


Stability of Human Telomeric G-Quadruplexes Complexed with Photosensitive Ligands and Irradiated with Visible Light
 , , , ,
, , , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
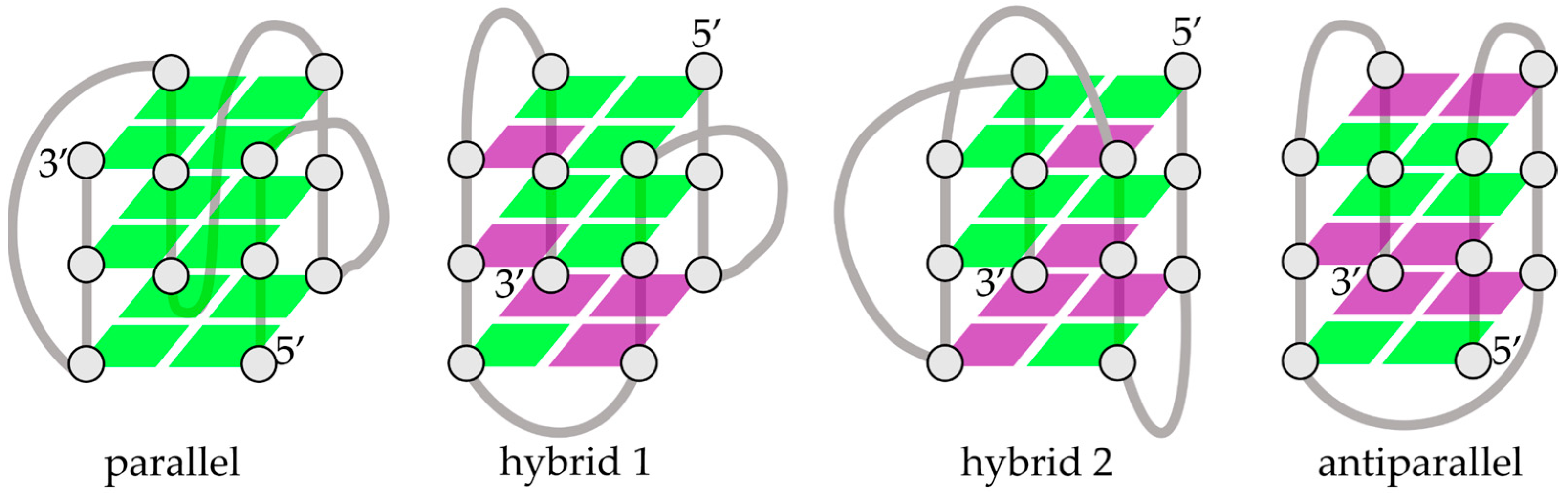
2.1. Light as a Driving Factor for Conformational Changes in G4
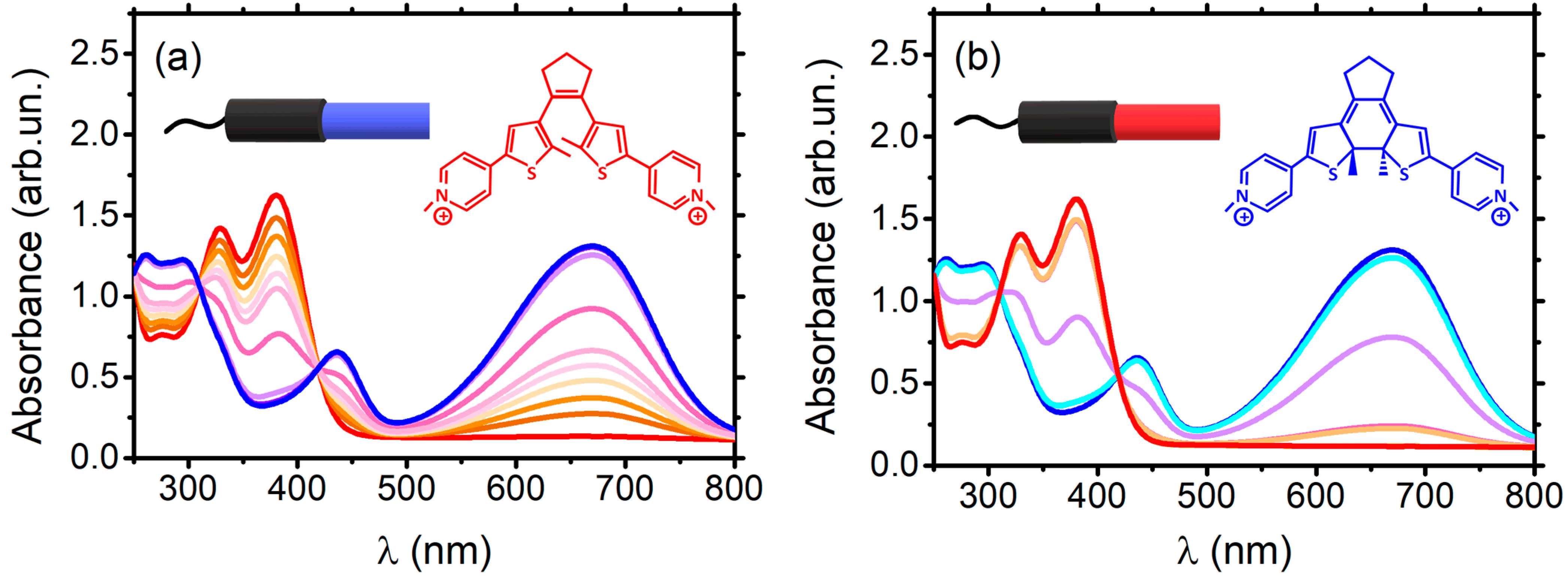
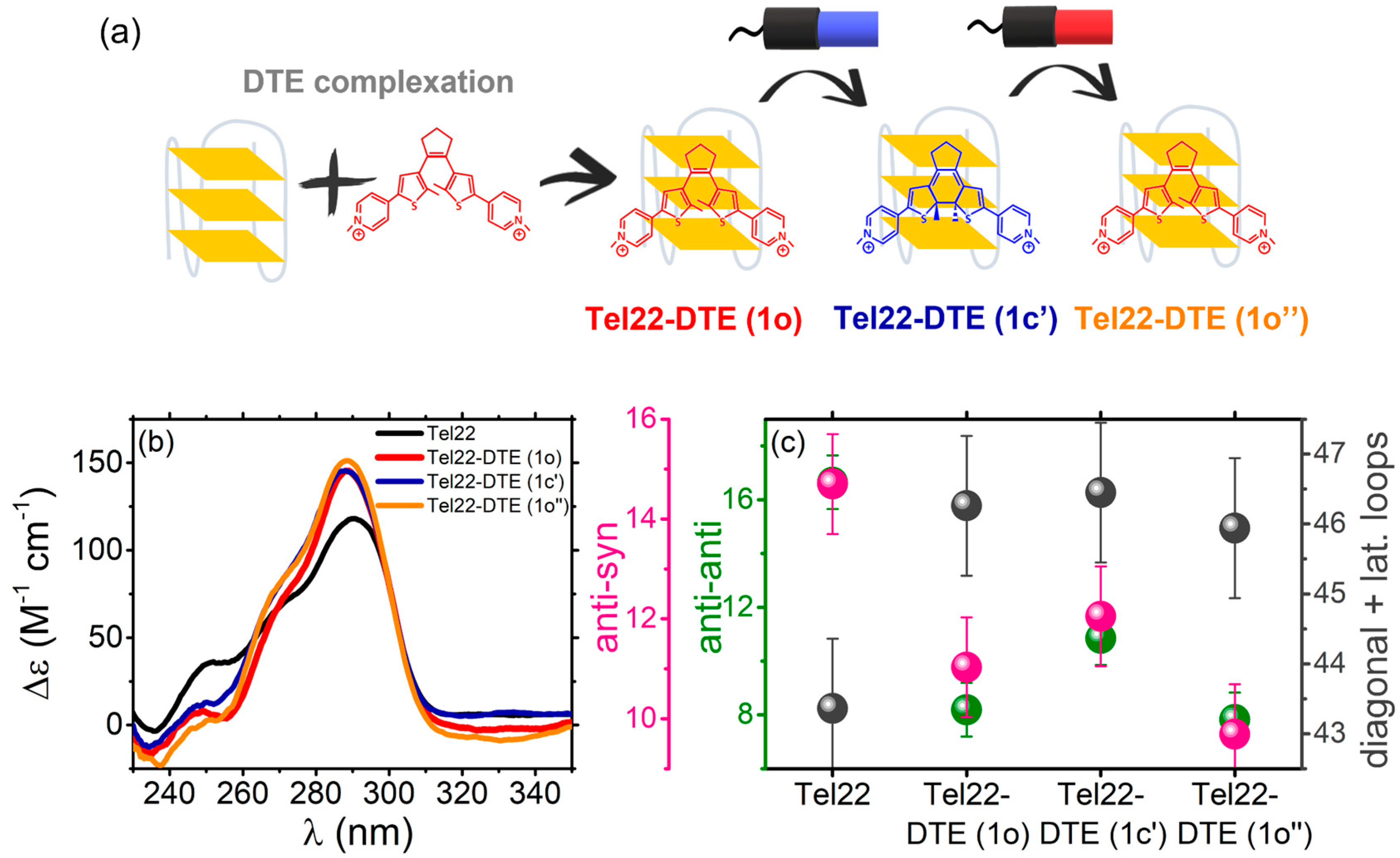
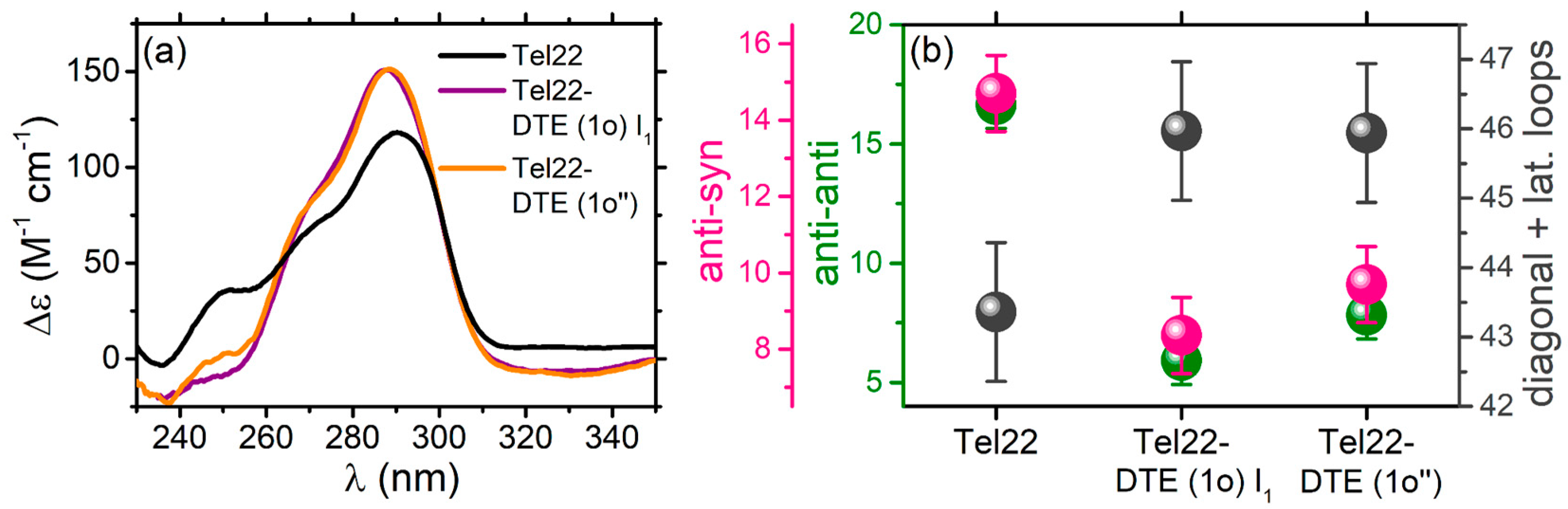
2.1.1. Tel22–DTE Complex
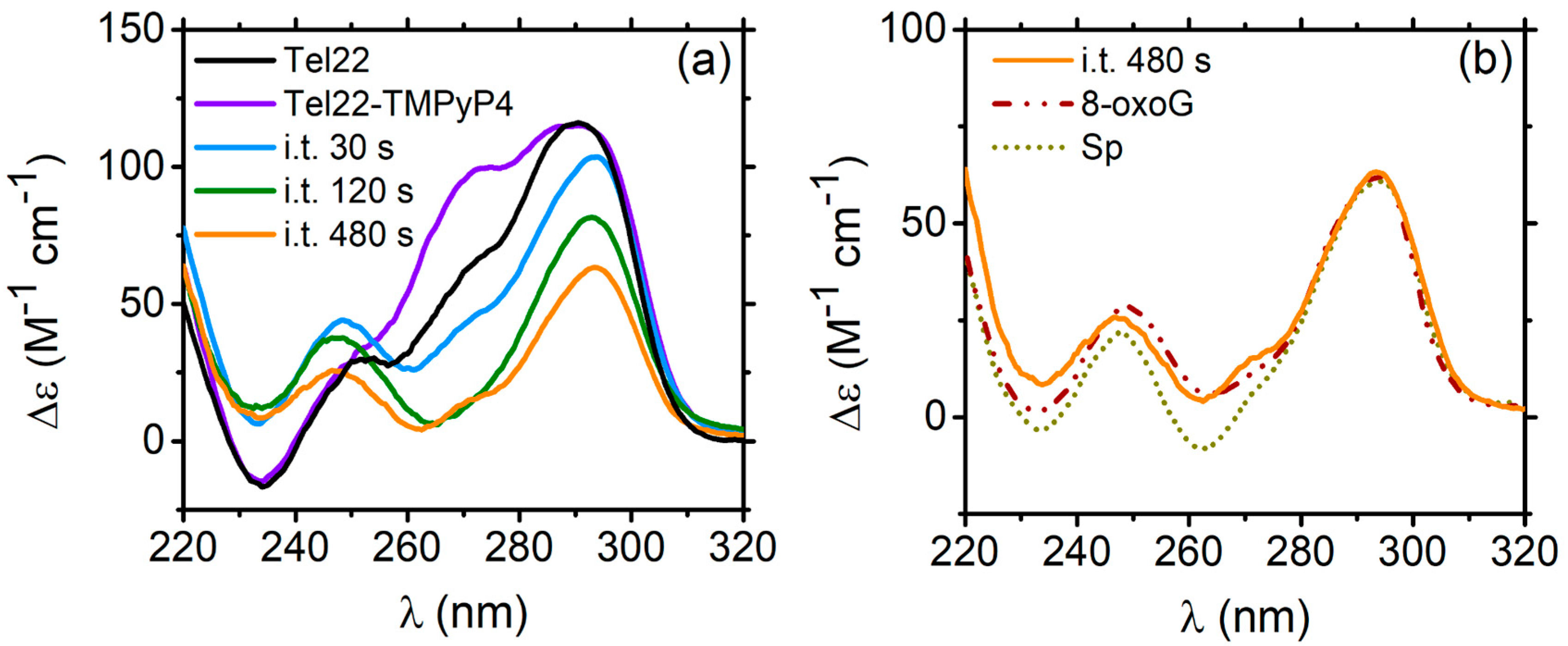
2.1.2. Tel22–TMPyP4 Complex
2.2. Conformational Changes upon Thermal Unfolding
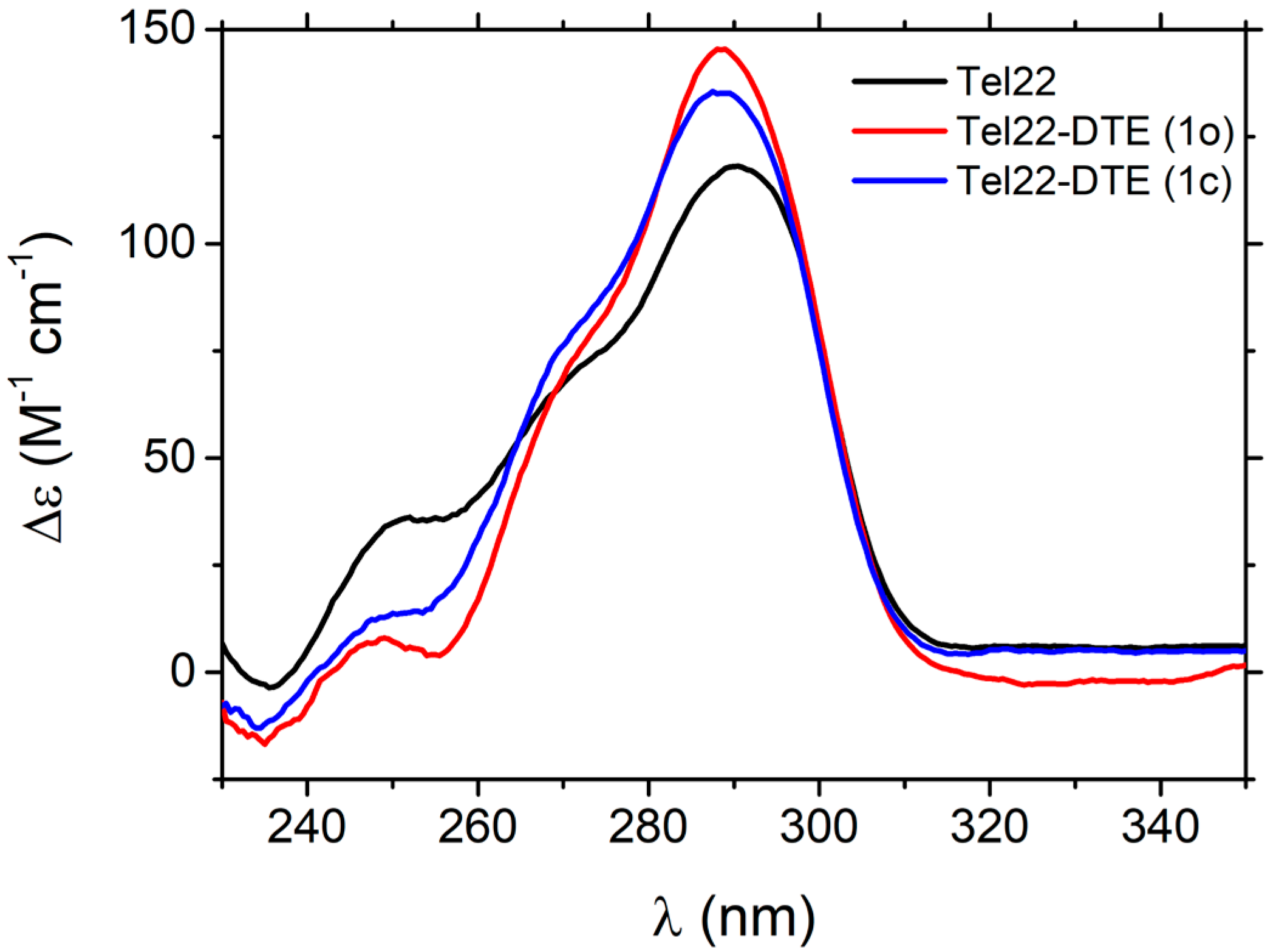
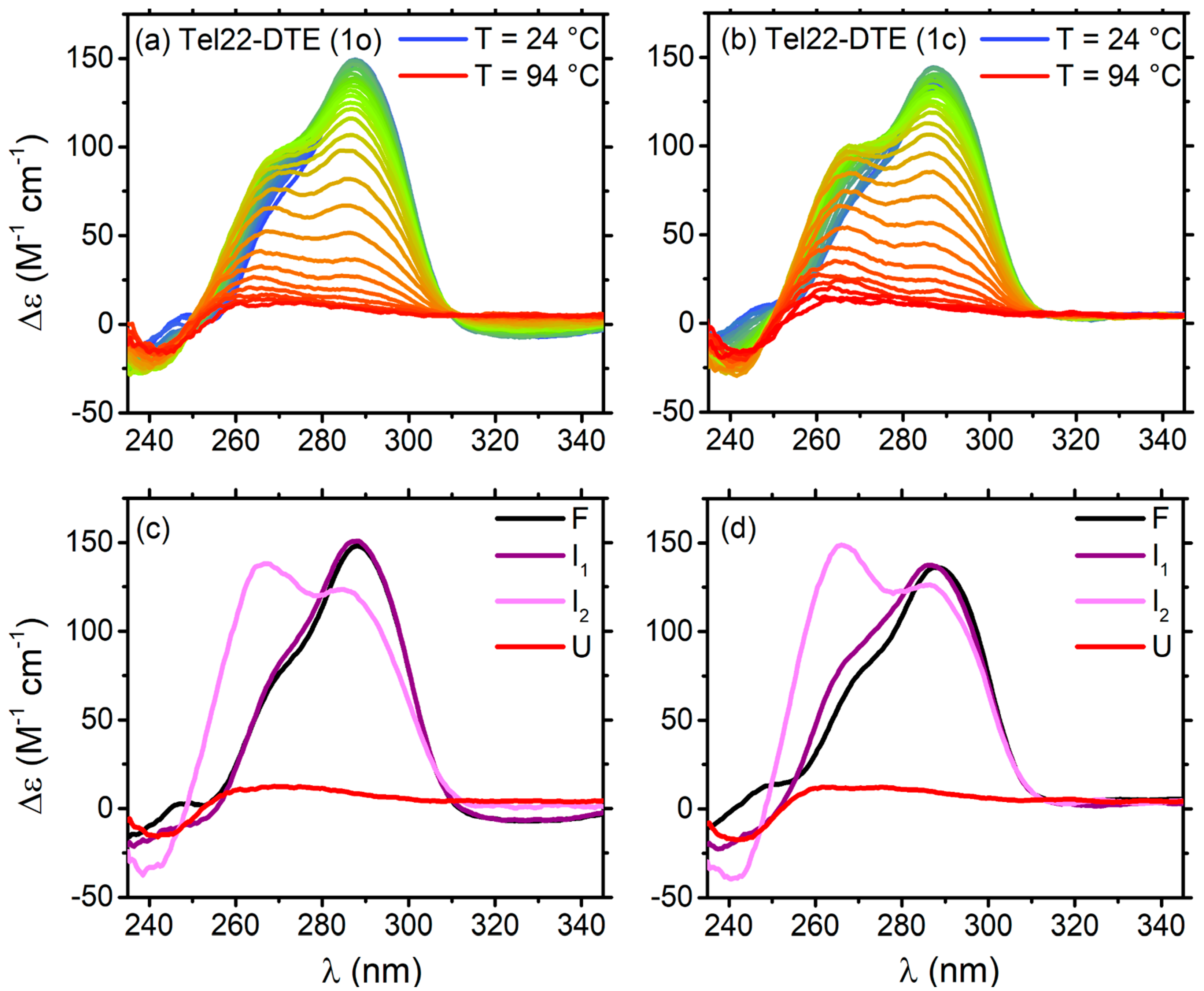
2.2.1. Thermal Unfolding of Tel22–DTE Complexes
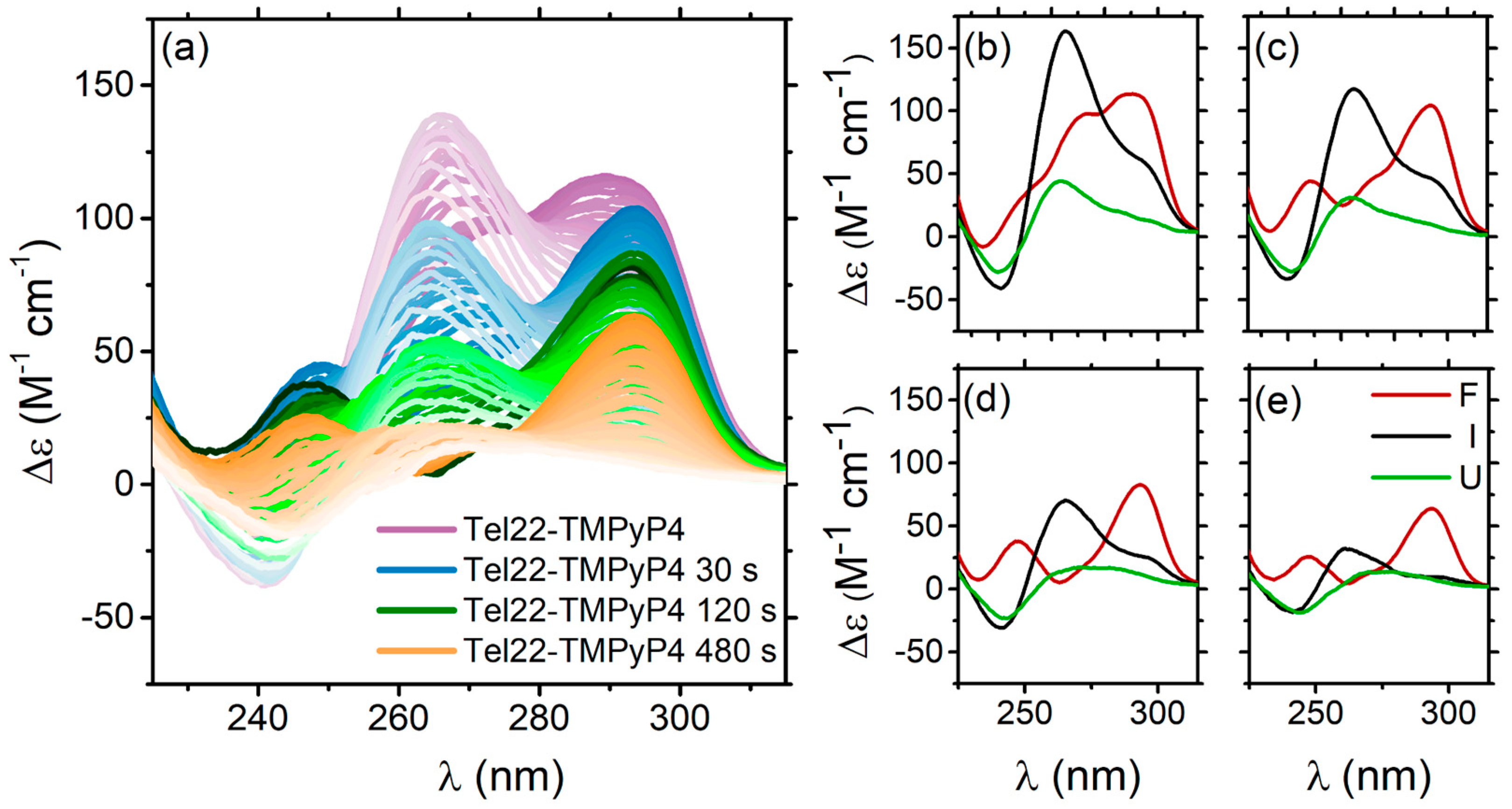
2.2.2. Thermal Unfolding of Tel22–TMPyP4 Systems
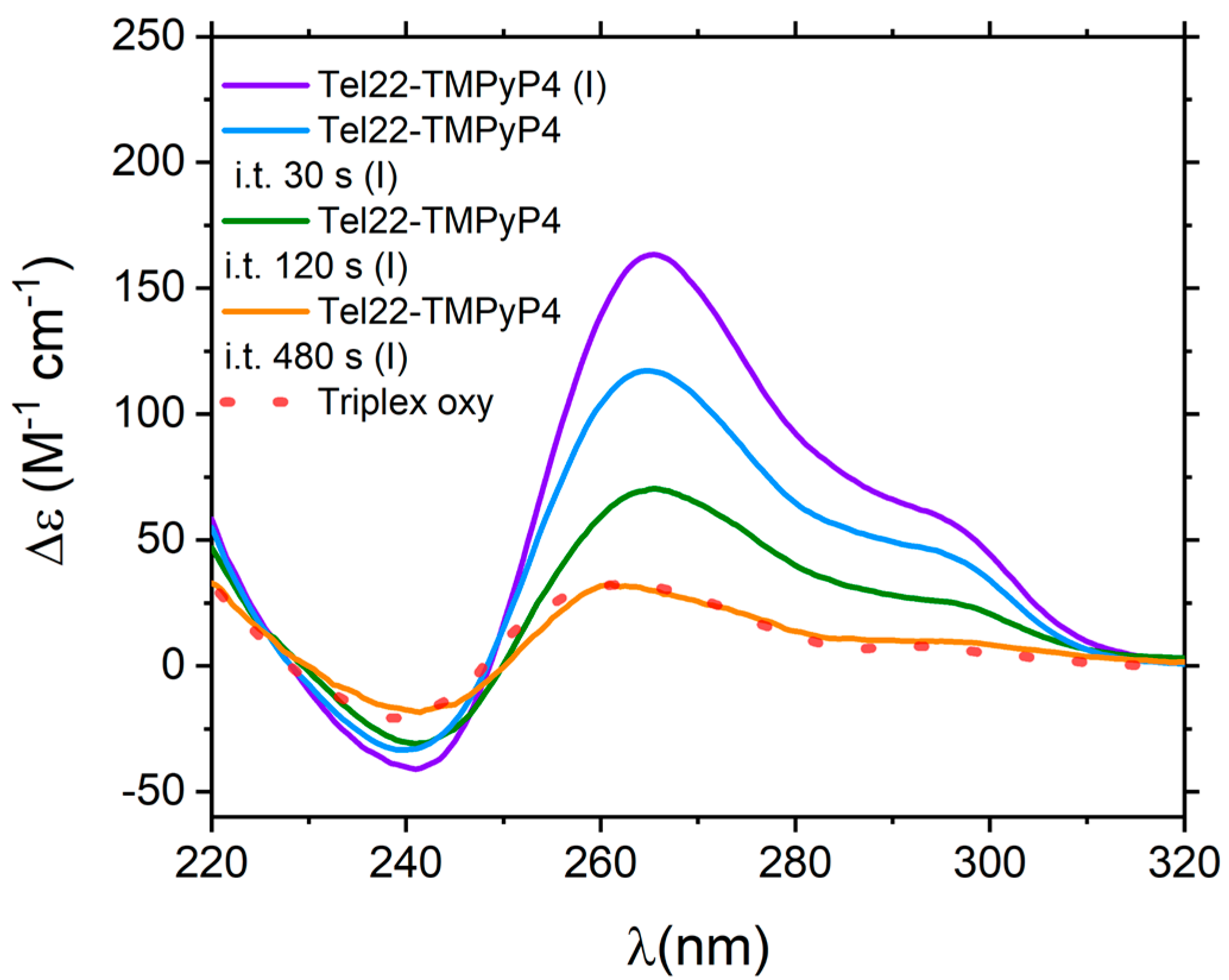
3. Discussions
4. Materials and Methods
4.1. UV-Vis Absorption Spectroscopy
4.2. CD Experiments
4.3. SVD Details
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bléger, D.; Hecht, S. Visible-Light-Activated Molecular Switches. Angew. Chem. Int. Ed Engl. 2015, 54, 11338–11349. [Google Scholar] [CrossRef] [PubMed]
- O’Hagan, M.P.; Ramos-Soriano, J.; Haldar, S.; Sheikh, S.; Morales, J.C.; Mulholland, A.J.; Galan, M.C. Visible-Light Photoswitching of Ligand Binding Mode Suggests G-Quadruplex DNA as a Target for Photopharmacology. Chem. Commun. 2020, 56, 5186–5189. [Google Scholar] [CrossRef] [PubMed]
- Sedighi, M.; Mahmoudi, Z.; Ghasempour, A.; Shakibaie, M.; Ghasemi, F.; Akbari, M.; Abbaszadeh, S.; Mostafavi, E.; Santos, H.A.; Shahbazi, M.-A. Nanostructured Multifunctional Stimuli-Responsive Glycopolypeptide-Based Copolymers for Biomedical Applications. J. Control. Release 2023, 354, 128–145. [Google Scholar] [CrossRef]
