
Potential Function of Testicular MicroRNAs in Heat-Stress-Induced Spermatogenesis Disorders
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Treatment
2.2. Thermal Imaging and Semen Quality Analysis
2.3. Data Analysis
2.4. Target Gene Prediction and miRNA mRNA Network Construction
2.5. Cell Culture
2.6. RT-q PCR
2.7. Statistical Analysis
3. Results
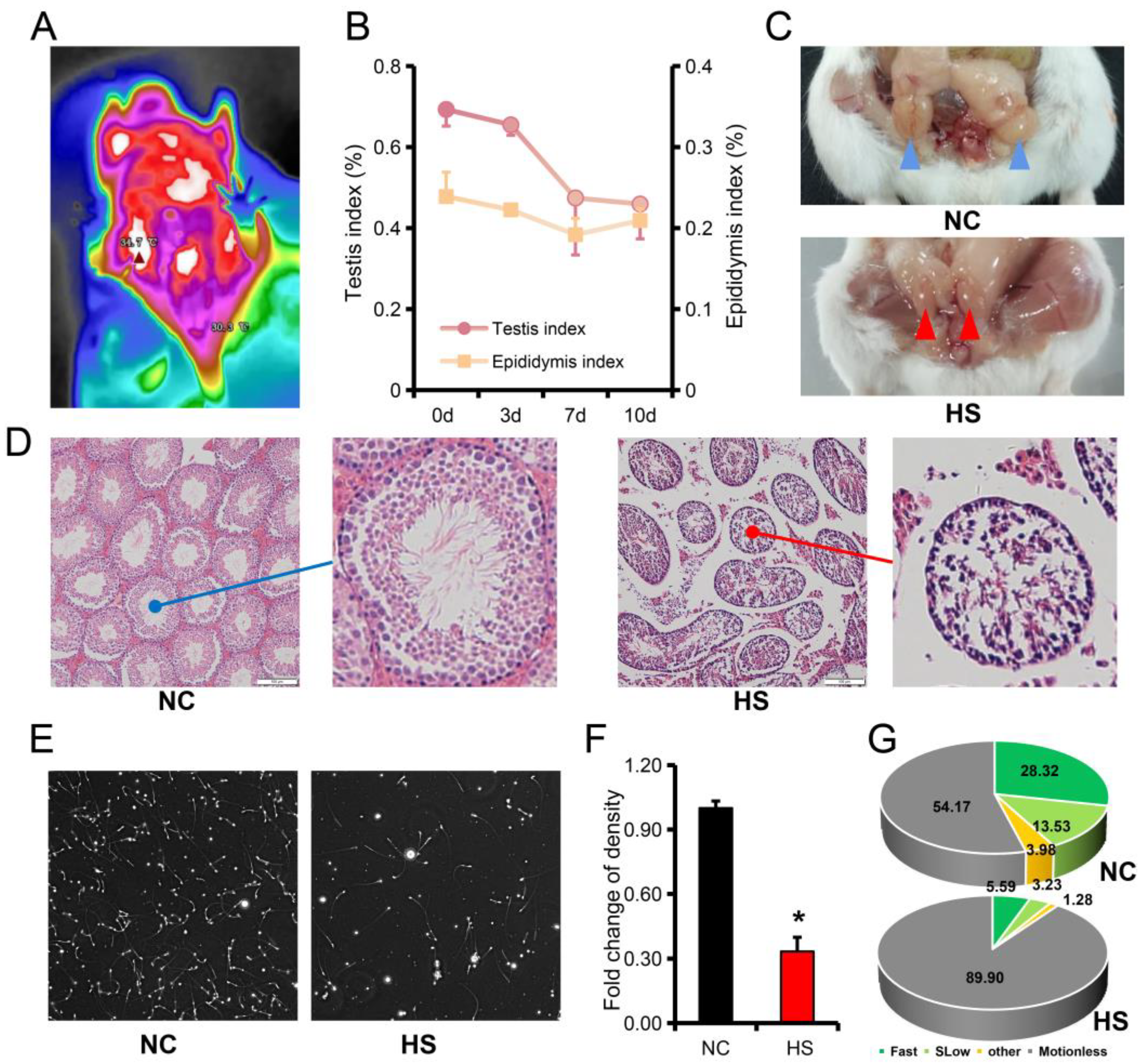
3.1. Testicular Heat Stress Leads to Testicular Atrophy and Reduced Spermatogenesis
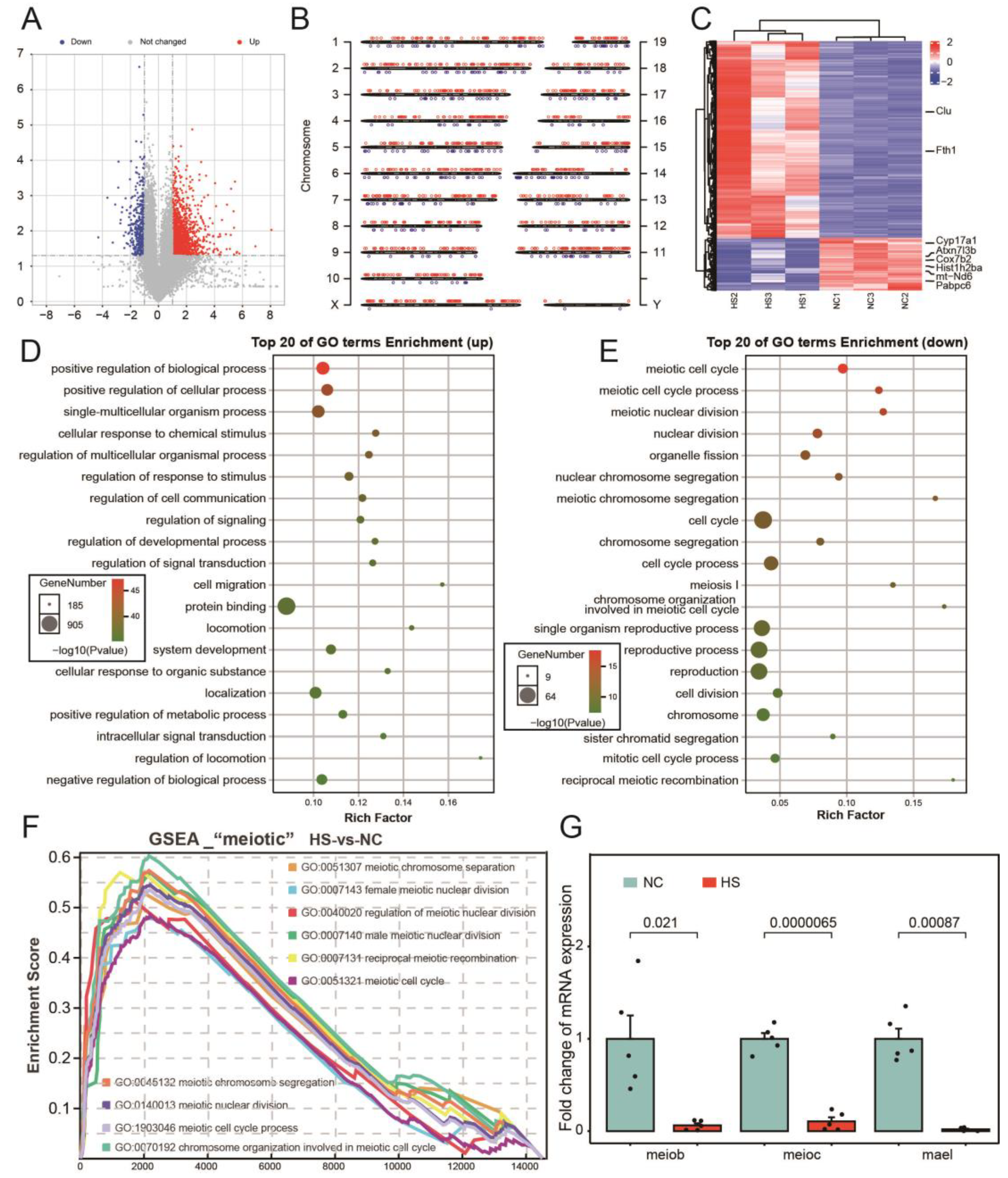
3.2. Abnormal Cell Cycle of Testicular Tissue after Testicular Heat Stress
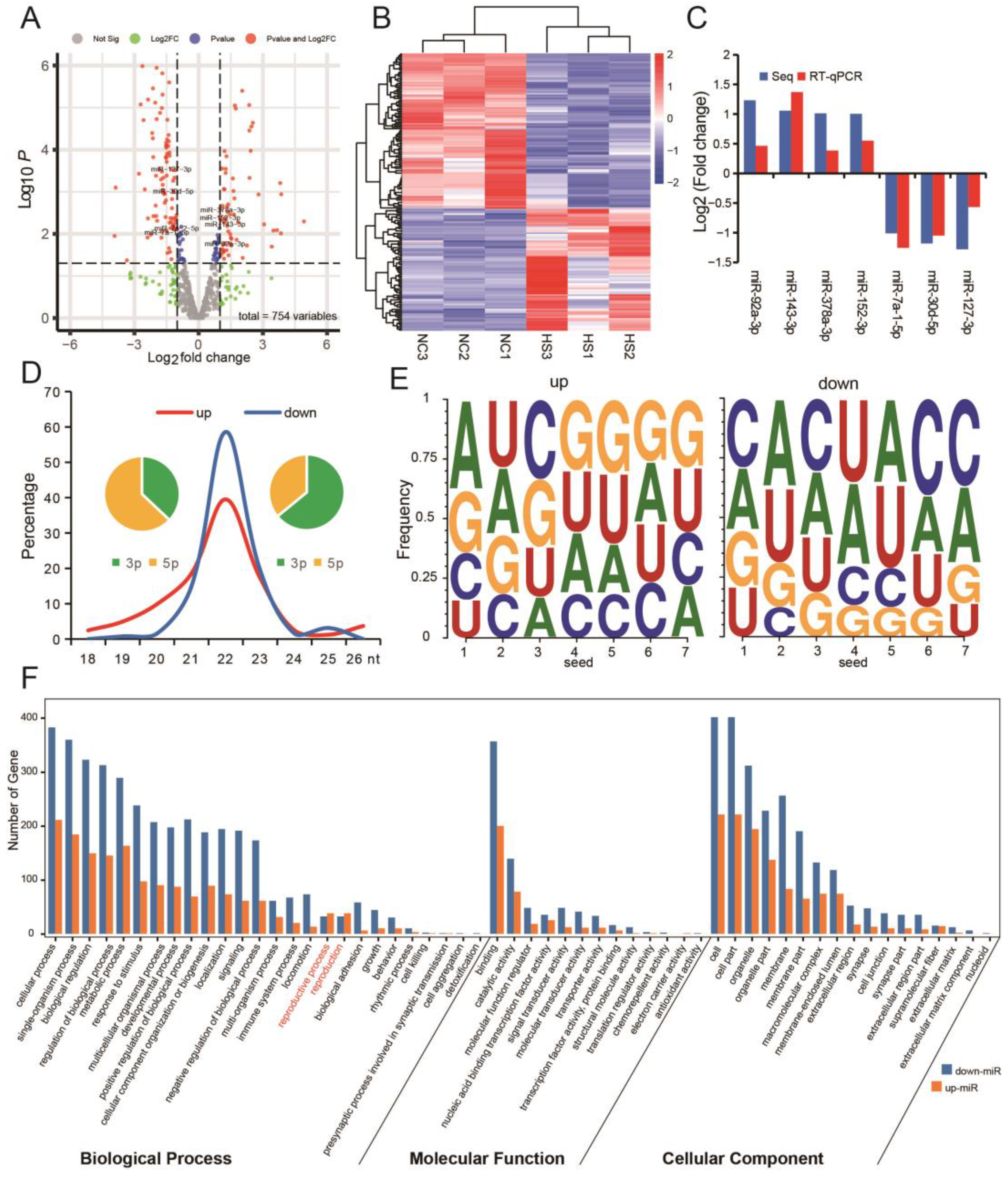
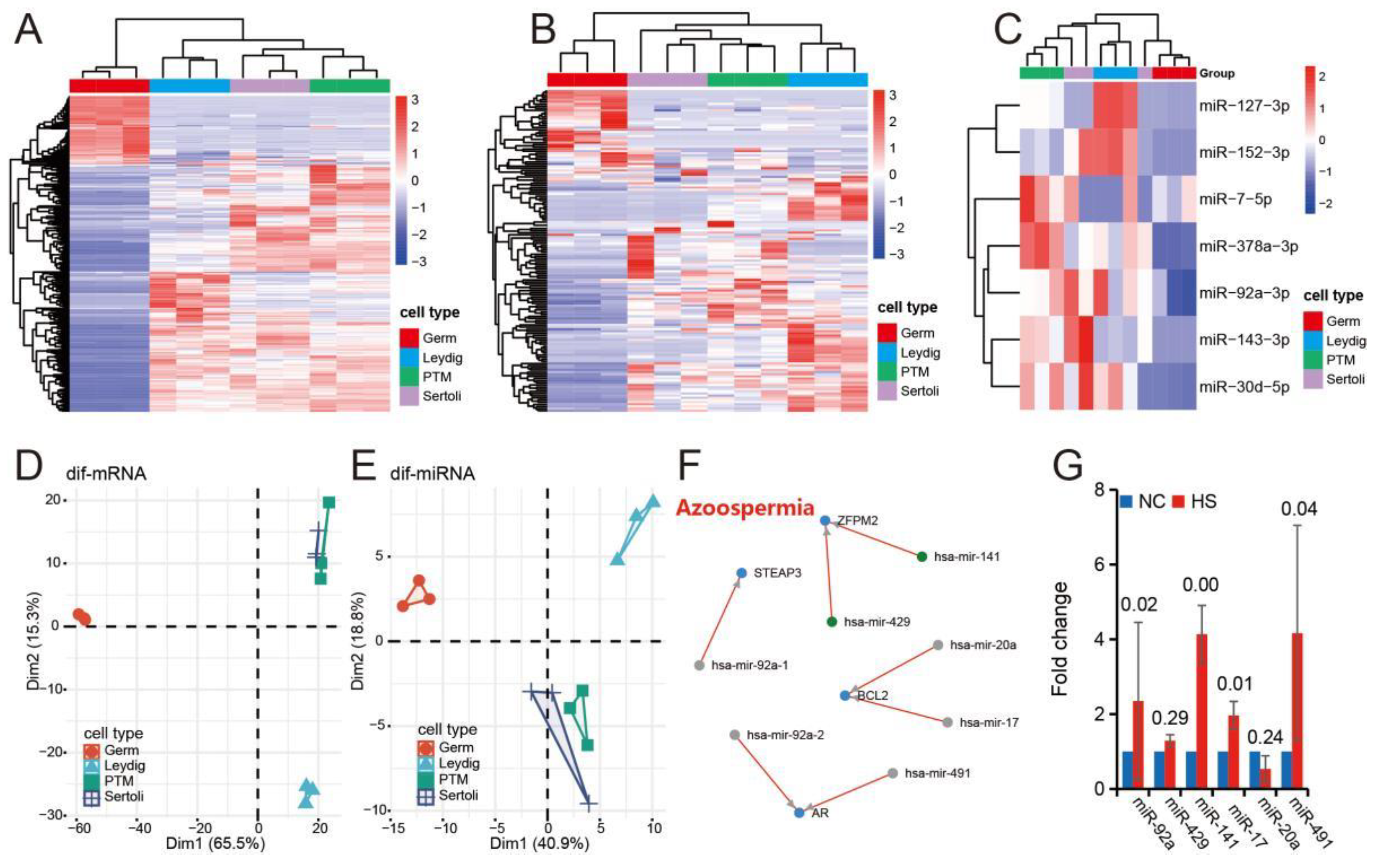
3.3. Testicular Heat Stress Changes the Expression Profile of miRNAs in Testicular Tissue
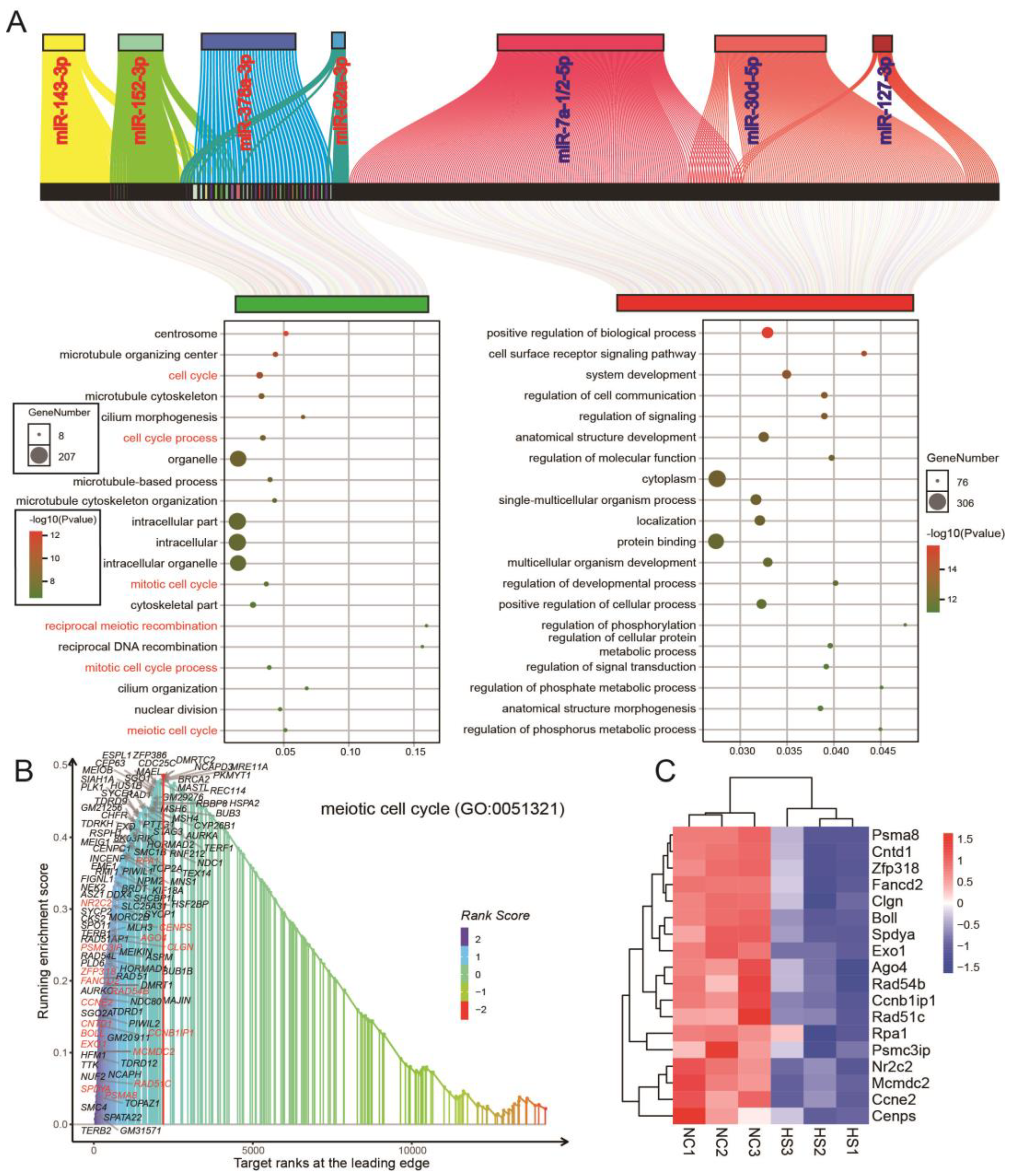
3.4. Differential miRNA and mRNA Co-Expression Networks in Testicular Tissue of Normal and Heat-Stressed Mice
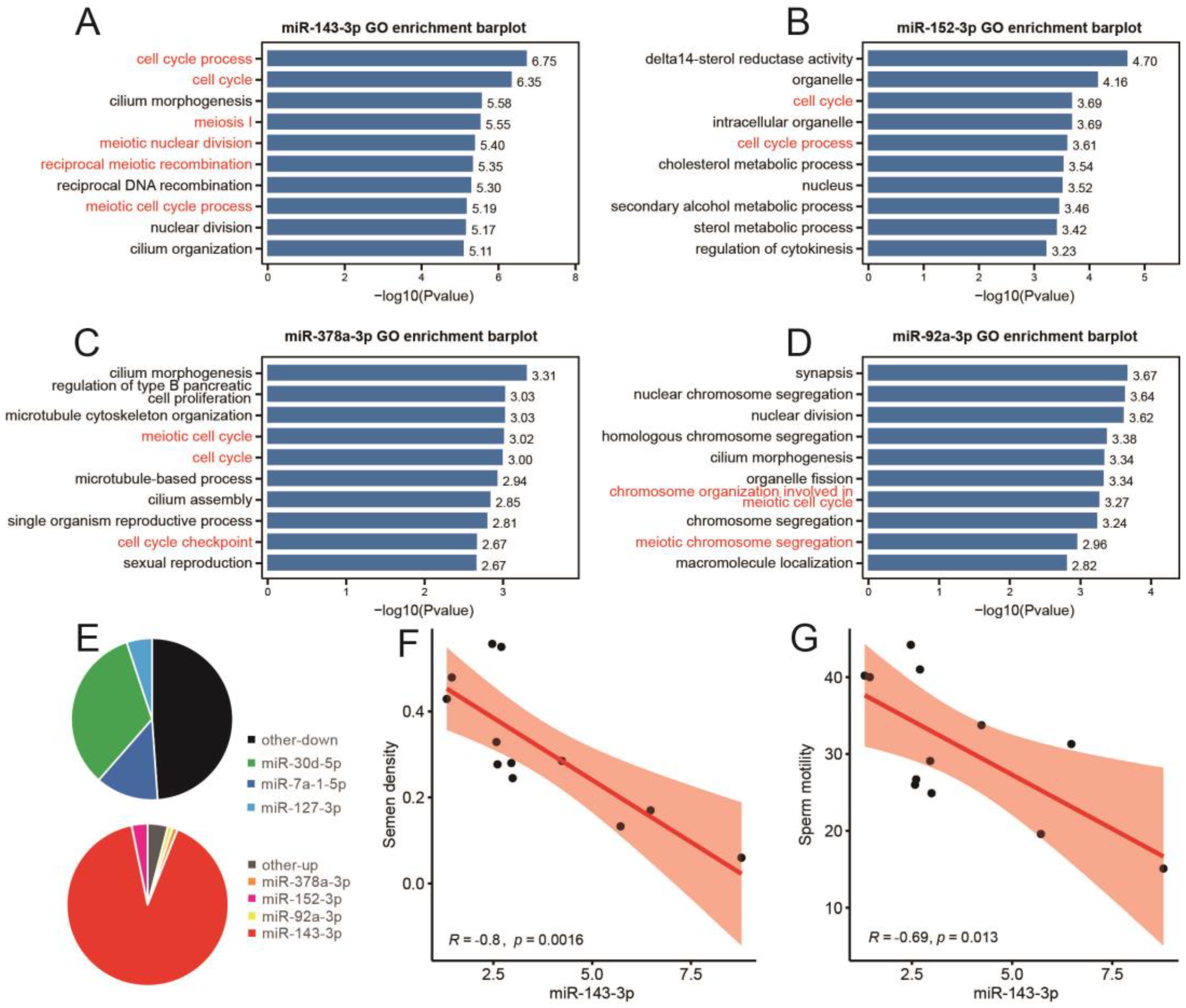
3.5. Expression Characteristics of Differentially Expressed miRNAs Induced by Heat Stress in Different Cell Groups of Mice Testis
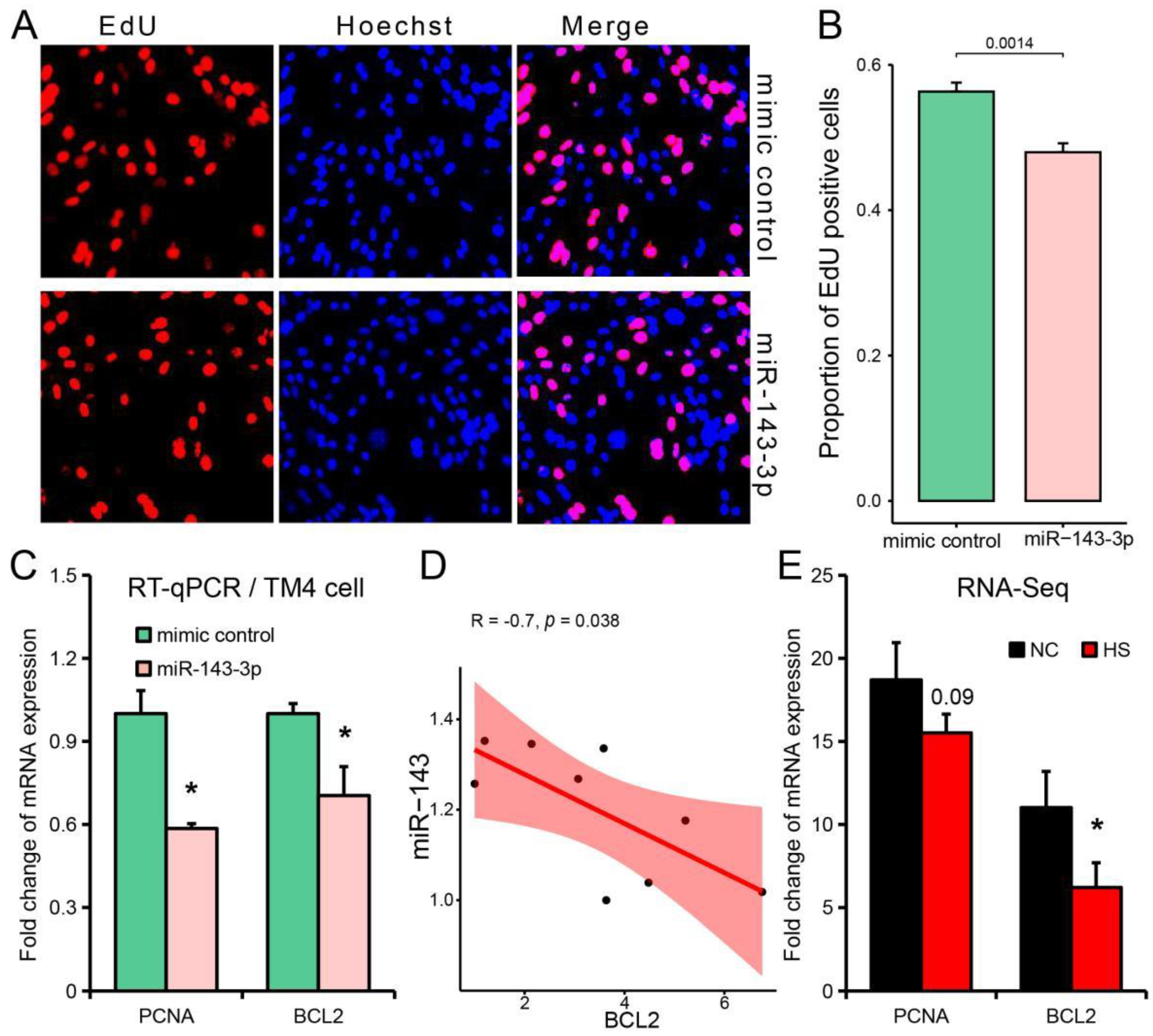
3.6. miR-143-3p Can Be Used as a Representative Potential Biomarker of Testicular Heat Stress
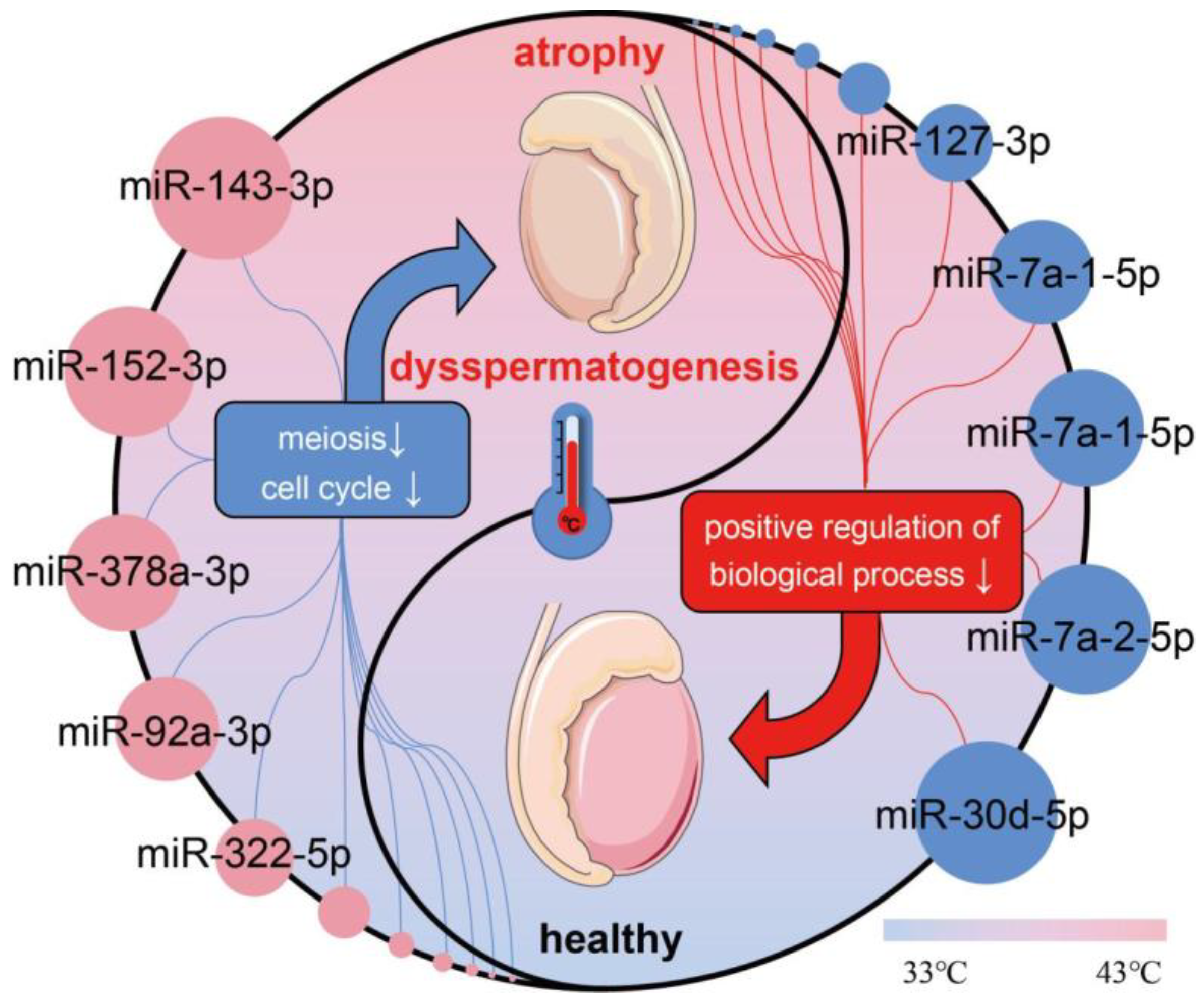
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Inhorn, M.; Patrizio, P. Infertility around the globe: New thinking on gender, reproductive technologies and global movements in the 21st century. Hum. Reprod. Update 2015, 21, 411–426. [Google Scholar] [CrossRef] [PubMed]
- Amirjannaty, S.; Gashti, N.; Mojtahedi, A.; Ashouri, A.; Bahadori, M. An In vitro Study on the Protective Effect of Melatonin on Human Sperm Parameters Treated by Cadmium. J. Hum. Reprod. Sci. 2022, 15, 21. [Google Scholar] [CrossRef] [PubMed]
- Mann, U.; Shiff, B.; Patel, P. Reasons for worldwide decline in male fertility. Curr. Opin. Urol. 2020, 30, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Nordkap, L.; Joensen, U.; Blomberg Jensen, M.; Jørgensen, N. Regional differences and temporal trends in male reproductive health disorders: Semen quality may be a sensitive marker of environmental exposures. Mol. Cell. Endocrinol. 2012, 355, 221–230. [Google Scholar] [CrossRef]
- Levine, H.; Jørgensen, N.; Martino-Andrade, A.; Mendiola, J.; Weksler-Derri, D.; Jolles, M.; Pinotti, R.; Swan, S. Temporal trends in sperm count: A systematic review and meta-regression analysis of samples collected globally in the 20th and 21st centuries. Hum. Reprod. Update 2022, 29, 157–176. [Google Scholar] [CrossRef]
