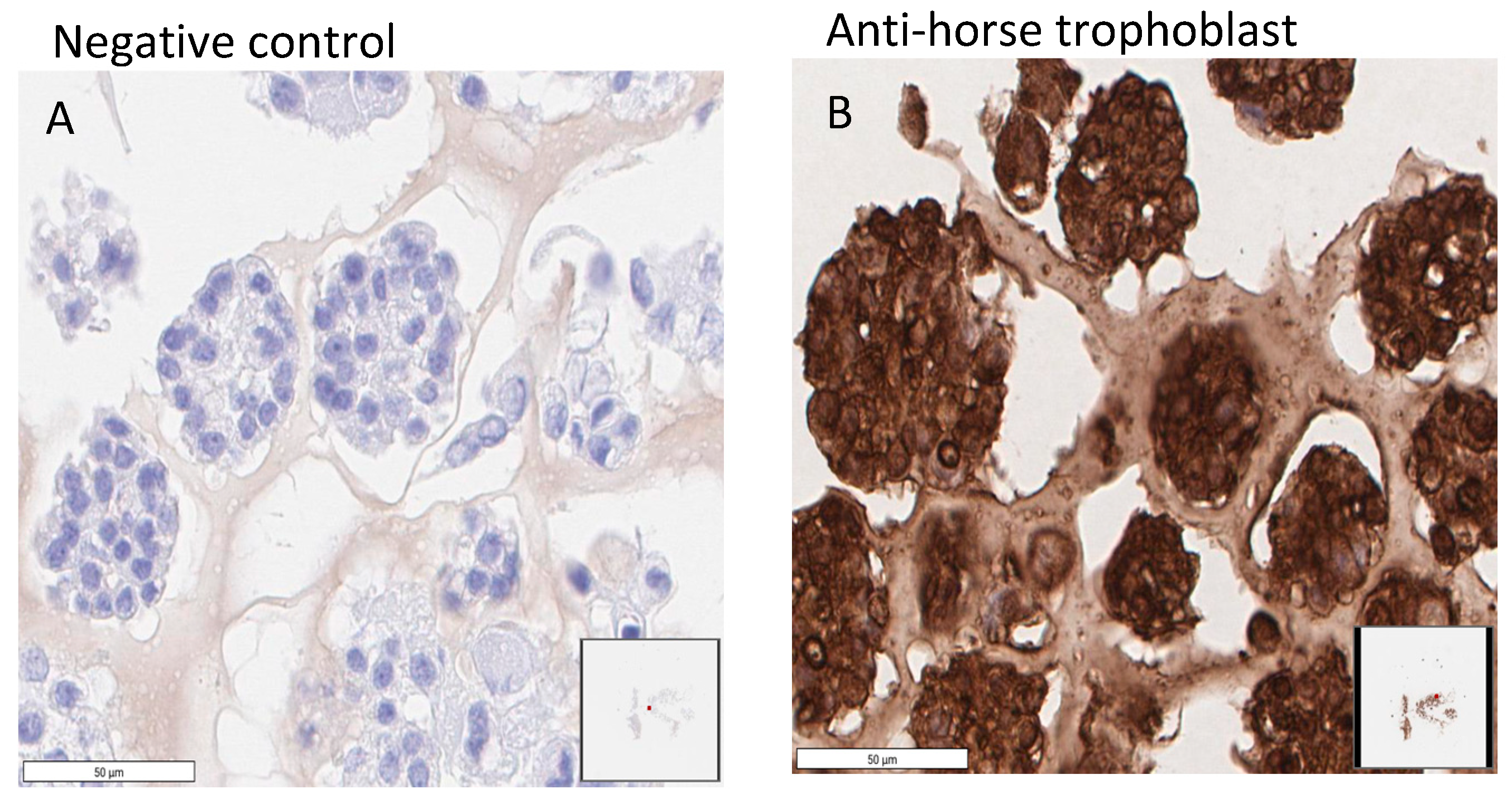
4.5. Immunohistochemistry (IHC)
4.6. Transmission Electron Microscopy (TEM)
4.7. Radioimmunoassay (RIA)
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Allen, W.R.; Moor, R.M. The origin of the equine endometrial cups. Reproduction 1972, 29, 313–316. [Google Scholar] [CrossRef] [PubMed]
- Wooding, F.B.P.; Morgan, G.; Fowden, A.L.; Allen, W.R. A Structural and Immunological Study of Chorionic Gonadotrophin Production by Equine Trophoblast Girdle and Cup Cells. Placenta 2001, 22, 749–767. [Google Scholar] [CrossRef] [PubMed]
- Allen, W.R. The immunological measurement of pregnant mare serum gonadotrophin. J. Endocrinol. 1969, 43, 593–598. [Google Scholar] [CrossRef]
- Allen, W.R.; Hamilton, D.W.; Moor, R.M. The origin of equine endometrial cups. II. Invasion of the endometrium by trophoblast. Anat. Rec. 1973, 177, 485–501. [Google Scholar] [CrossRef] [PubMed]
- Wilsher, S.; Allen, W.R. Factors influencing equine chorionic gonadotrophin production in the mare. Equine Vet. J. 2011, 43, 430–438. [Google Scholar] [CrossRef]
- Allen, W.R.; Stewart, F. Equine placentation. Reprod. Fertil. Dev. 2001, 13, 623–634. [Google Scholar] [CrossRef]
- Daels, P.F.; Albrecht, B.A.; Mohammed, H.O. Equine Chorionic Gonadotropin Regulates Luteal Steroidogenesis in Pregnant Mares. Biol. Reprod. 1998, 59, 1062–1068. [Google Scholar] [CrossRef]
- Liu, X.; Dai, Q.; Hart, E.J.; Barrett, D.M.W.; Rawlings, N.C.; Pierson, R.A.; Bartlewski, P.M. Ultrasonographic Characteristics of Ovulatory Follicles and Associated Endocrine Changes in Cyclic Ewes Treated with Medroxyprogesterone Acetate (MAP)-releasing Intravaginal Sponges and Equine Chorionic Gonadotropin (eCG). Reprod. Domest. Anim. 2007, 42, 393–401. [Google Scholar] [CrossRef]
- Lucia, T.; Corrêa, M.N.; Deschamps, J.C.; Peruzzo, I.A.; Matheus, J.E.M.; Aleixo, J.A.G. Influence of equine chorionic gonadotropin on weaning-to-estrus interval and estrus duration in early-weaned, primiparous, female swine. J. Anim. Sci. 1999, 77, 3163–3167. [Google Scholar] [CrossRef]
- De Rensis, F.; López-Gatius, F. Use of equine chorionic gonadotropin to control reproduction of the dairy cow: A review. Reprod. Domest. Anim. 2014, 49, 177–182. [Google Scholar] [CrossRef]
- Shahneh, Z.; Sadeghipanah, A.; Barfourooshi, H.J.; Emami-Mibody, M.A. Effects of equine chorionic gonadotropin (eCG) administration and flushing on reproductive performance in Nadooshan goats of Iran. Afr. J. Biotechnol. 2008, 7, 3373–3379. [Google Scholar]
- Pierce, J.G.; Parsons, T.F. Glycoprotein Hormones: Structure and Function. Annu. Rev. Biochem. 1981, 50, 465–495. [Google Scholar] [CrossRef] [PubMed]
- Sugino, H.; Bousfield, G.R.; Moore, W.T.; Ward, D.N. Structural studies on equine glycoprotein hormones. Amino acid sequence of equine chorionic gonadotropin beta-subunit. J. Biol. Chem. 1987, 262, 8603–8609. [Google Scholar] [CrossRef] [PubMed]
