
High-Fat Diet Consumption Induces Neurobehavioral Abnormalities and Neuronal Morphological Alterations Accompanied by Excessive Microglial Activation in the Medial Prefrontal Cortex in Adolescent Mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
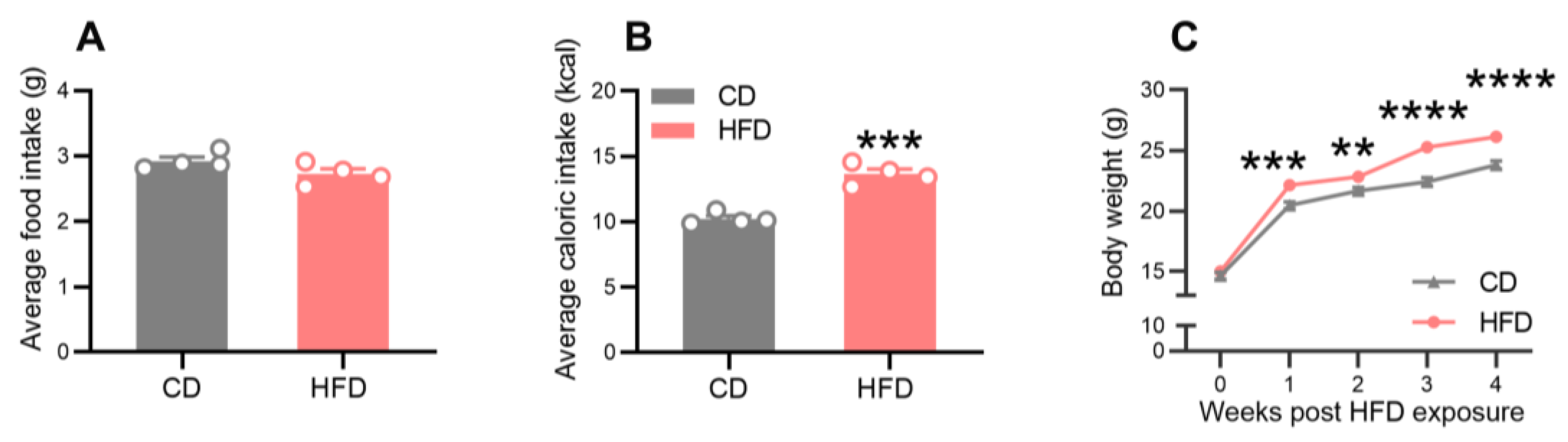
2.1. Effects of a HFD on Food and Energy Intake and Body Weight
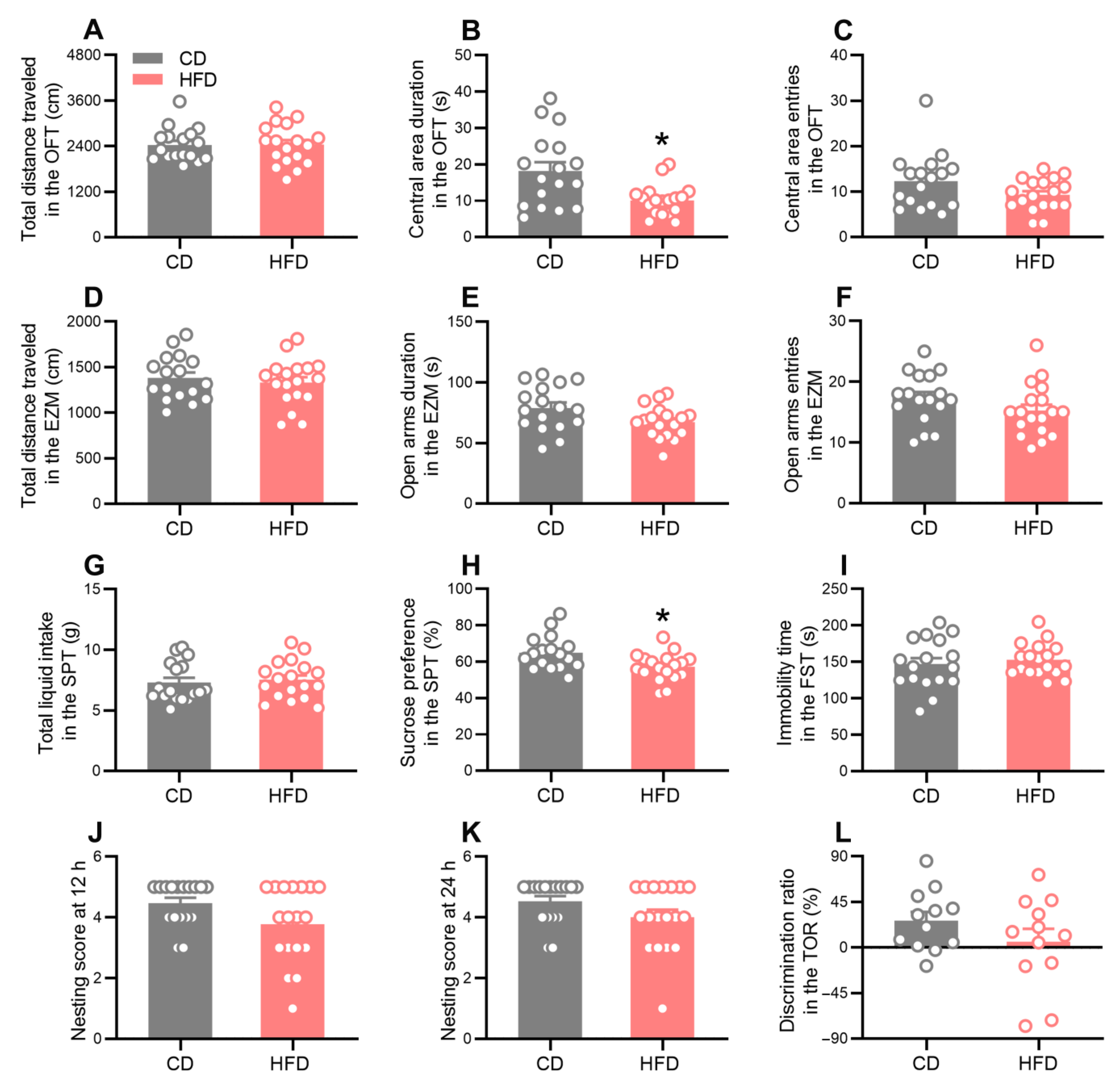
2.2. HFD-Fed Adolescent Mice Develop Anxiety- and Depression-Like Behavior
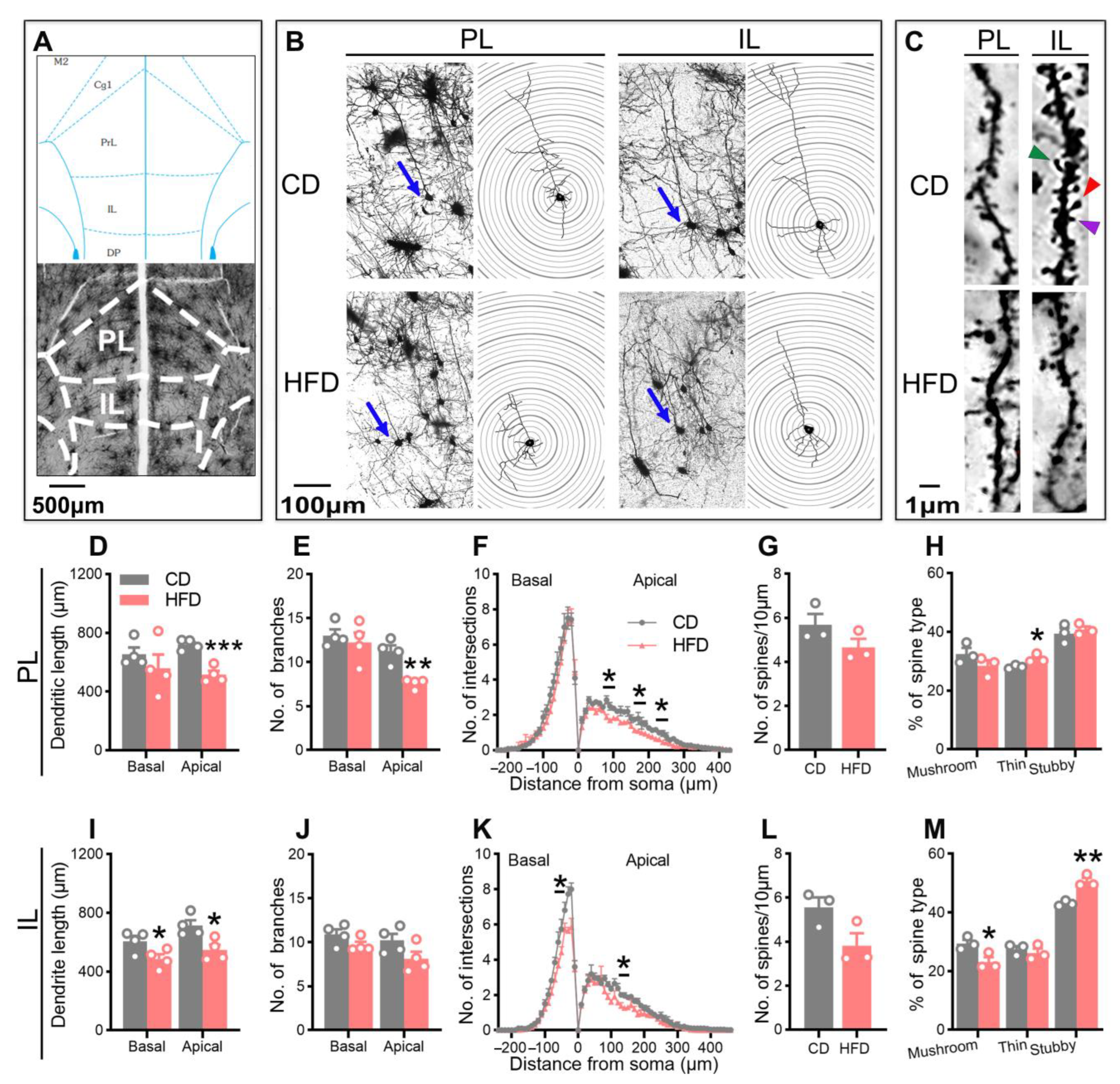
2.3. Adolescent HFD Consumption Alters Pyramidal Neuronal Morphology in the mPFC
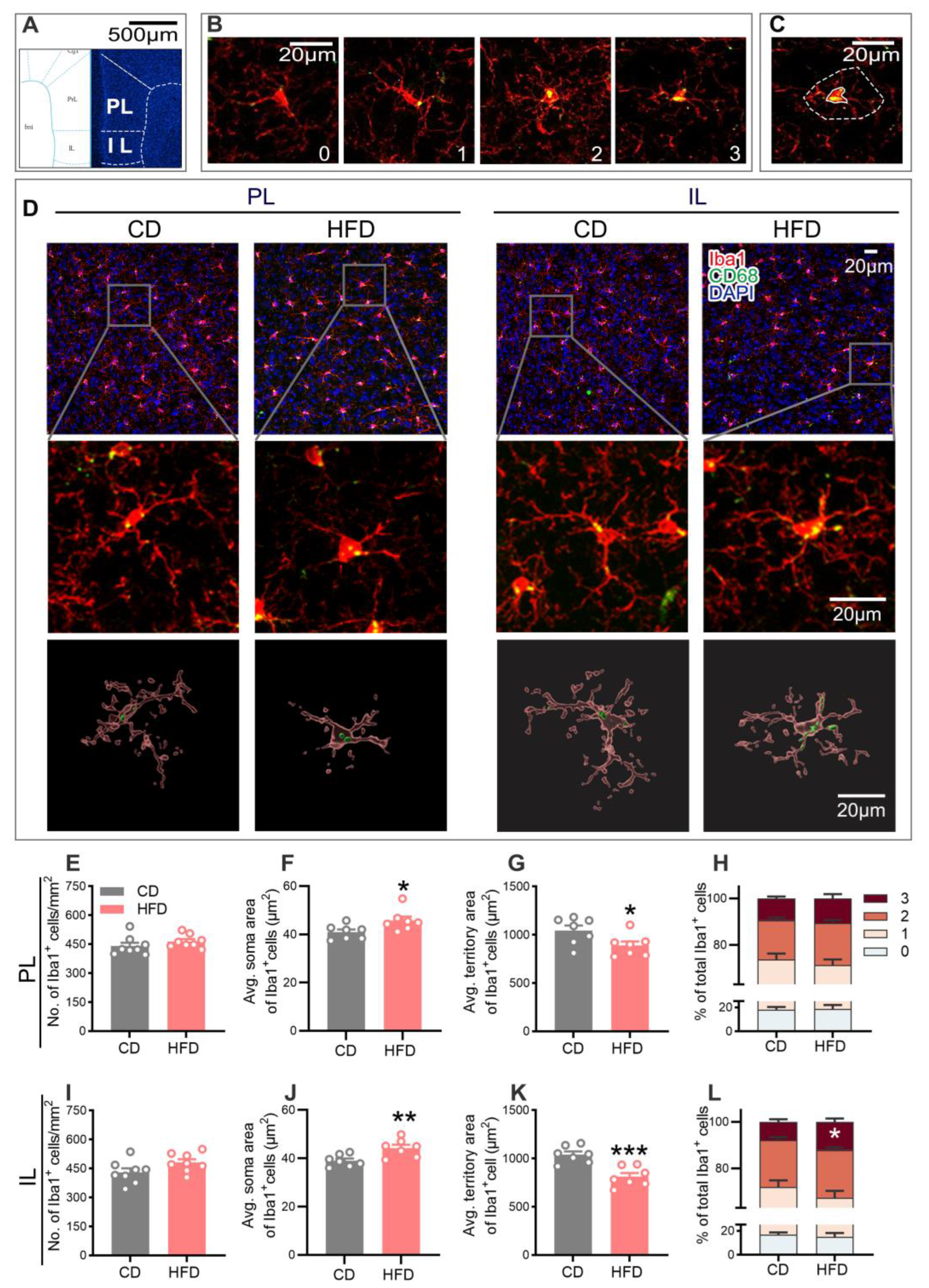
2.4. HFD-Fed Adolescent Mice Show Altered Microglial Activity in the mPFC
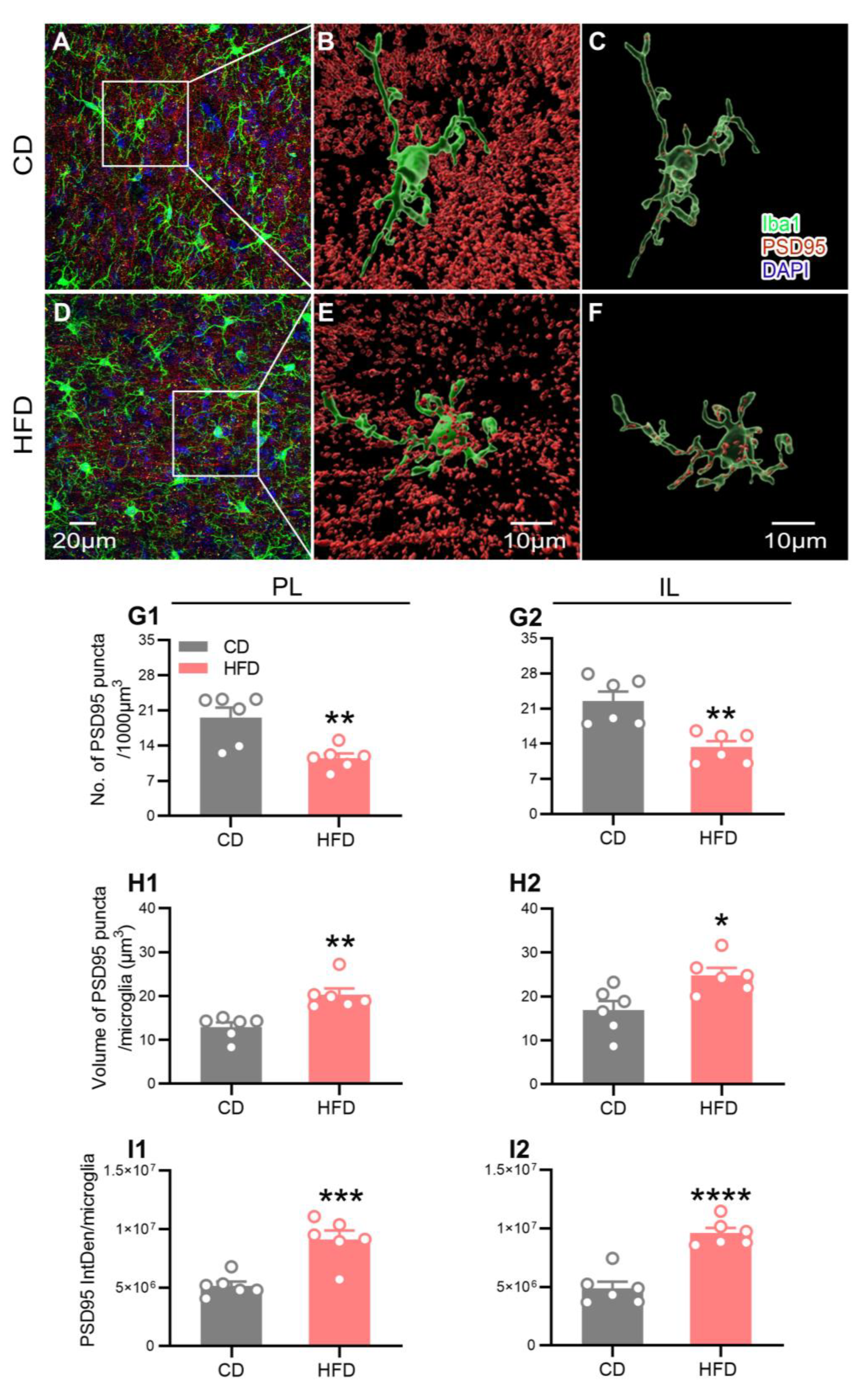
2.5. HFD Intake in Adolescence Alters Microglial Engulfment of PSD95
2.6. Correlations between Behavior, Neuroplasticity, and Microglia
3. Discussion
4. Materials and Methods
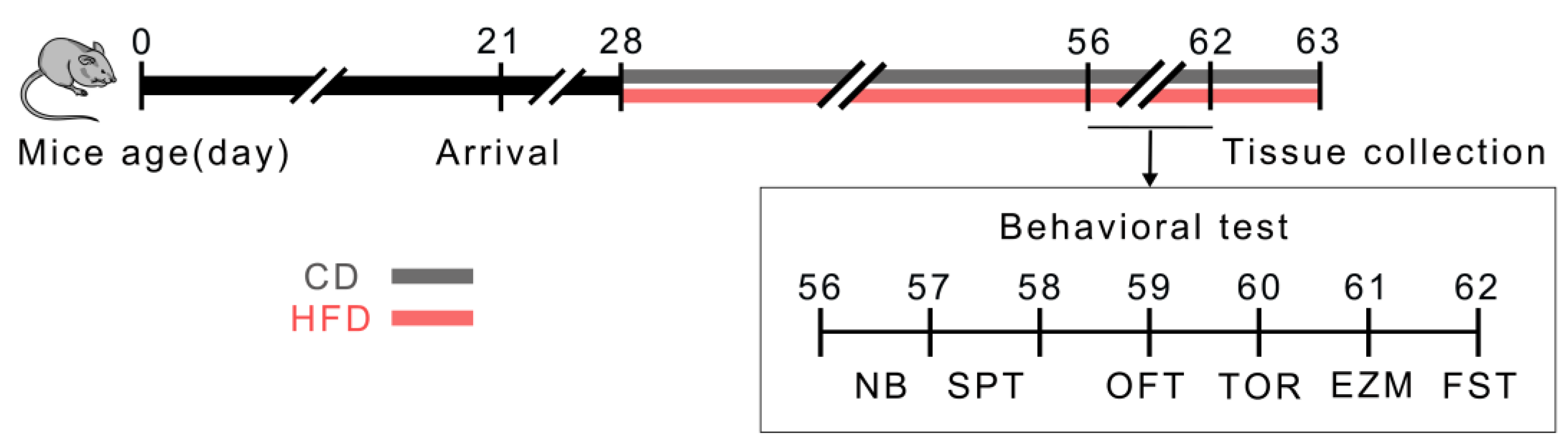
4.1. Animals and Experimental Design
4.2. Behavioral Tests
4.2.1. OFT
4.2.2. EZM Test
4.2.3. SPT
4.2.4. FST
4.2.5. NB Test
4.2.6. TOR Task
4.3. Tissue Collection
4.4. Golgi–Cox Impregnation and Morphometric Analyses
4.5. Immunohistochemistry
4.6. Quantitative Analysis of Immunohistological Data
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Valladolid-Acebes, I.; Fole, A.; Martín, M.; Morales, L.; Cano, M.V.; Ruiz-Gayo, M.; Del Olmo, N. Spatial memory impairment and changes in hippocampal morphology are triggered by high-fat diets in adolescent mice. Is there a role of leptin? Neurobiol. Learn. Mem. 2013, 106, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Yang, C.; Wang, C.; Li, H.; Zhao, J.; Kang, X.; Liu, Z.; Chen, L.; Chen, X.; Pu, T.; et al. High-Fat Diet Consumption in Adolescence Induces Emotional Behavior Alterations and Hippocampal Neurogenesis Deficits Accompanied by Excessive Microglial Activation. Int. J. Mol. Sci. 2022, 23, 8316. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Guo, C.; Wang, C.; Hu, C.; Chen, H. Effect of early life stress on anxiety and depressive behaviors in adolescent mice. Brain Behav. 2020, 10, e01526. [Google Scholar] [CrossRef] [PubMed]
- Blakemore, S.-J. Adolescence and mental health. Lancet 2019, 393, 2030–2031. [Google Scholar] [CrossRef] [PubMed]
- Andersen, S.L.; Teicher, M.H. Stress, sensitive periods and maturational events in adolescent depression. Trends Neurosci. 2008, 31, 183–191. [Google Scholar] [CrossRef]
- Miller, L.; Ropper, A.H.; Campo, J.V. Depression in Adolescents. N. Engl. J. Med. 2021, 385, 445–449. [Google Scholar] [CrossRef]
- Hemmati, A.; Ghoreishy, S.M.; Karami, K.; Imani, H.; Farsani, G.M.; Mousavi, S.E.; Asoudeh, F.; Shariati-Bafghi, S.-E.; Karamati, M. The association between dietary patterns and depression in adolescents: A cross-sectional study. Clin. Nutr. ESPEN 2021, 46, 271–275. [Google Scholar] [CrossRef]
