
Transcriptomic Analysis of the Effect of Torin-2 on the Central Nervous System of Drosophila melanogaster
 ,
,  , , , ,
, , , ,  and
and 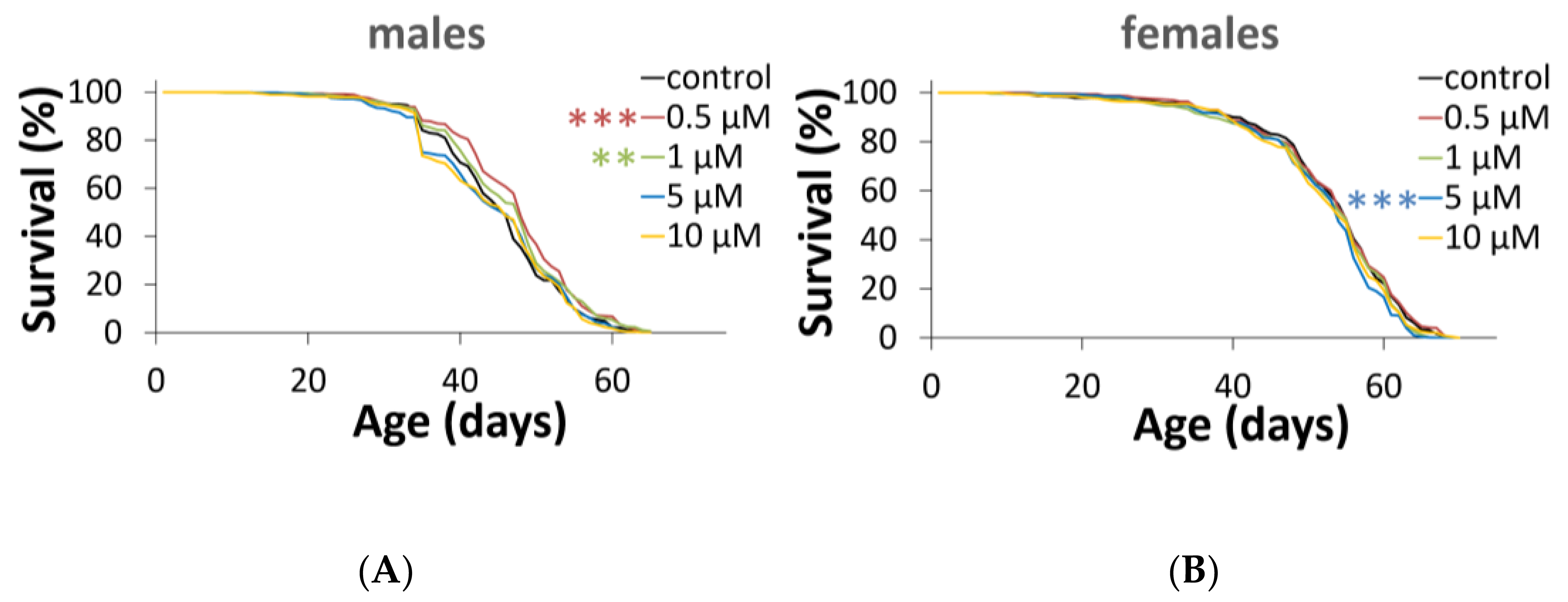{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
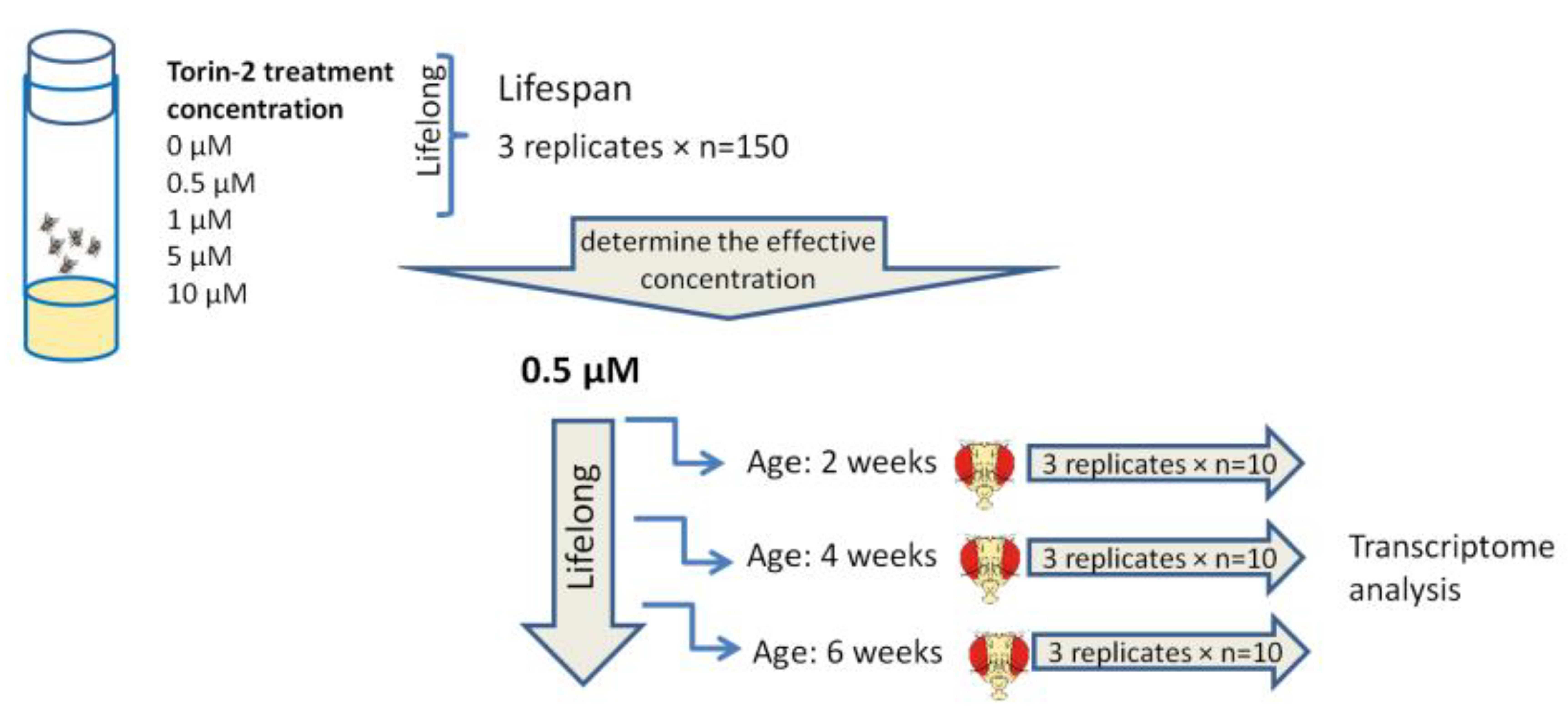
2.1. Selection of Concentrations and Longevity Analysis
2.2. Age-Associated Transcriptome Changes in Insects Receiving Torin-2 and Controls
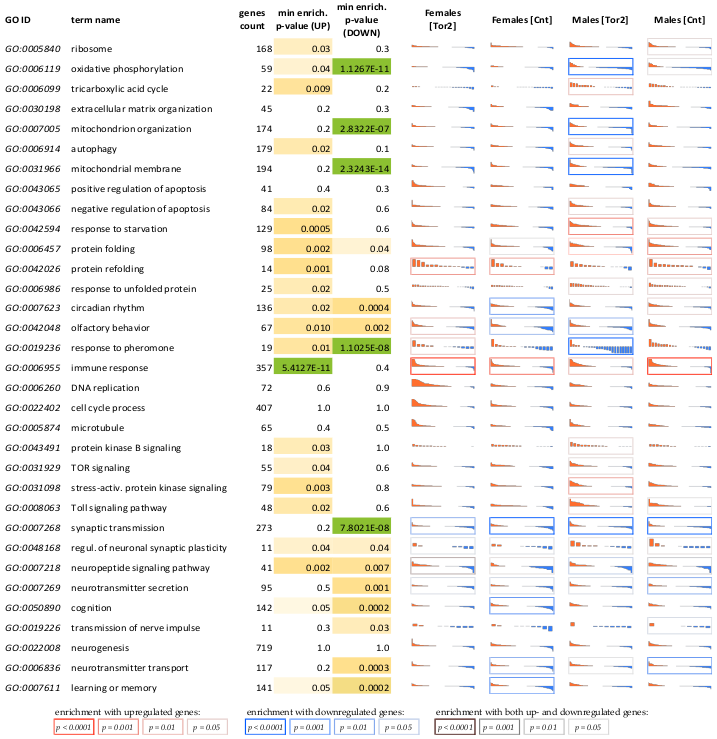
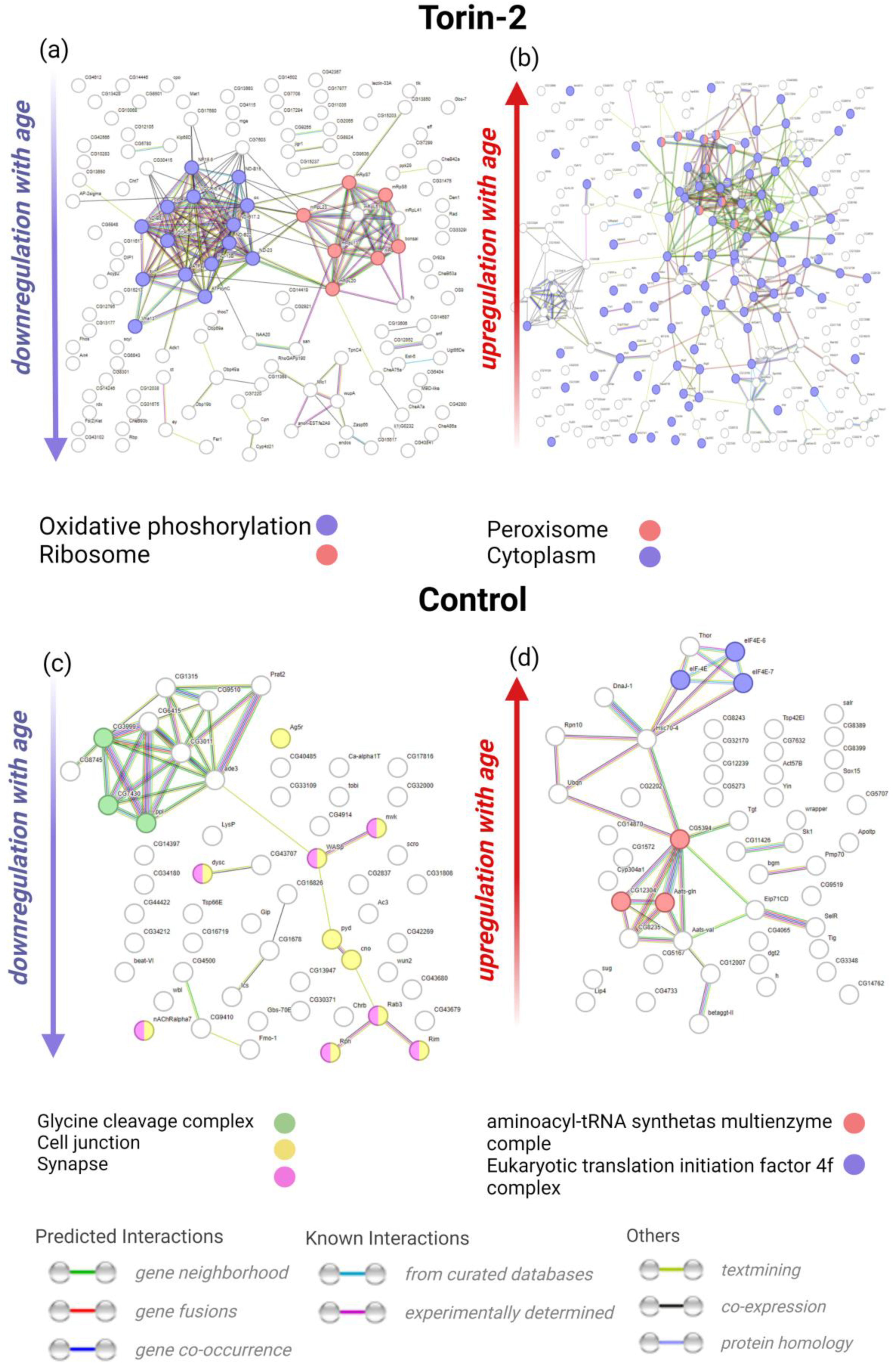
2.3. Analysis of Gene Sets Specifically Associated with Age in Either the Torin-2 Group or the Control Group
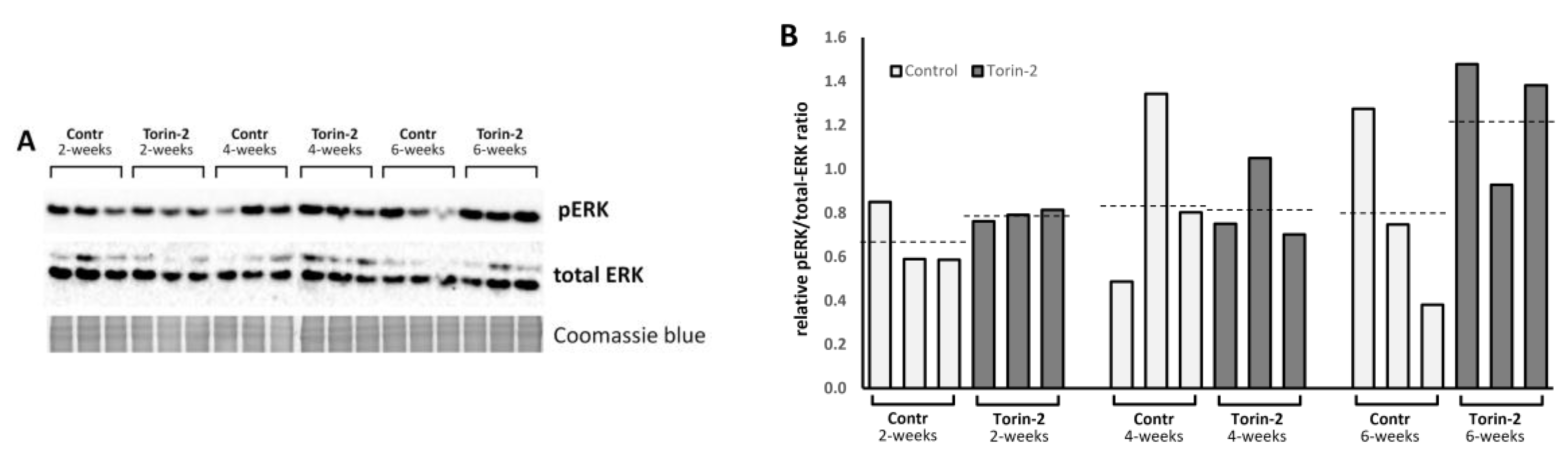
2.4. Expression of MAPK Pathway Genes (RNA-Seq Data)—Western Blot Analysis of Phosho- and Total ERK
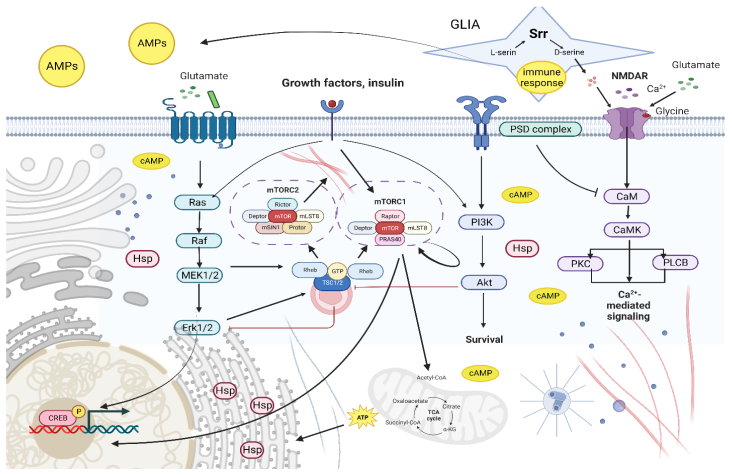
3. Discussion
4. Materials and Methods
4.1. Drosophila Strain and Experimental Conditions
4.2. Transcriptome Sequencing
4.3. Bioinformatic Analysis
4.4. Immunoblotting
4.5. Quantitative PCR
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| TORC1/2 | target of rapamycin complex1/complex2 |
| MAPK | a mitogen-activated protein kinase |
| NMDA | the N-methyl-D-aspartate receptor |
| LTP | long-term potentiation |
| BBB | blood–brain barrier |
| ERK1/2 extracellular signal | regulated kinase 1/2 |
| DEGs | differentially expressed genes |
| AMPs | antimicrobial peptides |
| cAMP | cyclic adenosine monophosphate |
| TCA | the citric acid cycle |
References
- Graham, W.V.; Bonito-Oliva, A.; Sakmar, T.P. Update on Alzheimer’s Disease Therapy and Prevention Strategies. Annu. Rev. Med. 2017, 68, 413–430. [Google Scholar] [CrossRef]
- Richardson, A.; Galvan, V.; Lin, A.-L.; Oddo, S. How Longevity Research Can Lead to Therapies for Alzheimer’s Disease: The Rapamycin Story. Exp. Gerontol. 2015, 68, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Saxton, R.A.; Sabatini, D.M. MTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 169, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Hoeffer, C.A.; Klann, E. MTOR Signaling: At the Crossroads of Plasticity, Memory and Disease. Trends Neurosci. 2010, 33, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.J.; Reis, G.; Kang, H.; Gingras, A.-C.; Sonenberg, N.; Schuman, E.M. A Rapamycin-Sensitive Signaling Pathway Contributes to Long-Term Synaptic Plasticity in the Hippocampus. Proc. Natl. Acad. Sci. USA 2002, 99, 467–472. [Google Scholar] [CrossRef]
