


Involvement of Nitric Oxide and Melatonin Enhances Cadmium Resistance of Tomato Seedlings through Regulation of the Ascorbate–Glutathione Cycle and ROS Metabolism

Abstract
:1. Introduction
2. Results
2.1. Effect of Different Concentrations of Cd on the Growth of Tomato Seedlings
2.2. Effect of Different Concentrations of MT and SNP on the Growth of Tomato Seedlings under Cd Stress
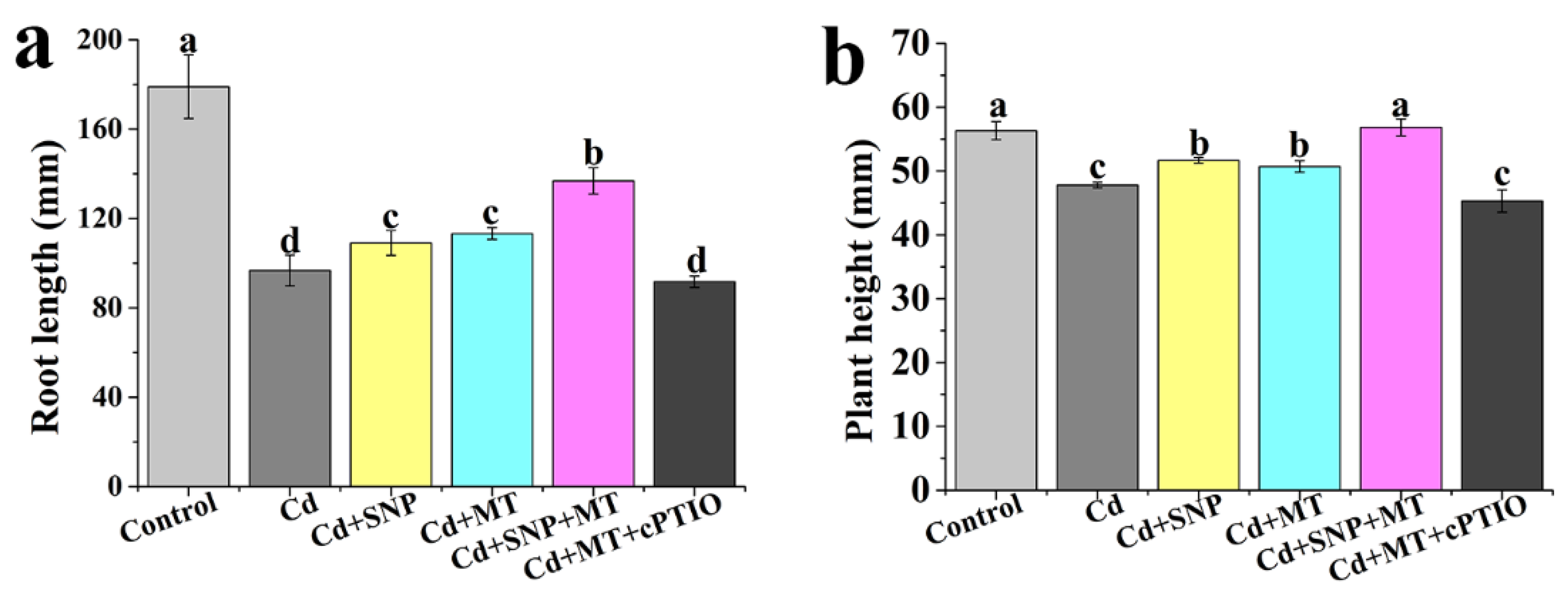
2.3. Effect of Different Treatments on the Growth of Tomato Seedlings under Cd Stress
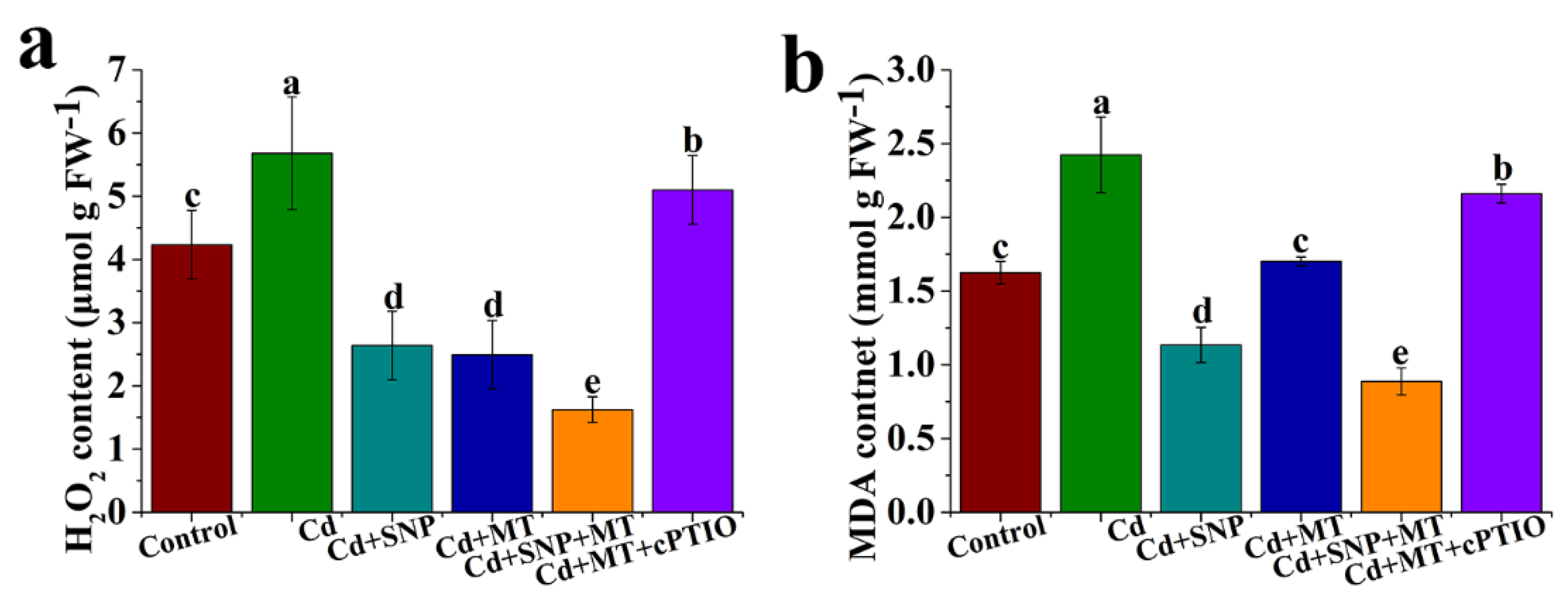
2.4. Different Treatments Alter the Levels of H2O2 and MDA under Cd Stress
2.5. Effect of Different Treatments on the Levels of AsA, DHA, GSH, and GSSG under Cd Stress
2.6. Effect of Different Treatments on the Levels of AsA/DHA and GSH/GSSG Ratios under Cd Stress
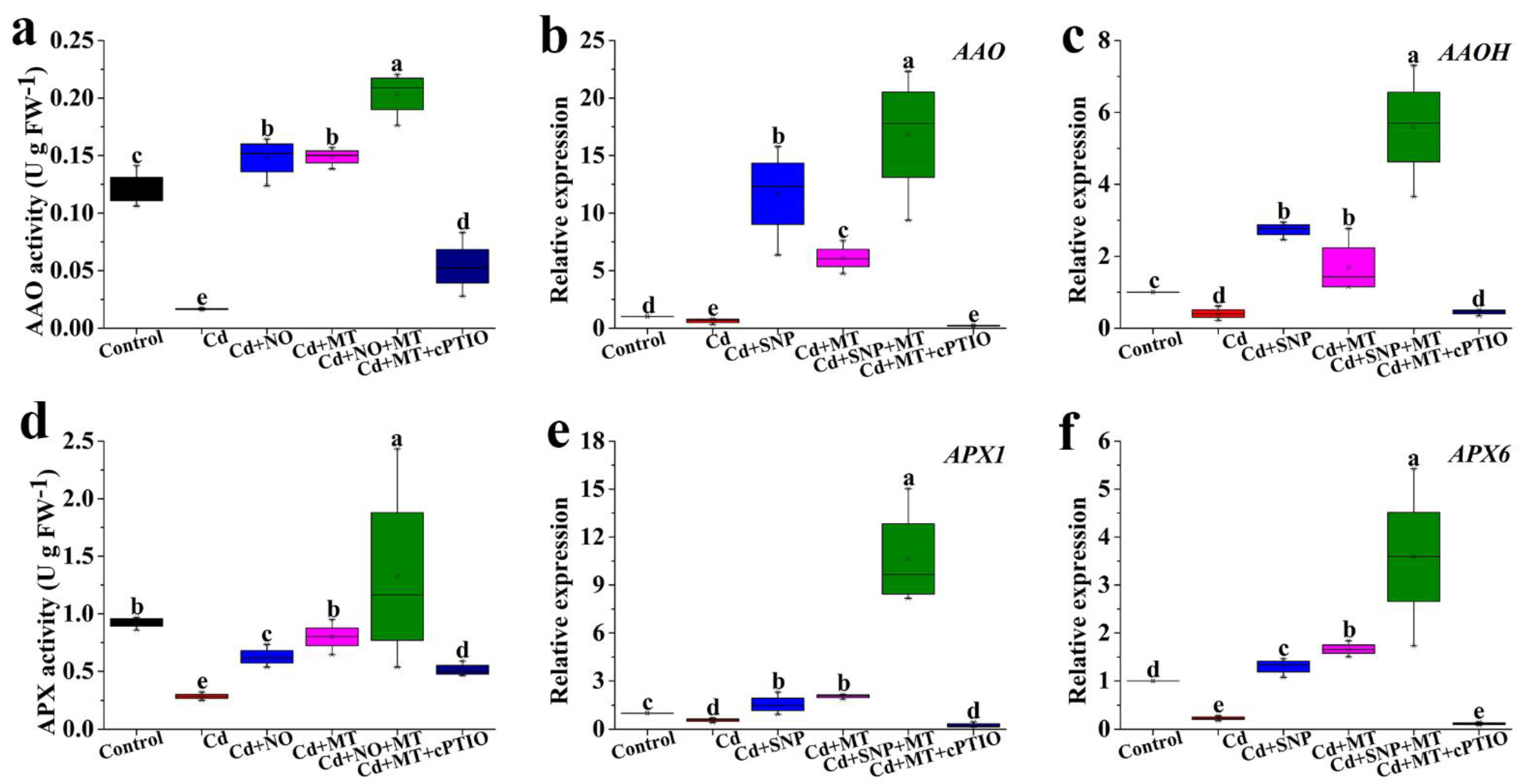
2.7. Effect of Different Treatments on the AAO and APX Activities and Gene Expression under Cd Stress
2.8. Effect of Different Treatments on the Activities of MDHAR, DHAR, and GR and Gene Expression under Cd Stress
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. H2O2 Content
4.3. Malonaldehyde (MDA) Content
4.4. Ascorbic Acid (AsA) and Dehydroascorbic Acid (DHA) Content
4.5. Reduced Glutathione (GSH) and Oxidized Glutathione (GSSG) Content
4.6. Enzymatic Activities
4.7. RNA Isolation and Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lu, C.W.; Ho, H.C.; Yao, C.L.; Tseng, T.Y.; Kao, C.M.; Chen, S.C. Bioremediation potential of cadmium by recombinant Escherichia coli surface expressing metallothionein MTT5 from Tetrahymena thermophila. Chemosphere 2023, 310, 136850. [Google Scholar] [CrossRef] [PubMed]
- Mojtahedi, N.; Zare-Dorabei, R.; Mosavi, S.H. A Zn-based metal-organic framework modified by CuCl2 under ambient conditions for simultaneous ultrasonic-assisted removal of Pb and Cd ions with fast kinetics from aqueous solution. Chemistryselect 2023, 8, e202204948. [Google Scholar] [CrossRef]
- Farhan, A.; Zulfiqar Samiah, M.; Rashid, E.U.; Nawaz, S.; Iqbal, H.M.N.; Jesionowski, T.; Bilal, M.; Zdarta, J. Removal of toxic metals from water by nanocomposites through advanced remediation processes and photocatalytic oxidation. Curr. Pollution Rep. 2023, 1–20. [Google Scholar] [CrossRef]
- Hu, Y.N.; Cheng, H.F.; Tao, S. The challenges and solutions for cadmium-contaminated rice in China: A critical review. Environ. Int. 2016, 92–93, 515–532. [Google Scholar] [CrossRef] [PubMed]
- Palansooriya, K.N.; Shaheen, S.M.; Chen, S.S.; Tsang, D.C.W.; Hashimoto, Y.; Hou, D.Y.; Bolan, N.S.; Rinklebe, J.; Ok, Y.S. Soil amendments for immobilization of potentially toxic elements in contaminated soils: A critical review. Environ. Int. 2020, 134, 105046. [Google Scholar] [CrossRef] [PubMed]
