
The Expression of the StNRAMP2 Gene Determined the Accumulation of Cadmium in Different Tissues of Potato
 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
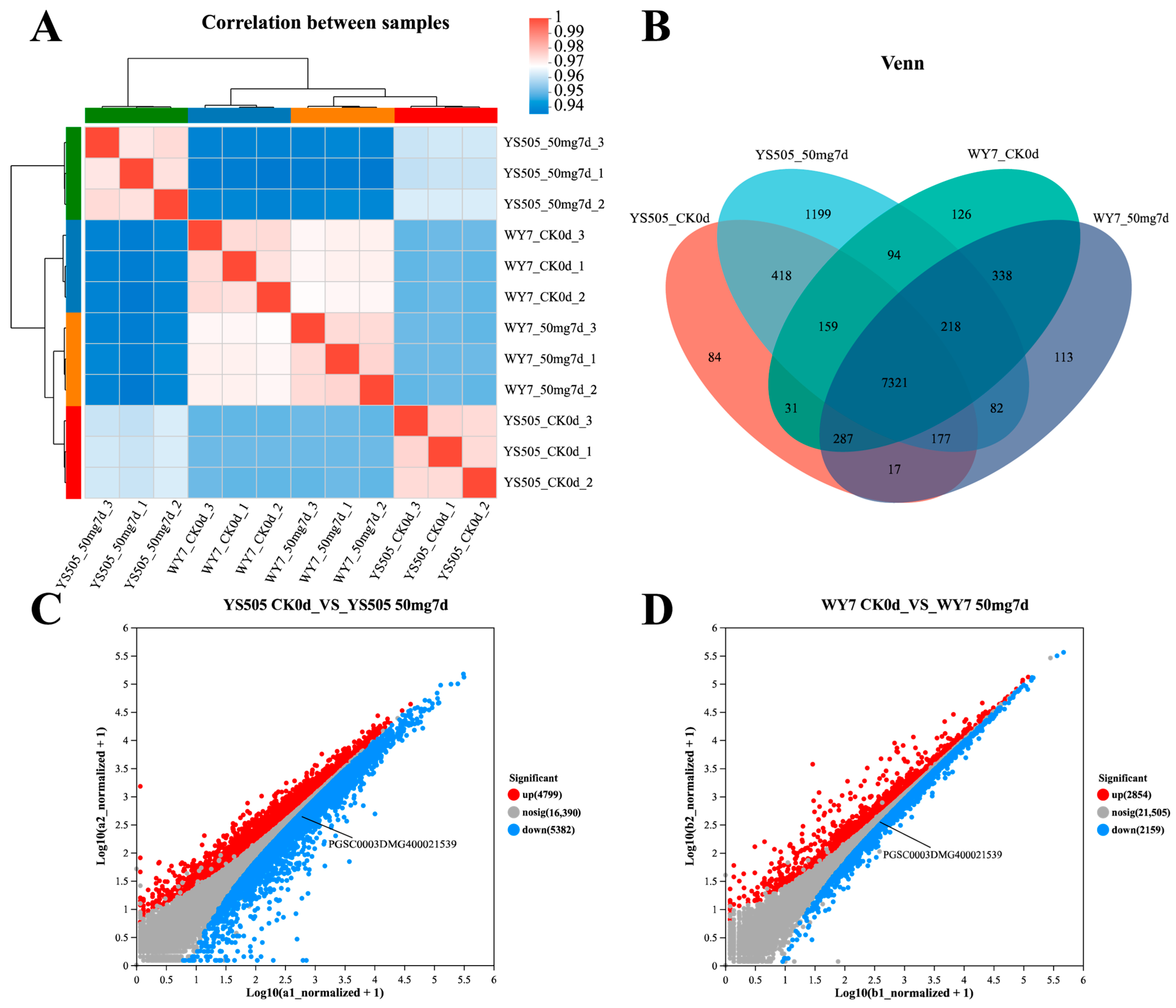
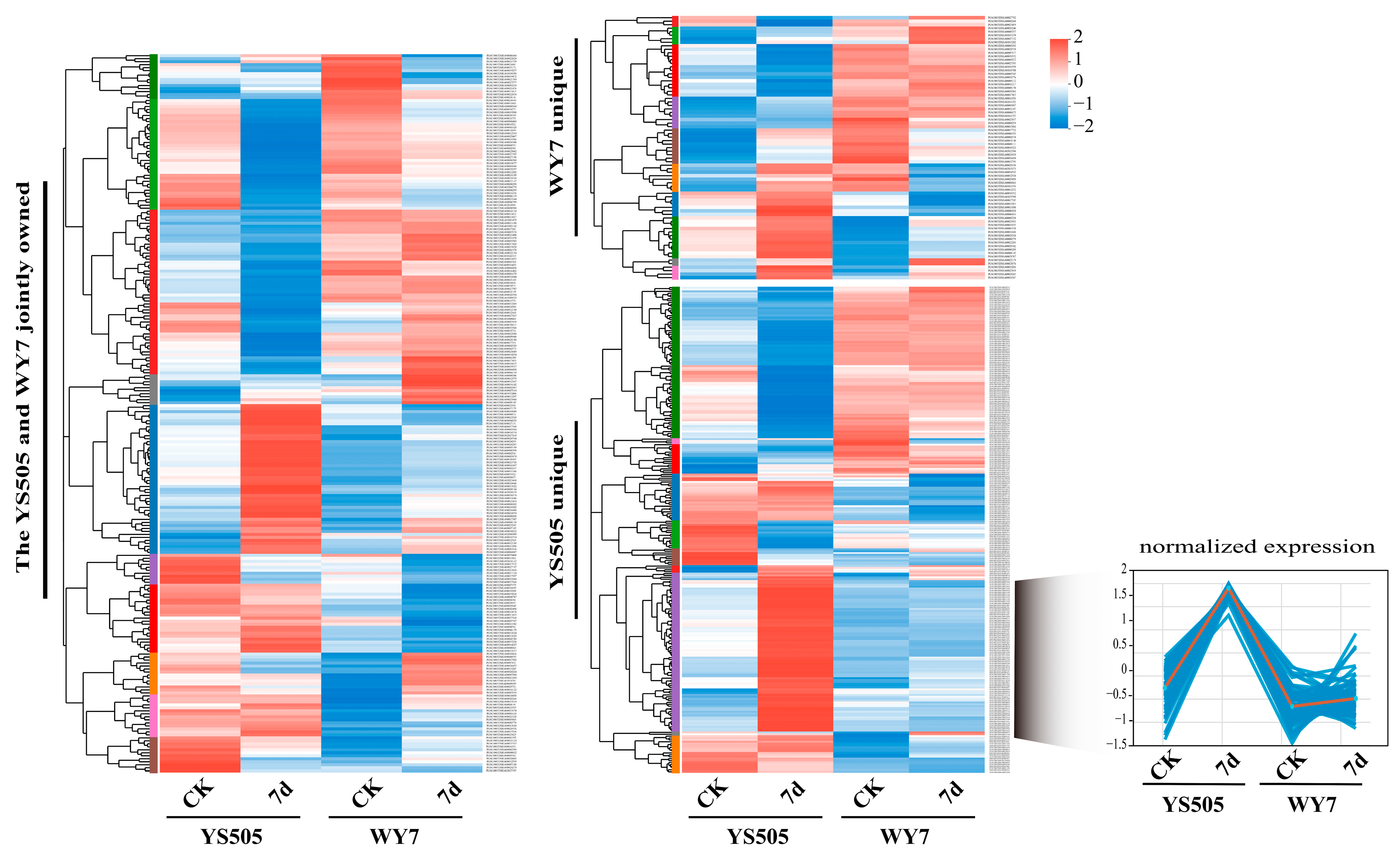
2.1. Gene Expression Patterns of Potato under Cadmium Stress for Different Days
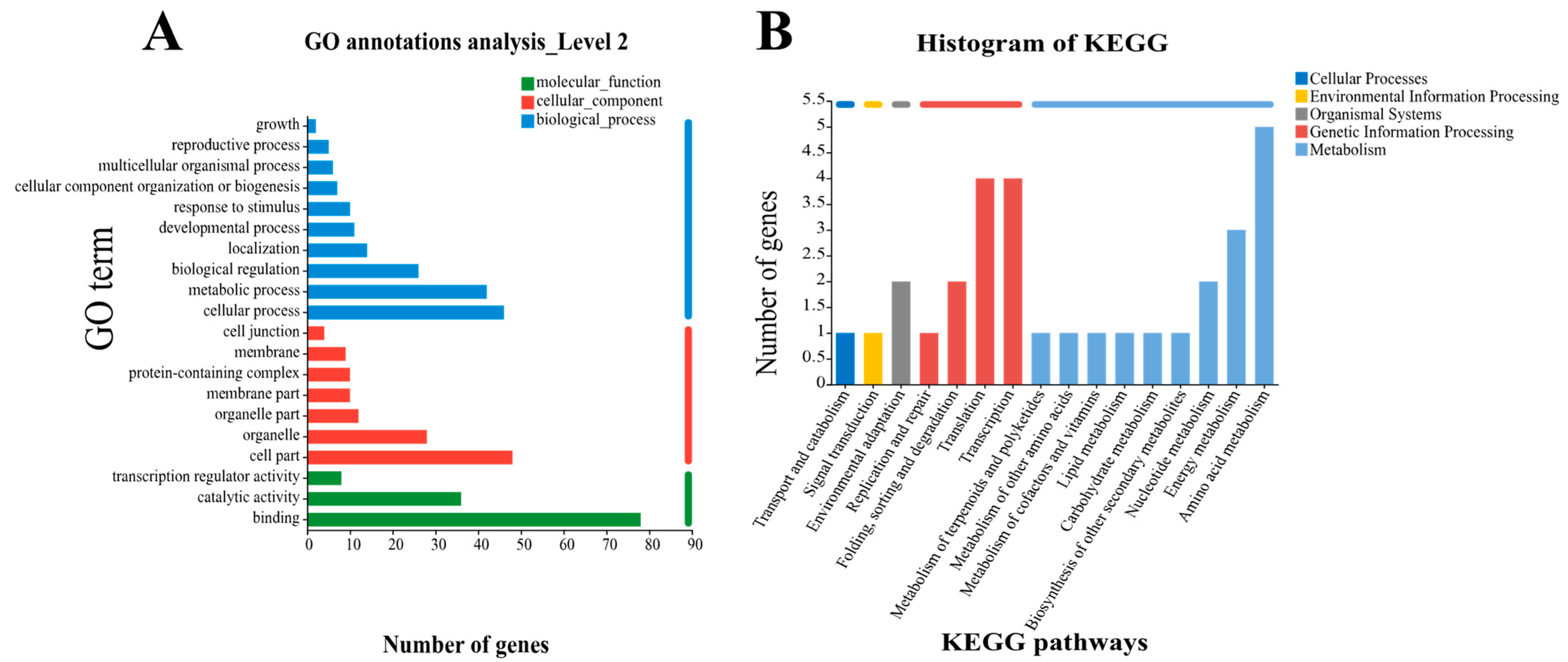
2.2. Identification of Metal Tolerance Genes
2.3. Qualitative and Quantitative Analysis of StNRAMP2
2.4. Cloning and Expression of StNRAMP2
2.5. Silencing StNRAMP2 Significantly Affected Potato REDOX Levels
2.6. StNRAMP2 Was Positively Correlated with Cadmium Tolerance in Plants
3. Discussion
3.1. RNA-Seq Screened 85 Cadmium Tolerance Genes in Potato
3.2. The Expression of StNRAMP2 Was Positively Correlated with the Accumulation of Cadmium in SOLANACEAE
3.3. Silencing StNRAMP2 Decreased the Activity of Antioxidant Enzymes in Different Tissues of Potato
4. Materials and Methods
4.1. Material Acquisition and Processing
4.1.1. Carriers and Bacteriocins
4.1.2. Plant Materials and Treatment
4.2. Transcriptome Screening of Cadmium Tolerance Candidate Genes in Potato
4.2.1. Transcriptome Sequencing
4.2.2. Transcriptome Analysis of Candidate Genes
4.3. Cloning and Transformation of StNRAMP2
4.4. Expression Patterns of StNRAMP2 in Different Tissues under Cadmium Stress
4.5. Determination of Antioxidant Activity of Potato
4.6. Determination of Cadmium Content in Plants
4.7. Statistic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vareda, J.P.; Valente, A.J.M.; Duraes, L. Assessment of heavy metal pollution from anthropogenic activities and remediation strategies: A review. J. Environ. Manag. 2019, 246, 101–118. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Su, J.; Li, Z.; Liu, B.; Cheng, G.; Jiang, Y.; Li, Y.; Zhou, S.; Yuan, W. Source apportionment of heavy metal and their health risks in soil-dustfall-plant system nearby a typical non-ferrous metal mining area of Tongling, Eastern China. Environ. Pollut. 2019, 254, 113089. [Google Scholar] [CrossRef] [PubMed]
