


Potential Benefits of Epidermal Growth Factor for Inhibiting Muscle Degrative Markers in Rats with Alcoholic Liver Damage
 and
and
Abstract
:1. Introduction
2. Results
2.1. Food Intake and Efficiency, Ethanol Consumption, and EGF Supplementation
2.2. Final BW, Relative Liver Weight, and Muscle Weight
2.3. Liver Damage
2.3.1. Plasma Aspartate Aminotransferase and Alanine Aminotransferase Activities and Ammonia Concentration
2.3.2. Hepatic TG and TC Levels
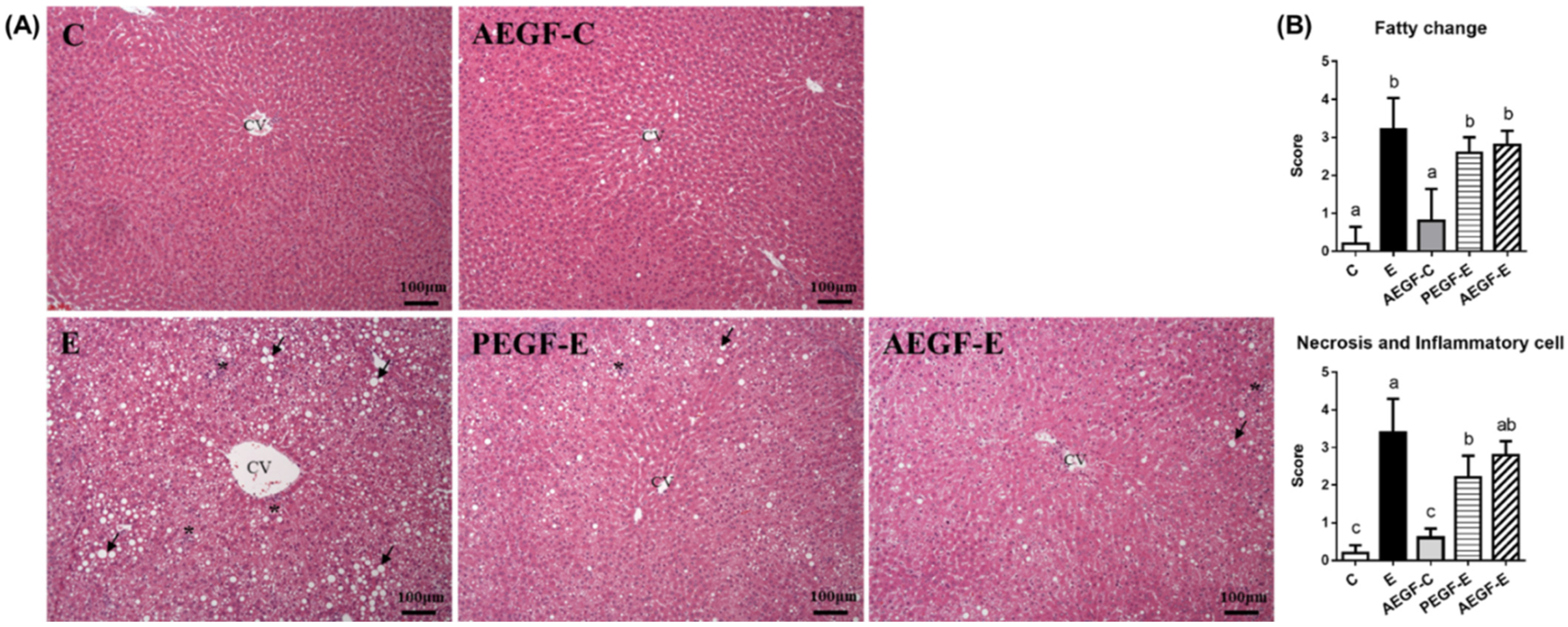
2.3.3. Hepatic Histopathology
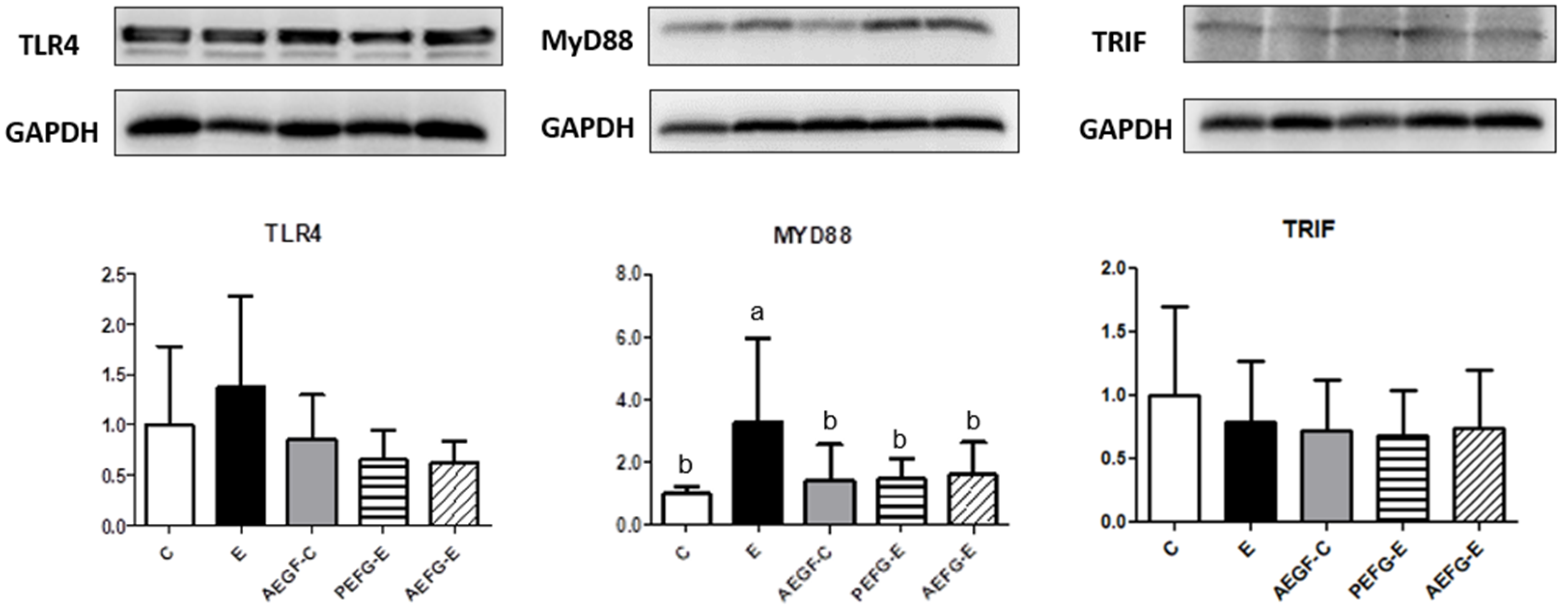
2.3.4. Hepatic Cytokines and TLR4 Signaling Pathway
2.4. Intestinal Injury
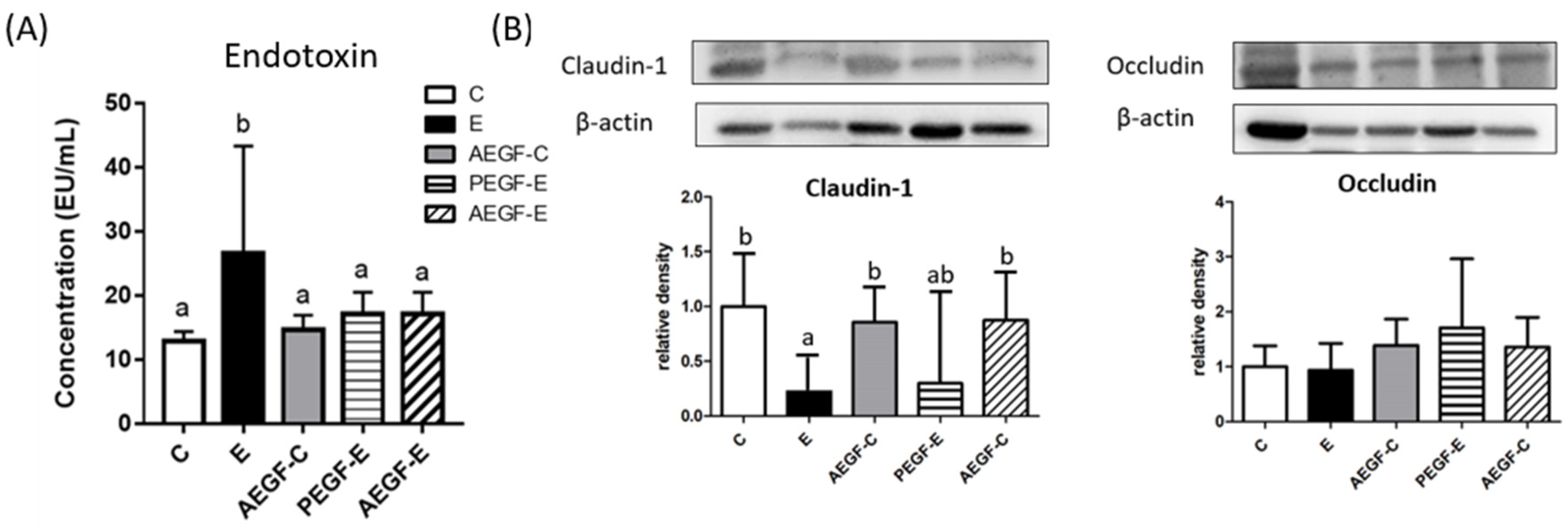
2.4.1. Endotoxin and Tight Junctions
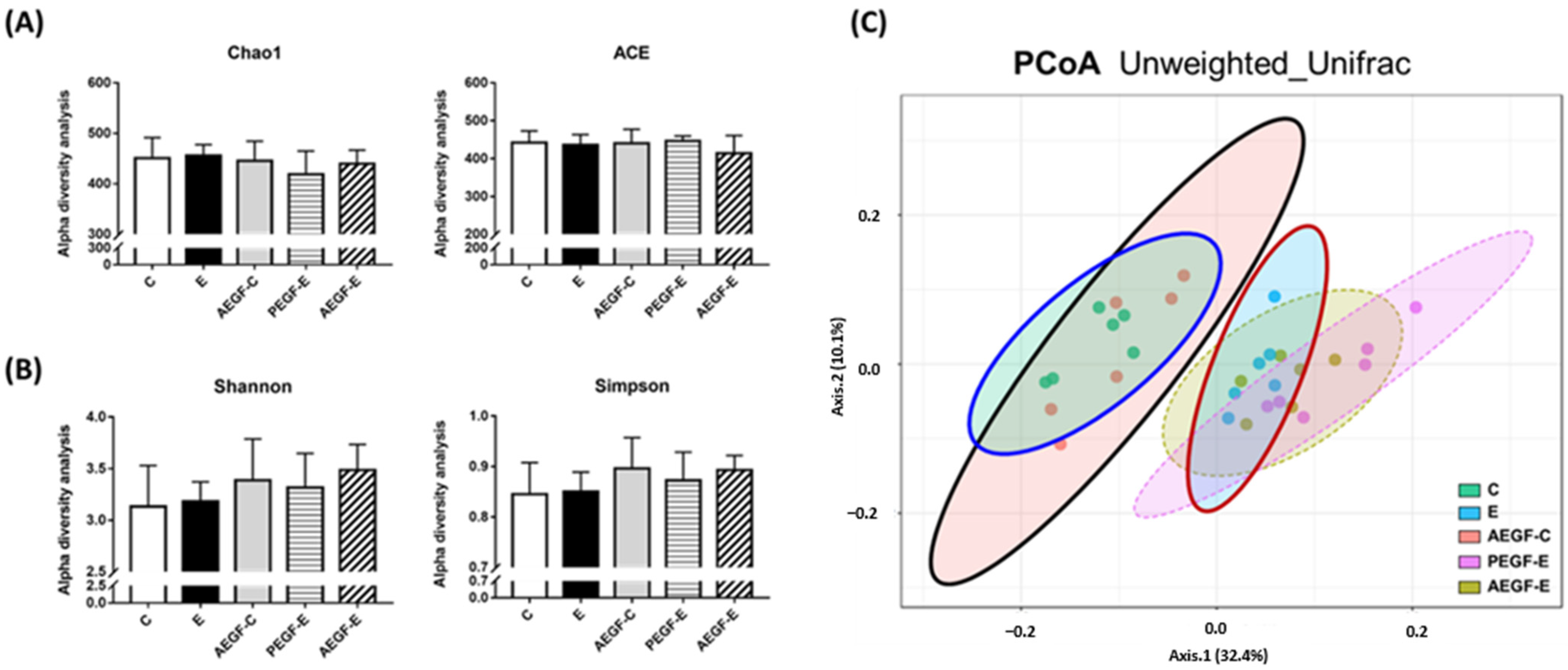
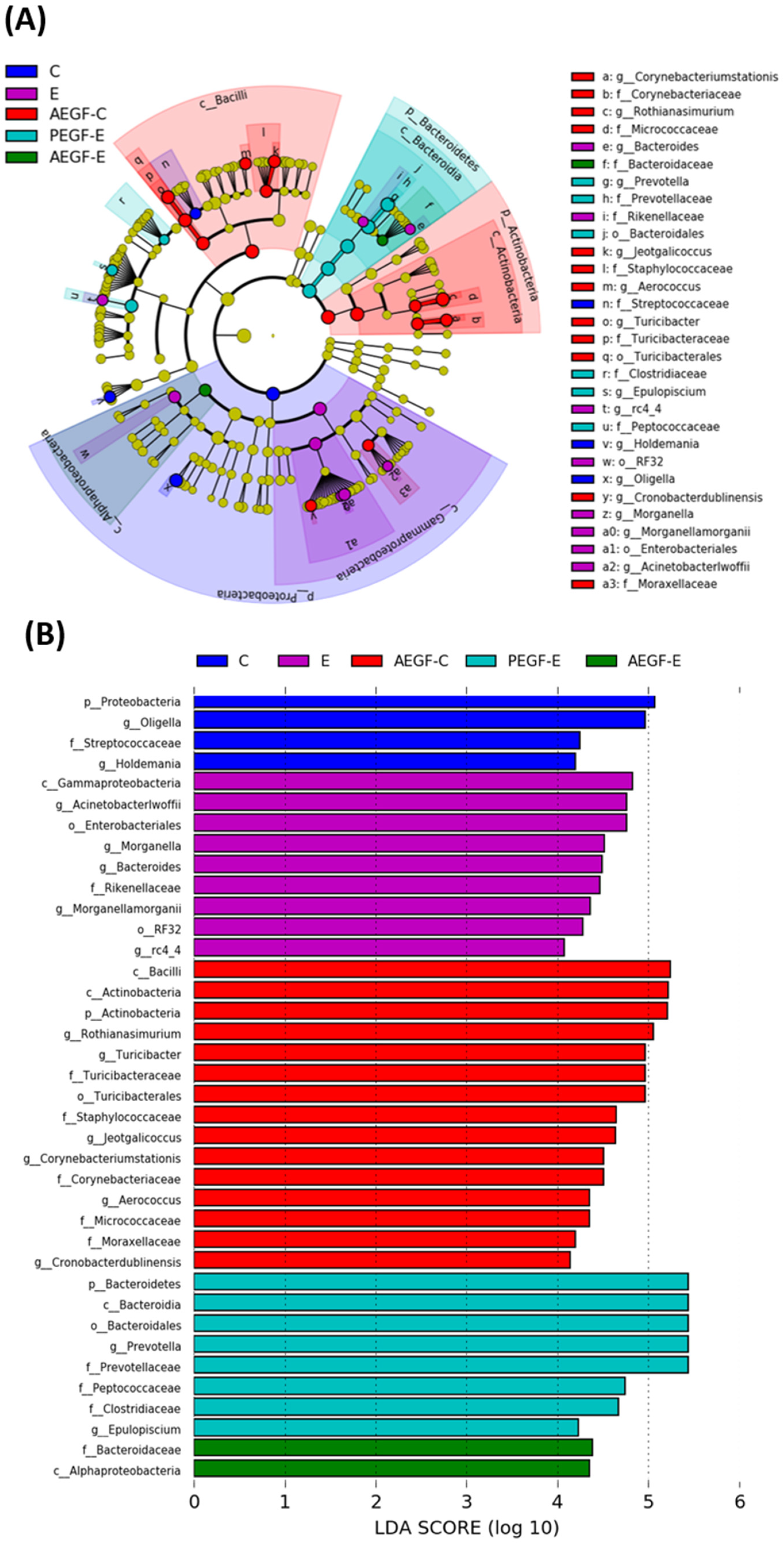
2.4.2. Fecal Microbial Diversity and Composition
Diversity Indices
Linear Discriminant Analysis of the Effect Size
2.5. Muscle Loss
2.5.1. Muscle Mass and Grip Strength
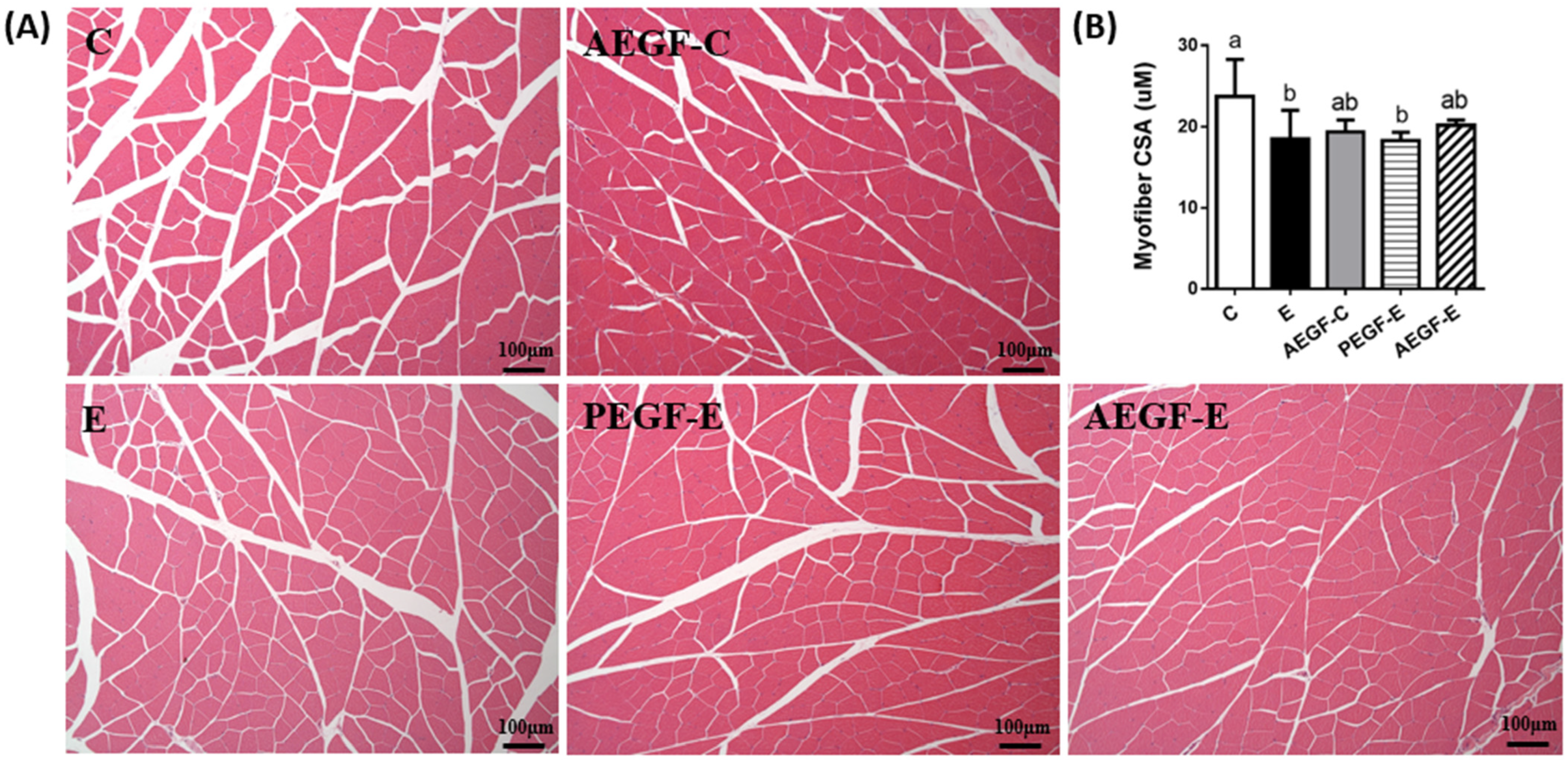
2.5.2. Muscle Histopathology
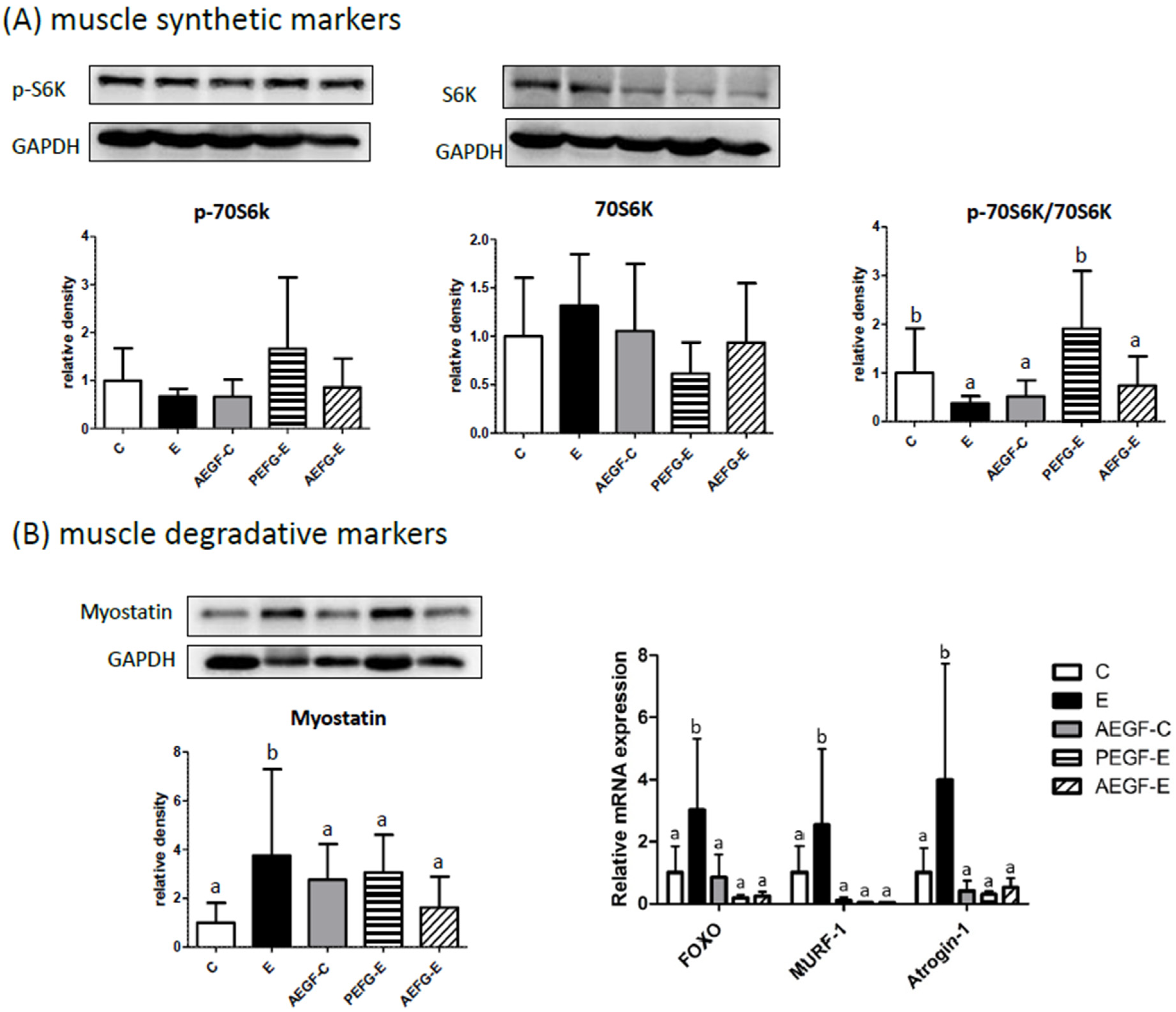
2.5.3. Muscular Protein Metabolism
2.5.4. Amino Acid Profile
3. Discussion
3.1. Intakes of Food, Ethanol, and EGF
3.2. BW and Relative Liver Weight
3.3. Liver Damage and EGF Supplementation
3.4. Intestinal Damage and EGF
3.5. Muscular Protein Metabolism and EGF Supplementation
3.6. Amino Acid Composition
3.7. Study Limitation
4. Materials and Methods
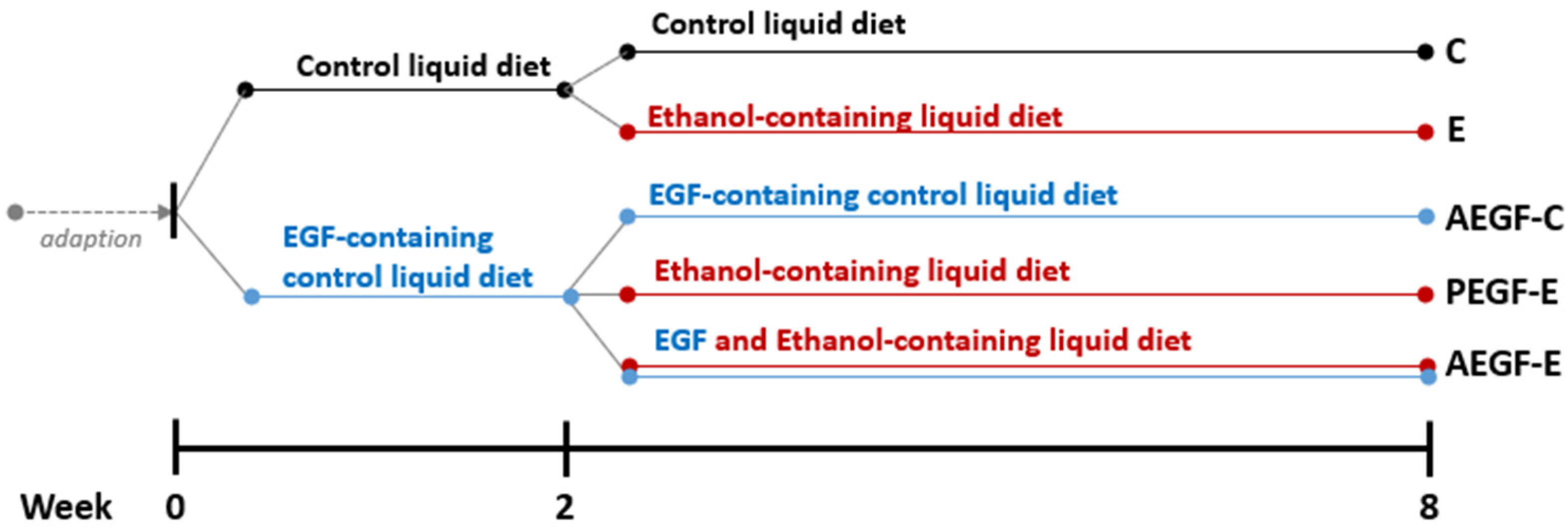
4.1. Animals and Study Protocol
4.2. Assessment of Liver Damage
4.2.1. Liver Function
4.2.2. Liver Histological Assessment
4.2.3. Hepatic Triglyceride (TG) and Total Cholesterol (TC)
4.2.4. Hepatic Cytokines
4.2.5. Hepatic Inflammatory Protein Level
4.3. Intestinal Health Status Examination
4.3.1. Tight Junction Protein Level
4.3.2. Microbiota Analysis
4.4. Muscle Mass and Related Protein Level
4.4.1. Grip Strength
4.4.2. Histological Examination and Muscle CSA
4.4.3. Protein Level of Muscular Protein Synthesis and Degradation Factors
4.4.4. mRNA Expression of Muscular Protein Synthesis and Degradation Factors
4.5. Plasma, Hepatic, and Muscular Amino Acids Profiles
4.6. Chemiclas and Kits
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rehm, J.; Shield, K.D. Global burden of alcohol use disorders and alcohol liver disease. Biomedicines 2019, 7, 99. [Google Scholar] [CrossRef]
- Osna, N.A.; Donohue, T.M., Jr.; Kharbanda, K.K. Alcoholic liver disease: Pathogenesis and current management. Alcohol Res. Curr. Rev. 2017, 38, 147. [Google Scholar]
- Hritz, I.; Mandrekar, P.; Velayudham, A.; Catalano, D.; Dolganiuc, A.; Kodys, K.; Kurt-Jones, E.; Szabo, G. The critical role of toll-like receptor (TLR) 4 in alcoholic liver disease is independent of the common TLR adapter MyD88. Hepatology 2008, 48, 1224–1231. [Google Scholar] [CrossRef]