- Kaczmarczyk, M.M.; Machaj, A.S.; Chiu, G.S.; Lawson, M.A.; Gainey, S.J.; York, J.M.; Meling, D.D.; Martin, S.A.; Kwakwa, K.A.; Newman, A.F.; et al. Methylphenidate prevents high-fat diet (HFD)-induced learning/memory impairment in juvenile mice. Psychoneuroendocrinology 2013, 38, 1553–1564. [Google Scholar] [CrossRef]
- Yang, C.; Yao, X.; Zhang, H.; Wang, C.; Zhao, J.; Xu, D.; Xiao, Y.; Li, Q.; Zhuang, H.; Kang, X.; et al. Effects of Prolonged High-Fat Diet Consumption Starting at Different Ages on Behavioral Parameters and Hippocampal Neuroplasticity in Male Mice. J. Integr. Neurosci. 2023, 22, 16. [Google Scholar] [CrossRef]
- Neutzling, M.B.; Araújo, C.L.P.; Vieira, M.d.F.A.; Hallal, P.C.; Menezes, A.M.B. Frequency of high-fat and low-fiber diets among adolescents. Rev. De Saude Publica 2007, 41, 336–342. [Google Scholar] [CrossRef]
- Reichelt, A.C.; Lemieux, C.A.; Princz-Lebel, O.; Singh, A.; Bussey, T.J.; Saksida, L.M. Age-dependent and region-specific alteration of parvalbumin neurons, perineuronal nets and microglia in the mouse prefrontal cortex and hippocampus following obesogenic diet consumption. Sci. Rep. 2021, 11, 5593. [Google Scholar] [CrossRef] [PubMed]
- Baker, K.D.; Loughman, A.; Spencer, S.J.; Reichelt, A.C. The impact of obesity and hypercaloric diet consumption on anxiety and emotional behavior across the lifespan. Neurosci. Biobehav. Rev. 2017, 83, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Hoover, W.B.; Vertes, R.P. Anatomical analysis of afferent projections to the medial prefrontal cortex in the rat. Brain Struct. Funct. 2007, 212, 149–179. [Google Scholar] [CrossRef] [PubMed]
- Duman, R.S.; Aghajanian, G.K. Synaptic Dysfunction in Depression: Potential Therapeutic Targets. Science 2012, 338, 68–72. [Google Scholar] [CrossRef]
- Deyama, S.; Kaneda, K. Role of neurotrophic and growth factors in the rapid and sustained antidepressant actions of ketamine. Neuropharmacology 2022, 224, 109335. [Google Scholar] [CrossRef]
- Lloyd, K.; Reyes, T. High fat diet consumption restricted to adolescence has minimal effects on adult executive function that vary by sex. Nutr. Neurosci. 2022, 25, 801–811. [Google Scholar] [CrossRef]
- Sun, Q.; Zhang, J.; Li, A.; Yao, M.; Liu, G.; Chen, S.; Luo, Y.; Wang, Z.; Gong, H.; Li, X.; et al. Acetylcholine deficiency disrupts extratelencephalic projection neurons in the prefrontal cortex in a mouse model of Alzheimer’s disease. Nat. Commun. 2022, 13, 998. [Google Scholar] [CrossRef]
- Yan, Z.; Rein, B. Mechanisms of synaptic transmission dysregulation in the prefrontal cortex: Pathophysiological implications. Mol. Psychiatry 2022, 27, 445–465. [Google Scholar] [CrossRef]