- Tchounwou, P.B.; Yedjou, C.G.; Patlolla, A.K.; Sutton, D.J. Heavy metal toxicity and the environment. Mol. Clin. Environ. Toxicol. 2012, 101, 133–164. [Google Scholar] [CrossRef]
- Islam, M.M.; Karim, M.R.; Zheng, X.; Li, X. Heavy Metal and Metalloid Pollution of Soil, Water and Foods in Bangladesh: A Critical Review. Int. J. Environ. Res. Public Health 2018, 15, 2825. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Sun, X.; Xu, J. Heavy metal pollution in the East China Sea: A review. Mar. Pollut. Bull. 2020, 159, 111473. [Google Scholar] [CrossRef]
- Pedro, C.A.; Bruno, C.S.; Sarly, M.S.; Meireles, G.; Moutinho, A.; Novais, S.C.; Marques, J.C.; Goncalves, S.C. Are tolerance processes limiting the responses of Hediste diversicolor to cadmium exposure? A multimarker approach. Aquat. Toxicol. 2022, 252, 106300. [Google Scholar] [CrossRef]
- Li, X.; Zhou, J.; Zhou, T.; Li, Z.; Hu, P.; Luo, Y.; Christie, P.; Wu, L. Potential mobilization of cadmium and zinc in soils spiked with smithsonite and sphalerite under different water management regimes. J. Environ. Manag. 2022, 324, 116336. [Google Scholar] [CrossRef]
- Cui, S.; Wang, Z.; Li, X.; Wang, H.; Wang, H.; Chen, W. A comprehensive assessment of heavy metal(loid) contamination in leafy vegetables grown in two mining areas in Yunnan, China-a focus on bioaccumulation of cadmium in Malabar spinach. Environ. Sci. Pollut. Res. Int. 2022, 30, 14959–14974. [Google Scholar] [CrossRef]
- Haider, F.U.; Farooq, M.; Naveed, M.; Cheema, S.A.; Ain, N.U.; Salim, M.A.; Liqun, C.; Mustafa, A. Influence of biochar and microorganism co-application on stabilization of cadmium (Cd) and improved maize growth in Cd-contaminated soil. Front. Plant Sci. 2022, 13, 983830. [Google Scholar] [CrossRef]
- Haider, F.U.; Coulter, J.A.; Cheema, S.A.; Farooq, M.; Wu, J.; Zhang, R.; Shuaijie, G.; Liqun, C. Co-application of biochar and microorganisms improves soybean performance and remediate cadmium-contaminated soil. Ecotoxicol. Environ. Saf. 2021, 214, 112112. [Google Scholar] [CrossRef]