- Dash, P.K.; Orsi, S.A.; Moore, A.N. Spatial Memory Formation and Memory-Enhancing Effect of Glucose Involves Activation of the Tuberous Sclerosis Complex–Mammalian Target of Rapamycin Pathway. J. Neurosci. 2006, 26, 8048–8056. [Google Scholar] [CrossRef]
- Costa-Mattioli, M.; Sossin, W.S.; Klann, E.; Sonenberg, N. Translational Control of Long-Lasting Synaptic Plasticity and Memory. Neuron 2009, 61, 10–26. [Google Scholar] [CrossRef]
- Sutton, M.A.; Schuman, E.M. Dendritic Protein Synthesis, Synaptic Plasticity, and Memory. Cell 2006, 127, 49–58. [Google Scholar] [CrossRef]
- Henry, F.E.; Hockeimer, W.; Chen, A.; Mysore, S.P.; Sutton, M.A. Mechanistic Target of Rapamycin Is Necessary for Changes in Dendritic Spine Morphology Associated with Long-Term Potentiation. Mol. Brain 2017, 10, 50. [Google Scholar] [CrossRef]
- Blagosklonny, M.V. Calorie Restriction: Decelerating MTOR-Driven Aging from Cells to Organisms (Including Humans). Cell Cycle 2010, 9, 683–688. [Google Scholar] [CrossRef]
- Roizen, M.F. Rapamycin Fed Late in Life Extends Lifespan in Genetically Heterogenous Mice. Yearb. Anesthesiol. Pain Manag. 2010, 2010, 15–16. [Google Scholar] [CrossRef]
- Lamming, D.W.; Ye, L.; Sabatini, D.M.; Baur, J.A. Rapalogs and MTOR Inhibitors as Anti-Aging Therapeutics. J. Clin. Investig. 2013, 123, 980–989. [Google Scholar] [CrossRef]
- Zhang, Y.; Bokov, A.; Gelfond, J.; Soto, V.; Ikeno, Y.; Hubbard, G.; Diaz, V.; Sloane, L.; Maslin, K.; Treaster, S.; et al. Rapamycin Extends Life and Health in C57BL/6 Mice. J. Gerontol. Ser. A 2014, 69, 119–130. [Google Scholar] [CrossRef]
- Colman, R.J.; Anderson, R.M.; Johnson, S.C.; Kastman, E.K.; Kosmatka, K.J.; Beasley, T.M.; Allison, D.B.; Cruzen, C.; Simmons, H.A.; Kemnitz, J.W.; et al. Caloric Restriction Delays Disease Onset and Mortality in Rhesus Monkeys. Science 2009, 325, 201–204. [Google Scholar] [CrossRef]
- Stanfel, M.N.; Shamieh, L.S.; Kaeberlein, M.; Kennedy, B.K. The TOR Pathway Comes of Age. Biochim. Biophys. Acta BBA Gen. Subj. 2009, 1790, 1067–1074. [Google Scholar] [CrossRef]
- Swiech, L.; Perycz, M.; Malik, A.; Jaworski, J. Role of MTOR in Physiology and Pathology of the Nervous System. Biochim. Biophys. Acta BBA Proteins Proteom. 2008, 1784, 116–132. [Google Scholar] [CrossRef]
- Sun, Q.; Wei, L.-L.; Zhang, M.; Li, T.-X.; Yang, C.; Deng, S.-P.; Zeng, Q.-C. Rapamycin Inhibits Activation of AMPK-MTOR Signaling Pathway-Induced Alzheimer’s Disease Lesion in Hippocampus of Rats with Type 2 Diabetes Mellitus. Int. J. Neurosci. 2019, 129, 184–193. [Google Scholar] [CrossRef]
- Li, J.; Kim, S.G.; Blenis, J. Rapamycin: One Drug, Many Effects. Cell Metab. 2014, 19, 373–379. [Google Scholar] [CrossRef]
