
Achieving High Expression of Cry in Green Tissues and Negligible Expression in Endosperm Simultaneously via rbcS Gene Fusion Strategy in Rice
 , ,
, ,
Abstract
:1. Introduction
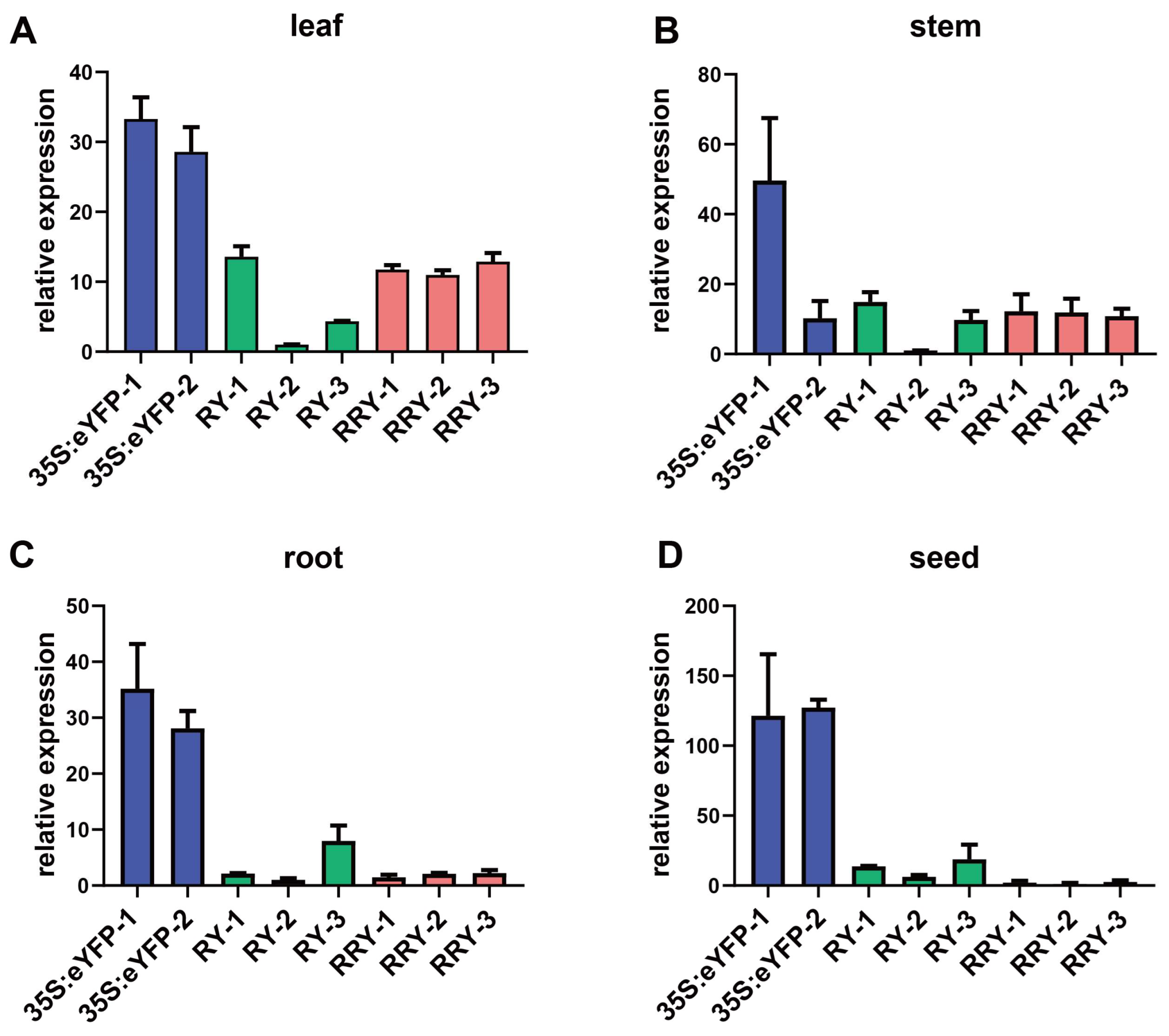
2. Results
2.1. Comparison of OsrbcS Fused and OsrbcS Unfused Transgenic Rice Plants
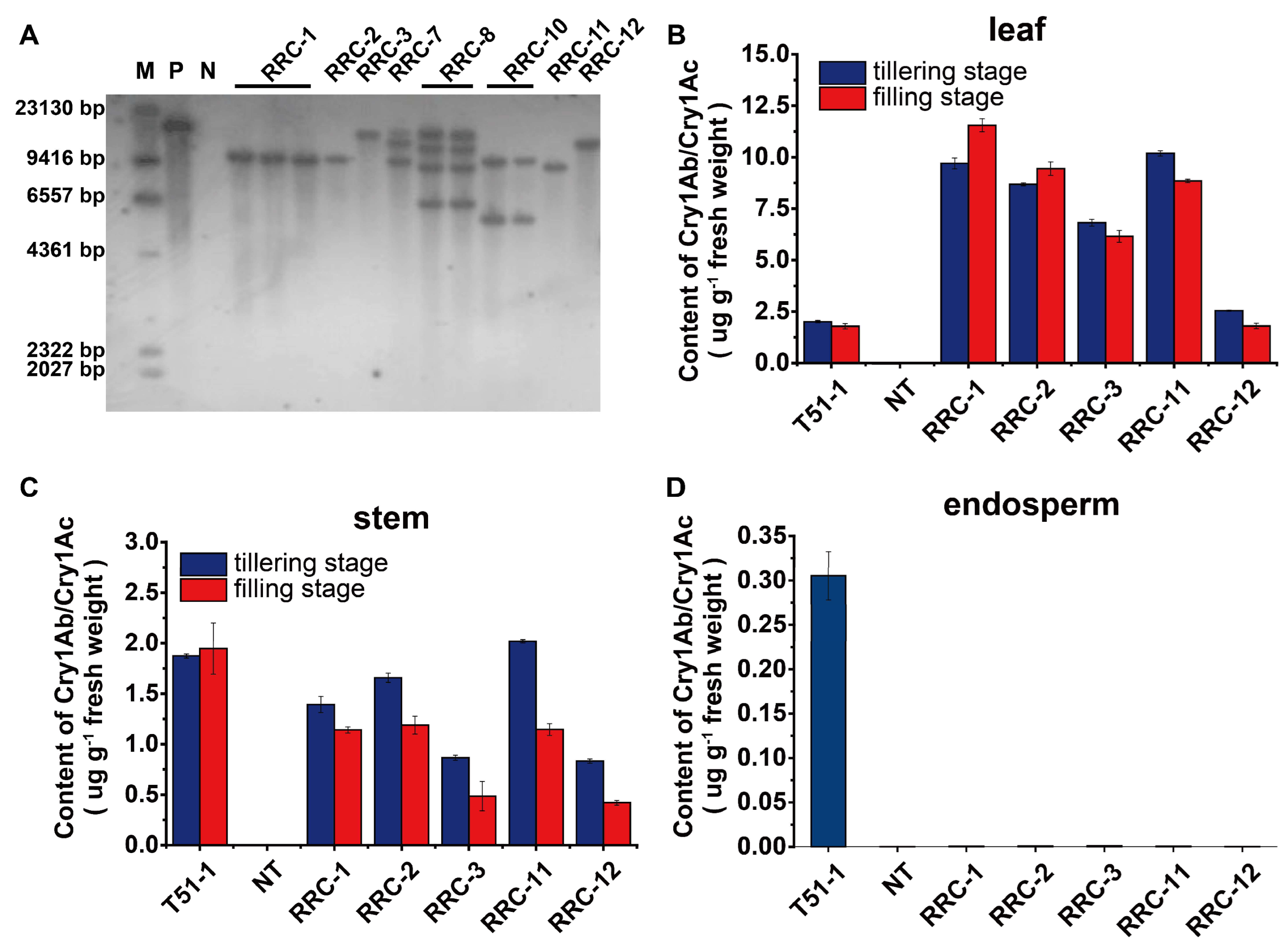
2.2. Generation and Protein Quantification of RRC Lines
2.3. Evaluation of Insect Resistance and Agronomic Performance in the Field
3. Discussion
4. Materials and Methods
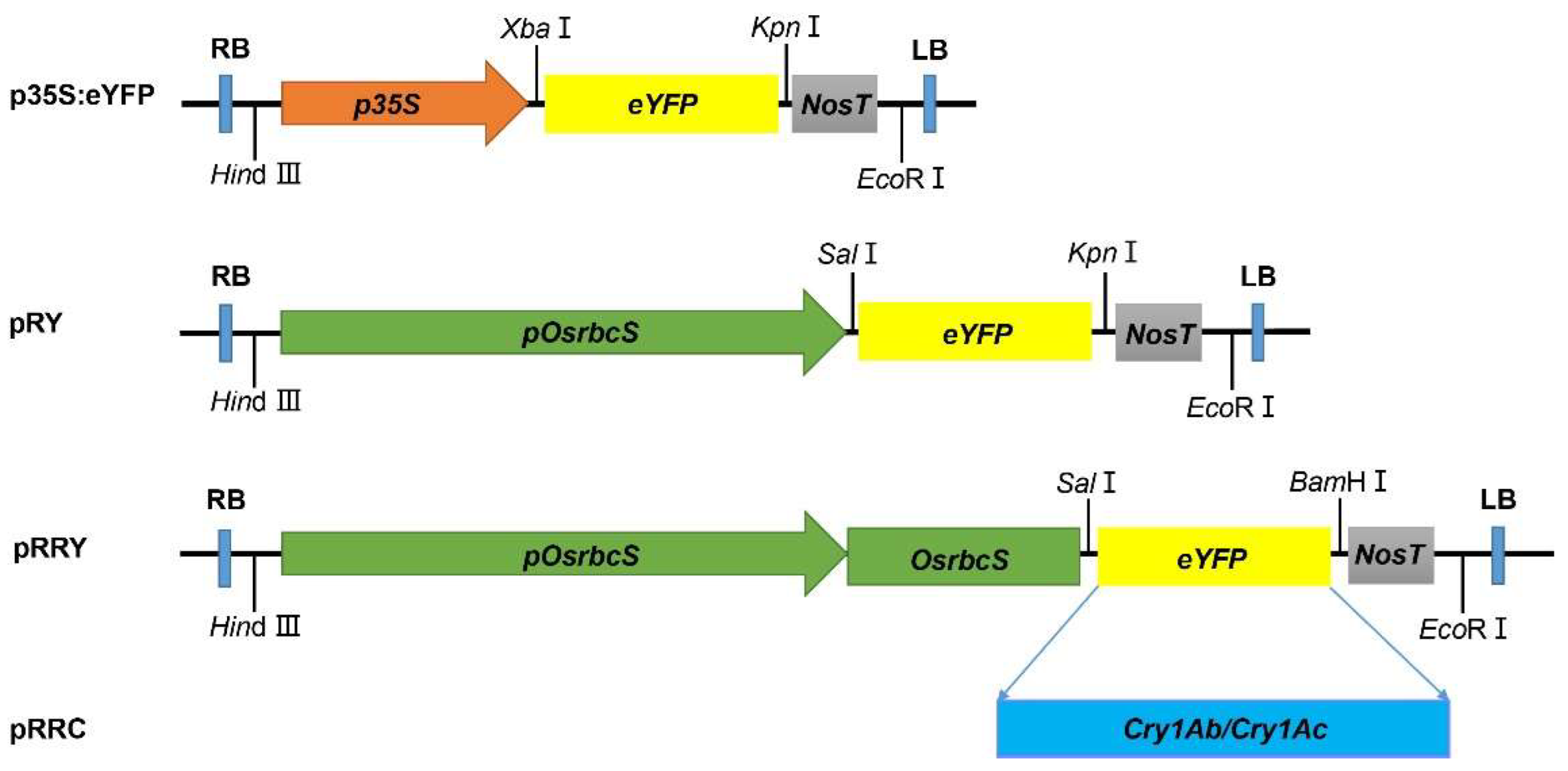
4.1. Plant Materials and Plasmid Construction
4.2. RNA Extraction and qRT-PCR Analysis
4.3. Tissue Localization of pOsrbcS:eYFP and pOsrbcS:OsrbcS-eYFP Line
4.4. Quantitative Estimation of Cry1Ab/Cry1Ac Protein Expressed in Transgenic Plants
4.5. Southern Blot
4.6. Field Tests for Insect Resistance and Agronomic Performance
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fahad, S.; Nie, L.; Hussain, S.; Khan, F.; Khan, F.A.; Saud, S.; Muhammad, H.; Li, L.; Liu, X.; Tabassum, A.; et al. Rice Pest Management and Biological Control. In Sustainable Agriculture Reviews: Cereals; Lichtfouse, E., Goyal, A., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 85–106. [Google Scholar]
- Prasad, R.; Shivay, Y.S.; Kumar, D. Current Status, Challenges, and Opportunities in Rice Production. In Rice Production Worldwide; Chauhan, B.S., Jabran, K., Mahajan, G., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 1–32. [Google Scholar]
- Chen, M.; Shelton, A.; Ye, G.-y. Insect-Resistant Genetically Modified Rice in China: From Research to Commercialization. Annu. Rev. Entomol. 2011, 56, 81–101. [Google Scholar] [CrossRef]
- Pigott, C.R.; Ellar, D.J. Role of Receptors in Bacillus thuringiensis Crystal Toxin Activity. Microbiol. Mol. Biol. Rev. 2007, 71, 255–281. [Google Scholar] [CrossRef]
- Guerchicoff, A.; Delécluse, A.; Rubinstein, C.P. The Bacillus thuringiensis cyt genes for hemolytic endotoxins constitute a gene family. Appl. Environ. Microbiol. 2001, 67, 1090–1096. [Google Scholar] [CrossRef]