- Zhou, J.-Q.; Jiang, Y.-R.; Ming, X.-Q.; Wang, J.-R.; Tang, W.-B.; Sun, L. Introgressing the allelic variation of a major locus in reducing the grain cadmium accumulation in indica rice hybrids. Mol. Breed. 2019, 39, 84. [Google Scholar] [CrossRef]
- Liu, K.; Liu, H.; Zhou, X.; Chen, Z.; Wang, X. Cadmium Uptake and Translocation By Potato in Acid and Calcareous Soils. Bull. Environ. Contam. Toxicol. 2021, 107, 1149–1154. [Google Scholar] [CrossRef] [PubMed]
- Tian, W.; Huang, Y.; Li, D.; Meng, L.; He, T.; He, G. Identification of StAP2/ERF genes of potato (Solanum tuberosum) and their multiple functions in detoxification and accumulation of cadmium in yest: Implication for Genetic-based phytoremediation. Sci. Total Environ. 2022, 810, 152322. [Google Scholar] [CrossRef] [PubMed]
- Mawari, G.; Kumar, N.; Sarkar, S.; Daga, M.K.; Singh, M.M.; Joshi, T.K.; Khan, N.A. Heavy Metal Accumulation in Fruits and Vegetables and Human Health Risk Assessment: Findings From Maharashtra, India. Environ. Health Insights 2022, 16, 11786302221119151. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhao, Y.; Gu, L.; Wang, S.; Li, Y.; Dong, F. Accumulation and localization of cadmium in potato (Solanum tuberosum) under different soil Cd levels. Bull. Environ. Contam. Toxicol. 2014, 92, 745–751. [Google Scholar] [CrossRef]
- Ashrafzadeh, S.; Gaw, S.; Genet, R.; Glover, C.N.; Leung, D.W.M. Natural variation in correlations between cadmium and micronutrients in potato tubers. J. Food Compos. Anal. 2017, 59, 55–60. [Google Scholar] [CrossRef]
- Mengist, M.F.; Milbourne, D.; Griffin, D.; McLaughlin, M.J.; Creedon, J.; Jones, P.W.; Alves, S. Cadmium uptake and partitioning in potato (Solanum tuberosum L.) cultivars with different tuber-Cd concentration. Environ. Sci. Pollut. Res. Int. 2017, 24, 27384–27391. [Google Scholar] [CrossRef]
- Bao, Z.; Li, C.; Li, G.; Wang, P.; Peng, Z.; Cheng, L.; Li, H.; Zhang, Z.; Li, Y.; Huang, W.; et al. Genome architecture and tetrasomic inheritance of autotetraploid potato. Mol. Plant 2022, 15, 1211–1226. [Google Scholar] [CrossRef]