- Waetzig, R.; Matthes, M.; Leister, J.; Penkivech, G.; Heise, T.; Corbacioglu, S.; Sommer, G. Comparing MTOR Inhibitor Rapamycin with Torin-2 within the RIST Molecular-Targeted Regimen in Neuroblastoma Cells. Int. J. Med. Sci. 2021, 18, 137–149. [Google Scholar] [CrossRef]
- Johnson, S.C.; Rabinovitch, P.S.; Kaeberlein, M. MTOR Is a Key Modulator of Ageing and Age-Related Disease. Nature 2013, 493, 338–345. [Google Scholar] [CrossRef]
- Ehninger, D.; Neff, F.; Xie, K. Longevity, Aging and Rapamycin. Cell. Mol. Life Sci. 2014, 71, 4325–4346. [Google Scholar] [CrossRef] [PubMed]
- Tee, A. The Target of Rapamycin and Mechanisms of Cell Growth. Ijms 2018, 19, 880. [Google Scholar] [CrossRef] [PubMed]
- McGuire, S.E.; Roman, G.; Davis, R.L. Gene Expression Systems in Drosophila: A Synthesis of Time and Space. Trends Genet. 2004, 20, 384–391. [Google Scholar] [CrossRef] [PubMed]
- Libert, S.; Zwiener, J.; Chu, X.; Vanvoorhies, W.; Roman, G.; Pletcher, S.D. Regulation of Drosophila Life Span by Olfaction and Food-Derived Odors. Science 2007, 315, 1133–1137. [Google Scholar] [CrossRef]
- Toivonen, J.M.; Partridge, L. Endocrine Regulation of Aging and Reproduction in Drosophila. Mol. Cell Endocrinol. 2009, 299, 39–50. [Google Scholar] [CrossRef]
- Partridge, L.; Barton, N.H. Evolution of Aging: Testing the Theory Using Drosophila. Genetica 1993, 91, 89–98. [Google Scholar] [CrossRef]
- Clancy, D.J.; Gems, D.; Harshman, L.G.; Oldham, S.; Stocker, H.; Hafen, E.; Leevers, S.J.; Partridge, L. Extension of Life-Span by Loss of CHICO, a Drosophila Insulin Receptor Substrate Protein. Science 2001, 292, 104–106. [Google Scholar] [CrossRef]
- Biteau, B.; Karpac, J.; Hwangbo, D.; Jasper, H. Regulation of Drosophila Lifespan by JNK Signaling. Exp. Gerontol. 2011, 46, 349–354. [Google Scholar] [CrossRef]
- Clancy, D.J.; Gems, D.; Hafen, E.; Leevers, S.J.; Partridge, L. Dietary Restriction in Long-Lived Dwarf Flies. Science 2002, 296, 319. [Google Scholar] [CrossRef]
- Tatar, M.; Kopelman, A.; Epstein, D.; Tu, M.P.; Yin, C.M.; Garofalo, R.S. A Mutant Drosophila Insulin Receptor Homolog That Extends Life-Span and Impairs Neuroendocrine Function. Science 2001, 292, 107–110. [Google Scholar] [CrossRef]
- Kapahi, P.; Zid, B.M.; Harper, T.; Koslover, D.; Sapin, V.; Benzer, S. Regulation of Lifespan in Drosophila by Modulation of Genes in the TOR Signaling Pathway. Curr. Biol. 2004, 14, 885–890. [Google Scholar] [CrossRef]