- Dado, T.B.; Kastro, C.; Fikire, Y.; Gudishe, K.; Bojago, E. ICP_OES determination of essential, trace essential, and non-essential elements in the anatomical parts of medicinal herb (Pentas Schimperiana) grown in Wolaita Zone, Ethiopia. J. Agr. Food Res. 2023, 12, 100547. [Google Scholar] [CrossRef]
- Lux, A.; Martinka, M.; Vaculík, M.; White, P.J. Root responses to cadmium in the rhizosphere: A review. J. Exp. Bot. 2011, 62, 21–37. [Google Scholar] [CrossRef]
- Ibaraki, T.; Fujitomi, S.I.; Ishitsuka, A.; Yanaka, M. Phytoextraction by high-Cd-accumulating rice to reduce Cd in wheat grains grown in Cd-polluted fields. Soil Sci. Plant Nutr. 2014, 60, 266–275. [Google Scholar] [CrossRef]
- Goncharuk, E.A.; Zagoskina, N.V. Heavy metals, their phytotoxicity, and the role of phenolic antioxidants in plant stress responses with focus on cadmium: Review. Molecules 2023, 28, 3921. [Google Scholar] [CrossRef]
- Liu, H.T.; Jiao, Q.J.; Fan, L.; Jiang, Y.; Alyemeni, M.N.; Ahmad, P.; Chen, Y.L.; Zhu, M.; Liu, H.P.; Zhao, Y.; et al. Integrated physio-biochemical and transcriptomic analysis revealed mechanism underlying of Si-mediated alleviation to cadmium toxicity in wheat. J. Hazard. Mater. 2023, 452, 131366. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Li, F.; Ji, C.L.; Wu, H.F. Toxicological mechanism of cadmium in the clam Ruditapes philippinarum using combined ionomic, metabolomic and transcriptomic analyses. Environ. Pollut. 2023, 323, 121286. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, M.E.A.; Agathokleous, E.; Nogueira, M.L.; Brunetto, G.; Brown, P.H.; Azevedo, R.A. Neutral-to-positive cadmium effects on germination and seedling vigor, with and without seed priming. J. Hazard. Mater. 2023, 448, 130813. [Google Scholar] [CrossRef] [PubMed]
- Ou, C.; Cheng, W.H.; Wang, Z.L.; Yao, X.M.; Yang, S.G. Exogenous melatonin enhances Cd stress tolerance in Platycladus orientalis seedlings by improving mineral nutrient uptake and oxidative stress. Ecotox. Environ. Safe 2023, 252, 114619. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.Y.; Hu, J.H.; Chen, L.; Wang, Z.; Sun, S.Y.; Zhang, W.M.; Jiang, H.; Luo, Y.; Wang, L.; Zeng, Y.; et al. Microplastics may increase the environmental risks of Cd via promoting Cd uptake by plants: A meta-analysis. J. Hazard. Mater. 2023, 448, 130887. [Google Scholar] [CrossRef]
- Peng, H.; Deng, K.; Shi, Y.; Liu, S.H.; Jian, Z.; Li, C.J.; Ji, X.H.; Li, S.N. Alleviation of Cd-polluted paddy soils through Si fertilizer application and its effects on the soil microbial community. Sci. Total Environ. 2023, 855, 158735. [Google Scholar] [CrossRef]
- Rasafi, T.E.; Oukarroum, A.; Haddioui, A.; Song, H.C.; Kwon, E.E.; Bolan, N.; Tack, F.M.G.; Sebastian, A.; Prasad, M.N.V.; Rinklebe, J. Cadmium stress in plants: A critical review of the effects, mechanisms, and tolerance strategies. Crit. Rev. Env. Sci. Tec. 2020, 52, 675–726. [Google Scholar] [CrossRef]