- Okabe, Y.; Ariizumi, T.; Ezura, H. Updating the Micro-Tom TILLING platform. Breed. Sci. 2013, 63, 42–48. [Google Scholar] [CrossRef]
- Kobayashi, M.; Nagasaki, H.; Garcia, V.; Just, D.; Bres, C.; Mauxion, J.P.; Le Paslier, M.C.; Brunel, D.; Suda, K.; Minakuchi, Y.; et al. Genome-wide analysis of intraspecific DNA polymorphism in ‘Micro-Tom’, a model cultivar of tomato (Solanum lycopersicum). Plant Cell Physiol. 2014, 55, 445–454. [Google Scholar] [CrossRef]
- Liu, Y.; Schiff, M.; Dinesh-Kumar, S.P. Virus-induced gene silencing in tomato. Plant J. 2002, 31, 777–786. [Google Scholar] [CrossRef]
- Hosseini Tafreshi, S.A.; Shariati, M.; Mofid, M.R.; Khayam Nekui, M.; Esmaeili, A. Heterologous virus-induced gene silencing as a promising approach in plant functional genomics. Mol. Biol. Rep. 2012, 39, 2169–2178. [Google Scholar] [CrossRef]
- Faivre-Rampant, O.; Gilroy, E.M.; Hrubikova, K.; Hein, I.; Millam, S.; Loake, G.J.; Birch, P.; Taylor, M.; Lacomme, C. Potato virus X-induced gene silencing in leaves and tubers of potato. Plant Physiol. 2004, 134, 1308–1316. [Google Scholar] [CrossRef]
- Tomar, M.; Singh, B.; Bhardwaj, V.; Sood, S.; Singh, B.; Salaria, N.; Thakur, K.; Kumar, A.; Sharma, N.; Goutam, U.; et al. Validation of molecular response of tuberization in response to elevated temperature by using a transient Virus Induced Gene Silencing (VIGS) in potato. Funct. Integr. Genom. 2021, 21, 215–229. [Google Scholar] [CrossRef]
- Chen, Y.; Li, G.; Yang, J.; Zhao, X.; Sun, Z.; Hou, H. Role of Nramp transporter genes of Spirodela polyrhiza in cadmium accumulation. Ecotoxicol. Environ. Saf. 2021, 227, 112907. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, M.; Song, H.; Zhao, J.; Shabala, S.; Tian, S.; Yang, X. A novel plasma membrane-based NRAMP transporter contributes to Cd and Zn hyperaccumulation in Sedum alfredii Hance. Environ. Exp. Bot. 2020, 176, 104121. [Google Scholar] [CrossRef]
- Yue, X.; Song, J.; Fang, B.; Wang, L.; Zou, J.; Su, N.; Cui, J. BcNRAMP1 promotes the absorption of cadmium and manganese in Arabidopsis. Chemosphere 2021, 283, 131113. [Google Scholar] [CrossRef]
