
Friunavirus Phage-Encoded Depolymerases Specific to Different Capsular Types of Acinetobacter baumannii
 , , ,
, , ,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
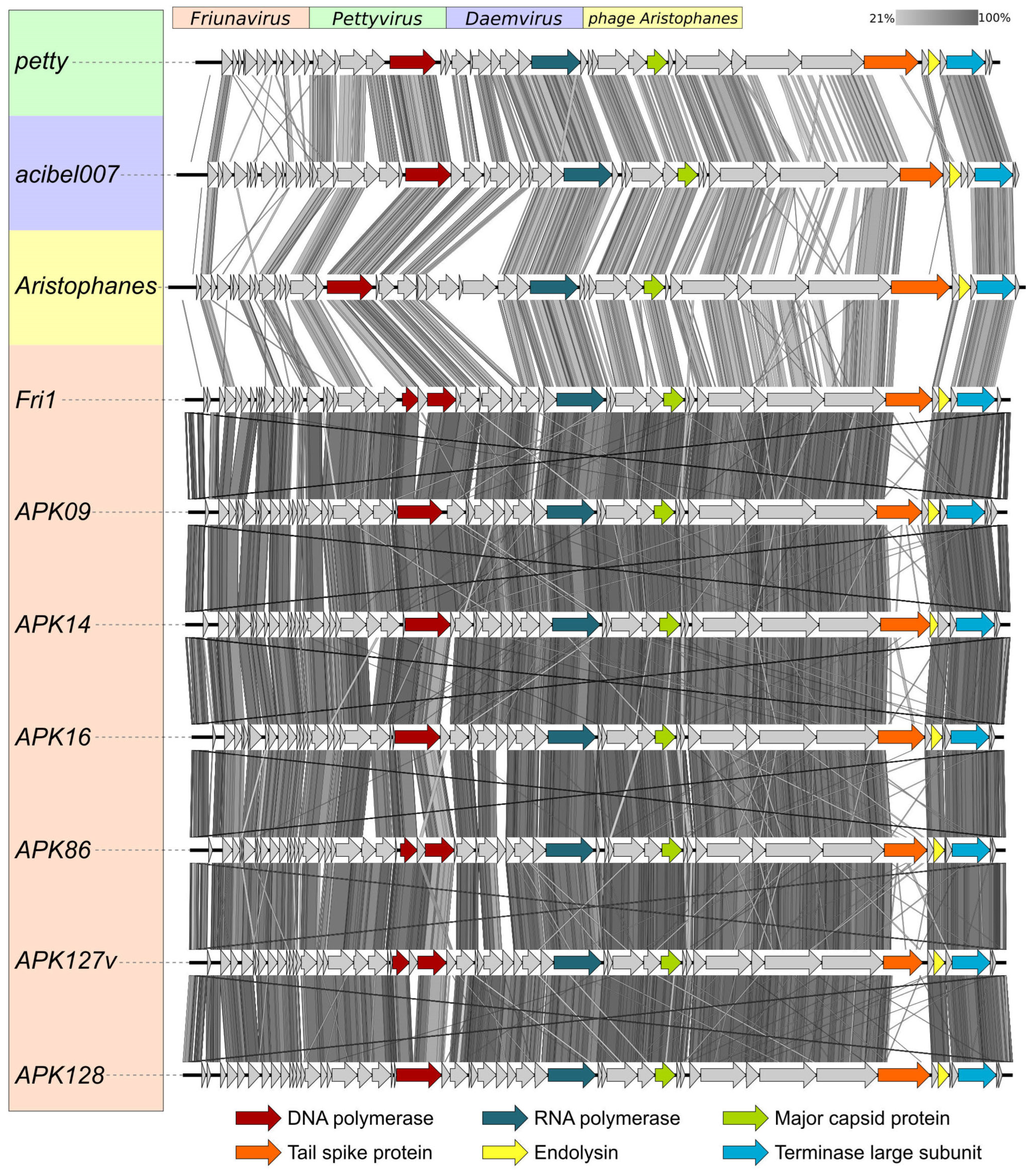
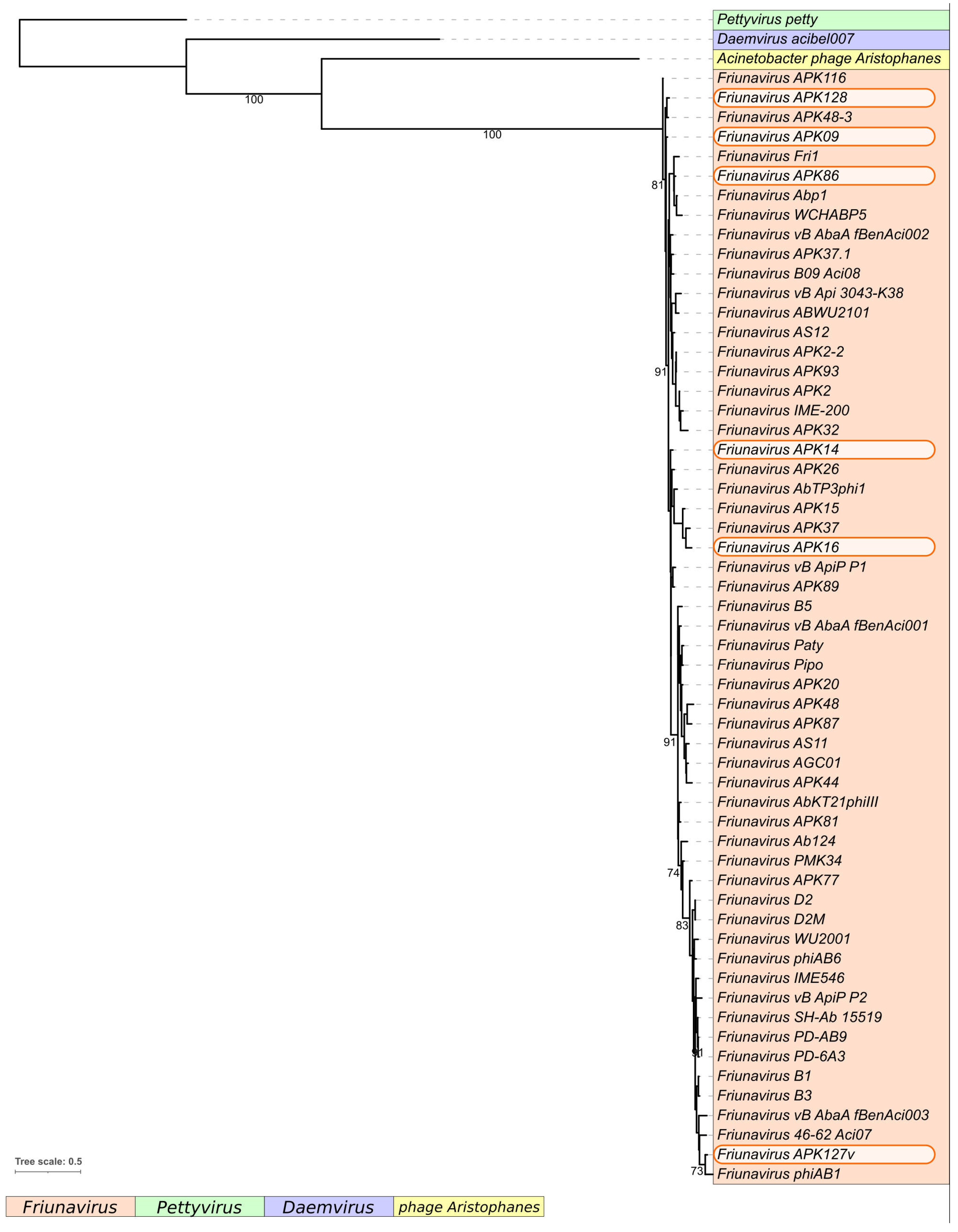
2.1. Characterization of Phages Encoding the Studied Depolymerases
2.2. Phage Tailspike Depolymerases
2.3. Mechanism of Cleavage of A. baumannii CPSs by Specific Phage TSDs
2.3.1. Cleavage of the A. baumannii B09 CPS by Recombinant TSD APK09_gp48

2.3.2. Cleavage of the A. baumannii AB5256 CPS by Recombinant TSD APK14_gp49

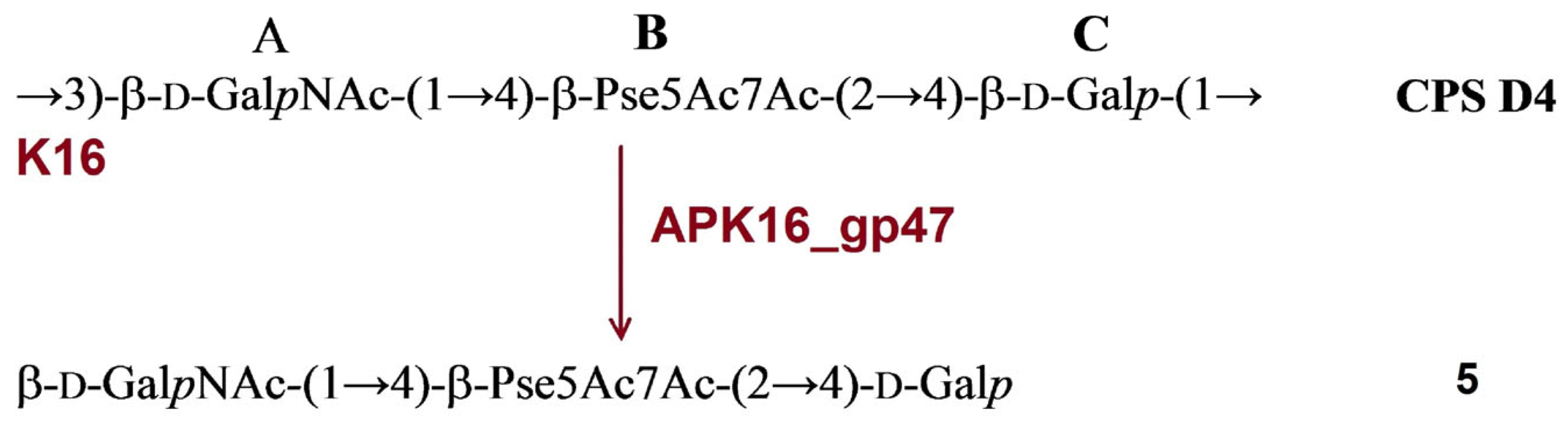
2.3.3. Cleavage of the A. baumannii D4 CPS by Recombinant TSD APK16_gp47

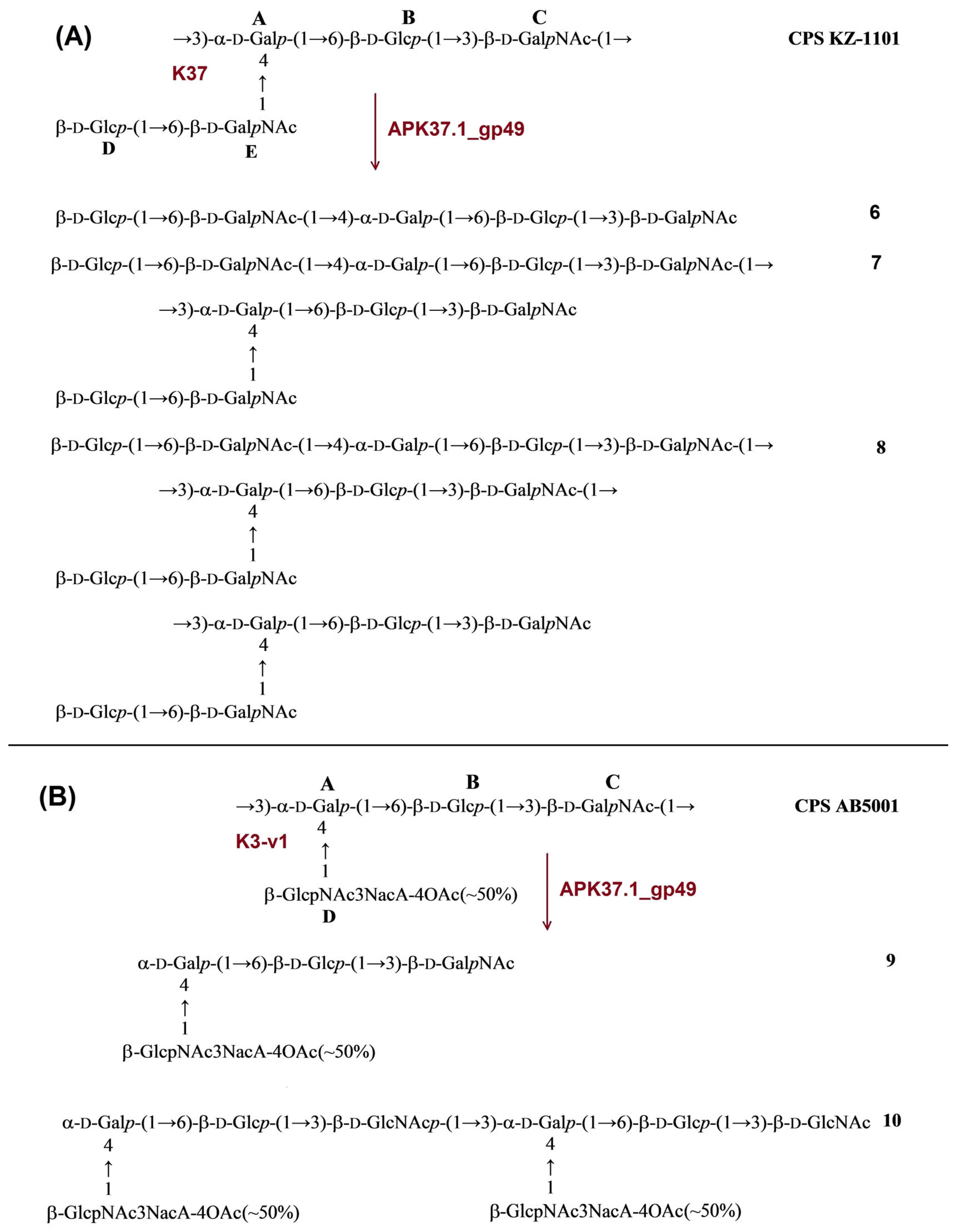
2.3.4. Cleavage of the A. baumannii KZ-1101 and AB5001 CPSs by Recombinant TSD APK37.1_gp49

2.3.5. Cleavage of the A. baumannii MAR55-66 CPS by Recombinant TSD APK86_gp49

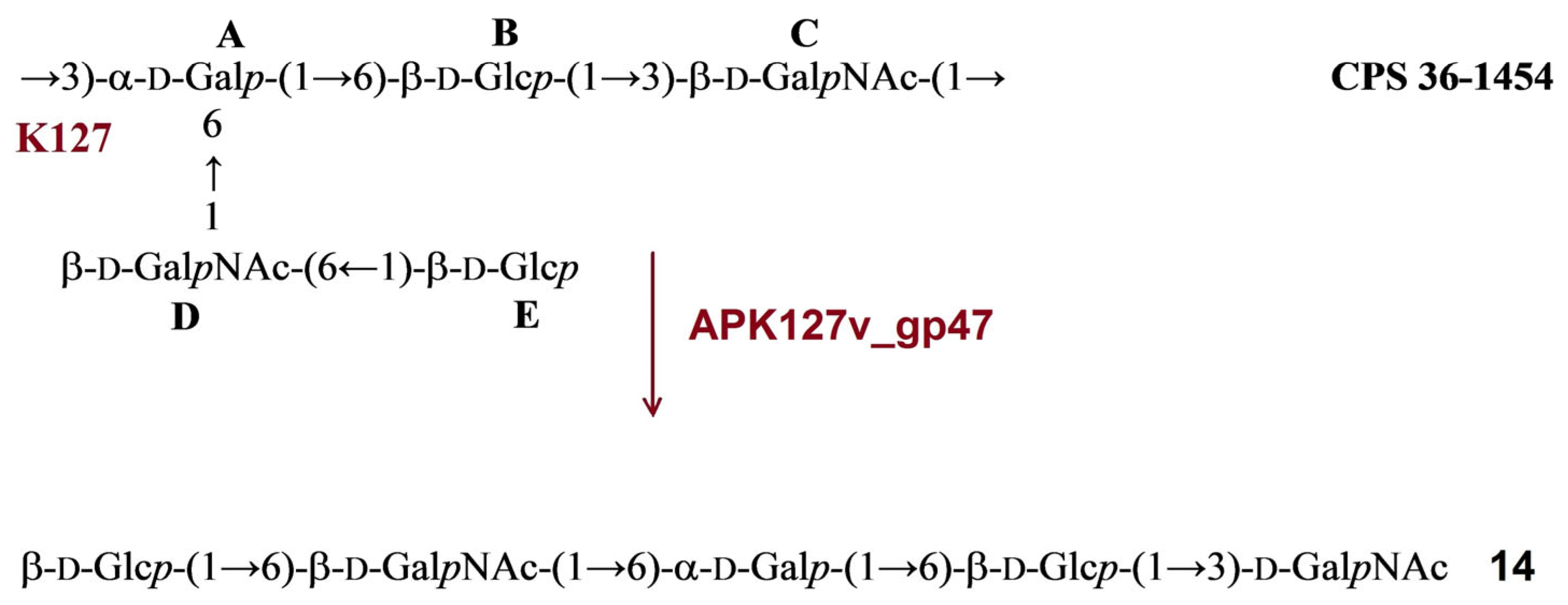
2.3.6. Cleavage of the A. baumannii 36-1454 CPS by Recombinant TSD APK127v_gp47

2.3.7. Cleavage of the A. baumannii KZ-1093 CPS by Recombinant TSD APK128_gp45

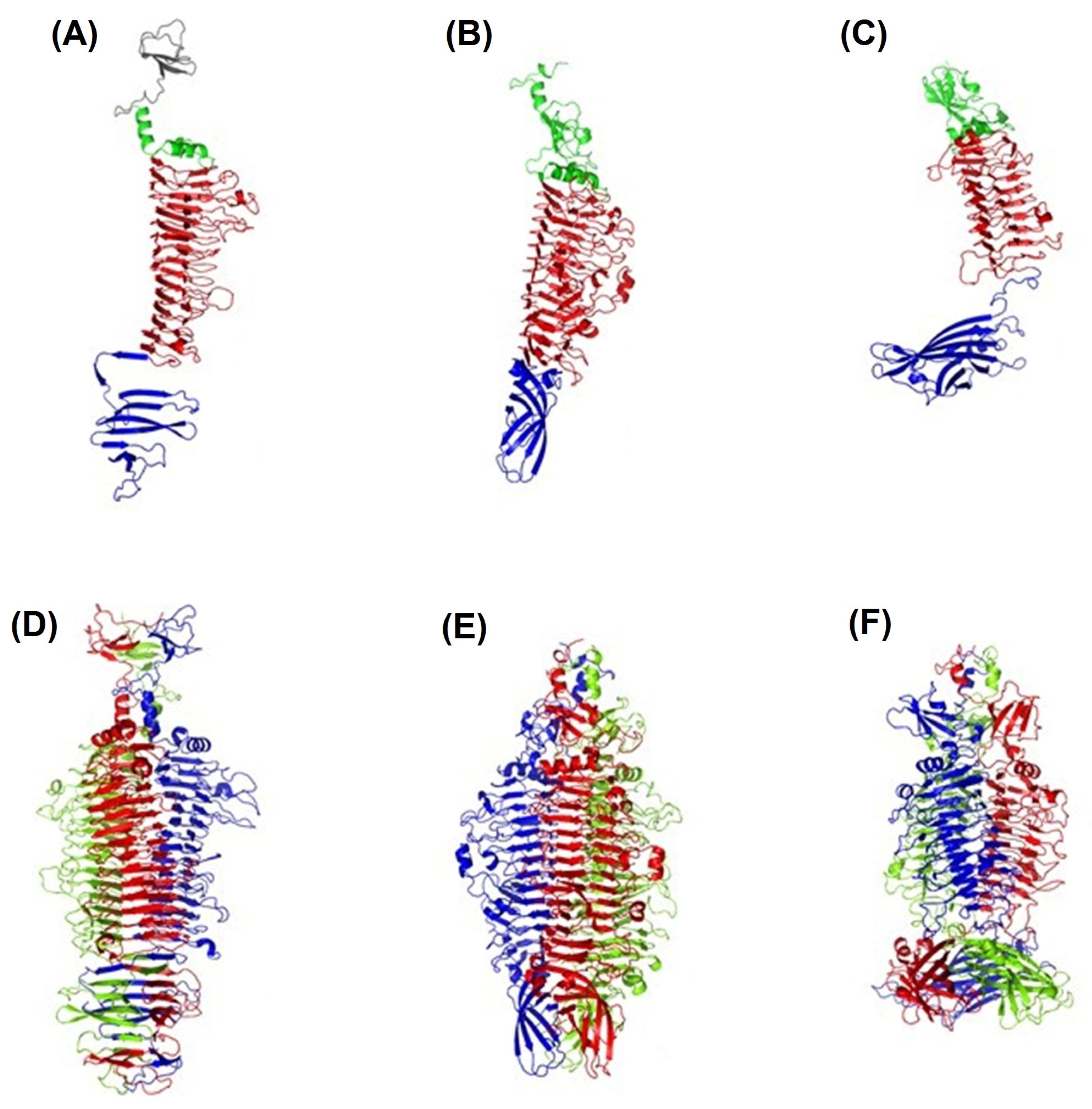
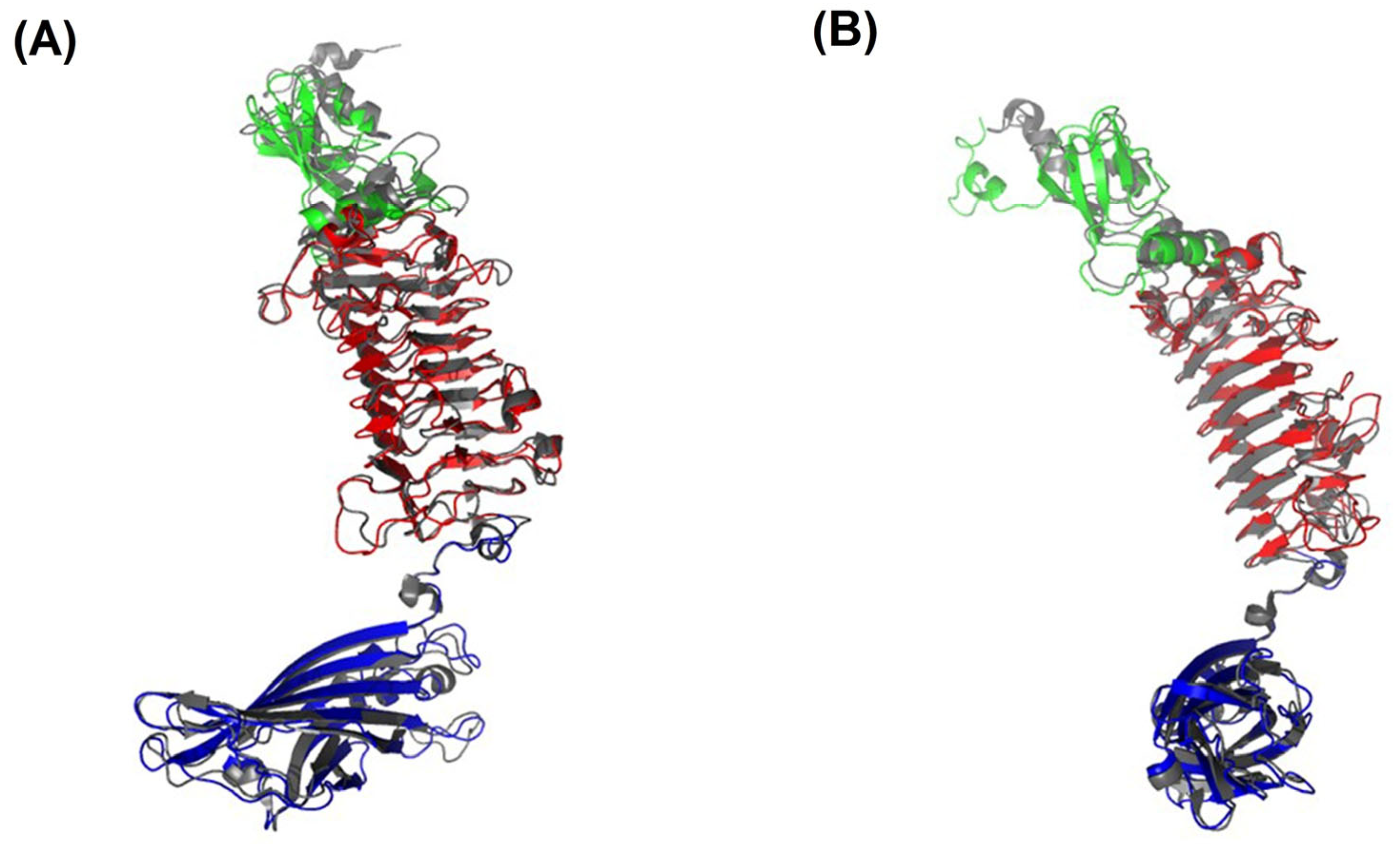
2.4. The Structure of the TSDs
2.4.1. Crystal Structure of TSD APK16_gp47
2.4.2. Crystal Structure of TSD APK14_gp49
2.4.3. Crystal Structure of TSD APK09_gp48
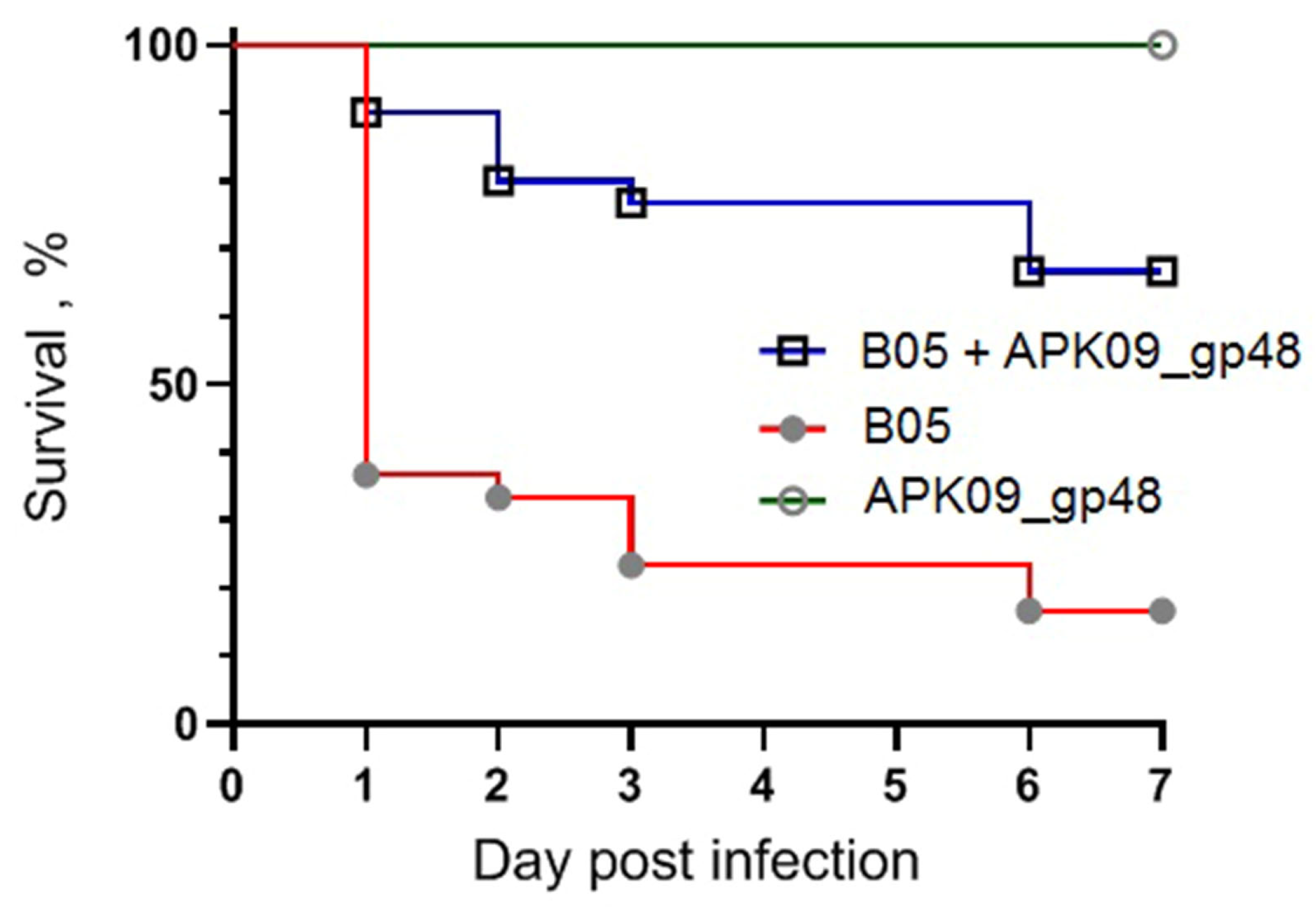
2.5. Evaluation of the Antivirulence Potential of TSD APK09_gp48 in a G. mellonella larvae Model of A. baumannii Infection
3. Discussion
4. Materials and Methods
4.1. Phage Isolation, Propagation, and Purification
4.2. Phage DNA Isolation and Sequencing
4.3. Phage Genome Analysis
4.4. Nucleotide Sequences Accession Numbers
4.5. Cloning, Expression, and Purification of the Recombinant Depolymerases
4.6. Lawn Spot Assay
4.7. Isolation, Purification, and Depolymerization of the CPSs by Recombinant Proteins
4.8. NMR Spectroscopy
4.9. Crystallization, Data Collection, Processing, Structure Solution, and Refinement for the Recombinant TSDs
4.10. Galleria mellonella Larvae Infection Experiments
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organisation. Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery and Development of New Antibiotics; WHO Press: Geneva, Switzerland, 2017; pp. 7–526. Available online: http://remed.org/wp-content/uploads/2017/03/lobal-priority-list-of-antibiotic-resistant-bacteria-2017.pdf (accessed on 2 April 2023).