- Gupta, M.; Kumar, H.; Kaur, S. Vegetative Insecticidal Protein (Vip): A Potential Contender from Bacillus thuringiensis for Efficient Management of Various Detrimental Agricultural Pests. Front. Microbiol. 2021, 12, 659736. [Google Scholar] [CrossRef] [PubMed]
- Donovan, W.P.; Engleman, J.T.; Donovan, J.C.; Baum, J.A.; Bunkers, G.J.; Chi, D.J.; Clinton, W.P.; English, L.; Heck, G.R.; Ilagan, O.M.; et al. Discovery and characterization of Sip1A: A novel secreted protein from Bacillus thuringiensis with activity against coleopteran larvae. Appl. Microbiol. Biotechnol. 2006, 72, 713–719. [Google Scholar] [CrossRef]
- Fabrick, J.A.; Li, X.; Carrière, Y.; Tabashnik, B.E. Molecular Genetic Basis of Lab- and Field-Selected Bt Resistance in Pink Bollworm. Insects 2023, 14, 201. [Google Scholar] [CrossRef]
- Brookes, G. Farm income and production impacts from the use of genetically modified (GM) crop technology 1996–2020. GM Crops Food 2022, 13, 171–195. [Google Scholar] [CrossRef] [PubMed]
- Cui, K.; Shoemaker, S.P. Public perception of genetically-modified (GM) food: A Nationwide Chinese Consumer Study. npj Sci. Food 2018, 2, 10. [Google Scholar] [CrossRef]
- Tu, J.; Zhang, G.; Datta, K.; Xu, C.; He, Y.; Zhang, Q.; Khush, G.S.; Datta, S.K. Field performance of transgenic elite commercial hybrid rice expressing Bacillus thuringiensis delta-endotoxin. Nat. Biotechnol. 2000, 18, 1101–1104. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Hallerman, E.M.; Peng, Y. Excessive Chinese concerns over genetically engineered food safety are unjustified. Nat. Plants 2020, 6, 590. [Google Scholar] [CrossRef] [PubMed]
- Fan, B.L.; Tang, X.Q.; Tian, H.; Sun, F.Z.; Liu, J.F. Chronic Toxicity of Huahui No. 1 Rice Toward Rats. Sci. Sin. 2014, 44, 911–919. [Google Scholar]
- Tabashnik, B.E.; Carrière, Y. Surge in insect resistance to transgenic crops and prospects for sustainability. Nat. Biotechnol. 2017, 35, 926–935. [Google Scholar] [CrossRef] [PubMed]