- Lea, R.; Byers, A.; Sumner, R.; Rhind, S.; Zhang, Z.; Freeman, S.; Moxon, R.; Richardson, H.; Green, M.; Craigon, J.; et al. Environmental chemicals impact dog semen quality in vitro and may be associated with a temporal decline in sperm motility and increased cryptorchidism. Sci. Rep. 2016, 6, 31281. [Google Scholar] [CrossRef]
- Wahl, R.; Reif, J. Temporal trends in bull semen quality: A comparative model for human health? Environ. Res. 2009, 109, 273–280. [Google Scholar] [CrossRef]
- Perrett, J.; Harris, I.; Maddock, C.; Farnworth, M.; Pyatt, A.; Sumner, R. Systematic Analysis of Breed, Methodological, and Geographical Impact on Equine Sperm Progressive Motility. Animals 2021, 11, e3088. [Google Scholar] [CrossRef]
- Hoang-Thi, A.-P.; Dang-Thi, A.-T.; Sang, P.; Nguyen Ba, T.; Truong-Thi, P.-L.; Le, M.T.; Nguyen Vu Quoc, H.; Nguyen Thanh, T. The Impact of High Ambient Temperature on Human Sperm Parameters: A Meta-Analysis. Iran. J. Public Health 2022, 51, 710–723. [Google Scholar] [CrossRef]
- Boni, R. Heat stress, a serious threat to reproductive function in animals and humans. Mol. Reprod. Dev. 2019, 86, 1307–1323. [Google Scholar] [CrossRef]
- Aldahhan, R.; Stanton, P. Heat stress response of somatic cells in the testis. Mol. Cell. Endocrinol. 2021, 527, 111216. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Crespo, M.; Pintado, B.; Gutiérrez-Adán, A. Scrotal heat stress effects on sperm viability, sperm DNA integrity, and the offspring sex ratio in mice. Mol. Reprod. Dev. 2008, 75, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, N.; Andersen, A.; Eustache, F.; Irvine, D.; Suominen, J.; Petersen, J.; Andersen, A.; Auger, J.; Cawood, E.; Horte, A.; et al. Regional differences in semen quality in Europe. Hum. Reprod. 2001, 16, 1012–1019. [Google Scholar] [CrossRef]
- Rizzoto, G.; Boe-Hansen, G.; Klein, C.; Thundathil, J.; Kastelic, J. Acute mild heat stress alters gene expression in testes and reduces sperm quality in mice. Theriogenology 2020, 158, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, Z.; Fang, T.; Zhang, Y.; Chen, L.; Du, Z.; Yang, C. Identification of internal reference genes for porcine immature Sertoli cells under heat stress. Reprod. Domest. Anim. 2022, 57, 1344–1352. [Google Scholar] [CrossRef] [PubMed]
- Rockett, J.C.; Mapp, F.L.; Garges, J.B.; Luft, J.C.; Mori, C.; Dix, D.J. Effects of Hyperthermia on Spermatogenesis, Apoptosis, Gene Expression, and Fertility in Adult Male Mice1. Biol. Reprod. 2001, 65, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Mainigi, M.; Sapienza, C.; Butts, S.; Coutifaris, C. A Molecular Perspective on Procedures and Outcomes with Assisted Reproductive Technologies. Cold Spring Harb. Perspect. Med. 2016, 6, a023416. [Google Scholar] [CrossRef]