- Bari, M.A.; El-Shehawi, A.M.; Elseehy, M.M.; Naheen, N.N.; Rahman, M.M.; Kabir, A.H. Molecular characterization and bioinformatics analysis of transporter genes associated with Cd-induced phytotoxicity in rice (Oryza sativa L.). Plant Physiol. Biochem. 2021, 167, 438–448. [Google Scholar] [CrossRef]
- Yokosho, K.; Yamaji, N.; Ma, J.F. Buckwheat FeNramp5 Mediates High Manganese Uptake in Roots. Plant Cell Physiol. 2021, 62, 600–609. [Google Scholar] [CrossRef]
- Zhang, W.; Yue, S.; Song, J.; Xun, M.; Han, M.; Yang, H. MhNRAMP1 from Malus hupehensis Exacerbates Cell Death by Accelerating Cd Uptake in Tobacco and Apple Calli. Front. Plant Sci. 2020, 11, 957. [Google Scholar] [CrossRef]
- Gao, H.; Xie, W.; Yang, C.; Xu, J.; Li, J.; Wang, H.; Chen, X.; Huang, C.F. NRAMP2, a trans-Golgi network-localized manganese transporter, is required for Arabidopsis root growth under manganese deficiency. New Phytol. 2018, 217, 179–193. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhao, D.; Xiao, S.; Zhang, A.; Deng, Y.; Dai, X.; Zhou, Z.; Ji, Z.; Cao, Q. Comparative Metabolomic and Transcriptomic Analyses of Phytochemicals in Two Elite Sweet Potato Cultivars for Table Use. Molecules 2022, 27, 8939. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Cai, Z.; Huang, J.; Wang, A.; Ntambiyukuri, A.; Chen, B.; Zheng, G.; Li, H.; Huang, Y.; Zhan, J.; et al. Transcriptomic analysis of tuberous root in two sweet potato varieties reveals the important genes and regulatory pathways in tuberous root development. BMC Genom. 2022, 23, 473. [Google Scholar] [CrossRef] [PubMed]
- Petek, M.; Zagorscak, M.; Ramsak, Z.; Sanders, S.; Tomaz, S.; Tseng, E.; Zouine, M.; Coll, A.; Gruden, K. Cultivar-specific transcriptome and pan-transcriptome reconstruction of tetraploid potato. Sci. Data 2020, 7, 249. [Google Scholar] [CrossRef]
- Enzingmuller-Bleyl, T.C.; Boden, J.S.; Herrmann, A.J.; Ebel, K.W.; Sanchez-Baracaldo, P.; Frankenberg-Dinkel, N.; Gehringer, M.M. On the trail of iron uptake in ancestral Cyanobacteria on early Earth. Geobiology 2022, 20, 776–789. [Google Scholar] [CrossRef]