- Wang, H.J.; Gao, B.; Zakhari, S.; Nagy, L.E. Inflammation in alcoholic liver disease. Annu. Rev. Nutr. 2012, 32, 343. [Google Scholar] [CrossRef]
- Bishehsari, F.; Magno, E.; Swanson, G.; Desai, V.; Voigt, R.M.; Forsyth, C.B.; Keshavarzian, A. Alcohol and gut-derived inflammation. Alcohol Res. Curr. Rev. 2017, 38, 163–171. [Google Scholar]
- Qiu, J.; Thapaliya, S.; Runkana, A.; Yang, Y.; Tsien, C.; Mohan, M.L.; Narayanan, A.; Eghtesad, B.; Mozdziak, P.E.; McDonald, C. Hyperammonemia in cirrhosis induces transcriptional regulation of myostatin by an NF-κB–mediated mechanism. Proc. Natl. Acad. Sci. USA 2013, 110, 18162–18167. [Google Scholar] [CrossRef]
- Gumucio, J.P.; Mendias, C.L. Atrogin-1, MuRF-1, and sarcopenia. Endocrine 2013, 43, 12–21. [Google Scholar] [CrossRef]
- Trendelenburg, A.U.; Meyer, A.; Rohner, D.; Boyle, J.; Hatakeyama, S.; Glass, D.J. Myostatin reduces Akt/TORC1/p70S6K signaling, inhibiting myoblast differentiation and myotube size. Am. J. Physiol. Cell Physiol. 2009, 296, C1258–C1270. [Google Scholar] [CrossRef]
- Dasarathy, J.; McCullough, A.J.; Dasarathy, S. Sarcopenia in alcoholic liver disease: Clinical and molecular advances. Alcohol. Clin. Exp. Res. 2017, 41, 1419–1431. [Google Scholar] [CrossRef]
- Prokopidis, K.; Witard, O.C. Understanding the role of smoking and chronic excess alcohol consumption on reduced caloric intake and the development of sarcopenia. Nutr. Res. Rev. 2021, 35, 197–206. [Google Scholar] [CrossRef]
- Boonstra, J.; Rijken, P.; Humbel, B.; Cremers, F.; Verkleij, A.; van en Bergen Henegouwen, P. The epidermal growth factor. Cell Biol. Int. 1995, 19, 413–430. [Google Scholar] [CrossRef]
- Fitzgerald, K.A.; O’Neill, L.A.; Gearing, A.J.; Callard, R.E. The Cytokine Factsbook and Webfacts; eBook; Elsevier: Amsterdam, The Netherlands, 2001; ISBN 9780080530208. [Google Scholar]
- Tang, X.; Liu, H.; Yang, S.; Li, Z.; Zhong, J.; Fang, R. Epidermal growth factor and intestinal barrier function. Mediat. Inflamm. 2016, 2016, 1927348. [Google Scholar] [CrossRef]
- Garay, C.; Judge, G.; Lucarelli, S.; Bautista, S.; Pandey, R.; Singh, T.; Antonescu, C.N. Epidermal growth factor–stimulated Akt phosphorylation requires clathrin or ErbB2 but not receptor endocytosis. Mol. Biol. Cell 2015, 26, 3504–3519. [Google Scholar] [CrossRef]
- Tang, X.; Liu, X.; Liu, H. Mechanisms of epidermal growth factor effect on animal intestinal phosphate absorption: A review. Front. Vet. Sci. 2021, 8, 670140. [Google Scholar] [CrossRef]