- Jodar, M.; Selvaraju, S.; Sendler, E.; Diamond, M.; Krawetz, S. The presence, role and clinical use of spermatozoal RNAs. Hum. Reprod. Update 2013, 19, 604–624. [Google Scholar] [CrossRef]
- Aliakbari, F.; Eshghifar, N.; Mirfakhraie, R.; Pourghorban, P.; Azizi, F. Coding and Non-Coding RNAs, as Male Fertility and Infertility Biomarkers. Int. J. Fertil. Steril. 2021, 15, 158–166. [Google Scholar] [CrossRef]
- Wu, D.; Khan, F.; Huo, L.; Sun, F.; Huang, C. Alternative splicing and MicroRNA: Epigenetic mystique in male reproduction. RNA Biol. 2022, 19, 162–175. [Google Scholar] [CrossRef]
- Walker, W. Regulation of mammalian spermatogenesis by miRNAs. Semin. Cell Dev. Biol. 2022, 121, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Loughlin, K.; Manson, K.; Foreman, R.; Schwartz, B.; Heuttner, P. The effect of intermittent scrotal hyperthermia on the Sprague-Dawley rat testicle. Adv. Exp. Med. Biol. 1991, 286, 183–185. [Google Scholar] [CrossRef] [PubMed]
- Rao, M.; Zeng, Z.; Tang, L.; Cheng, G.; Xia, W.; Zhu, C. Next-generation sequencing-based microRNA profiling of mice testis subjected to transient heat stress. Oncotarget 2017, 8, 111672–111682. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.; He, C.; Sun, X.; Li, L.; Xu, Y.; Zhang, K.; Liu, X.; Liang, M. Integrated study of circRNA, lncRNA, miRNA, and mRNA networks in mediating the effects of testicular heat exposure. Cell Tissue Res. 2021, 386, 127–143. [Google Scholar] [CrossRef] [PubMed]
- Sharma, U.; Sun, F.; Conine, C.; Reichholf, B.; Kukreja, S.; Herzog, V.; Ameres, S.; Rando, O. Small RNAs Are Trafficked from the Epididymis to Developing Mammalian Sperm. Dev. Cell 2018, 46, 481–494.e6. [Google Scholar] [CrossRef]
- Guang-Yu, L.; Hai-Yan, L.; Ji-Hong, L.; Yun-Cong, M.; Xue-Lian, D.; Chun-Yu, L.; Wen-Yong, S. MCL1 is a key regulator of steroidogenesis in mouse Leydig cells. Mol. Reprod. Dev. 2016, 83, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Lowe, T.; Chan, P. tRNAscan-SE On-line: Integrating search and context for analysis of transfer RNA genes. Nucleic Acids Res. 2016, 44, 54–57. [Google Scholar] [CrossRef]
- Betel, D.; Wilson, M.; Gabow, A.; Marks, D.; Sander, C. The microRNA.org resource: Targets and expression. Nucleic Acids Res. 2008, 36, 149–153. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Lin, C.; Shin, D.-G.; Park, S.G.; Chu, S.B.; Gwon, L.W.; Lee, J.-G.; Yon, J.-M.; Baek, I.-J.; Nam, S.-Y. Curcumin dose-dependently improves spermatogenic disorders induced by scrotal heat stress in mice. Food Funct. 2015, 6, 3770–3777. [Google Scholar] [CrossRef]