- Teissier, A.; Le Magueresse, C.; Olusakin, J.; da Costa, B.L.S.A.; De Stasi, A.M.; Bacci, A.; Imamura Kawasawa, Y.; Vaidya, V.A.; Gaspar, P. Early-life stress impairs postnatal oligodendrogenesis and adult emotional behaviour through activity-dependent mechanisms. Mol. Psychiatry 2020, 25, 1159–1174. [Google Scholar] [CrossRef]
- Klune, C.B.; Jin, B.; DeNardo, L.A. Linking mPFC circuit maturation to the developmental regulation of emotional memory and cognitive flexibility. eLife 2021, 10, e64567. [Google Scholar] [CrossRef]
- Hou, Y.; Wang, Y.; Song, S.; Zuo, Y.; Zhang, H.; Bai, C.; Zhao, H.; Zhang, T. Long-term variable photoperiod exposure impairs the mPFC and induces anxiety and depression-like behavior in male wistar rats. Exp. Neurol. 2022, 347, 113908. [Google Scholar] [CrossRef]
- Zhuang, P.-C.; Tan, Z.-N.; Jia, Z.-Y.; Wang, B.; Grady, J.J.; Ma, X.-M. Treadmill Exercise Reverses Depression Model-Induced Alteration of Dendritic Spines in the Brain Areas of Mood Circuit. Front. Behav. Neurosci. 2019, 13, 93. [Google Scholar] [CrossRef] [PubMed]
- Dingess, P.M.; Darling, R.A.; Kurt Dolence, E.; Culver, B.W.; Brown, T.E. Exposure to a diet high in fat attenuates dendritic spine density in the medial prefrontal cortex. Brain Struct. Funct. 2017, 222, 1077–1085. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Li, Y.; Li, X.; Zeng, F.; Rao, Y.; He, Y.; Wang, Y.; Liu, M.; Li, D.; Xu, Z.; et al. Microglial debris is cleared by astrocytes via C4b-facilitated phagocytosis and degraded via RUBICON-dependent noncanonical autophagy in mice. Nat. Commun. 2022, 13, 6233. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, H.; Yao, X.; Li, H.; Li, Q.; Yang, C.; Wang, C.; Xu, D.; Xiao, Y.; Gao, Y.; Gao, J.; et al. Long-term high-fat diet consumption by mice throughout adulthood induces neurobehavioral alterations and hippocampal neuronal remodeling accompanied by augmented microglial lipid accumulation. Brain Behav. Immun. 2022, 100, 155–171. [Google Scholar] [CrossRef] [PubMed]
- Folick, A.; Koliwad, S.K.; Valdearcos, M. Microglial Lipid Biology in the Hypothalamic Regulation of Metabolic Homeostasis. Front. Endocrinol. 2021, 12, 668396. [Google Scholar] [CrossRef] [PubMed]
- Wake, H.; Moorhouse, A.J.; Miyamoto, A.; Nabekura, J. Microglia: Actively surveying and shaping neuronal circuit structure and function. Trends Neurosci. 2013, 36, 209–217. [Google Scholar] [CrossRef]
- Dheen, S.T.; Kaur, C.; Ling, E.-A. Microglial Activation and its Implications in the Brain Diseases. Curr. Med. Chem. 2007, 14, 1189–1197. [Google Scholar] [CrossRef]
- Schafer, D.P.; Lehrman, E.K.; Kautzman, A.G.; Koyama, R.; Mardinly, A.R.; Yamasaki, R.; Ransohoff, R.M.; Greenberg, M.E.; Barres, B.A.; Stevens, B. Microglia sculpt postnatal neural circuits in an activity and complement-dependent manner. Neuron 2012, 74, 691–705. [Google Scholar] [CrossRef]