- Guo, Y.; Zhou, L.; Xu, L.; Zhou, X.; Hu, J.; Pei, R. Multiple Types of Logic Gates Based on a Single G-Quadruplex DNA Strand. Sci. Rep. 2014, 4, 7315. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Jiang, R.; He, H.; Ma, C.; Tang, Z. Recent Advances on G-Quadruplex for Biosensing, Bioimaging and Cancer Therapy. Trends Analyt. Chem. 2021, 139, 116257. [Google Scholar] [CrossRef]
- Sen, D.; Gilbert, W. Formation of Parallel Four-Stranded Complexes by Guanine-Rich Motifs in DNA and Its Implications for Meiosis. Nature 1988, 334, 364–366. [Google Scholar] [CrossRef]
- Neidle, S.; Parkinson, G. Telomere Maintenance as a Target for Anticancer Drug Discovery. Nat. Rev. Drug Discov. 2002, 1, 383–393. [Google Scholar] [CrossRef]
- Biffi, G.; Tannahill, D.; McCafferty, J.; Balasubramanian, S. Quantitative Visualization of DNA G-Quadruplex Structures in Human Cells. Nat. Chem. 2013, 5, 182–186. [Google Scholar] [CrossRef]
- Henderson, A.; Wu, Y.; Huang, Y.C.; Chavez, E.A.; Platt, J.; Johnson, F.B.; Brosh, R.M.; Sen, D.; Lansdorp, P.M. Detection of G-Quadruplex DNA in Mammalian Cells. Nucleic Acids Res. 2014, 42, 860–869. [Google Scholar] [CrossRef]
- Williamson, J.R.; Raghuraman, M.K.; Cech, T.R. Monovalent Cation-Induced Structure of Telomeric DNA: The G-Quartet Model. Cell 1989, 59, 871–880. [Google Scholar] [CrossRef]
- Ramos-Soriano, J.; Galan, M.C. Photoresponsive Control of G-Quadruplex DNA Systems. J. Am. Chem. Soc. Au 2021, 1, 1516–1526. [Google Scholar] [CrossRef] [PubMed]