- Miura, M.; Sasagawa, I.; Suzuki, Y.; Nakada, T.; Fujii, J. Apoptosis and expression of apoptosis-related genes in the mouse testis following heat exposure. Fertil. Steril. 2002, 77, 787–793. [Google Scholar] [CrossRef] [PubMed]
- Wang, S. PCNA: A silent housekeeper or a potential therapeutic target? Trends Pharmacol. Sci. 2014, 35, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Vermeulen, K.; Berneman, Z.; Van Bockstaele, D. Cell cycle and apoptosis. Cell Prolif. 2003, 36, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Huang, J.; Ma, P.; Yu, G. microRNA-143 acts as a suppressor of hemangioma growth by targeting Bcl-2. Gene 2017, 628, 211–217. [Google Scholar] [CrossRef]
- Qian, Y.; Teng, Y.; Li, Y.; Lin, X.; Guan, M.; Li, Y.; Cao, X.; Gao, Y. MiR-143-3p suppresses the progression of nasal squamous cell carcinoma by targeting Bcl-2 and IGF1R. Biochem. Biophys. Res. Commun. 2019, 518, 492–499. [Google Scholar] [CrossRef]
- Irvine, D.S. Epidemiology and aetiology of male infertility. Hum. Reprod. 1998, 13 (Suppl. S1), 33–44. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Xu, R.; Luan, Y.; Fan, H.; Yang, S.; Liu, J.; Zeng, H.; Shao, L. Rapamycin Ameliorates Radiation-Induced Testis Damage in Mice. Front. Cell Dev. Biol. 2022, 10, 783884. [Google Scholar] [CrossRef]
- Rato, L.; Alves, M.; Socorro, S.; Duarte, A.; Cavaco, J.; Oliveira, P. Metabolic regulation is important for spermatogenesis. Nat. Rev. Urol. 2012, 9, 330–338. [Google Scholar] [CrossRef]
- Parrish, J.J.; Willenburg, K.L.; Gibbs, K.M.; Yagoda, K.B.; Krautkramer, M.M.; Loether, T.M.; Melo, F.C.S.A. Scrotal insulation and sperm production in the boar. Mol. Reprod. Dev. 2017, 84, 969–978. [Google Scholar] [CrossRef]
- Mishra, R.K.; Singh, S.K. Safety assessment of Syzygium aromaticum flower bud (clove) extract with respect to testicular function in mice. Food Chem. Toxicol. 2008, 46, 3333–3338. [Google Scholar] [CrossRef]
- Johnson, L.; Welsh, T.H.; Curley, K.O.; Johnston, C.E. 11.02—Anatomy and Physiology of the Male Reproductive System and Potential Targets of Toxicants. In Comprehensive Toxicology, 2nd ed.; McQueen, C.A., Ed.; Elsevier: Oxford, UK, 2010; pp. 5–59. [Google Scholar]
- Li, Z.; Wang, S.; Gong, C.; Hu, Y.; Liu, J.; Wang, W.; Chen, Y.; Liao, Q.; He, B.; Huang, Y.; et al. Effects of Environmental and Pathological Hypoxia on Male Fertility. Front. Cell Dev. Biol. 2021, 9, 725933. [Google Scholar] [CrossRef] [PubMed]
- França, L.; Avelar, G.; Almeida, F. Spermatogenesis and sperm transit through the epididymis in mammals with emphasis on pigs. Theriogenology 2005, 63, 300–318. [Google Scholar] [CrossRef] [PubMed]