- Wohleb, E.S.; Terwilliger, R.; Duman, C.H.; Duman, R.S. Stress-Induced Neuronal Colony Stimulating Factor 1 Provokes Microglia-Mediated Neuronal Remodeling and Depressive-like Behavior. Biol. Psychiatry 2018, 83, 38–49. [Google Scholar] [CrossRef]
- Wright-Jin, E.C.; Gutmann, D.H. Microglia as Dynamic Cellular Mediators of Brain Function. Trends Mol. Med. 2019, 25, 967–979. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Ohgidani, M.; Kuwano, N.; Chrétien, F.; de la Grandmaison, G.L.; Onaya, M.; Tominaga, I.; Setoyama, D.; Kang, D.; Mimura, M.; et al. Suicide and Microglia: Recent Findings and Future Perspectives Based on Human Studies. Front. Cell Neurosci. 2019, 13, 31. [Google Scholar] [CrossRef]
- Nie, X.; Kitaoka, S.; Tanaka, K.; Segi-Nishida, E.; Imoto, Y.; Ogawa, A.; Nakano, F.; Tomohiro, A.; Nakayama, K.; Taniguchi, M.; et al. The Innate Immune Receptors TLR2/4 Mediate Repeated Social Defeat Stress-Induced Social Avoidance through Prefrontal Microglial Activation. Neuron 2018, 99, 464–479.e467. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Franklin, K.B. The Mouse Brain in Stereotaxic Coordinates, 3rd ed.; Academic Press: Cambridge, MA, USA, 2008. [Google Scholar]
- Salter, M.W.; Stevens, B. Microglia emerge as central players in brain disease. Nat. Med. 2017, 23, 1018–1027. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Beja-Glasser, V.F.; Nfonoyim, B.M.; Frouin, A.; Li, S.; Ramakrishnan, S.; Merry, K.M.; Shi, Q.; Rosenthal, A.; Barres, B.A.; et al. Complement and microglia mediate early synapse loss in Alzheimer mouse models. Science 2016, 352, 712–716. [Google Scholar] [CrossRef]
- Cardozo, P.L.; de Lima, I.B.Q.; Maciel, E.M.A.; Silva, N.C.; Dobransky, T.; Ribeiro, F.M. Synaptic Elimination in Neurological Disorders. Curr. Neuropharmacol. 2019, 17, 1071–1095. [Google Scholar] [CrossRef]
- Ljungberg, T.; Bondza, E.; Lethin, C. Evidence of the Importance of Dietary Habits Regarding Depressive Symptoms and Depression. Int. J. Environ. Res. Public Health 2020, 17, 1616. [Google Scholar] [CrossRef]
- Wu, H.; Lv, W.; Pan, Q.; Kalavagunta, P.K.; Liu, Q.; Qin, G.; Cai, M.; Zhou, L.; Wang, T.; Xia, Z.; et al. Simvastatin therapy in adolescent mice attenuates HFD-induced depression-like behavior by reducing hippocampal neuroinflammation. J. Affect. Disord. 2019, 243, 83–95. [Google Scholar] [CrossRef]
- Paus, T.; Keshavan, M.; Giedd, J.N. Why do many psychiatric disorders emerge during adolescence? Nat. Rev. Neurosci. 2008, 9, 947–957. [Google Scholar] [CrossRef]
- Thapar, A.; Collishaw, S.; Pine, D.S.; Thapar, A.K. Depression in adolescence. Lancet 2012, 379, 1056–1067. [Google Scholar] [CrossRef]