- Kawauchi, K.; Urano, R.; Kinoshita, N.; Kuwamoto, S.; Torii, T.; Hashimoto, Y.; Taniguchi, S.; Tsuruta, M.; Miyoshi, D. Photosensitizers Based on G-Quadruplex Ligand for Cancer Photodynamic Therapy. Genes 2020, 11, 1340. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.; Cui, Y.-X.; Wang, J.; Zhang, J.; Zhu, L.-N.; Kong, D.-M. G-Quadruplex/Porphyrin Composite Photosensitizer: A Facile Way to Promote Absorption Redshift and Photodynamic Therapy Efficacy. ACS Appl. Mater. Interfaces 2019, 11, 13158–13167. [Google Scholar] [CrossRef] [PubMed]
- Neidle, S.; Parkinson, G.N. The Structure of Telomeric DNA. Curr. Opin. Struct. Biol. 2003, 13, 275–283. [Google Scholar] [CrossRef]
- Largy, E.; Mergny, J.-L.; Gabelica, V. Role of Alkali Metal Ions in G-Quadruplex Nucleic Acid Structure and Stability. Alkali Met. Ions Role Life 2016, 203–258. [Google Scholar]
- Webba da Silva, M.; da Silva, M.W. Geometric Formalism for DNA Quadruplex Folding. Chem.-A Eur. J. 2007, 13, 9738–9745. [Google Scholar] [CrossRef]
- Lim, K.W.; Ng, V.C.M.; Martín-Pintado, N.; Heddi, B.; Phan, A.T. Structure of the Human Telomere in Na Solution: An Antiparallel (2 2) G-Quadruplex Scaffold Reveals Additional Diversity. Nucleic Acids Res. 2013, 41, 10556–10562. [Google Scholar] [CrossRef]
- Wang, Y.; Patel, D.J. Solution Structure of a Parallel-Stranded G-Quadruplex DNA. J. Mol. Biol. 1993, 234, 1171–1183. [Google Scholar] [CrossRef]
- Gray, R.D.; Trent, J.O.; Arumugam, S.; Chaires, J.B. Folding Landscape of a Parallel G-Quadruplex. J. Phys. Chem. Lett. 2019, 10, 1146–1151. [Google Scholar] [CrossRef]
- Luu, K.N.; Phan, A.T.; Kuryavyi, V.; Lacroix, L.; Patel, D.J. Structure of the Human Telomere in K Solution: An Intramolecular (3 1) G-Quadruplex Scaffold. J. Am. Chem. Soc. 2006, 128, 9963–9970. [Google Scholar] [CrossRef]