- Sherman, G.B. A single gene encodes the beta-subunits of equine luteinizing hormone and chorionic gonadotropin. Mol. Endocrinol. 1992, 6, 951–959. [Google Scholar] [CrossRef] [PubMed]
- Murphy, B.D. Equine chorionic gonadotrophin: An enigmatic but essential tool. Anim. Reprod. 2012, 9, 223–230. [Google Scholar]
- Martinuk, S.D.; Manning, A.W.; Black, W.D.; Murphy, B.D. Effects of Carbohydrates on the Pharmacokinetics and Biological Activity of Equine Chorionic Gonadotropin in Vivo1. Biol. Reprod. 1991, 45, 598–604. [Google Scholar] [CrossRef]
- Vilanova, X.M.; De Briyne, N.; Beaver, B.; Turner, P.V. Horse Welfare During Equine Chorionic Gonadotropin (eCG) Production. Animals 2019, 9, 1053. [Google Scholar] [CrossRef]
- Cabrera-Sharp, V.; Read, J.E.; Richardson, S.; Kowalski, A.A.; Antczak, D.F.; Cartwright, J.E.; Mukherjee, A.; De Mestre, A.M. SMAD1/5 Signaling in the Early Equine Placenta Regulates Trophoblast Differentiation and Chorionic Gonadotropin Secretion. Endocrinology 2014, 155, 3054–3064. [Google Scholar] [CrossRef]
- Salman, S.; Asghar, A.; Magee, C.; Winger, Q.; Bouma, G.; Bruemmer, J. 90 Establishment and characterization of Day 30 equine chorionic girdle and allantochorion cell lines. Reprod. Fertil. Dev. 2020, 32, 171. [Google Scholar] [CrossRef]
- Thway, T.M.; Clay, C.M.; Maher, J.K.; Reed, D.K.; McDowell, K.J.; Antczak, D.F.; Eckert, R.L.; Nilson, J.H.; Wolfe, M.W. Immortalization of equine trophoblast cell lines of chorionic girdle cell lineage by simian virus-40 large T antigen. J. Endocrinol. 2001, 171, 45–55. [Google Scholar] [CrossRef]
- Gu, Z.-Y.; Jia, S.-Z.; Liu, S.; Leng, J.-H. Endometrial Organoids: A New Model for the Research of Endometrial-Related Diseases. Biol. Reprod. 2020, 103, 918–926. [Google Scholar] [CrossRef]
- Lancaster, M.A.; Knoblich, J.A. Organogenesis in a dish: Modeling development and disease using organoid technologies. Science 2014, 345, 1247125. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H. Modeling Development and Disease with Organoids. Cell 2016, 165, 1586–1597. [Google Scholar] [CrossRef]
- Song, Y.; Fazleabas, A.T. Endometrial Organoids: A Rising Star for Research on Endometrial Development and Associated Diseases. Reprod. Sci. 2021, 28, 1626–1636. [Google Scholar] [CrossRef] [PubMed]
- Huch, M.; Dorrell, C.; Boj, S.F.; Van Es, J.H.; Li, V.S.W.; Van De Wetering, M.; Sato, T.; Hamer, K.; Sasaki, N.; Finegold, M.J.; et al. In vitro expansion of single Lgr5+ liver stem cells induced by Wnt-driven regeneration. Nature 2013, 494, 247–250. [Google Scholar] [CrossRef] [PubMed]
- Karthaus, W.R.; Iaquinta, P.J.; Drost, J.; Gracanin, A.; van Boxtel, R.; Wongvipat, J.; Dowling, C.M.; Gao, D.; Begthel, H.; Sachs, N.; et al. Identification of Multipotent Luminal Progenitor Cells in Human Prostate Organoid Cultures. Cell 2014, 159, 163–175. [Google Scholar] [CrossRef]