- Lindemans, C.A.; Calafiore, M.; Mertelsmann, A.M.; O′connor, M.H.; Dudakov, J.A.; Jenq, R.R.; Velardi, E.; Young, L.F.; Smith, O.M.; Lawrence, G. Interleukin-22 promotes intestinal-stem-cell-mediated epithelial regeneration. Nature 2015, 528, 560–564. [Google Scholar] [CrossRef]
- Basuroy, S.; Seth, A.; Elias, B.; Naren, A.P.; Rao, R. MAPK interacts with occludin and mediates EGF-induced prevention of tight junction disruption by hydrogen peroxide. Biochem. J. 2006, 393, 69–77. [Google Scholar] [CrossRef]
- Suzuki, T.; Seth, A.; Rao, R. Role of phospholipase Cγ-induced activation of protein kinase Cϵ (PKCϵ) and PKCβI in epidermal growth factor-mediated protection of tight junctions from acetaldehyde in Caco-2 cell monolayers. J. Biol. Chem. 2008, 283, 3574–3583. [Google Scholar] [CrossRef]
- Guntaka, S.R.; Samak, G.; Seth, A.; LaRusso, N.F.; Rao, R. Epidermal growth factor protects the apical junctional complexes from hydrogen peroxide in bile duct epithelium. Lab. Investig. 2011, 91, 1396–1409. [Google Scholar] [CrossRef]
- Chen, Y.L.; Peng, H.C.; Hsieh, Y.C.; Yang, S.C. Epidermal growth factor improved alcohol-induced inflammation in rats. Alcohol 2014, 48, 701–706. [Google Scholar] [CrossRef]
- Crabb, D.W.; Bataller, R.; Chalasani, N.P.; Kamath, P.S.; Lucey, M.; Mathurin, P.; McClain, C.; McCullough, A.; Mitchell, M.C.; Morgan, T.R. Standard definitions and common data elements for clinical trials in patients with alcoholic hepatitis: Recommendation from the NIAAA Alcoholic Hepatitis Consortia. Gastroenterology 2016, 150, 785–790. [Google Scholar] [CrossRef]
- Chen, Y.H.; Chiu, W.C.; Xiao, Q.; Chen, Y.L.; Shirakawa, H.; Yang, S.C. Synbiotics alleviate hepatic damage, intestinal injury and muscular beclin-1 elevation in rats after chronic ethanol administration. Int. J. Mol. Sci. 2021, 22, 12547. [Google Scholar] [CrossRef]
- McClain, C.J.; Rios, C.D.; Condon, S.; Marsano, L.S. Malnutrition and alcohol-associated hepatitis. Clin. Liver Dis. 2021, 25, 557–570. [Google Scholar] [CrossRef]
- Holmuhamedov, E.L.; Czerny, C.; Beeson, C.C.; Lemasters, J.J. Ethanol suppresses ureagenesis in rat hepatocytes: Role of acetaldehyde. J. Nutr. Biochem. 2012, 287, 7692–7700. [Google Scholar]
- Alsahli, M.A.; Almatroodi, S.A.; Almatroudi, A.; Khan, A.A.; Anwar, S.; Almutary, A.G.; Alrumaihi, F.; Rahmani, A.H. 6-gingerol, a major ingredient of ginger attenuates diethylnitrosamine-induced liver injury in rats through the modulation of oxidative stress and anti-inflammatory activity. Mediators. Inflamm. 2021, 2021, 6661937. [Google Scholar] [CrossRef]
- Chen, Y.L.; Shirakawa, H.; Lu, N.S.; Peng, H.C.; Xiao, Q.; Yang, S.C. Impacts of fish oil on the gut microbiota of rats with alcoholic liver damage. J. Nutr. Biochem. 2020, 86, 108491. [Google Scholar] [CrossRef]