- Dvorkin, S.A.; Karsisiotis, A.I.; Webba da Silva, M. Encoding Canonical DNA Quadruplex Structure. Sci Adv 2018, 4, eaat3007. [Google Scholar] [CrossRef] [PubMed]
- del Villar-Guerra, R.; del Villar-Guerra, R.; Trent, J.O.; Chaires, J.B. G-Quadruplex Secondary Structure Obtained from Circular Dichroism Spectroscopy. Angew. Chem. 2018, 57, 7171–7175. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, F.; Comez, L.; Biehl, R.; D’Amico, F.; Gessini, A.; Longo, M.; Masciovecchio, C.; Petrillo, C.; Radulescu, A.; Rossi, B.; et al. Structure of Human Telomere G-Quadruplex in the Presence of a Model Drug along the Thermal Unfolding Pathway. Nucleic Acids Res. 2018, 46, 11927–11938. [Google Scholar] [CrossRef] [PubMed]
- Parkinson, G.N.; Lee, M.P.H.; Neidle, S. Crystal Structure of Parallel Quadruplexes from Human Telomeric DNA. Nature 2002, 417, 876–880. [Google Scholar] [CrossRef]
- Comez, L.; Bianchi, F.; Libera, V.; Longo, M.; Petrillo, C.; Sacchetti, F.; Sebastiani, F.; D’Amico, F.; Rossi, B.; Gessini, A.; et al. Polymorphism of Human Telomeric Quadruplexes with Drugs: A Multi-Technique Biophysical Study. Phys. Chem. Chem. Phys. 2020, 22, 11583–11592. [Google Scholar] [CrossRef]
- Bertini, L.; Libera, V.; Ripanti, F.; Natali, F.; Paolantoni, M.; Orecchini, A.; Nucara, A.; Petrillo, C.; Comez, L.; Paciaroni, A. Polymorphism and Ligand Binding Modulate Fast Dynamics of Human Telomeric G-Quadruplexes. Int. J. Mol. Sci. 2023, 24, 4280. [Google Scholar] [CrossRef]
- Wang, Y.; Patel, D.J. Solution Structure of the Human Telomeric Repeat d[AG3(T2AG3)3] G-Tetraplex. Structure 1993, 1, 263–282. [Google Scholar] [CrossRef]