- Oliveira, H.; Costa, A.R.; Konstantinides, N.; Ferreira, A.; Akturk, E.; Sillankorva, S.; Nemec, A.; Shneider, M.; Dötsch, A.; Azeredo, J. Ability of phages to infect Acinetobacter calcoaceticus-Acinetobacter baumannii complex species through acquisition of different pectate lyase depolymerase domains. Environ. Microbiol. 2017, 19, 5060–5077. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.-M.; Tu, I.-F.; Yang, F.-L.; Ko, T.-P.; Liao, J.-H.; Lin, N.-T.; Wu, C.-Y.; Ren, C.-T.; Wang, A.H.-J.; Chang, C.-M.; et al. Structural basis for fragmenting the exopolysaccharide of Acinetobacter baumannii by bacteriophage ΦAB6 tailspike protein. Sci. Rep. 2017, 7, 42711. [Google Scholar] [CrossRef] [PubMed]
- Popova, A.V.; Shneider, M.M.; Arbatsky, N.P.; Kasimova, A.A.; Senchenkova, S.N.; Shashkov, A.S.; Dmitrenok, A.S.; Chizhov, A.O.; Mikhailova, Y.V.; Shagin, D.A.; et al. Specific Interaction of Novel Friunavirus Phages Encoding Tailspike Depolymerases with Corresponding Acinetobacter baumannii Capsular Types. J. Virol. 2021, 95, e01714-20. [Google Scholar] [CrossRef] [PubMed]
- Kasimova, A.A.; Arbatsky, N.P.; Timoshina, O.Y.; Shneider, M.M.; Shashkov, A.S.; Chizhov, A.O.; Popova, A.V.; Hall, R.M.; Kenyon, J.J.; Knirel, Y.A. The K26 capsular polysaccharide from Acinetobacter baumannii KZ-1098: Structure and cleavage by a specific phage depolymerase. Int. J. Biol. Macromol. 2021, 191, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Arbatsky, N.P.; Kasimova, A.A.; Shashkov, A.S.; Shneider, M.M.; Popova, A.V.; Shagin, D.A.; Shelenkov, A.A.; Mikhailova, Y.V.; Yanushevich, Y.G.; Hall, R.M.; et al. Involvement of a Phage-Encoded Wzy Protein in the Polymerization of K127 Units to Form the Capsular Polysaccharide of Acinetobacter baumannii Isolate 36-1454. Microbiol. Spectr. 2022, 10, e0150321. [Google Scholar] [CrossRef] [PubMed]
- Wyres, K.L.; Cahill, S.M.; Holt, K.E.; Hall, R.M.; Kenyon, J.J. Identification of Acinetobacter baumannii loci for capsular polysaccharide (KL) and lipooligosaccharide outer core (OCL) synthesis in genome assemblies using curated reference databases compatible with Kaptive. Microb. Genom. 2020, 6, e000339. [Google Scholar] [CrossRef]
- Kenyon, J.J.; Hall, R.M. Updated Analysis of the Surface Carbohydrate Gene Clusters in a Diverse Panel of Acinetobacter baumannii Isolates. Antimicrob. Agents Chemother. 2022, 66, e0180721. [Google Scholar] [CrossRef]