- Rao, R. Endotoxemia and gut barrier dysfunction in alcoholic liver disease. Hepatology 2009, 50, 638. [Google Scholar] [CrossRef]
- Piras, V.; Selvarajoo, K. Beyond MyD88 and TRIF pathways in Toll-like receptor signaling. Fronti. Immunol. 2014, 5, 70. [Google Scholar] [CrossRef]
- Tseng, P.-H.; Matsuzawa, A.; Zhang, W.; Mino, T.; Vignali, D.A.; Karin, M. Different modes of ubiquitination of the adaptor TRAF3 selectively activate the expression of type I interferons and proinflammatory cytokines. Nat. Immunol. 2010, 11, 70–75. [Google Scholar] [CrossRef]
- McCarthy, G.M.; Warden, A.S.; Bridges, C.R.; Blednov, Y.A.; Harris, R.A. Chronic ethanol consumption: Role of TLR3/TRIF-dependent signaling. Addict. Biol. 2018, 23, 889–903. [Google Scholar] [CrossRef]
- Anderson, C.; Andersson, T.; Molander, M. Ethanol absorption across human skin measured by in vivo microdialysis tech-nique. Acta Derm. Vener. 1991, 71, 389–393. [Google Scholar]
- Chaudhry, K.K.; Samak, G.; Shukla, P.K.; Mir, H.; Gangwar, R.; Manda, B.; Isse, T.; Kawamoto, T.; Salaspuro, M.; Kaihovaara, P.; et al. ALDH2 deficiency promotes ethanol-induced gut barrier dysfunction and fatty liver in mice. Alcohol. Clin. Exp. Res. 2015, 39, 1465–1475. [Google Scholar] [CrossRef]
- Kim, B.K.; Lee, I.O.; Tan, P.L.; Eor, J.Y.; Hwang, J.K.; Kim, S.H. Protective effect of Lactobacillus fermentum la12 in an alcohol-induced rat model of alcoholic steatohepatitis. Korean J. Food Sci. Anim. Resour. 2017, 37, 931–939. [Google Scholar] [CrossRef]
- Rao, R.K. Acetaldehyde-induced increase in paracellular permeability in Caco-2 cell monolayer. Alcohol. Clin. Exp. Res. 1998, 22, 1724–1730. [Google Scholar] [CrossRef]
- Duh, G.; Mouri, N.; Warburton, D.; Thomas, D.W. EGF regulates early embryonic mouse gut development in chemically defined organ culture. Pediatr. Res. 2000, 48, 794–802. [Google Scholar] [CrossRef]
- Clark, J.A.; Clark, A.T.; Hotchkiss, R.S.; Buchman, T.G.; Coopersmith, C.M. Epidermal growth factor treatment decreases mortality and is associated with improved gut integrity in sepsis. Shock 2008, 30, 36. [Google Scholar]
- Xiao, Q.; Chen, Y.H.; Pratama, S.A.; Chen, Y.L.; Shirakawa, H.; Peng, H.C.; Yang, S.C. The prophylactic effects of glutamine on muscle protein synthesis and degradation in rats with ethanol-induced liver damage. Nutrients 2021, 13, 2788. [Google Scholar] [CrossRef]
- Wang, H.; Yan, Y.; Yi, X.; Duan, Y.; Wang, J.; Li, S.; Luo, L.; Huang, T.; Inglis, B.; Li, X.; et al. Histopathological features and composition of gut microbiota in rhesus monkey of alcoholic liver disease. Front. Microbiol. 2019, 10, 705. [Google Scholar] [CrossRef]