- Tripathy, S.; Mohanty, P.K. Reactive oxygen species (ROS) are boon or bane. Int. J. Pharm. Sci. Res. 2017, 8, 1. [Google Scholar]
- He, Z.L.; Xu, Q.; Newland, B.; Foley, R.; Lara-Sáez, I.; Curtin, J.F.; Wang, W.X. Reactive oxygen species (ROS): Utilizing injectable antioxidative hydrogels and ROS-producing therapies to manage the double-edged sword. J. Mater. Chem. B 2021, 9, 6326–6346. [Google Scholar] [CrossRef]
- Cuypers, A.; Plusquin, M.; Remans, T.; Jozefczak, M.; Keunen, E.; Gielen, H.; Opdenakker, K.; Nair, A.R.; Munters, E.; Artois, T.J.; et al. Cadmium stress: An oxidative challenge. Biometals 2010, 23, 927–940. [Google Scholar] [CrossRef]
- Kapoor, D.; Singh, S.; Kumar, V.; Romero, R.; Prasad, R.; Singh, J. Antioxidant enzymes regulation in plants in reference to reactive oxygen species (ROS) and reactive nitrogen species (RNS). Plant Gene 2019, 19, 100182. [Google Scholar] [CrossRef]
- Hall, J.L. Cellular mechanisms for heavy metal detoxification and tolerance. J. Exp. Bot. 2002, 53, 1–11. [Google Scholar] [CrossRef]
- Viehweger, K. How plants cope with heavy metal. Bot. Stud. 2014, 55, 35. [Google Scholar] [CrossRef] [PubMed]
- Fancy, N.N.; Bahlmann, A.K.; Loake, G.J. Nitric oxide function in plant abiotic stress. Plant Cell Environ. 2017, 40, 462–472. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.J.; Huo, J.Q.; Zhang, J.; Wang, C.L.; Wang, B.; Fang, H.; Liao, W.B. Protein S-nitrosylation in programmed cell death in plants. Cell Mol. Life Sci. 2019, 76, 1877–1887. [Google Scholar] [CrossRef] [PubMed]
- Qi, F.; Xiang, Z.X.; Kou, N.H.; Cui, W.T.; Xu, D.K.; Wang, R.; Zhu, D.; Shen, W.B. Nitric oxide is involved in methane-induced adventitious root formation in cucumber. Physiol. Plant 2017, 159, 366–377. [Google Scholar] [CrossRef]
- Li, C.X.; Huang, D.J.; Wang, C.L.; Wang, N.; Yao, Y.D.; Li, W.F.; Liao, W.B. NO is involved in H2-induced adventitious rooting in cucumber by regulating the expression and interaction of plasma membrane H+-ATPase and 14-3-3. Planta 2020, 252, 9. [Google Scholar] [CrossRef]
- Cao, Z.Y.; Duan, X.L.; Yao, P.; Cui, W.T.; Cheng, D.; Zhang, J.; Jin, Q.J.; Chen, J.; Dai, T.H.; Shen, W.B. Hydrogen gas is involved in auxin-induced lateral root formation by modulating nitric oxide synthesis. Int. J. Mol. Sci. 2017, 18, 2084. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Li, Q.T.; Chu, Y.N.; Reiter, R.J.; Yu, X.M.; Zhu, D.H.; Zhang, W.K.; Ma, B.; Lin, Q.; Zhang, J.S.; et al. Melatonin enhances plant growth and abiotic stress tolerance in soybean plants. J. Exp. Bot. 2015, 66, 695–707. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin in flowering, fruit set and fruit ripening. Plant Reprod. 2020, 33, 77–87. [Google Scholar] [CrossRef]