- Tabashnik, B.E.; Fabrick, J.A.; Carrière, Y. Global Patterns of Insect Resistance to Transgenic Bt Crops: The First 25 Years. J. Econ. Entomol. 2023, 116, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Bates, S.L.; Zhao, J.-Z.; Roush, R.T.; Shelton, A.M. Insect resistance management in GM crops: Past, present and future. Nat. Biotechnol. 2005, 23, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Tabashnik, B.E.; Brévault, T.; Carrière, Y. Insect resistance to Bt crops: Lessons from the first billion acres. Nat. Biotechnol. 2013, 31, 510–521. [Google Scholar] [CrossRef]
- Manikandan, R.; Balakrishnan, N.; Sudhakar, D.; Udayasuriyan, V. Development of leaffolder resistant transgenic rice expressing cry2AX1 gene driven by green tissue-specific rbcS promoter. World J. Microbiol. Biotechnol. 2016, 32, 37. [Google Scholar] [CrossRef]
- Feng, Z.; Yuan, M.; Zou, J.; Wu, L.B.; Wei, L.; Chen, T.; Zhou, N.; Xue, W.; Zhang, Y.; Chen, Z.; et al. Development of marker-free rice with stable and high resistance to rice black-streaked dwarf virus disease through RNA interference. Plant Biotechnol. J. 2021, 19, 212–214. [Google Scholar] [CrossRef]
- Foissac, X.; Thi Loc, N.; Christou, P.; Gatehouse, A.M.; Gatehouse, J.A. Resistance to green leafhopper (Nephotettix virescens) and brown planthopper (Nilaparvata lugens) in transgenic rice expressing snowdrop lectin (Galanthus nivalis agglutinin; GNA). J. Insect Physiol. 2000, 46, 573–583. [Google Scholar] [CrossRef]
- Ye, R.; Huang, H.; Yang, Z.; Chen, T.; Liu, L.; Li, X.; Chen, H.; Lin, Y. Development of insect-resistant transgenic rice with Cry1C*-free endosperm. Pest Manag. Sci. 2009, 65, 1015–1020. [Google Scholar] [CrossRef]
- Qi, Y.; Chen, L.; He, X.; Jin, Q.; Zhang, X.; He, Z. Marker-free, tissue-specific expression of Cry1Ab as a safe transgenic strategy for insect resistance in rice plants. Pest Manag. Sci. 2013, 69, 135–141. [Google Scholar] [CrossRef]
- Chen, H.; Luo, J.; Zheng, P.; Zhang, X.; Zhang, C.; Li, X.; Wang, M.; Huang, Y.; Liu, X.; Jan, M.; et al. Application of Cre-lox gene switch to limit the Cry expression in rice green tissues. Sci. Rep. 2017, 7, 14505. [Google Scholar] [CrossRef] [PubMed]