- Cahill, S.M.; Hall, R.M.; Kenyon, J.J. An update to the database for Acinetobacter baumannii capsular polysaccharide locus typing extends the extensive and diverse repertoire of genes found at and outside the K locus. Microb. Genom. 2022, 8, mgen000878. [Google Scholar] [CrossRef]
- Oliveira, H.; Domingues, R.; Evans, B.; Sutton, J.M.; Adriaenssens, E.M.; Turner, D. Genomic Diversity of Bacteriophages Infecting the Genus Acinetobacter. Viruses 2022, 14, 181. [Google Scholar] [CrossRef]
- Timoshina, O.Y.; Kasimova, A.A.; Shneider, M.M.; Arbatsky, N.P.; Shashkov, A.S.; Shelenkov, A.A.; Mikhailova, Y.V.; Popova, A.V.; Hall, R.M.; Knirel, Y.A.; et al. Loss of a Branch Sugar in the Acinetobacter baumannii K3-Type Capsular Polysaccharide Due to Frameshifts in the gtr6 Glycosyltransferase Gene Leads to Susceptibility To Phage APK37.1. Microbiol. Spectr. 2023, 11, e0363122. [Google Scholar] [CrossRef]
- Popova, A.; Lavysh, D.; Klimuk, E.; Edelstein, M.; Bogun, A.; Shneider, M.; Goncharov, A.; Leonov, S.; Severinov, K. Novel Fri1-like Viruses Infecting Acinetobacter baumannii—vB_AbaP_AS11 and vB_AbaP_AS12—Characterization, Comparative Genomic Analysis, and Host-Recognition Strategy. Viruses 2017, 9, 188. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Hernandez-Morales, A.; Clark, J.; Le, T.; Biswas, B.; Bishop-Lilly, K.A.; Henry, M.; Quinones, J.; Voegtly, L.J.; Cer, R.Z.; et al. Comparative genomics of Acinetobacter baumannii and therapeutic bacteriophages from a patient undergoing phage therapy. Nat. Commun. 2022, 13, 3776. [Google Scholar] [CrossRef] [PubMed]
- Popova, A.V.; Shneider, M.M.; Myakinina, V.P.; Bannov, V.A.; Edelstein, M.V.; Rubalskii, E.O.; Aleshkin, A.V.; Fursova, N.K.; Volozhantsev, N.V. Characterization of myophage AM24 infecting Acinetobacter baumannii of the K9 capsular type. Arch. Virol. 2019, 164, 1493–1497. [Google Scholar] [CrossRef] [PubMed]
- Shchurova, A.S.; Shneider, M.M.; Arbatsky, N.P.; Shashkov, A.S.; Chizhov, A.O.; Skryabin, Y.P.; Mikhaylova, Y.V.; Sokolova, O.S.; Shelenkov, A.A.; Miroshnikov, K.A.; et al. Novel Acinetobacter baumannii Myovirus TaPaz Encoding Two Tailspike Depolymerases: Characterization and Host-Recognition Strategy. Viruses 2021, 13, 978. [Google Scholar] [CrossRef] [PubMed]
- Timoshina, O.Y.; Shneider, M.M.; Evseev, P.V.; Shchurova, A.S.; Shelenkov, A.A.; Mikhaylova, Y.V.; Sokolova, O.S.; Kasimova, A.A.; Arbatsky, N.P.; Dmitrenok, A.S.; et al. Novel Acinetobacter baumannii Bacteriophage Aristophanes Encoding Structural Polysaccharide Deacetylase. Viruses 2021, 13, 1688. [Google Scholar] [CrossRef] [PubMed]