- Zou, X.; Huang, R.; Wang, L.; Wang, G.; Miao, Y.; Rao, I.; Liu, G.; Chen, Z. SgNramp1, a plasma membrane-localized transporter, involves in manganese uptake in Stylosanthes guianensis. Front. Plant Sci. 2022, 13, 1027551. [Google Scholar] [CrossRef]
- Wu, D.; Yamaji, N.; Yamane, M.; Kashino-Fujii, M.; Sato, K.; Feng Ma, J. The HvNramp5 Transporter Mediates Uptake of Cadmium and Manganese, But Not Iron. Plant Physiol. 2016, 172, 1899–1910. [Google Scholar] [CrossRef]
- Hussain, B.; Li, J.; Ma, Y.; Tahir, N.; Ullah, A. Effects of Fe and Mn cations on Cd uptake by rice plant in hydroponic culture experiment. PLoS ONE 2020, 15, e0243174. [Google Scholar] [CrossRef]
- Tang, L.; Dong, J.; Qu, M.; Lv, Q.; Zhang, L.; Peng, C.; Hu, Y.; Li, Y.; Ji, Z.; Mao, B.; et al. Knockout of OsNRAMP5 enhances rice tolerance to cadmium toxicity in response to varying external cadmium concentrations via distinct mechanisms. Sci. Total Environ. 2022, 832, 155006. [Google Scholar] [CrossRef]
- Robas Mora, M.; Fernandez Pastrana, V.M.; Gonzalez Reguero, D.; Gutierrez Oliva, L.L.; Probanza Lobo, A.; Jimenez Gomez, P.A. Oxidative stress protection and growth promotion activity of Pseudomonas mercuritolerans sp. nov., in forage plants under mercury abiotic stress conditions. Front. Microbiol. 2022, 13, 1032901. [Google Scholar] [CrossRef]
- Elsalahy, H.H.; Bellingrath-Kimura, S.D.; Ross, C.L.; Kautz, T.; Doring, T.F. Crop Resilience to Drought with and without Response Diversity. Front. Plant Sci. 2020, 11, 721. [Google Scholar] [CrossRef]
- Zhang, F.Q.; Wang, Y.S.; Lou, Z.P.; Dong, J.D. Effect of heavy metal stress on antioxidative enzymes and lipid peroxidation in leaves and roots of two mangrove plant seedlings (Kandelia candel and Bruguiera gymnorrhiza). Chemosphere 2007, 67, 44–50. [Google Scholar] [CrossRef]
- Laskos, K.; Myskow, B.; Dziurka, M.; Warchol, M.; Dziurka, K.; Juzon, K.; Czyczylo-Mysza, I.M. Variation between glaucous and non-glaucous near-isogenic lines of rye (Secale cereale L.) under drought stress. Sci. Rep. 2022, 12, 22486. [Google Scholar] [CrossRef]