- Padhi, A.; Ghaly, M.M.; Ma, L. Testis-enriched heat shock protein A2 (HSPA2): Adaptive advantages of the birds with internal testes over the mammals with testicular descent. Sci. Rep. 2016, 6, 18770. [Google Scholar] [CrossRef] [PubMed]
- Shahat, A.M.; Rizzoto, G.; Kastelic, J.P. Amelioration of heat stress-induced damage to testes and sperm quality. Theriogenology 2020, 158, 84–96. [Google Scholar] [CrossRef]
- He, C.; Wang, K.; Gao, Y.; Wang, C.; Li, L.; Liao, Y.; Hu, K.; Liang, M. Roles of Noncoding RNA in Reproduction. Front. Genet. 2021, 12, 777510. [Google Scholar] [CrossRef] [PubMed]
- Van Pelt, A.; Rooij, D. Synchronization of the Seminiferous Epithelium after Vitamin A Replacement in Vitamin A-Deficient Mice. Biol. Reprod. 1990, 43, 363–367. [Google Scholar] [CrossRef]
- Cai, H.; Qin, D.; Peng, S. Responses and coping methods of different testicular cell types to heat stress: Overview and perspectives. Biosci. Rep. 2021, 41, BSR20210443. [Google Scholar] [CrossRef]
- Hirano, K.; Nonami, Y.; Nakamura, Y.; Sato, T.; Sato, T.; Ishiguro, K.-I.; Ogawa, T.; Yoshida, S. Temperature sensitivity of DNA double-strand break repair underpins heat-induced meiotic failure in mouse spermatogenesis. Commun. Biol. 2022, 5, 504. [Google Scholar] [CrossRef]
- Levitas, E.; Lunenfeld, E.; Weisz, N.; Friger, M.; Har-Vardi, I. Seasonal variations of human sperm cells among 6455 semen samples: A plausible explanation of a seasonal birth pattern. Am. J. Obstet. Gynecol. 2013, 208, 406.e1–406.e6. [Google Scholar] [CrossRef]
- Henkel, R.; Menkveld, R.; Kleinhappl, M.; Schill, W.B. Seasonal changes in human sperm chromatin condensation. J. Assist. Reprod. Genet. 2001, 18, 371–377. [Google Scholar] [CrossRef]
- Gruhot, T.; Gray, K.; Brown, V.; Huang, Y.; Kachman, S.D.; Spangler, M.L.; Mote, B. Genetic relationships among sperm quality traits of Duroc boars collected during the summer season. Anim. Reprod. Sci. 2019, 206, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Malama, E.; Zeron, Y.; Janett, F.; Siuda, M.; Roth, Z.; Bollwein, H. Use of computer-assisted sperm analysis and flow cytometry to detect seasonal variations of bovine semen quality. Theriogenology 2017, 87, 79–90. [Google Scholar] [CrossRef]
- Ahmad, E.; Naseer, Z.; Uçan, U.; Serin, İ.; Ceylan, A.; Aksoy, M. Seasonal variations in sperm acrosome reaction, osmotic tolerance and serum testosterone concentrations in rams. Anim. Reprod. Sci. 2018, 198, 112–120. [Google Scholar] [CrossRef]