- Cui, G.B.; Zhao, X.X.; Liu, S.D.; Sun, F.L.; Zhang, C.; Xi, Y.J. Beneficial effects of melatonin in overcoming drought stress in wheat seedlings. Plant Physiol. Biochem. 2017, 118, 138–149. [Google Scholar] [CrossRef]
- Jiang, C.Q.; Cui, Q.R.; Feng, K.; Xu, D.F.; Li, C.F.; Zheng, Q.S. Melatonin improves antioxidant capacity and ion homeostasis and enhances salt tolerance in maize seedlings. Acta Physiol. Plant 2016, 38, 82. [Google Scholar] [CrossRef]
- Shi, H.T.; Jiang, C.; Ye, T.T.; Tan, D.X.; Reiter, R.J.; Zhang, H.; Liu, R.Y.; Chen, Z. Comparative physiological, metabolomic, and transcriptomic analyses reveal mechanisms of improved abiotic stress resistance in bermudagrass [Cynodon dactylon (L). Pers.] by exogenous melatonin. J. Exp. Bot. 2015, 66, 681–694. [Google Scholar] [CrossRef] [PubMed]
- Aghdam, M.S.; Luo, Z.S.; Jannatizadeh, A.; Sheikh-Assadi, M.; Sharafi, Y.; Farmani, B.; Fard, J.R.; Razavi, F. Employing exogenous melatonin applying confers chilling tolerance in tomato fruits by upregulating ZAT2/6/12 giving rise to promoting endogenous polyamines, proline, and nitric oxide accumulation by triggering arginine pathway activity. Food Chem. 2019, 275, 549–556. [Google Scholar] [CrossRef]
- Lee, H.Y.; Back, K. Melatonin induction and its role in high light stress tolerance in Arabidopsis thaliana. J. Pineal Res. 2018, 65, e12504. [Google Scholar] [CrossRef]
- Gu, Q.; Chen, Z.P.; Yu, X.L.; Cui, W.T.; Pan, J.C.; Zhao, G.; Xu, S.; Wang, R.; Shen, W.B. Melatonin confers plant tolerance against cadmium stress via the decrease of cadmium accumulation and reestablishment of microRNA-mediated redox homeostasis. Plant Sci. 2017, 261, 28–37. [Google Scholar] [CrossRef]
- Zhan, H.S.; Nie, X.J.; Zhang, T.; Li, S.; Wang, X.Y.; Du, X.H.; Tong, W.; Song, W.N. Melatonin: A small molecule but important for salt stress tolerance in plants. Int. J. Mol. Sci. 2019, 20, 709. [Google Scholar] [CrossRef]
- Tan, D.X.; Reiter, R.J.; Manchester, L.C.; Yan, M.T.; El-Sawi, M.; Sainz, R.M.; Mayo, J.C.; Kohen, R.; Allegra, M.; Hardeland, R. Chemical and physical properties and potential mechanisms: Melatonin as a broad spectrum antioxidant and free radical scavenger. Curr. Top. Med. Chem. 2002, 2, 181–197. [Google Scholar] [CrossRef]
- Kaur, H.; Mukherjee, S.; Baluska, F.; Bhatla, S.C. Regulatory roles of serotonin and melatonin in abiotic stress tolerance in plants. Plant Signal. Behav. 2015, 10, e1049788. [Google Scholar] [CrossRef]
- Khan, A.; Numan, M.; Khan, A.L.; Lee, I.J.; Imran, M.; Asaf, S.; Al-Harrasi, A. Melatonin: Awakening the defense mechanisms during plant oxidative stress. Plants 2020, 9, 407. [Google Scholar] [CrossRef]
- Kaur, H.; Bhatla, S.C. Melatonin and nitric oxide modulate glutathione content and glutathione reductase activity in sunflower seedling cotyledons accompanying salt stress. Nitric Oxide 2016, 59, 42–53. [Google Scholar] [CrossRef]