- Hartmann, P.; Seebauer, C.T.; Schnabl, B. Alcoholic liver disease: The gut microbiome and liver cross talk. Alcohol. Clin. Exp. Res. 2015, 39, 763–775. [Google Scholar] [CrossRef]
- Bluemel, S.; Wang, L.; Kuelbs, C.; Moncera, K.; Torralba, M.; Singh, H.; Fouts, D.E.; Schnabl, B. Intestinal and hepatic microbiota changes associated with chronic ethanol administration in mice. Gut Microbes 2020, 11, 265–275. [Google Scholar] [CrossRef]
- Michail, S.; Lin, M.; Frey, M.R.; Fanter, R.; Paliy, O.; Hilbush, B.; Reo, N.V. Altered gut microbial energy and metabolism in children with non-alcoholic fatty liver disease. FEMS Microbiol. Ecol. 2015, 91, 1. [Google Scholar] [CrossRef]
- Pedersen, C.; Ijaz, U.Z.; Gallagher, E.; Horton, F.; Ellis, R.J.; Jaiyeola, E.; Duparc, T.; Russell-Jones, D.; Hinton, P.; Cani, P.D. Fecal Enterobacteriales enrichment is associated with increased in vivo intestinal permeability in humans. Physiol. Rep. 2018, 6, e13649. [Google Scholar] [CrossRef] [PubMed]
- Castro-Mejia, J.; Jakesevic, M.; Krych, Ł.; Nielsen, D.S.; Hansen, L.H.; Sondergaard, B.C.; Kvist, P.H.; Hansen, A.K.; Holm, T.L. Treatment with a monoclonal anti-IL-12p40 antibody induces substantial gut microbiota changes in an experimental colitis model. Gastroenterol. Res. Pract. 2016, 2016, 4953120. [Google Scholar] [CrossRef] [PubMed]
- Noda, M.; Maruyama, M.; Danshiitsoodol, N.; Higashikawa, F.; Sugiyama, M. Improvement of alcohol-poisoning symptoms in mice by the oral administration of live Lactobacillus plantarum SN13T cells. Int. J. Mol. Sci. 2020, 21, 1896. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Chen, S.; Fu, Y.; Wu, W.; Chen, T.; Chen, J.; Yang, B.; Ou, Q. Gut microbiota dysbiosis in patients with hepatitis B virus–induced chronic liver disease covering chronic hepatitis, liver cirrhosis and hepatocellular carcinoma. J. Viral Hepat. 2020, 27, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Pailhoriès, H.; Boursier, J.; Diehl, A.M. Determinants of the Severity of Fatty Liver Diseases: Need all the Pieces to Solve the Puzzle; Wiley Online Library: Hoboken, NJ, USA, 2022; Volume 75, pp. 782–784. [Google Scholar]
- Mohanty, A.; Kabi, A.; Mohanty, A. Acinetobacterlwoffii-Emerging pathogen causing liver abscess: A Case Report. Natl. J. Integr. Res. Med. 2018, 9, 53–54. [Google Scholar]
- Regalado, N.G.; Martin, G.; Antony, S.J. Acinetobacter lwoffii: Bacteremia associated with acute gastroenteritis. Travel Med. Infect. Dis. 2009, 7, 316–317. [Google Scholar] [CrossRef] [PubMed]
- Takakura, W.; Pimentel, M. Small intestinal bacterial overgrowth and irritable bowel syndrome–an update. Front. Psychiatry 2020, 11, 664. [Google Scholar] [CrossRef]