- Desai, P.N.; Shrivastava, N.; Padh, H. Production of heterologous proteins in plants: Strategies for optimal expression. Biotechnol. Adv. 2010, 28, 427–435. [Google Scholar] [CrossRef]
- Viana, J.F.; Dias, S.C.; Franco, O.L.; Lacorte, C. Heterologous production of peptides in plants: Fusion proteins and beyond. Curr. Protein Pept. Sci. 2013, 14, 568–579. [Google Scholar] [CrossRef] [PubMed]
- Takaiwa, F.; Yang, L.; Wakasa, Y.; Ozawa, K. Compensatory rebalancing of rice prolamins by production of recombinant prolamin/bioactive peptide fusion proteins within ER-derived protein bodies. Plant Cell Rep. 2018, 37, 209–223. [Google Scholar] [CrossRef]
- Formighieri, C.; Melis, A. A phycocyanin·phellandrene synthase fusion enhances recombinant protein expression and β-phellandrene (monoterpene) hydrocarbons production in Synechocystis (cyanobacteria). Metab. Eng. 2015, 32, 116–124. [Google Scholar] [CrossRef]
- McElroy, D.; Zhang, W.; Cao, J.; Wu, R. Isolation of an efficient actin promoter for use in rice transformation. Plant Cell 1990, 2, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.X.; Tang, T.; Liu, F.X.; Lu, C.L.; Hu, X.L.; Ji, L.L.; Liu, Q.Q. Unintended Changes in Genetically Modified Rice Expressing the Lysine-Rich Fusion Protein Gene Revealed by a Proteomics Approach. J. Integr. Agric. 2013, 12, 2013–2021. [Google Scholar] [CrossRef]
- Zhou, X.H.; Dong, Y.; Wang, Y.; Xiao, X.; Xu, Y.; Xu, B.; Li, X.; Wei, X.S.; Liu, Q.Q. A three generation study with high-lysine transgenic rice in Sprague–Dawley rats. Food Chem. Toxicol. 2012, 50, 1902–1910. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.H.; Suh, S.C.; Park, B.S.; Shin, K.S.; Kweon, S.J.; Han, E.J.; Park, S.H.; Kim, Y.S.; Kim, J.K. Chloroplast-targeted expression of synthetic cry1Ac in transgenic rice as an alternative strategy for increased pest protection. Planta 2009, 230, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, S.; Chakraborty, A.; Sikdar, N.; Chakraborty, S.; Bhattacharyya, J.; Mitra, J.; Manna, A.; Dutta Gupta, S.; Sen, S.K. Marker-free transgenic rice expressing the vegetative insecticidal protein (Vip) of Bacillus thuringiensis shows broad insecticidal properties. Planta 2016, 244, 789–804. [Google Scholar] [CrossRef]