- Saitoe, M.; Horiuchi, J.; Tamura, T.; Ito, N. Drosophila as a Novel Animal Model for Studying the Genetics of Age-Related Memory Impairment. Rev. Neurosci. 2005, 16, 137–149. [Google Scholar] [CrossRef]
- Davie, K.; Janssens, J.; Koldere, D.; De Waegeneer, M.; Pech, U.; Kreft, Ł.; Aibar, S.; Makhzami, S.; Christiaens, V.; Bravo González-Blas, C.; et al. A Single-Cell Transcriptome Atlas of the Aging Drosophila Brain. Cell 2018, 174, 982–998.e20. [Google Scholar] [CrossRef]
- Moskalev, A.A.; Shaposhnikov, M.V.; Zemskaya, N.V.; Koval, L.A.; Schegoleva, E.V.; Guvatova, Z.G.; Krasnov, G.S.; Solovev, I.A.; Sheptyakov, M.A.; Zhavoronkov, A.; et al. Transcriptome Analysis of Long-Lived Drosophila Melanogaster E(z) Mutants Sheds Light on the Molecular Mechanisms of Longevity. Sci. Rep. 2019, 9, 9151. [Google Scholar] [CrossRef]
- Koch, S.C.; Nelson, A.; Hartenstein, V. Structural Aspects of the Aging Invertebrate Brain. Cell Tissue Res. 2021, 383, 931–947. [Google Scholar] [CrossRef]
- Partridge, L.; Alic, N.; Bjedov, I.; Piper, M.D.W. Ageing in Drosophila: The Role of the Insulin/Igf and TOR Signalling Network. Exp. Gerontol. 2011, 46, 376–381. [Google Scholar] [CrossRef]
- Whelan, J.T.; Hollis, S.E.; Cha, D.S.; Asch, A.S.; Lee, M.-H. Post-Transcriptional Regulation of the Ras-ERK/MAPK Signaling Pathway. J. Cell Physiol. 2012, 227, 1235–1241. [Google Scholar] [CrossRef]
- Landis, G.; Shen, J.; Tower, J. Gene Expression Changes in Response to Aging Compared to Heat Stress, Oxidative Stress and Ionizing Radiation in Drosophila Melanogaster. Aging 2012, 4, 768–789. [Google Scholar] [CrossRef]
- Garschall, K.; Flatt, T. The Interplay between Immunity and Aging in Drosophila. F1000Res 2018, 7, 160. [Google Scholar] [CrossRef]
- Hanson, M.A.; Lemaitre, B. Antimicrobial Peptides Do Not Directly Contribute to Aging in Drosophila, but Improve Lifespan by Preventing Dysbiosis. Dis. Model Mech. 2023, 16, dmm.049965. [Google Scholar] [CrossRef]
- Badinloo, M.; Nguyen, E.; Suh, W.; Alzahrani, F.; Castellanos, J.; Klichko, V.I.; Orr, W.C.; Radyuk, S.N. Overexpression of Antimicrobial Peptides Contributes to Aging through Cytotoxic Effects in Drosophila Tissues. Arch. Insect. Biochem. Physiol. 2018, 98, e21464. [Google Scholar] [CrossRef] [PubMed]
- Hanson, M.A.; Lemaitre, B. New Insights on Drosophila Antimicrobial Peptide Function in Host Defense and Beyond. Curr. Opin. Immunol. 2020, 62, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Mogk, A.; Schlieker, C.; Friedrich, K.L.; Schönfeld, H.-J.; Vierling, E.; Bukau, B. Refolding of Substrates Bound to Small Hsps Relies on a Disaggregation Reaction Mediated Most Efficiently by ClpB/DnaK. J. Biol. Chem. 2003, 278, 31033–31042. [Google Scholar] [CrossRef] [PubMed]