- Kenyon, J.J.; Arbatsky, N.P.; Sweeney, E.L.; Shashkov, A.S.; Shneider, M.M.; Popova, A.V.; Hall, R.M.; Knirel, Y.A. Production of the K16 capsular polysaccharide by Acinetobacter baumannii ST25 isolate D4 involves a novel glycosyltransferase encoded in the KL16 gene cluster. Int. J. Biol. Macromol. 2019, 128, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Arbatsky, N.P.; Shashkov, A.S.; Chizhov, A.O.; Timoshina, O.Y.; Shneider, M.M.; Knirel, Y. A Structure of the capsular polysaccharide of Acinetobacter baumannii MAR 55–66. Russ. Chem. Bull. 2021, 70, 592–599. [Google Scholar] [CrossRef]
- Kenyon, J.J.; Kasimova, A.A.; Sviridova, A.N.; Shpirt, A.M.; Shneider, M.M.; Mikhaylova, Y.V.; Shelenkov, A.A.; Popova, A.V.; Perepelov, A.V.; Shashkov, A.S.; et al. Correlation of Acinetobacter baumannii K144 and K86 capsular polysaccharide structures with genes at the K locus reveals the involvement of a novel multifunctional rhamnosyltransferase for structural synthesis. Int. J. Biol. Macromol. 2021, 193, 1294–1300. [Google Scholar] [CrossRef]
- Arbatsky, N.P.; Kasimova, A.A.; Shashkov, A.S.; Shneider, M.M.; Popova, A.V.; Shagin, D.A.; Shelenkov, A.A.; Mikhailova, Y.V.; Yanushevich, Y.G.; Azizov, I.S.; et al. Structure of the K128 capsular polysaccharide produced by Acinetobacter baumannii KZ-1093 from Kazakhstan. Carbohydr Res. 2019, 485, 107814. [Google Scholar] [CrossRef]
- Pires, D.P.; Oliveira, H.; Melo, L.; Sillankorva, S.; Azeredo, J. Bacteriophage-encoded depolymerases: Their diversity and biotechnological applications. Appl. Microbiol. Biotechnol. 2016, 100, 2141–2151. [Google Scholar] [CrossRef]
- Farrugia, D.N.; Elbourne, L.D.H.; Hassan, K.A.; Eijkelkamp, B.A.; Tetu, S.G.; Brown, M.H.; Shah, B.S.; Peleg, A.Y.; Mabbutt, B.C.; Paulsen, I.T. The Complete Genome and Phenome of a Community-Acquired Acinetobacter baumannii. PLoS ONE 2013, 8, e58628. [Google Scholar] [CrossRef] [PubMed]
- Roucourt, B.; Lavigne, R. The role of interactions between phage and bacterial proteins within the infected cell: A diverse and puzzling interactome. Environ. Microbiol. 2009, 11, 2789–2805. [Google Scholar] [CrossRef] [PubMed]
- Latka, A.; Maciejewska, B.; Majkowska-Skrobek, G.; Briers, Y.; Drulis-Kawa, Z. Bacteriophage-encoded virion-associated enzymes to overcome the carbohydrate barriers during the infection process. Appl. Microbiol. Biotechnol. 2017, 101, 3103–3119. [Google Scholar] [CrossRef] [PubMed]