- Liu, N.; Jin, Z.Y.; Wang, S.S.; Gong, B.A.; Wen, D.; Wang, X.F.; Wei, M.; Shi, Q.H. Sodic alkaline stress mitigation with exogenous melatonin involves reactive oxygen metabolism and ion homeostasis in tomato. Sci. Hortic. 2015, 181, 18–25. [Google Scholar] [CrossRef]
- Yu, Y.C.; Wang, A.M.; Li, X.; Kou, M.; Wang, W.J.; Chen, X.Y.; Xu, T.; Zhu, M.K.; Ma, D.F.; Li, Z.Y.; et al. Melatonin-stimulated triacylglycerol breakdown and energy turnover under salinity stress contributes to the maintenance of plasma membrane H+-ATPase activity and K+/Na+ homeostasis in sweet potato. Front. Plant Sci. 2018, 9, 256. [Google Scholar] [CrossRef]
- Ahammed, G.J.; Xu, W.; Liu, A.R.; Chen, S.C. Endogenous melatonin deficiency aggravates high temperature-induced oxidative stress in Solanum lycopersicum L. Environ. Exp. Bot. 2019, 161, 303–311. [Google Scholar] [CrossRef]
- Jahan, M.S.; Shu, S.; Wang, Y.; Chen, Z.; He, M.M.; Tao, M.Q.; Sun, J.; Guo, S.R. Melatonin alleviates heat-induced damage of tomato seedlings by balancing redox homeostasis and modulating polyamine and nitric oxide biosynthesis. BMC Plant Biol. 2019, 19, 414. [Google Scholar] [CrossRef]
- Li, Z.G.; Xu, Y.; Bai, L.K.; Zhang, S.Y.; Wang, Y. Melatonin enhances thermotolerance of maize seedlings (Zea mays L.) by modulating antioxidant defense, methylglyoxal detoxification, and osmoregulation systems. Protoplasma 2019, 256, 471–490. [Google Scholar] [CrossRef]
- Liang, D.; Gao, F.; Ni, Z.Y.; Lin, L.J.; Deng, Q.X.; Tang, Y.; Wang, X.; Luo, X.; Xia, H. Melatonin improves heat tolerance in kiwifruit seedlings through promoting antioxidant enzymatic activity and glutathione S-transferase transcription. Molecules 2018, 23, 584. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.L.; Ye, L.; Wang, Y.P.; Zhou, X.T.; Yang, J.W.; Wang, J.W.; Cao, K.; Zou, Z.R. Melatonin increases the chilling tolerance of chloroplast in cucumber seedlings by regulating photosynthetic electron flux and the ascorbate-glutathione cycle. Front. Plant Sci. 2016, 7, 1314. [Google Scholar] [CrossRef] [PubMed]
- Dawood, M.G.; Sadak, M.S. Physiological role of glycinebetaine in alleviating the deleterious effects of drought stress on canola plants (Brassica napus L.). Middle East J. Agric. Res. 2014, 3, 943–954. [Google Scholar]
- Kim, T.Y.; Jo, M.H.; Hong, J.H. Protective effect of nitric oxide against oxidative stress under UV-B radiation in maize leaves. J. Environ. Sci. 2010, 19, 1323–1334. [Google Scholar]
- Sharma, A.; Wang, J.F.; Xu, D.B.; Tao, S.C.; Chong, S.L.; Yan, D.L.; Li, Z.; Yuan, H.W.; Zheng, B.S. Melatonin regulates the functional components of photosynthesis, antioxidant system, gene expression, and metabolic pathways to induce drought resistance in grafted Carya cathayensis plants. Sci. Total Environ. 2020, 713, 136675. [Google Scholar] [CrossRef]