- Liu, M.-Y.; Yin, C.-Y.; Zhu, L.-J.; Zhu, X.-H.; Xu, C.; Luo, C.-X.; Chen, H.; Zhu, D.-Y.; Zhou, Q.-G. Sucrose preference test for measurement of stress-induced anhedonia in mice. Nat. Protoc. 2018, 13, 1686–1698. [Google Scholar] [CrossRef]
- Wu, J.; Zhu, Y.; Zhou, L.; Lu, Y.; Feng, T.; Dai, M.; Liu, J.; Xu, W.; Cheng, W.; Sun, F.; et al. Parasite-Derived Excretory-Secretory Products Alleviate Gut Microbiota Dysbiosis and Improve Cognitive Impairment Induced by a High-Fat Diet. Front. Immunol. 2021, 12, 710513. [Google Scholar] [CrossRef]
- Chen, Z.; Sui, G.; Wang, L.; Yang, C.; Wang, F. High-fat diet induced hippocampal CREB dysfunction, cognitive impairment and depression-like behaviors via downregulation of interleukin-2 in the mice. Metab. Brain Dis. 2022, 37, 1163–1174. [Google Scholar] [CrossRef] [PubMed]
- Yoshizaki, K.; Asai, M.; Hara, T. High-Fat Diet Enhances Working Memory in the Y-Maze Test in Male C57BL/6J Mice with Less Anxiety in the Elevated Plus Maze Test. Nutrients 2020, 12, 2036. [Google Scholar] [CrossRef] [PubMed]
- Giménez-Llort, L.; Torres-Lista, V. Social Nesting, Animal Welfare, and Disease Monitoring. Animals 2021, 11, 1079. [Google Scholar] [CrossRef] [PubMed]
- Suwaluk, A.; Chutabhakdikul, N. Long-term effects of prenatal stress on the development of prefrontal cortex in the adolescent offspring. J. Chem. Neuroanat. 2022, 125, 102169. [Google Scholar] [CrossRef]
- Liu, L.; Roquet, D.; Ahmed, R.M.; Hodges, J.R.; Piguet, O.; Irish, M. Examining prefrontal contributions to past- and future-oriented memory disturbances in daily life in dementia. Cortex 2021, 134, 307–319. [Google Scholar] [CrossRef]
- Duman, R.S.; Aghajanian, G.K.; Sanacora, G.; Krystal, J.H. Synaptic plasticity and depression: New insights from stress and rapid-acting antidepressants. Nat. Med. 2016, 22, 238–249. [Google Scholar] [CrossRef]
- Lyttle, K.; Ohmura, Y.; Konno, K.; Yoshida, T.; Izumi, T.; Watanabe, M.; Yoshioka, M. Repeated fluvoxamine treatment recovers juvenile stress-induced morphological changes and depressive-like behavior in rats. Brain Res. 2015, 1616, 88–100. [Google Scholar] [CrossRef]
- Xu, H.; Yu, Z.-H.; Ge, M.-J.; Shen, J.-X.; Han, F.; Pan, C.; Chen, J.-J.; Zhu, X.-L.; Hou, W.-Y.; Hou, Y.-Q.; et al. Estradiol attenuates chronic restraint stress-induced dendrite and dendritic spine loss and cofilin1 activation in ovariectomized mice. Horm. Behav. 2021, 135, 105040. [Google Scholar] [CrossRef]
- Lee, K.J.; Kim, H.; Kim, T.S.; Park, S.-H.; Rhyu, I.J. Morphological analysis of spine shapes of Purkinje cell dendrites in the rat cerebellum using high-voltage electron microscopy. Neurosci. Lett. 2004, 359, 21–24. [Google Scholar] [CrossRef]