- Lancaster, M.A.; Renner, M.; Martin, C.-A.; Wenzel, D.; Bicknell, L.S.; Hurles, M.E.; Homfray, T.; Penninger, J.M.; Jackson, A.P.; Knoblich, J.A. Cerebral organoids model human brain development and microcephaly. Nature 2013, 501, 373–379. [Google Scholar] [CrossRef]
- Boretto, M.; Cox, B.; Noben, M.; Hendriks, N.; Fassbender, A.; Roose, H.; Amant, F.; Timmerman, D.; Tomassetti, C.; Vanhie, A.; et al. Development of organoids from mouse and human endometrium showing endometrial epithelium physiology and long-term expandability. Development 2017, 144, 1775–1786. [Google Scholar] [CrossRef]
- Turco, M.Y.; Gardner, L.; Hughes, J.; Cindrova-Davies, T.; Gomez, M.J.; Farrell, L.; Hollinshead, M.; Marsh, S.G.E.; Brosens, J.J.; Critchley, H.O.; et al. Long-term, hormone-responsive organoid cultures of human endometrium in a chemically defined medium. Nat. Cell Biol. 2017, 19, 568–577. [Google Scholar] [CrossRef]
- Turco, M.Y.; Gardner, L.; Kay, R.G.; Hamilton, R.S.; Prater, M.; Hollinshead, M.S.; McWhinnie, A.; Esposito, L.; Fernando, R.; Skelton, H.; et al. Trophoblast organoids as a model for maternal–fetal interactions during human placentation. Nature 2018, 564, 263–267. [Google Scholar] [CrossRef]
- Thompson, R.E.; Johnson, A.K.; Dini, P.; Turco, M.Y.; Prado, T.M.; Premanandan, C.; Burton, G.J.; Ball, B.A.; Whitlock, B.K.; Pukazhenthi, B.S. Hormone-responsive organoids from domestic mare and endangered Przewalski’s horse endometrium. Reproduction 2020, 160, 819–831. [Google Scholar] [CrossRef] [PubMed]
- Thompson, R.E.; Meyers, M.A.; Pukazhenthi, B.S.; Hollinshead, F.K. Evaluation of growth, viability, and structural integrity of equine endometrial organoids following cryopreservation. Cryobiology 2021, 104, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Thompson, R.E.; Meyers, M.A.; Veeramachaneni, D.N.R.; Pukazhenthi, B.S.; Hollinshead, F.K. Equine Oviductal Organoid Generation and Cryopreservation. Methods Protoc. 2022, 5, 51. [Google Scholar] [CrossRef] [PubMed]
- de Mestre, A.M.; Bacon, S.J.; Costa, C.C.; Leadbeater, J.C.; Noronha, L.E.; Stewart, F.; Antczak, D.F. Modeling Trophoblast Differentiation using Equine Chorionic Girdle Vesicles. Placenta 2008, 29, 158–169. [Google Scholar] [CrossRef]
- Legardinier, S.; Klett, D.; Poirier, J.-C.; Combarnous, Y.; Cahoreau, C. Mammalian-like nonsialyl complex-type N-glycosylation of equine gonadotropins in Mimic™ insect cells. Glycobiology 2005, 15, 776–790. [Google Scholar] [CrossRef] [PubMed]
- Villarraza, C.J.; Antuña, S.; Tardivo, M.B.; Rodríguez, M.C.; Mussio, P.; Cattaneo, L.; Fontana, D.; Díaz, P.U.; Ortega, H.H.; Tríbulo, A.; et al. Development of a suitable manufacturing process for production of a bioactive recombinant equine chorionic gonadotropin (reCG) in CHO-K1 cells. Theriogenology 2021, 172, 8–19. [Google Scholar] [CrossRef]