- Liu, J.L.; Zhang, W.L.; Hu, M.J.; Pan, Y.G.; Jiang, Y.M.; Zhang, Z.K.; Jiang, G.X. Nitric oxide is involved in melatonin-induced cold tolerance in postharvest litchi fruit. Postharvest Biol. Tec. 2023, 196, 112157. [Google Scholar] [CrossRef]
- Liu, J.L.; Yang, J.; Zhang, H.Q.; Cong, L.; Zhai, R.; Yang, C.Q.; Wang, Z.G.; Ma, F.W.; Xu, L.F. Melatonin inhibits ethylene synthesis via nitric oxide regulation to delay postharvest senescence in pears. J. Agric. Food Chem. 2019, 67, 2279–2288. [Google Scholar] [CrossRef]
- Zhu, Y.; Gao, H.; Lu, M.; Hao, C.; Pu, Z.; Guo, M.; Hou, D.; Chen, L.; Huang, X. Melatonin-nitric oxide crosstalk and their roles in the redox network in plants. Int. J. Mol. Sci. 2019, 20, 6200. [Google Scholar] [CrossRef] [PubMed]
- Allah, E.F.; Alqarawi, A.A. Mitigation of cadmium induced stress in tomato (Solanum lycopersicum L.) by selenium. Pak. J. Bot. 2016, 48, 953–961. [Google Scholar]
- Rady, M.M.; Osman, A.S. Response of growth and antioxidant system of heavy metal-contaminated tomato plants to 24-epibrassinolide. Afr. J. Agr. Res. 2012, 7, 3249–3254. [Google Scholar]
- Chmielowska-Bak, J.; Gzyl, J.; Rucinska-Sobkowiak, R.; Arasimowicz-Jelonek, M.; Deckert, J. The new insights into cadmium sensing. Front. Plant Sci. 2014, 5, 245. [Google Scholar]
- Per, T.S.; Masood, A.; Khan, N.A. Nitric oxide improves S-assimilation and GSH production to prevent inhibitory effects of cadmium stress on photosynthesis in mustard (Brassica juncea L.). Nitric Oxide 2017, 68, 111–124. [Google Scholar] [CrossRef]
- Gu, Q.; Wang, C.Y.; Xiao, Q.Q.; Chen, Z.P.; Han, Y. Melatonin confers plant cadmium tolerance: An update. Int. J. Mol. Sci. 2021, 22, 11704. [Google Scholar] [CrossRef]
- Zhao, G.; Zhao, Y.Y.; Yu, X.L.; Kiprotich, F.; Han, H.; Guan, R.Z.; Wang, R.; Shen, W.B. Nitric oxide is required for melatonin-enhanced tolerance against salinity stress in rapeseed (Brassica napus L.) seedlings. Int. J. Mol. Sci. 2018, 19, 1912. [Google Scholar] [CrossRef] [PubMed]
- Okant, M.; Kaya, C. The role of endogenous nitric oxide in melatonin-improved tolerance to lead toxicity in maize plants. Environ. Sci. Pollut. Res. 2019, 26, 11864–11874. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Liu, Z.; Zhu, L.; Ma, Z.Y.; Wang, J.F.; Zhu, J. Exogenous melatonin improves plant iron deficiency tolerance via increased accumulation of polyamine-mediated nitric oxide. Int. J. Mol. Sci. 2016, 17, 1777. [Google Scholar] [CrossRef]
- Gupta, D.K.; Pena, L.B.; Romero-Puertas, M.C.; Hernandez, A.; Inouhe, M.; Sandalio, L.M. NADPH oxidases differentially regulate ROS metabolism and nutrient uptake under cadmium toxicity. Plant Cell Environ. 2017, 40, 509–526. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.Q.; Fu, X.; Han, L.J.; Xu, C.X.; Liu, C.Y.; Bi, H.G.; Ai, X.Z. Nitric Oxide functions as a downstream signal for melatonin-induced cold tolerance in cucumber seedlings. Front. Plant Sci. 2021, 12, 686545. [Google Scholar] [CrossRef] [PubMed]