- Popova, A.V.; Shneider, M.M.; Mikhailova, Y.V.; Shelenkov, A.A.; Shagin, D.A.; Edelstein, M.V.; Kozlov, R.S. Complete Genome Sequence of Acinetobacter baumannii Phage BS46. Microbiol. Resour. Announc. 2020, 9, e00398-20. [Google Scholar] [CrossRef]
- Knirel, Y.A.; Shneider, M.M.; Popova, A.V.; Kasimova, A.A.; Senchenkova, S.N.; Shashkov, A.S.; Chizhov, A.O. Mechanisms of Acinetobacter baumannii Capsular Polysaccharide Cleavage by Phage Depolymerases. Biochemistry 2020, 85, 567–574. [Google Scholar] [CrossRef]
- Vinogradov, E.V.; Pantophlet, R.; Dijkshoorn, L.; Brade, L.; Holst, O.; Brade, H. Structural and serological characterisation of two O-specific polysaccharides of Acinetobacter. Eur. J. Biochem. 1996, 239, 602–610. [Google Scholar] [CrossRef]
- Kenyon, J.J.; Hall, R.M. Variation in the Complex Carbohydrate Biosynthesis Loci of Acinetobacter baumannii Genomes. PLoS ONE 2013, 8, e62160. [Google Scholar] [CrossRef]
- Haseley, S.R.; Wilkinson, S.G. Structural studies of the putative O-specific polysaccharide of Acinetobacter baumannii O11. Eur. J. Biochem. 1996, 237, 266–271. [Google Scholar] [CrossRef]
- Kenyon, J.J.; Hall, R.M.; De Castro, C. Structural determination of the K14 capsular polysaccharide from an ST25 Acinetobacter baumannii isolate, D46. Carbohydr. Res. 2015, 417, 52–56. [Google Scholar] [CrossRef]
- Shashkov, A.S.; Cahill, S.M.; Arbatsky, N.P.; Westacott, A.C.; Kasimova, A.A.; Shneider, M.M.; Popova, A.V.; Shagin, D.A.; Shelenkov, A.A.; Mikhailova, Y.V.; et al. Acinetobacter baumannii K116 capsular polysaccharide structure is a hybrid of the K14 and revised K37 structures. Carbohydr. Res. 2019, 484, 107774. [Google Scholar] [CrossRef]
- Oliveira, H.; Mendes, A.; Fraga, A.G.; Ferreira, A.; Pimenta, A.I.; Mil-Homens, D.; Fialho, A.M.; Pedrosa, J.; Azeredo, J. K2 Capsule Depolymerase Is Highly Stable, Is Refractory to Resistance, and Protects Larvae and Mice from Acinetobacter baumannii Sepsis. Appl. Environ. Microbiol. 2019, 85, e00934-19. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Leung, S.S.Y.; Guo, Y.; Zhao, L.; Jiang, N.; Mi, L.; Li, P.; Wang, C.; Qin, Y.; Mi, Z.; et al. The Capsule Depolymerase Dpo48 Rescues Galleria mellonella and Mice from Acinetobacter baumannii Systemic Infections. Front. Microbiol. 2019, 10, 545. [Google Scholar] [CrossRef] [PubMed]