- Young, J.C.; Moarefi, I.; Hartl, F.U. Hsp90. J. Cell Biol. 2001, 154, 267–274. [Google Scholar] [CrossRef]
- Zatsepina, O.G.; Evgen’ev, M.B.; Garbuz, D.G. Role of a Heat Shock Transcription Factor and the Major Heat Shock Protein Hsp70 in Memory Formation and Neuroprotection. Cells 2021, 10, 1638. [Google Scholar] [CrossRef]
- Sandry, J.; Trafimow, D.; Marks, M.J.; Rice, S. Adaptive Memory: Evaluating Alternative Forms of Fitness-Relevant Processing in the Survival Processing Paradigm. PLoS ONE 2013, 8, e60868. [Google Scholar] [CrossRef]
- Forest, J.; Sunada, H.; Dodd, S.; Lukowiak, K. Training Lymnaea in the Presence of a Predator Scent Results in a Long-Lasting Ability to Form Enhanced Long-Term Memory. J. Comp. Physiol. A 2016, 202, 399–409. [Google Scholar] [CrossRef]
- Lukowiak, K.; Orr, M.; De Caigny, P.; Lukowiak, K.S.; Rosenegger, D.; Han, J.I.; Dalesman, S. Ecologically Relevant Stressors Modify Long-Term Memory Formation in a Model System. Behav. Brain Res. 2010, 214, 18–24. [Google Scholar] [CrossRef]
- Zhao, H.; Bucci, D.J.; Weltzin, M.; Drew, K.L. Effects of Aversive Stimuli on Learning and Memory in Arctic Ground Squirrels. Behav. Brain Res. 2004, 151, 219–224. [Google Scholar] [CrossRef]
- Ciechanover, A.; Kwon, Y.T. Protein Quality Control by Molecular Chaperones in Neurodegeneration. Front. Neurosci. 2017, 11, 185. [Google Scholar] [CrossRef]
- King, V.; Tower, J. Aging-Specific Expression OfDrosophila Hsp22. Dev. Biol. 1999, 207, 107–118. [Google Scholar] [CrossRef]
- Morrow, G.; Tanguay, R.M. Heat Shock Proteins and Aging in Drosophila Melanogaster. Semin. Cell Dev. Biol. 2003, 14, 291–299. [Google Scholar] [CrossRef]
- Wang, H.-D.; Kazemi-Esfarjani, P.; Benzer, S. Multiple-Stress Analysis for Isolation of Drosophila Longevity Genes. Proc. Natl. Acad. Sci. USA 2004, 101, 12610–12615. [Google Scholar] [CrossRef]
- Kurapati, R.; Passananti, H.B.; Rose, M.R.; Tower, J. Increased Hsp22 RNA Levels in Drosophila Lines Genetically Selected for Increased Longevity. J. Gerontol. A Biol. Sci. Med. Sci. 2000, 55, B552–B559. [Google Scholar] [CrossRef]
- Xiao, C.; Hull, D.; Qiu, S.; Yeung, J.; Zheng, J.; Barwell, T.; Robertson, R.M.; Seroude, L. Expression of Heat Shock Protein 70 Is Insufficient To Extend Drosophila Melanogaster Longevity. G3 2019, 9, 4197–4207. [Google Scholar] [CrossRef]