- Jiao, N.; Baker, S.S.; Nugent, C.A.; Tsompana, M.; Cai, L.; Wang, Y.; Buck, M.J.; Genco, R.J.; Baker, R.D.; Zhu, R. Gut microbiome may contribute to insulin resistance and systemic inflammation in obese rodents: A meta-analysis. Physiol. Genom. 2018, 50, 244–254. [Google Scholar] [CrossRef]
- Posteraro, B.; Paroni Sterbini, F.; Petito, V.; Rocca, S.; Cubeddu, T.; Graziani, C.; Arena, V.; Vassallo, G.A.; Mosoni, C.; Lopetuso, L.; et al. Liver injury, endotoxemia, and their relationship to intestinal microbiota composition in alcohol-preferring rats. Alcohol. Clin. Exp. Res. 2018, 42, 2313–2325. [Google Scholar] [CrossRef]
- Bermingham, E.N.; Maclean, P.; Thomas, D.G.; Cave, N.J.; Young, W. Key bacterial families (Clostridiaceae, Erysipelotrichaceae and Bacteroidaceae) are related to the digestion of protein and energy in dogs. Peer J. 2017, 5, e3019. [Google Scholar] [CrossRef]
- Posey, A.D., Jr.; Demonbreun, A.; McNally, E.M. Ferlin proteins in myoblast fusion and muscle growth. Curr. Top. Dev. Biol. 2011, 96, 203–230. [Google Scholar] [CrossRef]
- Shishkin, S.S.; Krokhina, T.B.; Akhunov, V.S.; Makarov, A.A.; Popov, V.O. Effects of myostatin and other growth factors on cultured human cells. Prikl. Biokhim. Mikrobiol. 2004, 40, 630–633. [Google Scholar] [PubMed]
- Song, D.S.; Chang, U.I.; Choi, S.; Jung, Y.D.; Han, K.; Ko, S.H.; Ahn, Y.B.; Yang, J.M. Heavy alcohol consumption with alcoholic liver disease accelerates sarcopenia in elderly korean males: The Korean national health and nutrition examination survey 2008–2010. PLoS ONE 2016, 11, e0163222. [Google Scholar] [CrossRef] [PubMed]
- Thapaliya, S.; Runkana, A.; McMullen, M.R.; Nagy, L.E.; McDonald, C.; Naga Prasad, S.V.; Dasarathy, S. Alcohol-induced autophagy contributes to loss in skeletal muscle mass. Autophagy 2014, 10, 677–690. [Google Scholar] [CrossRef]
- Lang, C.H.; Frost, R.A.; Svanberg, E.; Vary, T.C. IGF-I/IGFBP-3 ameliorates alterations in protein synthesis, eIF4E availability, and myostatin in alcohol-fed rats. Am. J. Physiol. Endocrinol. Metab. 2004, 286, E916–E926. [Google Scholar] [CrossRef] [PubMed]
- Korzick, D.H.; Sharda, D.R.; Pruznak, A.M.; Lang, C.H. Aging accentuates alcohol-induced decrease in protein synthesis in gastrocnemius. Am. J. Physiol. Regul. Integr. Compa. Physiol. 2013, 304, R887–R898. [Google Scholar] [CrossRef]
- Przewłócka, K.; Folwarski, M.; Kaźmierczak-Siedlecka, K.; Skonieczna-Żydecka, K.; Kaczor, J.J. Gut-muscle axis exists and may affect skeletal muscle adaptation to training. Nutrients 2020, 12, 1451. [Google Scholar] [CrossRef]
- Kant, S.; Davuluri, G.; Alchirazi, K.A.; Welch, N.; Heit, C.; Kumar, A.; Gangadhariah, M.; Kim, A.; McMullen, M.R.; Willard, B.; et al. Ethanol sensitizes skeletal muscle to ammo- nia-induced molecular perturbations. J. Biol. Chem. 2019, 294, 7231–7244. [Google Scholar] [CrossRef]