- Drobiazko, A.Y.; Kasimova, A.A.; Evseev, P.V.; Shneider, M.M.; Klimuk, E.I.; Shashkov, A.S.; Dmitrenok, A.S.; Chizhov, A.O.; Slukin, P.V.; Skryabin, Y.P.; et al. Capsule-Targeting Depolymerases Derived from Acinetobacter baumannii Prophage Regions. Int. J. Mol. Sci. 2022, 23, 4971. [Google Scholar] [CrossRef]
- Adams, M.D. Bacteriophages; Interscience Publishers, Inc.: New York, NY, USA, 1959. [Google Scholar]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989; ISBN 0-87969-309-6. [Google Scholar]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST server: Rapid annotations using subsystems technology. BMC Genomics 2008, 9, 550. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Anderson, J.B.; Chitsaz, F.; Derbyshire, M.K.; DeWeese-Scott, C.; Fong, J.H.; Geer, L.Y.; Geer, R.C.; Gozales, N.R.; Gwadz, M.; et al. CDD: Specific functional annotation with the Conserved Domain Database. Nucleic Acids Res. 2009, 37, D205–D210. [Google Scholar] [CrossRef] [PubMed]
- Söding, J.; Biegert, A.; Lupas, A.N. The HHpred interactive server for protein homology detection and structure prediction. Nucleic Acids Res. 2005, 33, W244–W248. [Google Scholar] [CrossRef]
- Schattner, P.; Brooks, A.N.; Lowe, T.M. The TRNAscan-SE, Snoscan and SnoGPS Web Servers for the Detection of TRNAs and SnoRNAs. Nucleic Acids Res. 2005, 33, W686–W689. [Google Scholar] [CrossRef]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A Genome Comparison Visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef]
- Moraru, C.; Varsani, A.; Kropinski, A.M. VIRIDIC—A Novel Tool to Calculate the Intergenomic Similarities of Prokaryote-Infecting Viruses. Viruses 2020, 12, 1268. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.I.; Miyata, T. MAFFT: A Novel Method for Rapid Multiple Sequence Alignment Based on Fast Fourier Transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Kozlov, A.M.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML-NG: A Fast, Scalable and User-Friendly Tool for Maximum Likelihood Phylogenetic Inference. Bioinformatics 2019, 35, 4453–4455. [Google Scholar] [CrossRef] [PubMed]
- Taylor, N.M.I.; Prokhorov, N.S.; Guerrero-Ferreira, R.C.; Shneider, M.M.; Browning, C.; Goldie, K.N.; Stahlberg, H.; Leiman, P.G. Structure of the T4 Baseplate and Its Function in Triggering Sheath Contraction. Nature 2016, 533, 346–352. [Google Scholar] [CrossRef]
- Westphal, O.; Jann, K. Bacterial lipopolysaccharides. Extraction with phenol-water and further applications of the procedure. Methods Carbohydr. Chem. 1965, 5, 83–91. [Google Scholar]