- Zhang, P.; Zheng, Y.; Lv, Y.; Li, F.; Su, L.; Qin, Y.; Zeng, W. Melatonin protects the mouse testis against heat-induced damage. Mol. Hum. Reprod. 2020, 26, 65–79. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Wang, C.; Wang, K.; He, C.; Hu, K.; Liang, M. The effects and molecular mechanism of heat stress on spermatogenesis and the mitigation measures. Syst. Biol. Reprod. Med. 2022, 68, 331–347. [Google Scholar] [CrossRef] [PubMed]
- Kheradmand, A.; Dezfoulian, O.; Tarrahi, M. Ghrelin attenuates heat-induced degenerative effects in the rat testis. Regul. Pept. 2011, 167, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Melo, I.; Angelo Mendes Tenorio, F.; da Silva Gomes, J.; da Silva Junior, V.; de Albuquerque Nogueira, R.; Tenorio, B. Fractal methods applied to the seminiferous lumen images can quantify testicular changes induced by heat stress. Acta Histochem. 2022, 124, 151949. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Oliveros, L.N.; Arruda, R.P.d.; Batissaco, L.; Gonzaga, V.H.G.; Nogueira, V.J.M.; Florez-Rodriguez, S.A.; Almeida, F.d.S.; Alves, M.B.R.; Pinto, S.C.C.; Nichi, M.; et al. Chronological characterization of sperm morpho-functional damage and recovery after testicular heat stress in Nellore bulls. J. Therm. Biol. 2022, 106, 103237. [Google Scholar] [CrossRef]
- Wang, J.; Gao, W.; Deng, S.; Liu, X.; Jia, H.; Ma, W. High temperature suppressed SSC self-renewal through S phase cell cycle arrest but not apoptosis. Stem Cell Res. Ther. 2019, 10, 227. [Google Scholar] [CrossRef]
- Zhu, Q.; Kirby, J.; Chu, C.; Gou, L. Small Noncoding RNAs in Reproduction and Infertility. Biomedicines 2021, 9, 1884. [Google Scholar] [CrossRef]
- Ren, X.; Chen, X.; Wang, Z.; Wang, D. Is transcription in sperm stationary or dynamic? J. Reprod. Dev. 2017, 63, 439–443. [Google Scholar] [CrossRef] [PubMed]
- Turri, F.; Capra, E.; Lazzari, B.; Cremonesi, P.; Stella, A.; Pizzi, F. A Combined Flow Cytometric Semen Analysis and miRNA Profiling as a Tool to Discriminate Between High- and Low-Fertility Bulls. Front. Vet. Sci. 2021, 8, 703101. [Google Scholar] [CrossRef] [PubMed]
- Kawata, R.; Kagawa, T.; Koya, Y.; Kajiyama, H.; Oda, S.; Yokoi, T. Exploration of small RNA biomarkers for testicular injury in the serum exosomes of rats. Toxicology 2020, 440, 152490. [Google Scholar] [CrossRef]
- Zhang, R.; Zuo, Y.; Cao, S. Upregulated microRNA-423-5p promotes oxidative stress through targeting glutathione S-transferase mu 1 in asthenozoospermia. Mol. Reprod. Dev. 2021, 88, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Zou, L.; Cheng, G.; Xu, C.; Liu, H.; Wang, Y.; Li, N.; Zhu, C.; Xia, W. The role of miR-128-3p through MAPK14 activation in the apoptosis of GC2 spermatocyte cell line following heat stress. Andrology 2021, 9, 665–672. [Google Scholar] [CrossRef]