- Chakraborty, M.; Reddy, P.S.; Mustafa, G.; Rajesh, G.; Narasu, V.M.L.; Udayasuriyan, V.; Rana, D. Transgenic rice expressing the cry2AX1 gene confers resistance to multiple lepidopteran pests. Transgenic Res. 2016, 25, 665–678. [Google Scholar] [CrossRef]
- Yang, Y.Y.; Mei, F.; Zhang, W.; Shen, Z.; Fang, J. Creation of Bt rice expressing a fusion protein of Cry1Ac and Cry1I-like using a green tissue-specific promoter. J. Econ. Entomol. 2014, 107, 1674–1679. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-m.; Chiu, C.-C. Protein Transport into Chloroplasts. Annu. Rev. Plant Biol. 2010, 61, 157–180. [Google Scholar] [CrossRef] [PubMed]
- Daniell, H.; Lin, C.S.; Yu, M.; Chang, W.J. Chloroplast genomes: Diversity, evolution, and applications in genetic engineering. Genome Biol. 2016, 17, 134. [Google Scholar] [CrossRef]
- Lopez-Juez, E.; Pyke, K.A. Plastids unleashed: Their development and their integration in plant development. Int. J. Dev. Biol. 2005, 49, 557–577. [Google Scholar] [CrossRef] [PubMed]
- Pillay, P.; Schlüter, U.; van Wyk, S.; Kunert, K.J.; Vorster, B.J. Proteolysis of recombinant proteins in bioengineered plant cells. Bioengineered 2014, 5, 15–20. [Google Scholar] [CrossRef]
- Miziorko, H.M.; Lorimer, G.H. Ribulose-1,5-bisphosphate carboxylase-oxygenase. Annu. Rev. Biochem. 1983, 52, 507–535. [Google Scholar] [CrossRef]
- Hwang, I.K.H.; Sohn, E.J.; Lee, Y.J.; Berggren, P.-O.; Yoo, Y.J.; Kim, J.Y. Method of Highly Expressing Target Protein from Plants Using RbcS Fusion Protein and Method of Preparing Composition for Oral Adminstration of Medical Protein Using Target Protein Expression Plant Body. U.S. Patent Application No. US2017/0335361 A1, 23 November 2017. [Google Scholar]