- Jindal, A.; Jagdish, R.K. Sarcopenia: Ammonia metabolism and hepatic encephalopathy. Clin. Mol. Hepatol. 2019, 25, 270–279. [Google Scholar] [CrossRef]
- Bazgir, B.; Fathi, R.; Rezazadeh Valojerdi, M.; Mozdziak, P.; Asgari, A. Satellite cells contribution to exercise mediated muscle hypertrophy and repair. Cell J. 2017, 18, 473–484. [Google Scholar] [CrossRef]
- Pascual-Fernández, J.; Fernández-Montero, A.; Córdova-Martínez, A.; Pastor, D.; Martínez-Rodríguez, A.; Roche, E. Sarcopenia: Molecular pathways and potential targets for intervention. Int. J. Mol. Sci. 2020, 21, 8844. [Google Scholar] [CrossRef]
- Navolanic, P.M.; Steelman, L.S.; McCubrey, J.A. EGFR family signaling and its association with breast cancer development and resistance to chemotherapy (Review). Int. J. Oncol. 2003, 22, 237–252. [Google Scholar] [CrossRef]
- Vuorela, P.; Sarkola, T.; Alfthan, H.; Halmesmäki, E. Hepatocyte growth factor, epidermal growth factor, and placenta growth factor concentrations in peripheral blood of pregnant women with alcohol abuse. Alcohol Clin. Exp. Res. 2002, 26, 682–687. [Google Scholar] [CrossRef]
- Shaw, S.; Lieber, C.S. Plasma amino acid abnormalities in the alcoholic: Respective role of alcohol, nutrition, and liver injury. Gastroenterology 1978, 74, 677–682. [Google Scholar] [CrossRef] [PubMed]
- Dam, G.; Sørensen, M.; Buhl, M.; Sandahl, T.D.; Møller, N.; Ott, P.; Vilstrup, H. Muscle metabolism and whole blood amino acid profile in patients with liver disease. Scand. J. Clin. Lab. 2015, 75, 674–680. [Google Scholar]
- Shepard, B.D.; Tuma, P.L. Alcohol-induced protein hyperacetylation: Mechanisms and consequences. World J. Gastroenterol. 2009, 15, 1219–1230. [Google Scholar] [CrossRef]
- Wang, H.Y.; Peng, H.C.; Chien, Y.W.; Chen, Y.L.; Lu, N.S.; Yang, S.C. Effects of fish oil on lipid metabolism and its molecular biological regulators in chronic ethanol-fed rats. Nutrients 2018, 10, 802. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Breitkopf, S.B.; Yang, X.; Asara, J.M. A positive/negative ion-switching, targeted mass spectrometry-based metabolomics platform for bodily fluids, cells, and fresh and fixed tissue. Nat. Protoc. 2012, 7, 872–881. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Food Intake (g/100 g BW/Day) | Ethanol Intake (g/100 g BW/Day) | Food Efficiency (%) |
|---|---|---|---|
| C | 23.7 ± 0.7 | - | 4.4 ± 0.4 b |
| E | 24.7 ± 0.7 | 1.14 ± 0.04 | 3.2 ± 0.6 a |
| AEGF-C | 23.4 ± 1.1 | - | 4.9 ± 0.6 b |
| PEGF-E | 24.1 ± 0.5 | 1.10 ± 0.04 | 3.1 ± 0.4 a |
| AEGF-E | 23.9 ± 0.6 | 1.09 ± 0.03 | 3.5 ± 0.5 a |
| Groups | Final Body Weight (g) | Liver Weight (g) | Relative Liver Weight (%) |