- Karney-Grobe, S.; Russo, A.; Frey, E.; Milbrandt, J.; DiAntonio, A. HSP90 Is a Chaperone for DLK and Is Required for Axon Injury Signaling. Proc. Natl. Acad. Sci. USA 2018, 115, E9899–E9908. [Google Scholar] [CrossRef]
- Ardestani, A.; Maedler, K. MTORC1 and IRS1: Another Deadly Kiss. Trends Endocrinol. Metab. 2018, 29, 737–739. [Google Scholar] [CrossRef]
- Shilova, V.; Zatsepina, O.; Zakluta, A.; Karpov, D.; Chuvakova, L.; Garbuz, D.; Evgen’ev, M. Age-Dependent Expression Profiles of Two Adaptogenic Systems and Thermotolerance in Drosophila Melanogaster. Cell Stress Chaperones 2020, 25, 305–315. [Google Scholar] [CrossRef]
- Markmiller, S.; Soltanieh, S.; Server, K.L.; Mak, R.; Jin, W.; Fang, M.Y.; Luo, E.-C.; Krach, F.; Yang, D.; Sen, A.; et al. Context-Dependent and Disease-Specific Diversity in Protein Interactions within Stress Granules. Cell 2018, 172, 590–604.e13. [Google Scholar] [CrossRef]
- Zhong, Y.; Wu, C.-F. Neuronal Activity and Adenylyl Cyclase in Environment-Dependent Plasticity of Axonal Outgrowth in Drosophila. J. Neurosci. 2004, 24, 1439–1445. [Google Scholar] [CrossRef]
- Villa, A.; Gelosa, P.; Castiglioni, L.; Cimino, M.; Rizzi, N.; Pepe, G.; Lolli, F.; Marcello, E.; Sironi, L.; Vegeto, E.; et al. Sex-Specific Features of Microglia from Adult Mice. Cell Rep. 2018, 23, 3501–3511. [Google Scholar] [CrossRef] [PubMed]
- Hanamsagar, R.; Alter, M.D.; Block, C.S.; Sullivan, H.; Bolton, J.L.; Bilbo, S.D. Generation of a Microglial Developmental Index in Mice and in Humans Reveals a Sex Difference in Maturation and Immune Reactivity. Glia 2017, 65, 1504–1520. [Google Scholar] [CrossRef] [PubMed]
- Angele, M.K.; Schwacha, M.G.; Ayala, A.; Chaudry, I.H. Effect of Gender and Sex Hormones on Immune Responses following Shock. Shock 2000, 14, 81–90. [Google Scholar] [CrossRef]
- Pettersson, U.S.; Waldén, T.B.; Carlsson, P.-O.; Jansson, L.; Phillipson, M. Female Mice Are Protected against High-Fat Diet Induced Metabolic Syndrome and Increase the Regulatory T Cell Population in Adipose Tissue. PLoS ONE 2012, 7, e46057. [Google Scholar] [CrossRef] [PubMed]
- Christensen, R.H.; Von Scholten, B.J.; Hansen, C.S.; Jensen, M.T.; Vilsbøll, T.; Rossing, P.; Jørgensen, P.G. Epicardial Adipose Tissue Predicts Incident Cardiovascular Disease and Mortality in Patients with Type 2 Diabetes. Cardiovasc. Diabetol. 2019, 18, 114. [Google Scholar] [CrossRef]
- Fontes, G.S.; McCarthy, R.J.; Kutzler, M.A.; Zitek-Morrison, E. The Effects of Sex and Neuter Status on Trauma Survival in Dogs: A Veterinary Committee on Trauma Registry Study. J. Vet. Emergen. Crit. Care 2022, 32, 756–763. [Google Scholar] [CrossRef]