- Rawat, P.; Singh, A.K.; Ray, K.; Chaudhary, B.; Kumar, S.; Gautam, T.; Kanoria, S.; Kaur, G.; Kumar, P.; Pental, D.; et al. Detrimental effect of expression of Bt endotoxin Cry1Ac on in vitro regeneration, in vivo growth and development of tobacco and cotton transgenics. J. Biosci. 2011, 36, 363–376. [Google Scholar] [CrossRef]
- Maqbool, S.B.; Husnain, T.; Riazuddin, S.; Masson, L.; Christou, P. Effective control of yellow stem borer and rice leaf folder in transgenic rice indica varieties Basmati 370 and M 7 using the novel δ-endotoxin cry2A Bacillus thuringiensis gene. Mol. Breed. 1998, 4, 501–507. [Google Scholar] [CrossRef]
- Staehelin, L.A. Chloroplast Structure and Supramolecular Organization of Photosynthetic Membranes. In Photosynthesis III: Photosynthetic Membranes and Light Harvesting Systems; Staehelin, L.A., Arntzen, C.J., Eds.; Springer: Berlin/Heidelberg, Germany, 1986; pp. 1–84. [Google Scholar]
- Kirchhoff, H. Chloroplast ultrastructure in plants. New Phytol. 2019, 223, 565–574. [Google Scholar] [CrossRef]
- Khan, M.S.; Maliga, P. Fluorescent antibiotic resistance marker for tracking plastid transformation in higher plants. Nat. Biotechnol. 1999, 17, 910–915. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Kang, K.; Chung, H.; Yoo, S.H.; Xu, X.M.; Lee, S.B.; Cheong, J.J.; Daniell, H.; Kim, M. Plastid transformation in the monocotyledonous cereal crop, rice (Oryza sativa) and transmission of transgenes to their progeny. Mol. Cells 2006, 21, 401–410. [Google Scholar] [PubMed]
- Wang, Y.; Wei, Z.; Xing, S. Stable plastid transformation of rice, a monocot cereal crop. Biochem. Biophys. Res. Commun. 2018, 503, 2376–2379. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, S.; Fomo, D.A.; Cock, J.H.; Gomez, K.A. Routine procedures for growing rice plants in culture solution. Lab. Man. Physiol. Stud. Rice 1976, 61–66. [Google Scholar]
- Hoopes, L.L.M. Nucleic Acid Blotting: Southern and Northern. Curr. Protoc. Essent. Lab. Tech. 2012, 6, 8.2.1–8.2.26. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Line | Damaged by Stem Borers Number of Tillers Affected (%) | Damaged by Leaffolders Number of Damaged Leaves per Plant |