- Libera, V.; Bianchi, F.; Rossi, B.; D’Amico, F.; Masciovecchio, C.; Petrillo, C.; Sacchetti, F.; Paciaroni, A.; Comez, L. Solvent Vibrations as a Proxy of the Telomere G-Quadruplex Rearrangements across Thermal Unfolding. Int. J. Mol. Sci. 2022, 23, 5123. [Google Scholar] [CrossRef]
- Renciuk, D.; Kejnovská, I.; Skoláková, P.; Bednárová, K.; Motlová, J.; Vorlícková, M. Arrangements of Human Telomere DNA Quadruplex in Physiologically Relevant K+ Solutions. Nucleic Acids Res. 2009, 37, 6625–6634. [Google Scholar] [CrossRef]
- Bao, H.-L.; Liu, H.-S.; Xu, Y. Hybrid-Type and Two-Tetrad Antiparallel Telomere DNA G-Quadruplex Structures in Living Human Cells. Nucleic Acids Res. 2019, 47, 4940–4947. [Google Scholar] [CrossRef]
- Mashimo, T.; Yagi, H.; Sannohe, Y.; Rajendran, A.; Sugiyama, H. Folding Pathways of Human Telomeric Type-1 and Type-2 G-Quadruplex Structures. J. Am. Chem. Soc. 2010, 132, 14910–14918. [Google Scholar] [CrossRef] [PubMed]
- Gray, R.D.; Trent, J.O.; Chaires, J.B. Folding and Unfolding Pathways of the Human Telomeric G-Quadruplex. J. Mol. Biol. 2014, 426, 1629–1650. [Google Scholar] [CrossRef] [PubMed]
- Criscuolo, A.; Napolitano, E.; Riccardi, C.; Musumeci, D.; Platella, C.; Montesarchio, D. Insights into the Small Molecule Targeting of Biologically Relevant G-Quadruplexes: An Overview of NMR and Crystal Structures. Pharmaceutics 2022, 14, 2361. [Google Scholar] [CrossRef]
- Brcic, J.; Plavec, J. NMR Structure of a G-Quadruplex Formed by Four d(G4C2) Repeats: Insights into Structural Polymorphism. Nucleic Acids Res. 2018, 46, 11605–11617. [Google Scholar] [PubMed]
- Reddy Sannapureddi, R.K.; Mohanty, M.K.; Gautam, A.K.; Sathyamoorthy, B. Characterization of DNA G-Quadruplex Topologies with NMR Chemical Shifts. J. Phys. Chem. Lett. 2020, 11, 10016–10022. [Google Scholar] [CrossRef] [PubMed]