- Yamamoto, D.; Sato, K.; Koganezawa, M. Neuroethology of Male Courtship in Drosophila: From the Gene to Behavior. J. Comp. Physiol. A 2014, 200, 251–264. [Google Scholar] [CrossRef]
- Sumien, N.; Cunningham, J.T.; Davis, D.L.; Engelland, R.; Fadeyibi, O.; Farmer, G.E.; Mabry, S.; Mensah-Kane, P.; Trinh, O.T.P.; Vann, P.H.; et al. Neurodegenerative Disease: Roles for Sex, Hormones, and Oxidative Stress. Endocrinology 2021, 162, bqab185. [Google Scholar] [CrossRef]
- Okamoto, N.; Watanabe, A. Interorgan Communication through Peripherally Derived Peptide Hormones in Drosophila. Fly 2022, 16, 152–176. [Google Scholar] [CrossRef]
- Pawate, S.; Shen, Q.; Fan, F.; Bhat, N.R. Redox Regulation of Glial Inflammatory Response to Lipopolysaccharide and Interferon? J. Neurosci. Res. 2004, 77, 540–551. [Google Scholar] [CrossRef]
- Qin, L.; Li, G.; Qian, X.; Liu, Y.; Wu, X.; Liu, B.; Hong, J.-S.; Block, M.L. Interactive Role of the Toll-like Receptor 4 and Reactive Oxygen Species in LPS-Induced Microglia Activation. Glia 2005, 52, 78–84. [Google Scholar] [CrossRef]
- Gomes-Leal, W. Microglial Physiopathology: How to Explain the Dual Role of Microglia after Acute Neural Disorders? Brain Behav. 2012, 2, 345–356. [Google Scholar] [CrossRef]
- Billard, J.-M. D-Serine Signalling as a Prominent Determinant of Neuronal-Glial Dialogue in the Healthy and Diseased Brain. J. Cell. Mol. Med. 2008, 12, 1872–1884. [Google Scholar] [CrossRef]
- Nickels, S.; Schwamborn, J. Is Serine Racemase (SRR) a Second Hit Target for LRRK2-G2019S Induced Parkinson’s Disease? Neural. Regen. Res. 2021, 16, 498. [Google Scholar] [CrossRef]
- Williams, S.M.; Diaz, C.M.; Macnab, L.T.; Sullivan, R.K.P.; Pow, D.V. Immunocytochemical Analysis OfD-Serine Distribution in the Mammalian Brain Reveals Novel Anatomical Compartmentalizations in Glia and Neurons. Glia 2006, 53, 401–411. [Google Scholar] [CrossRef]
- Steinmetz, R.D.; Fava, E.; Nicotera, P.; Steinhilber, D. A Simple Cell Line Based in Vitro Test System for N-Methyl-d-Aspartate (NMDA) Receptor Ligands. J. Neurosci. Methods 2002, 113, 99–110. [Google Scholar] [CrossRef]
- Katsuki, H.; Watanabe, Y.; Fujimoto, S.; Kume, T.; Akaike, A. Contribution of Endogenous Glycine and D-Serine to Excitotoxic and Ischemic Cell Death in Rat Cerebrocortical Slice Cultures. Life Sci. 2007, 81, 740–749. [Google Scholar] [CrossRef]
- Billard, J.-M. D-Serine in the Aging Hippocampus. J. Pharm. Biomed. Anal. 2015, 116, 18–24. [Google Scholar] [CrossRef]