- Qiao, H.; Li, M.-X.; Xu, C.; Chen, H.-B.; An, S.-C.; Ma, X.-M. Dendritic Spines in Depression: What We Learned from Animal Models. Neural Plast. 2016, 2016, 8056370. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Cheng, Y.; Gong, M.; Liang, B.; Zhang, M.; Chen, Y.; Zhang, C.; Yuan, X.; Xu, J. Systematic correlation between spine plasticity and the anxiety/depression-like phenotype induced by corticosterone in mice. Neuroreport 2013, 24 Pt 2, 682–687. [Google Scholar] [CrossRef] [PubMed]
- Soztutar, E.; Colak, E.; Ulupinar, E. Gender- and anxiety level-dependent effects of perinatal stress exposure on medial prefrontal cortex. Exp. Neurol. 2016, 275, 274–284. [Google Scholar] [CrossRef]
- Leem, Y.-H.; Yoon, S.-S.; Jo, S.A. Imipramine Ameliorates Depressive Symptoms by Blocking Differential Alteration of Dendritic Spine Structure in Amygdala and Prefrontal Cortex of Chronic Stress-Induced Mice. Biomol. Ther. 2020, 28, 230–239. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; He, Y.; Sun, Z.; Ren, S.; Liu, M.; Wang, G.; Yang, J. Microglia in depression: An overview of microglia in the pathogenesis and treatment of depression. J. Neuroinflamm. 2022, 19, 132. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, R.; Soares-Cunha, C.; Domingues, A.V.; Coimbra, B.; Baptista, F.I.; Pinto, L.; Ambrósio, A.F.; Rodrigues, A.J.; Gomes, C.A. Resilience to stress and sex-specific remodeling of microglia and neuronal morphology in a rat model of anxiety and anhedonia. Neurobiol. Stress 2021, 14, 100302. [Google Scholar] [CrossRef]
- Sominsky, L.; De Luca, S.; Spencer, S.J. Microglia: Key players in neurodevelopment and neuronal plasticity. Int. J. Biochem. Cell Biol. 2018, 94, 56–60. [Google Scholar] [CrossRef]
- Block, C.L.; Eroglu, O.; Mague, S.D.; Smith, C.J.; Ceasrine, A.M.; Sriworarat, C.; Blount, C.; Beben, K.A.; Malacon, K.E.; Ndubuizu, N.; et al. Prenatal environmental stressors impair postnatal microglia function and adult behavior in males. Cell Rep. 2022, 40, 111161. [Google Scholar] [CrossRef]
- Fernández-Arjona, M.D.M.; Grondona, J.M.; Granados-Durán, P.; Fernández-Llebrez, P.; López-Ávalos, M.D. Microglia Morphological Categorization in a Rat Model of Neuroinflammation by Hierarchical Cluster and Principal Components Analysis. Front. Cell Neurosci. 2017, 11, 235. [Google Scholar] [CrossRef]
- Kim, H.-J.; Cho, M.-H.; Shim, W.H.; Kim, J.K.; Jeon, E.-Y.; Kim, D.-H.; Yoon, S.-Y. Deficient autophagy in microglia impairs synaptic pruning and causes social behavioral defects. Mol. Psychiatry 2017, 22, 1576–1584. [Google Scholar] [CrossRef] [PubMed]
- Paolicelli, R.C.; Ferretti, M.T. Function and Dysfunction of Microglia during Brain Development: Consequences for Synapses and Neural Circuits. Front. Synaptic Neurosci. 2017, 9, 9. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Zhang, W.; Huang, M.; Lin, X.; Chiou, J. Prolonged High-Fat Diet Consumption throughout Adulthood in Mice Induced Neurobehavioral Deterioration via Gut-Brain Axis. Nutrients 2023, 15, 392. [Google Scholar] [CrossRef]