- Winter, G.; Waterman, D.G.; Parkhurst, J.M.; Brewster, A.S.; Gildea, R.J.; Gerstel, M.; Fuentes-Montero, L.; Vollmar, M.; Michels-Clark, T.; Young, I.D.; et al. DIALS: Implementation and evaluation of a new integration package. Acta Crystallogr. D Struct. Biol. 2018, 74, 85–97. [Google Scholar] [CrossRef]
- Kabsch, W. XDS. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 125–132. [Google Scholar] [CrossRef]
- Evans, P. Scaling and assessment of data quality. Acta Crystallogr. D Biol. Crystallogr. 2006, 62, 72–82. [Google Scholar] [CrossRef]
- Skubák, P.; Pannu, N.S. Automatic protein structure solution from weak X-ray data. Nat. Commun. 2013, 4, 2777. [Google Scholar] [CrossRef]
- Vagin, A.; Teplyakov, A. MOLREP: An Automated Program for Molecular Replacement. J. Appl. Cryst. 1997, 30, 1022–1025. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Collaborative Computational Project, Number 4 The CCP4 suite: Programs for protein crystallography. Acta Crystallogr. D Biol. Crystallogr. 1994, 50, 760–763. [CrossRef] [PubMed]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Crystallogr. D Biol. Crystallogr. 2004, 60, 2126–2132. [Google Scholar] [CrossRef]
- Krissinel, E.; Henrick, K. Secondary-structure matching (SSM), a new tool for fast protein structure alignment in three dimensions. Acta Crystallogr. D Biol. Crystallogr. 2004, 60, 2256–2268. [Google Scholar] [CrossRef] [PubMed]
- Krissinel, E.; Henrick, K.J. Inference of macromolecular assemblies from crystalline state. Mol. Biol. 2007, 372, 774–797. [Google Scholar] [CrossRef] [PubMed]
- Gorodnichev, R.B.; Volozhantsev, N.V.; Krasilnikova, V.M.; Bodoev, I.N.; Kornienko, M.A.; Kuptsov, N.S.; Popova, A.V.; Makarenko, G.I.; Manolov, A.I.; Slukin, P.V.; et al. Novel Klebsiella pneumoniae K23-Specific Bacteriophages from Different Families: Similarity of Depolymerases and Their Therapeutic Potential. Front. Microbiol. 2021, 12, 669618. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phage Name | Genome Length (bp) | DTR Length (bp) | G+C Content (%) | Total No of Genes | Genbank Accession Number | TSD Designation (ID) | Reference |
|---|---|---|---|---|---|---|---|
| APK09 | 41477 | 409 | 39.2 | 56 | MZ868724 | APK09_gp48 (UAW09804) | this work |
| APK14 | 41767 | 405 | 39.2 | 55 | MK089780 | APK14_gp49 (AYR04394) | this work |
| APK16 | 41135 | 357 | 39.4 | 54 | MZ868725 | APK16_gp47 (UAW09859) | this work |