- Grand, C.L.; Han, H.; Muñoz, R.M.; Weitman, S.; Von Hoff, D.D.; Hurley, L.H.; Bearss, D.J. The Cationic Porphyrin TMPyP4 down-Regulates c-MYC and Human Telomerase Reverse Transcriptase Expression and Inhibits Tumor Growth in Vivo. Mol. Cancer Ther. 2002, 1, 565–573. [Google Scholar]
- Libera, V.; Andreeva, E.A.; Martel, A.; Thureau, A.; Longo, M.; Petrillo, C.; Paciaroni, A.; Schirò, G.; Comez, L. Porphyrin Binding and Irradiation Promote G-Quadruplex DNA Dimeric Structure. J. Phys. Chem. Lett. 2021, 12, 8096–8102. [Google Scholar] [CrossRef]
- Maharjan, P.S.; Bhattarai, H.K. Singlet Oxygen, Photodynamic Therapy, and Mechanisms of Cancer Cell Death. J. Oncol. 2022, 2022, 7211485. [Google Scholar] [CrossRef]
- Martino, L.; Pagano, B.; Fotticchia, I.; Neidle, S.; Giancola, C. Shedding Light on the Interaction between TMPyP4 and Human Telomeric Quadruplexes. J. Phys. Chem. B 2009, 113, 14779–14786. [Google Scholar] [CrossRef]
- Beniaminov, A.D.; Novikov, R.A.; Mamaeva, O.K.; Mitkevich, V.A.; Smirnov, I.P.; Livshits, M.A.; Shchyolkina, A.K.; Kaluzhny, D.N. Light-Induced Oxidation of the Telomeric G4 DNA in Complex with Zn(II) Tetracarboxymethyl Porphyrin. Nucleic Acids Res. 2016, 44, 10031–10041. [Google Scholar] [CrossRef]
- Kadish, K.; Smith, K.M.; Guilard, R. The Porphyrin Handbook: Phthalocyanines: Synthesis; Academic Press: Cambridge, MA, USA, 2000; ISBN 9780123932259. [Google Scholar]
- Fleming, A.M.; Burrows, C.J. G-Quadruplex Folds of the Human Telomere Sequence Alter the Site Reactivity and Reaction Pathway of Guanine Oxidation Compared to Duplex DNA. Chem. Res. Toxicol. 2013, 26, 593–607. [Google Scholar] [CrossRef]