- Marx, W.; Lane, M.; Hockey, M.; Aslam, H.; Berk, M.; Walder, K.; Borsini, A.; Firth, J.; Pariante, C.M.; Berding, K.; et al. Diet and depression: Exploring the biological mechanisms of action. Mol. Psychiatry 2021, 26, 134–150. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Cheng, Y.; Zhou, Y.; Du, H.; Zhang, C.; Zhao, Z.; Chen, Y.; Zhou, Z.; Mei, J.; Wu, W.; et al. High fat diet-induced obesity leads to depressive and anxiety-like behaviors in mice via AMPK/mTOR-mediated autophagy. Exp. Neurol. 2022, 348, 113949. [Google Scholar] [CrossRef]
- Dutta, S.; Sengupta, P. Men and mice: Relating their ages. Life Sci. 2016, 152, 244–248. [Google Scholar] [CrossRef] [PubMed]
- Brust, V.; Schindler, P.M.; Lewejohann, L. Lifetime development of behavioural phenotype in the house mouse (Mus musculus). Front. Zoöl. 2015, 12 (Suppl. S1), S17. [Google Scholar] [CrossRef]
- Nizinska, K.; Szydlowska, K.; Vouros, A.; Kiryk, A.; Stepniak, A.; Vasilaki, E.; Lukasiuk, K. Behavioral characteristics as potential biomarkers of the development and phenotype of epilepsy in a rat model of temporal lobe epilepsy. Sci. Rep. 2021, 11, 8665. [Google Scholar] [CrossRef]
- Li, M.; Li, C.; Yu, H.; Cai, X.; Shen, X.; Sun, X.; Wang, J.; Zhang, Y.; Wang, C. Lentivirus-mediated interleukin-1β (IL-1β) knock-down in the hippocampus alleviates lipopolysaccharide (LPS)-induced memory deficits and anxiety- and depression-like behaviors in mice. J. Neuroinflamm. 2017, 14, 190. [Google Scholar] [CrossRef]
- Mi, X.; Zeng, G.-R.; Liu, J.-Q.; Luo, Z.-S.; Zhang, L.; Dai, X.-M.; Fang, W.-T.; Zhang, J.; Chen, X.-C. Ganoderma Lucidum Triterpenoids Improve Maternal Separation-Induced Anxiety- and Depression-like Behaviors in Mice by Mitigating Inflammation in the Periphery and Brain. Nutrients 2022, 14, 2268. [Google Scholar] [CrossRef]
- Yu, X.; Bai, Y.; Han, B.; Ju, M.; Tang, T.; Shen, L.; Li, M.; Yang, L.; Zhang, Z.; Hu, G.; et al. Extracellular vesicle-mediated delivery of circDYM alleviates CUS-induced depressive-like behaviours. J. Extracell. Vesicles 2022, 11, e12185. [Google Scholar] [CrossRef] [PubMed]
- Harauma, A.; Sagisaka, T.; Horii, T.; Watanabe, Y.; Moriguchi, T. The influence of n-3 fatty acids on maternal behavior and brain monoamines in the perinatal period. Prostaglandins Leukot. Essent. Fat. Acids 2016, 107, 1–7. [Google Scholar] [CrossRef]
- Warburton, E.C.; Brown, M.W. Findings from animals concerning when interactions between perirhinal cortex, hippocampus and medial prefrontal cortex are necessary for recognition memory. Neuropsychologia 2010, 48, 2262–2272. [Google Scholar] [CrossRef] [PubMed]