| APK37.1 | 40966 | 339 | 39.2 | 56 | MZ967493 | APK37.1_gp49 (UAW07728) | [11] |
| APK86 | 41297 | 383 | 39.2 | 56 | MZ936314 | APK86_gp49 (UAW09972) | this work |
| APK127v | 41380 | 422 | 39.2 | 53 | ON210142 | APK127v_gp47 (URQ05189) | this work |
| APK128 | 42013 | 428 | 39.2 | 52 | MW459163 | APK128_gp45 (QVD48888) | this work |
| Phage | TSDs | A. baumannii Host Strain | K Type | Linkage in CPS Cleaved by a Depolymerase | Depolymerization Products | |||
|---|---|---|---|---|---|---|---|---|
| Mono- mer | Di- mer | Tri- mer | Struc- tures | |||||
| APK09 | APK09_gp48 | B05 | K9 | β-d-GlcpNAc-(1→3)-α-d-GalpNAcA | 1 | 2 | Figure 4 | |
| APK14 | APK14_gp49 | AB5256 | K14 | α-d-GalpNAc-(1→4)-β-d-GalpNAc | 3 | 4 | Figure 5 | |
| APK16 | APK16_gp47 | D4 | K16 | β-d-Galp-(1→3)-β-d-GalpNAc | 5 | Figure 6 | ||
| APK37.1 | APK37.1_gp49 | KZ-1101 | K37 | β-d-GalpNAc-(1→3)-α-d-Galp | 6 | 7 | 8 | Figure 7A |
| AB5001 | K3-v1 | β-d-GalpNAc-(1→3)-α-d-Galp | 9 | 10 | Figure 7B | |||
| APK86 | APK86_gp49 | MAR55-66 | K86 | β-d-GalpNAc-(1→3)-α-L-Rhap | 11 | 12 | 13 | Figure 8 |
| APK127v | APK127v_gp47 | 36-1454 | K127 | β-d-GalpNAc-(1→3)-α-d-Galp | 14 | Figure 9 | ||
| APK128 | APK128_gp45 | KZ-1093 | K128 | β-d-GalpNAc-(1→4)-α-d-Galp | 15 | Figure 10 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Timoshina, O.Y.; Kasimova, A.A.; Shneider, M.M.; Matyuta, I.O.; Nikolaeva, A.Y.; Evseev, P.V.; Arbatsky, N.P.; Shashkov, A.S.; Chizhov, A.O.; Shelenkov, A.A.; et al. Friunavirus Phage-Encoded Depolymerases Specific to Different Capsular Types of Acinetobacter baumannii. Int. J. Mol. Sci. 2023, 24, 9100. https://doi.org/10.3390/ijms24109100
Timoshina OY, Kasimova AA, Shneider MM, Matyuta IO, Nikolaeva AY, Evseev PV, Arbatsky NP, Shashkov AS, Chizhov AO, Shelenkov AA, et al. Friunavirus Phage-Encoded Depolymerases Specific to Different Capsular Types of Acinetobacter baumannii. International Journal of Molecular Sciences. 2023; 24(10):9100. https://doi.org/10.3390/ijms24109100
Chicago/Turabian StyleTimoshina, Olga Y., Anastasia A. Kasimova, Mikhail M. Shneider, Ilya O. Matyuta, Alena Y. Nikolaeva, Peter V. Evseev, Nikolay P. Arbatsky, Alexander S. Shashkov, Alexander O. Chizhov, Andrey A. Shelenkov, and et al. 2023. "Friunavirus Phage-Encoded Depolymerases Specific to Different Capsular Types of Acinetobacter baumannii" International Journal of Molecular Sciences 24, no. 10: 9100. https://doi.org/10.3390/ijms24109100