- Huang, Z.; Shi, J.; Gao, Y.; Cui, C.; Zhang, S.; Li, J.; Zhou, Y.; Cui, Q. HMDD v3.0: A database for experimentally supported human microRNA-disease associations. Nucleic Acids Res. 2019, 47, D1013–D1017. [Google Scholar] [CrossRef]
- Ding, N.; Zhang, Y.; Huang, M.; Liu, J.; Wang, C.; Zhang, C.; Cao, J.; Zhang, Q.; Jiang, L. Circ-CREBBP inhibits sperm apoptosis via the PI3K-Akt signaling pathway by sponging miR-10384 and miR-143-3p. Commun. Biol. 2022, 5, 1339. [Google Scholar] [CrossRef]
- Salas-Huetos, A.; Blanco, J.; Vidal, F.; Godo, A.; Grossmann, M.; Pons, M.; F-Fernández, S.; Garrido, N.; Anton, E. Spermatozoa from patients with seminal alterations exhibit a differential micro-ribonucleic acid profile. Fertil. Steril. 2015, 104, 591–601. [Google Scholar] [CrossRef]
- Kamijo, S.; Hamatani, T.; Sasaki, H.; Suzuki, H.; Abe, A.; Inoue, O.; Iwai, M.; Ogawa, S.; Odawara, K.; Tanaka, K.; et al. MicroRNAs secreted by human preimplantation embryos and IVF outcome. Reprod. Biol. Endocrinol. 2022, 20, 130. [Google Scholar] [CrossRef]
- Corral-Vazquez, C.; Blanco, J.; Salas-Huetos, A.; Vidal, F.; Anton, E. Normalization matters: Tracking the best strategy for sperm miRNA quantification. Mol. Hum. Reprod. 2017, 23, 45–53. [Google Scholar] [CrossRef]
- Bizuayehu, T.; Babiak, I. Heterogenic Origin of Micro RNAs in Atlantic Salmon (Salmo salar) Seminal Plasma. Int. J. Mol. Sci. 2020, 21, 2723. [Google Scholar] [CrossRef] [PubMed]
- Alves, M.B.R.; Arruda, R.P.d.; Batissaco, L.; Garcia-Oliveros, L.N.; Gonzaga, V.H.G.; Nogueira, V.J.M.; Almeida, F.D.S.; Pinto, S.C.C.; Andrade, G.M.; Perecin, F.; et al. Changes in miRNA levels of sperm and small extracellular vesicles of seminal plasma are associated with transient scrotal heat stress in bulls. Theriogenology 2021, 161, 26–40. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Chen, M.; Ji, X.; Yao, M.; Mao, Z.; Zhou, K.; Xia, Y.; Han, X.; Tang, W. Metabolomic profiles reveal key metabolic changes in heat stress-treated mouse Sertoli cells. Toxicol. Vitr. 2015, 29, 1745–1752. [Google Scholar] [CrossRef]
- Li, B.; Fan, J.; Chen, N. A Novel Regulator of Type II Diabetes: MicroRNA-143. Trends Endocrinol. Metab. TEM 2018, 29, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Vacante, F.; Denby, L.; Sluimer, J.; Baker, A. The function of miR-143, miR-145 and the MiR-143 host gene in cardiovascular development and disease. Vasc. Pharmacol. 2019, 112, 24–30. [Google Scholar] [CrossRef]