- Hunsaker, M.R.; Goodrich-Hunsaker, N.J.; Willemsen, R.; Berman, R.F. Temporal ordering deficits in female CGG KI mice heterozygous for the fragile X premutation. Behav. Brain Res. 2010, 213, 263–268. [Google Scholar] [CrossRef]
- Liu, L.; Xuan, C.; Shen, P.; He, T.; Chang, Y.; Shi, L.; Tao, S.; Yu, Z.; Brown, R.E.; Wang, J. Hippocampal Mechanisms Underlying Impairment in Spatial Learning Long After Establishment of Noise-Induced Hearing Loss in CBA Mice. Front. Syst. Neurosci. 2018, 12, 35. [Google Scholar] [CrossRef] [PubMed]
- Cao, P.; Chen, C.; Liu, A.; Shan, Q.; Zhu, X.; Jia, C.; Peng, X.; Zhang, M.; Farzinpour, Z.; Zhou, W.; et al. Early-life inflammation promotes depressive symptoms in adolescence via microglial engulfment of dendritic spines. Neuron 2021, 109, 2573–2589.e2579. [Google Scholar] [CrossRef]
- Fernández-Mendívil, C.; Luengo, E.; Trigo-Alonso, P.; García-Magro, N.; Negredo, P.; López, M.G. Protective role of microglial HO-1 blockade in aging: Implication of iron metabolism. Redox Biol. 2021, 38, 101789. [Google Scholar] [CrossRef]
- Filipello, F.; Morini, R.; Corradini, I.; Zerbi, V.; Canzi, A.; Michalski, B.; Erreni, M.; Markicevic, M.; Starvaggi-Cucuzza, C.; Otero, K.; et al. The Microglial Innate Immune Receptor TREM2 Is Required for Synapse Elimination and Normal Brain Connectivity. Immunity 2018, 48, 979–991.e978. [Google Scholar] [CrossRef]
- Schafer, D.P.; Lehrman, E.K.; Heller, C.T.; Stevens, B. An engulfment assay: A protocol to assess interactions between CNS phagocytes and neurons. J. Vis. Exp. 2014, 88, e51482. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, C.; Li, H.; Chen, C.; Yao, X.; Yang, C.; Yu, Z.; Ren, J.; Ming, Y.; Huang, Y.; Rong, Y.; et al. High-Fat Diet Consumption Induces Neurobehavioral Abnormalities and Neuronal Morphological Alterations Accompanied by Excessive Microglial Activation in the Medial Prefrontal Cortex in Adolescent Mice. Int. J. Mol. Sci. 2023, 24, 9394. https://doi.org/10.3390/ijms24119394
Wang C, Li H, Chen C, Yao X, Yang C, Yu Z, Ren J, Ming Y, Huang Y, Rong Y, et al. High-Fat Diet Consumption Induces Neurobehavioral Abnormalities and Neuronal Morphological Alterations Accompanied by Excessive Microglial Activation in the Medial Prefrontal Cortex in Adolescent Mice. International Journal of Molecular Sciences. 2023; 24(11):9394. https://doi.org/10.3390/ijms24119394
Chicago/Turabian StyleWang, Conghui, Hong Li, Chen Chen, Xiuting Yao, Chenxi Yang, Zhehao Yu, Jiayi Ren, Yue Ming, Yi Huang, Yi Rong, and et al. 2023. "High-Fat Diet Consumption Induces Neurobehavioral Abnormalities and Neuronal Morphological Alterations Accompanied by Excessive Microglial Activation in the Medial Prefrontal Cortex in Adolescent Mice" International Journal of Molecular Sciences 24, no. 11: 9394. https://doi.org/10.3390/ijms24119394