- Fang, S.; Yang, H.; Wei, G.; Shen, T.; Wan, Z.; Wang, M.; Wang, X.; Wu, Z. Potassium application enhances drought tolerance in sesame by mitigating oxidative damage and regulating osmotic adjustment. Front. Plant Sci. 2022, 13, 1096606. [Google Scholar] [CrossRef]
- Cui, B.; Liu, C.; Hu, C.; Liang, S. Transcriptomic Sequencing Analysis on Key Genes and Pathways Regulating Cadmium (Cd) in Ryegrass (Lolium perenne L.) under Different Cadmium Concentrations. Toxics 2022, 10, 734. [Google Scholar] [CrossRef]
- Xu, T.; Xi, J.; Ke, J.; Wang, Y.; Chen, X.; Zhang, Z.; Lin, Y. Deciphering soil amendments and actinomycetes for remediation of cadmium (Cd) contaminated farmland. Ecotoxicol. Environ. Saf. 2022, 249, 114388. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2010, 26, 136–138. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Klopfenstein, D.V.; Zhang, L.; Pedersen, B.S.; Ramirez, F.; Warwick Vesztrocy, A.; Naldi, A.; Mungall, C.J.; Yunes, J.M.; Botvinnik, O.; Weigel, M.; et al. GOATOOLS: A Python library for Gene Ontology analyses. Sci. Rep. 2018, 8, 10872. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; He, T.; Tian, W.; Xia, Y.; He, Y.; Su, M.; He, G. The Expression of the StNRAMP2 Gene Determined the Accumulation of Cadmium in Different Tissues of Potato. Int. J. Mol. Sci. 2023, 24, 9322. https://doi.org/10.3390/ijms24119322
Zhang Y, He T, Tian W, Xia Y, He Y, Su M, He G. The Expression of the StNRAMP2 Gene Determined the Accumulation of Cadmium in Different Tissues of Potato. International Journal of Molecular Sciences. 2023; 24(11):9322. https://doi.org/10.3390/ijms24119322
Chicago/Turabian StyleZhang, Yule, Tengbing He, Weijun Tian, Yabei Xia, Yeqing He, Minmin Su, and Guandi He. 2023. "The Expression of the StNRAMP2 Gene Determined the Accumulation of Cadmium in Different Tissues of Potato" International Journal of Molecular Sciences 24, no. 11: 9322. https://doi.org/10.3390/ijms24119322