- Cordes, K.R.; Sheehy, N.T.; White, M.P.; Berry, E.C.; Morton, S.U.; Muth, A.N.; Lee, T.-H.; Miano, J.M.; Ivey, K.N.; Srivastava, D. miR-145 and miR-143 regulate smooth muscle cell fate and plasticity. Nature 2009, 460, 705–710. [Google Scholar] [CrossRef]
- Ma, W.-Y.; Song, R.-J.; Xu, B.-B.; Xu, Y.; Wang, X.-X.; Sun, H.-Y.; Li, S.-N.; Liu, S.-Z.; Yu, M.-X.; Yang, F.; et al. Melatonin promotes cardiomyocyte proliferation and heart repair in mice with myocardial infarction via miR-143-3p/Yap/Ctnnd1 signaling pathway. Acta Pharmacol. Sin. 2021, 42, 921–931. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Jiang, J.; He, G.; Zhou, H.; Ji, C. miR-143-3p represses leukemia cell proliferation by inhibiting KAT6A expression. Anticancer Drugs 2022, 33, e662. [Google Scholar] [CrossRef]
- Yan, X.; Chen, X.; Liang, H.; Deng, T.; Chen, W.; Zhang, S.; Liu, M.; Gao, X.; Liu, Y.; Zhao, C.; et al. miR-143 and miR-145 synergistically regulate ERBB3 to suppress cell proliferation and invasion in breast cancer. Mol. Cancer 2014, 13, 220. [Google Scholar] [CrossRef]
- Sofikitis, N.; Giotitsas, N.; Tsounapi, P.; Baltogiannis, D.; Giannakis, D.; Pardalidis, N. Hormonal regulation of spermatogenesis and spermiogenesis. J. Steroid Biochem. Mol. Biol. 2008, 109, 323–330. [Google Scholar] [CrossRef]
- Dimitriadis, F.; Tsiampali, C.; Chaliasos, N.; Tsounapi, P.; Takenaka, A.; Sofikitis, N. The Sertoli cell as the orchestra conductor of spermatogenesis: Spermatogenic cells dance to the tune of testosterone. Hormones 2015, 14, 479–503. [Google Scholar] [CrossRef] [PubMed]
- Menezes, E.S.B.; Badial, P.R.; El Debaky, H.; Husna, A.U.; Ugur, M.R.; Kaya, A.; Topper, E.; Bulla, C.; Grant, K.E.; Bolden-Tiller, O.; et al. Sperm miR-15a and miR-29b are associated with bull fertility. Andrologia 2020, 52, e13412. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gan, M.; Jing, Y.; Xie, Z.; Ma, J.; Chen, L.; Zhang, S.; Zhao, Y.; Niu, L.; Wang, Y.; Li, X.; et al. Potential Function of Testicular MicroRNAs in Heat-Stress-Induced Spermatogenesis Disorders. Int. J. Mol. Sci. 2023, 24, 8809. https://doi.org/10.3390/ijms24108809
Gan M, Jing Y, Xie Z, Ma J, Chen L, Zhang S, Zhao Y, Niu L, Wang Y, Li X, et al. Potential Function of Testicular MicroRNAs in Heat-Stress-Induced Spermatogenesis Disorders. International Journal of Molecular Sciences. 2023; 24(10):8809. https://doi.org/10.3390/ijms24108809
Chicago/Turabian StyleGan, Mailin, Yunhong Jing, Zhongwei Xie, Jianfeng Ma, Lei Chen, Shunhua Zhang, Ye Zhao, Lili Niu, Yan Wang, Xuewei Li, and et al. 2023. "Potential Function of Testicular MicroRNAs in Heat-Stress-Induced Spermatogenesis Disorders" International Journal of Molecular Sciences 24, no. 10: 8809. https://doi.org/10.3390/ijms24108809