|---|---|---|
| Nip-WT | 68.03 ± 4.27 | 24.00 ± 7.05 |
| RRC-1 | 2.39 ± 2.12 ** | 0.17 ± 0.21 ** |
| RRC-2 | 1.28 ± 1.28 ** | 0.13 ± 0.12 ** |
| RRC-3 | 2.88 ± 2.66 ** | 0.06 ± 0.11 ** |
| RRC-11 | 0.46 ± 0.79 ** | 0.00 ± 0.00 ** |
| RRC-12 | 1.08 ± 1.86 ** | 0.37 ± 0.15 ** |
| Line | Plant Height (cm) | Panicles per Plant | Panicle Length (cm) | Grains per Panicle | Grains per Plant | Seed-Set Rate (%) | 1000-Grain Weight (g) | Yield per Plant (g) |
|---|---|---|---|---|---|---|---|---|
| Nip-WT | 96.83 ± 1.17 | 11.53 ± 0.23 | 17.13 ± 0.62 | 83.43 ± 2.40 | 961.93 ± 14.44 | 87.68 ± 0.76 | 25.75 ± 0.05 | 24.77 ± 0.36 |
| RRC-1 | 96.30 ± 0.52 | 10.93 ± 0.64 | 17.80 ± 1.23 | 87.23 ± 2.78 | 952.6 ± 31.20 | 89.67 ± 0.86 | 25.73 ± 0.13 | 24.5 ± 0.91 |
| RRC-2 | 94.57 ± 1.02 * | 11.87 ± 0.76 | 16.99 ± 0.30 | 81.21 ± 3.06 | 963.27 ± 63.97 | 89.8 ± 0.89 | 25.76 ± 0.30 | 24.79 ± 1.43 |
| RRC-3 | 93.8 ± 0.40 ** | 11.93 ± 1.03 | 17.53 ± 0.69 | 27.93 ± 1.44 ** | 332.33 ± 12.35 ** | 26.59 ± 1.95 ** | 25.81 ± 0.12 | 8.56 ± 0.29 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, H.; Huang, Y.; Ye, M.; Wang, Y.; He, X.; Tu, J. Achieving High Expression of Cry in Green Tissues and Negligible Expression in Endosperm Simultaneously via rbcS Gene Fusion Strategy in Rice. Int. J. Mol. Sci. 2023, 24, 9045. https://doi.org/10.3390/ijms24109045
Chen H, Huang Y, Ye M, Wang Y, He X, Tu J. Achieving High Expression of Cry in Green Tissues and Negligible Expression in Endosperm Simultaneously via rbcS Gene Fusion Strategy in Rice. International Journal of Molecular Sciences. 2023; 24(10):9045. https://doi.org/10.3390/ijms24109045
Chicago/Turabian StyleChen, Hao, Yuqing Huang, Mengnan Ye, Ya Wang, Xiuying He, and Jumin Tu. 2023. "Achieving High Expression of Cry in Green Tissues and Negligible Expression in Endosperm Simultaneously via rbcS Gene Fusion Strategy in Rice" International Journal of Molecular Sciences 24, no. 10: 9045. https://doi.org/10.3390/ijms24109045