- Zhou, J.; Fleming, A.M.; Averill, A.M.; Burrows, C.J.; Wallace, S.S. The NEIL Glycosylases Remove Oxidized Guanine Lesions from Telomeric and Promoter Quadruplex DNA Structures. Nucleic Acids Res. 2015, 43, 4039–4054. [Google Scholar] [CrossRef] [PubMed]
- Oikawa, S.; Tada-Oikawa, S.; Kawanishi, S. Site-Specific DNA Damage at the GGG Sequence by UVA Involves Acceleration of Telomere Shortening. Biochemistry 2001, 40, 4763–4768. [Google Scholar] [CrossRef] [PubMed]
- Gray, R.D.; Buscaglia, R.; Chaires, J.B. Populated Intermediates in the Thermal Unfolding of the Human Telomeric Quadruplex. J. Am. Chem. Soc. 2012, 134, 16834–16844. [Google Scholar] [CrossRef] [PubMed]
- Koirala, D.; Mashimo, T.; Sannohe, Y.; Yu, Z.; Mao, H.; Sugiyama, H. Intramolecular Folding in Three Tandem Guanine Repeats of Human Telomeric DNA. Chem. Comm. 2012, 48, 2006. [Google Scholar] [CrossRef]
- Dai, J.; Carver, M.; Yang, D. Polymorphism of Human Telomeric Quadruplex Structures. Biochimie 2008, 90, 1172–1183. [Google Scholar] [CrossRef]
- Su, D.G.T.; Fang, H.; Gross, M.L.; Taylor, J.-S.A. Photocrosslinking of Human Telomeric G-Quadruplex Loops by anti Cyclobutane Thymine Dimer Formation. Proc. Nat. Acad. Sci. USA 2009, 106, 12861–12866. [Google Scholar] [CrossRef]
- Mashimo, T.; Sannohe, Y.; Yagi, H.; Sugiyama, H. Folding Pathways of Hybrid-1 and Hybrid-2 G-Quadruplex Structures. In NUCLEIC Acids Symposium Series; Oxford University Press: Oxford, UK, 2008; pp. 409–410. [Google Scholar]
- Gray, R.D.; Li, J.; Chaires, J.B. Energetics and Kinetics of a Conformational Switch in G-Quadruplex DNA. J. Phys. Chem. B 2009, 113, 2676–2683. [Google Scholar] [CrossRef]