- Adams, A.P.; Antczak, D.F. Ectopic Transplantation of Equine Invasive Trophoblast1. Biol. Reprod. 2001, 64, 753–763. [Google Scholar] [CrossRef]
- Antczak, D.F.; Oriol, J.G.; Donaldson, W.L.; Poleman, C.; Stenzler, L.; Volsen, S.G.; Allen, W.R. Differentiation molecules of the equine trophoblast. J. Reprod. Fertil. Suppl. 1987, 35, 371–378. [Google Scholar]
- Thompson, R.E.; Johnson, A.K.; Prado, T.M.; Premanandan, C.; Brown, M.E.; Whitlock, B.K.; Pukazhenthi, B.S. Dimethyl sulfoxide maintains structure and function of cryopreserved equine endometrial explants. Cryobiology 2019, 91, 90–96. [Google Scholar] [CrossRef]
- Thompson, R.E.; Brown, M.E.; Helmick, K.; Whitlock, B.K.; Pukazhenthi, B.S. Persian onager (Equus hemionus onager) endometrial explant cryopreservation and in vitro culture. Anim. Reprod. Sci. 2020, 217, 106459. [Google Scholar] [CrossRef]
- Oriol, J.G.; Poleman, J.C.; Antczak, D.F.; Allen, W.R. A monoclonal antibody specific for equine trophoblast. Equine Vet. J. 1989, 21, 14–18. [Google Scholar] [CrossRef]
- Nett, T.; Holtan, D.; Estergreen, V. Levels of LH, prolactin and estrogens in the serum of postpartum mares. J. Reprod. Fertil. 1975, 23, 201–206. [Google Scholar]
- Nett, T.M.; Holtan, D.W.; Estergreen, V.L. Oestrogens, LH, PMSG, and prolactin in serum of pregnant mares. J. Reprod. Fertil. Suppl. 1975, 23, 457–462. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trial | Cell Isolation | Culture Method | Culture Medium | Incubator Settings |
|---|---|---|---|---|
| 1 | Enzymatic digestion | Organoid | Human trophoblast organoid medium [30] | 37 °C, 5% CO2 |
| 2a | Enzymatic digestion | Organoid | Human trophoblast organoid medium [30] | 37 °C, 5% CO2 |
| 2b | Mechanical isolation | Organoid | Human trophoblast organoid medium [30] | 37 °C, 8% CO2 |
| 2c | Mechanical isolation | Monolayer | Equine chorionic girdle monolayer medium [34] | 37 °C, 8% CO2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thompson, R.E.; Meyers, M.A.; Palmer, J.; Veeramachaneni, D.N.R.; Magee, C.; de Mestre, A.M.; Antczak, D.F.; Hollinshead, F.K. Production of Mare Chorionic Girdle Organoids That Secrete Equine Chorionic Gonadotropin. Int. J. Mol. Sci. 2023, 24, 9538. https://doi.org/10.3390/ijms24119538
Thompson RE, Meyers MA, Palmer J, Veeramachaneni DNR, Magee C, de Mestre AM, Antczak DF, Hollinshead FK. Production of Mare Chorionic Girdle Organoids That Secrete Equine Chorionic Gonadotropin. International Journal of Molecular Sciences. 2023; 24(11):9538. https://doi.org/10.3390/ijms24119538
Chicago/Turabian StyleThompson, Riley E., Mindy A. Meyers, Jennifer Palmer, D. N. Rao Veeramachaneni, Christianne Magee, Amanda M. de Mestre, Douglas F. Antczak, and Fiona K. Hollinshead. 2023. "Production of Mare Chorionic Girdle Organoids That Secrete Equine Chorionic Gonadotropin" International Journal of Molecular Sciences 24, no. 11: 9538. https://doi.org/10.3390/ijms24119538