|---|---|---|---|
| C | 422.0 ± 28.4 b | 10.4 ± 0.8 a | 2.5 ± 0.0 a |
| E | 375.8 ± 22.1 a | 13.2 ± 1.5 c | 3.5 ± 0.5 c |
| AEGF-C | 434.5 ± 25.1 b | 10.8 ± 0.9 a | 2.5 ± 0.1 a |
| PEGF-E | 364.3 ± 17.1 a | 11.3 ± 0.9 ab | 3.1 ± 0.2 b |
| AEGF-E | 367.8 ± 32.1 a | 12.7 ± 1.6 bc | 3.5 ± 0.4 c |
| Groups | AST (U/L) | ALT (U/L) | Ammonia (μg/dL) | TG mg/g Liver | TC mg/g Liver |
|---|---|---|---|---|---|
| C | 78 ± 2 a | 38 ± 1 a | 104 ± 10 b | 49.2 ± 6.5 c | 3.01 ± 0.19 b |
| E | 298 ± 93 b | 180 ± 83 b | 175 ± 40 c | 99.9 ± 8.2 a | 4.31 ± 0.30 a |
| AEGF-C | 75 ± 2 a | 38 ± 2 a | 63 ± 7 ab | 65.2 ± 7.6 bc | 3.49 ± 0.33 ab |
| PEGF-E | 141 ± 11 a | 97 ± 7 ab | 24 ± 8 a | 77.4 ± 10.3 ab | 3.47 ± 0.20 ab |
| AEGF-E | 164 ± 20 a | 108 ± 10 ab | 34 ± 10 a | 68.2 ± 8.2 bc | 3.66 ± 0.31 ab |
| Groups | TNF-α (pg/mg Protein) | IL-1β (pg/mg Protein) | IL-6 (pg/mg Protein) | IL-10 (pg/mg Protein) |
|---|---|---|---|---|
| C | 16.39 ± 2.86 | 70.40 ± 13.36 ab | 175.29 ± 35.56 b | 85.86 ± 21.64 b |
| E | 18.89 ± 2.64 | 101.21 ± 20.89 c | 186.39 ± 35.43 b | 110.16 ± 29.05 ab |
| AEGF-C | 19.25 ± 4.40 | 54.51 ± 15.82 a | 114.18 ± 27.65 a | 69.28 ± 20.11 a |
| PEGF-E | 18.06 ± 4.50 | 58.07 ± 15.60 a | 87.17 ± 32.12 a | 77.69 ± 21.41 a |
| AEGF-E | 16.57 ± 4.02 | 80.54 ± 15.03 b | 76.84 ± 17.57 a | 86.80 ± 12.05 ab |
| Groups | Quadriceps Weight (g) | Gastrocnemius (g) | Grip Strength (g) |
|---|---|---|---|
| C | 4.4 ± 1.6 | 6.9 ± 2.1 | 1673.31 ± 75.54 c |
| E | 5.9 ± 1.5 | 5.1 ± 1.2 | 1350.37 ± 150.88 a |
| AEGF-C | 6.8 ± 1.6 | 5.6 ± 0.6 | 1562.29 ± 69.73 bc |
| PEGF-E | 5.4 ± 1.1 | 4.9 ± 0.4 | 1572.16 ± 109.75 bc |
| AEGF-E | 5.7 ± 1.5 | 5.2 ± 1.1 | 1484.23 ± 105.29 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, Q.; Chen, Y.-H.; Chen, Y.-L.; Chien, Y.-S.; Hsieh, L.-H.; Shirakawa, H.; Yang, S.-C. Potential Benefits of Epidermal Growth Factor for Inhibiting Muscle Degrative Markers in Rats with Alcoholic Liver Damage. Int. J. Mol. Sci. 2023, 24, 8845. https://doi.org/10.3390/ijms24108845
Xiao Q, Chen Y-H, Chen Y-L, Chien Y-S, Hsieh L-H, Shirakawa H, Yang S-C. Potential Benefits of Epidermal Growth Factor for Inhibiting Muscle Degrative Markers in Rats with Alcoholic Liver Damage. International Journal of Molecular Sciences. 2023; 24(10):8845. https://doi.org/10.3390/ijms24108845
Chicago/Turabian StyleXiao, Qian, Yi-Hsiu Chen, Ya-Ling Chen, Yu-Shan Chien, Li-Hsuan Hsieh, Hitoshi Shirakawa, and Suh-Ching Yang. 2023. "Potential Benefits of Epidermal Growth Factor for Inhibiting Muscle Degrative Markers in Rats with Alcoholic Liver Damage" International Journal of Molecular Sciences 24, no. 10: 8845. https://doi.org/10.3390/ijms24108845