- Phan, A.T.; Kuryavyi, V.; Luu, K.N.; Patel, D.J. Structure of Two Intramolecular G-Quadruplexes Formed by Natural Human Telomere Sequences in K+ Solution. Nucleic Acids Res. 2007, 35, 6517–6525. [Google Scholar] [CrossRef]
- Gray, R.D.; Chaires, J.B. Analysis of Multidimensional G-Quadruplex Melting Curves. Curr. Protoc. Nucleic Acid Chem. 2011, 45, 17-4. [Google Scholar] [CrossRef]
- Privalov, P.L.; Khechinashvili, N.N. A Thermodynamic Approach to the Problem of Stabilization of Globular Protein Structure: A Calorimetric Study. J. Mol. Biol. 1974, 86, 665–684. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Libera, V.; Ripanti, F.; Petrillo, C.; Sacchetti, F.; Ramos-Soriano, J.; Galan, M.C.; Schirò, G.; Paciaroni, A.; Comez, L. Stability of Human Telomeric G-Quadruplexes Complexed with Photosensitive Ligands and Irradiated with Visible Light. Int. J. Mol. Sci. 2023, 24, 9090. https://doi.org/10.3390/ijms24109090
Libera V, Ripanti F, Petrillo C, Sacchetti F, Ramos-Soriano J, Galan MC, Schirò G, Paciaroni A, Comez L. Stability of Human Telomeric G-Quadruplexes Complexed with Photosensitive Ligands and Irradiated with Visible Light. International Journal of Molecular Sciences. 2023; 24(10):9090. https://doi.org/10.3390/ijms24109090
Chicago/Turabian StyleLibera, Valeria, Francesca Ripanti, Caterina Petrillo, Francesco Sacchetti, Javier Ramos-Soriano, Maria Carmen Galan, Giorgio Schirò, Alessandro Paciaroni, and Lucia Comez. 2023. "Stability of Human Telomeric G-Quadruplexes Complexed with Photosensitive Ligands and Irradiated with Visible Light" International Journal of Molecular Sciences 24, no. 10: 9090. https://doi.org/10.3390/ijms24109090