- Kaya, C.; Okant, M.; Ugurlar, F.; Alyemeni, M.N.; Ashraf, M.; Ahmad, P. Melatonin-mediated nitric oxide improves tolerance to cadmium toxicity by reducing oxidative stress in wheat plants. Chemosphere 2019, 225, 627–638. [Google Scholar] [CrossRef]
- Liu, N.; Gong, B.; Jin, Z.Y.; Wang, X.F.; Wei, M.; Yang, F.J.; Li, Y.; Shi, Q.H. Sodic alkaline stress mitigation by exogenous melatonin in tomato needs nitric oxide as a downstream signal. J. Plant Physiol. 2015, 186, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Jiao, J.Q.; Jin, M.J.; Liu, H.; Suo, J.T.; Yin, X.R.; Zhu, Q.G.; Rao, J.P. Application of melatonin in kiwifruit (Actinidia chinensis) alleviated chilling injury during cold storage. Sci. Hortic. 2022, 296, 110876. [Google Scholar] [CrossRef]
- Li, X.Y.; Wang, X.F.; Lu, L.F.; Yin, B.; Zhang, M.; Cui, X.M. Effects of exogenous nitric oxide on ascorbate-glutathione cycle in tomato seedlings roots under copper stress. Ying Yong Sheng Tai Xue Bao 2013, 24, 1023–1030. [Google Scholar]
- Shan, C.J.; Zhou, Y.; Liu, M.J. Nitric oxide participates in the regulation of the ascorbate-glutathione cycle by exogenous jasmonic acid in the leaves of wheat seedlings under drought stress. Protoplasma 2015, 252, 1397–1405. [Google Scholar] [CrossRef]
- Shalata, A.; Mittova, V.; Volokita, M.; Guy, M.; Tal, M. Response of the cultivated tomato and its wild salt-tolerant relative Lycopersiconpennellii to salt-dependent oxidative stress: The root antioxidative system. Physiol. Plant 2001, 112, 487–494. [Google Scholar] [CrossRef]
- Wei, L.J.; Zhang, J.; Wei, S.H.; Hu, D.L.; Liu, Y.Y.; Feng, L.; Li, C.X.; Qi, N.N.; Wang, C.L.; Liao, W.B. Nitric oxide enhanced salt stress tolerance in tomato seedlings, involving phytohormone equilibrium and photosynthesis. Int. J. Mol. Sci. 2022, 23, 4539. [Google Scholar] [CrossRef]
- Junglee, S.; Urban, L.; Sallanon, H.; Lopez-Lauri, F. Optimized assay for hydrogen peroxide determination in plant tissue using potassium iodide. Am. J. Analyt. Chem. 2014, 05, 730–736. [Google Scholar] [CrossRef]
- Fazeli, F.; Ghorbanli, M.; Niknam, V. Effect of drought on biomass, protein content, lipid peroxidation and antioxidant enzymes in two sesame cultivars. Biol. Plant 2007, 51, 98–103. [Google Scholar] [CrossRef]
- Arakawa, N.; Tsutsumi, K.; Sanceda, N.G.; Kurata, T.; Inagaki, C. A rapid and sensitive method for the determination of ascorbic acid using 4,7-diphenyl-l,10- phenanthroline. Agric. Biol. Chem. 1981, 45, 1289–1290. [Google Scholar] [CrossRef]