- Ploux, E.; Freret, T.; Billard, J.-M. D-Serine in Physiological and Pathological Brain Aging. Biochim. Biophys. Acta BBA Proteins Proteom. 2021, 1869, 140542. [Google Scholar] [CrossRef]
- Wu, S.; Zhou, J.; Zhang, H.; Barger, S.W. Serine Racemase Expression Differentiates Aging from Alzheimer’s Brain. CAR 2022, 19, 494–502. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, S.; Li, W. RSeQC: Quality Control of RNA-Seq Experiments. Bioinformatics 2012, 28, 2184–2185. [Google Scholar] [CrossRef] [PubMed]
- Sigurgeirsson, B.; Emanuelsson, O.; Lundeberg, J. Sequencing Degraded RNA Addressed by 3′ Tag Counting. PLoS ONE 2014, 9, e91851. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. ClusterProfiler: An R Package for Comparing Biological Themes Among Gene Clusters. OMICS A J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Wang, C.; Li, Q.; Redden, D.T.; Weindruch, R.; Allison, D.B. Statistical Methods for Testing Effects on “Maximum Lifespan”. Mech. Ageing Dev. 2004, 125, 629–632. [Google Scholar] [CrossRef]
- Han, S.K.; Lee, D.; Lee, H.; Kim, D.; Son, H.G.; Yang, J.-S.; Lee, S.-J.V.; Kim, S. OASIS 2: Online Application for Survival Analysis 2 with Features for the Analysis of Maximal Lifespan and Healthspan in Aging Research. Oncotarget 2016, 7, 56147–56152. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vershinina, Y.S.; Krasnov, G.S.; Garbuz, D.G.; Shaposhnikov, M.V.; Fedorova, M.S.; Pudova, E.A.; Katunina, I.V.; Kornev, A.B.; Zemskaya, N.V.; Kudryavtsev, A.A.; et al. Transcriptomic Analysis of the Effect of Torin-2 on the Central Nervous System of Drosophila melanogaster. Int. J. Mol. Sci. 2023, 24, 9095. https://doi.org/10.3390/ijms24109095
Vershinina YS, Krasnov GS, Garbuz DG, Shaposhnikov MV, Fedorova MS, Pudova EA, Katunina IV, Kornev AB, Zemskaya NV, Kudryavtsev AA, et al. Transcriptomic Analysis of the Effect of Torin-2 on the Central Nervous System of Drosophila melanogaster. International Journal of Molecular Sciences. 2023; 24(10):9095. https://doi.org/10.3390/ijms24109095
Chicago/Turabian StyleVershinina, Yulia S., George S. Krasnov, David G. Garbuz, Mikhail V. Shaposhnikov, Maria S. Fedorova, Elena A. Pudova, Irina V. Katunina, Alexey B. Kornev, Nadezhda V. Zemskaya, Alexander A. Kudryavtsev, and et al. 2023. "Transcriptomic Analysis of the Effect of Torin-2 on the Central Nervous System of Drosophila melanogaster" International Journal of Molecular Sciences 24, no. 10: 9095. https://doi.org/10.3390/ijms24109095