- Rao, I.M.; Terry, N. Leaf phosphate status, photosynthesis, and carbon partitioning in sugar beet. Plant Physiol. 1989, 90, 814–819. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.P.; Li, C.X.; Liu, H.W.; Zhao, Z.X.; Liao, W.B. Hydrogen gas improves seed germination in cucumber by regulating sugar and starch metabolisms. Horticulturae 2021, 7, 456. [Google Scholar] [CrossRef]
- Compton, M.E. Statistical methods suitable for the analysis of plant tissue culture data. Plant Cell Tissue Organ Cult. 1994, 37, 217–242. [Google Scholar] [CrossRef]
- Jonassen, T.M.; Imsland, A.K.; Kadowaki, S.; Stefansson, S.O. Interaction of temperature and photoperiod on growth of Atlantic halibut Hippoglossus hippoglossus L. Aquac. Res. 2000, 31, 219–227. [Google Scholar] [CrossRef]
- Gómez, M.J.R.; Maestro-Gaitán, I.; Magro, P.C.; Sobrado, V.C.; Blázquez, M.R.; Prieto, J.M. Unique nutritional features that distinguish Amaranthus cruentus L. and Chenopodium quinoa Willd seeds. Food Res. Int. 2023, 164, 112160. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Accession Number | Forward Primer | Reverse Primer |
|---|---|---|---|
| AAO | LOC101258916 | 5′-GAGCAATACGCACCTCAGATTCTCC-3′ | 5′-GCCAAGTTGAGTGAAGCCAATGC-3′ |
| AAOH | LOC101263934 | 5′- GTGCTCTCATCCCTGTTCCTTTCG-3′ | 5′-CATCAGGTCTGCCAACGGTGTG-3′ |
| APX1 | LOC778224 | 5′-AGCAGTTTCCCACTCTCTCCCATG-3′ | 5′-CAACAGCAACAACACCAGCCAAC-3′ |
| APX6 | LOC778341 | 5′-GCTACCAGGTTCATTGCTCTTCTCC-3′ | 5′-TCGGCGAAGCGAATGTACTGAATC-3′ |
| MDHAR | LOC778288 | 5′-CCATTTGGCGATTTCGGCTTGTAAG-3′ | 5′-ACACCCGCTCGCTCTCATCC-3′ |
| DHAR1 | LOC778229 | 5′-AAGAAGTGGAGTGTGCCTGAAAGC-3′ | 5′-CACGCATACAAGGACACGGTGAG-3′ |
| DHAR2 | LOC778343 | 5′-CACGAAGTTCAGAGCACCCAGAAG-3′ | 5′-CAGTCACCGAGCTTGTTAGGAGTTG-3′ |
| GR | LOC100301931 | 5′-CCGCCCATTTATCCCAGACATTCC-3′ | 5′-GTCAGGCTTCGTTGGCAAATCAAG-3′ |
| Actin | NC_015447 | 5′-AATGAACTTCGTGTGGCTCCAGAG-3′ | 5′-ATGGCAGGGGTGTTGAAGGTTTC-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, J.; Wei, Z.; Lu, X.; Liu, Y.; Yu, W.; Li, C. Involvement of Nitric Oxide and Melatonin Enhances Cadmium Resistance of Tomato Seedlings through Regulation of the Ascorbate–Glutathione Cycle and ROS Metabolism. Int. J. Mol. Sci. 2023, 24, 9526. https://doi.org/10.3390/ijms24119526
Xu J, Wei Z, Lu X, Liu Y, Yu W, Li C. Involvement of Nitric Oxide and Melatonin Enhances Cadmium Resistance of Tomato Seedlings through Regulation of the Ascorbate–Glutathione Cycle and ROS Metabolism. International Journal of Molecular Sciences. 2023; 24(11):9526. https://doi.org/10.3390/ijms24119526
Chicago/Turabian StyleXu, Junrong, Zhien Wei, Xuefang Lu, Yunzhi Liu, Wenjin Yu, and Changxia Li. 2023. "Involvement of Nitric Oxide and Melatonin Enhances Cadmium Resistance of Tomato Seedlings through Regulation of the Ascorbate–Glutathione Cycle and ROS Metabolism" International Journal of Molecular Sciences 24, no. 11: 9526. https://doi.org/10.3390/ijms24119526