
Carcino-Evo-Devo, A Theory of the Evolutionary Role of Hereditary Tumors


1
Vavilov Institute of General Genetics, Russian Academy of Sciences, 3 Gubkina Street, 117971 Moscow, Russia
2
Peter the Great St. Petersburg Polytechnic University, 29 Polytekhnicheskaya Street, 195251 St. Petersburg, Russia
Int. J. Mol. Sci. 2023, 24(10), 8611; https://doi.org/10.3390/ijms24108611
Submission received: 16 March 2023
/
Revised: 8 May 2023
/
Accepted: 9 May 2023
/
Published: 11 May 2023
(This article belongs to the Special Issue Systems Biology Approach in Cancer)
Abstract
:A theory of the evolutionary role of hereditary tumors, or the carcino-evo-devo theory, is being developed. The main hypothesis of the theory, the hypothesis of evolution by tumor neofunctionalization, posits that hereditary tumors provided additional cell masses during the evolution of multicellular organisms for the expression of evolutionarily novel genes. The carcino-evo-devo theory has formulated several nontrivial predictions that have been confirmed in the laboratory of the author. It also suggests several nontrivial explanations of biological phenomena previously unexplained by the existing theories or incompletely understood. By considering three major types of biological development—individual, evolutionary, and neoplastic development—within one theoretical framework, the carcino-evo-devo theory has the potential to become a unifying biological theory.
1. Introduction
The theory of the evolutionary role of hereditary tumors, or the carcino-evo-devo theory, was developed in order to explain the sources of additional cell masses, which were necessary for the origins of new cell types, tissues, and organs during the progressive evolution of multicellular organisms. The content of the theory has been published in the theoretical papers of the author [1,2,3,4,5,6,7,8,9,10,11,12,13] and as a monograph in English [14], Russian [15], and Chinese [16]. Many experimental papers from the author’s lab were devoted to the confirmation of nontrivial predictions of the carcino-evo-devo theory [17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39].
The basic statements of the carcino-evo-devo theory are the following:
- Tumor processes participate in the evolution of development;
- Hereditary tumors provide evolving multicellular organisms with extra cell masses for the expression of evolutionarily novel genes and gene combinations, and thereby participate in the origins of new cell types, tissues, and organs;
- Populations of tumor-bearing organisms served as transitional forms in progressive evolution;
- Tumors may be considered search engines for new gene combinations in the space of biological possibilities.
Currently, the carcino-evo-devo theory consists of several parts describing different aspects of the theory. The main divisions of the theory include the following: (1) the analysis of the previous concepts; (2) the analysis of tumor features that might be used in evolution; (3) the main hypothesis (i.e., the hypothesis of evolution by tumor neofunctionalization); (4) nontrivial predictions and the results of their experimental confirmation; (5) nontrivial explanations of the carcino-evo-devo theory and the analysis of its relationship to other biological theories; (6) carcino-evo-devo diagrams; (7) the concept of tumors as search engines in the space of biological possibilities and their relationship to biological computational processes; and (8) tumors and the biocomputational formulation of the increase in complexity principle.
In this paper, the author will present the current state of the carcino-evo-devo theory by describing each of the theory’s principal divisions.
2. Preceding and Related Concepts
The term “carcino-evo-devo” was first introduced in [14], and it was used for the first time as a name for the theory of the evolutionary role of tumors in [9]. The new term was coined from two other terms: “carcinoembryonic” and “evo-devo”.
The term “carcinoembryonic” is used to designate embryonal proteins produced by tumor cells. The first examples of such proteins were alpha-fetoprotein [40,41] and carcinoembryonic antigen [42]. The concept of carcinoembryonic proteins was further developed in the concept of the convergence of embryonic and cancer signaling pathways [43,44,45,46]. These concepts point to molecular links between embryonal and tumor processes.
The abbreviation “evo-devo” corresponds to evolutionary developmental biology, which examines connections of normal individual development and evolution [47].
The carcino-evo-devo theory interconnects evolutionary, individual, and neoplastic development within one unified consideration. This theory studies the role of hereditary tumors in the evolution of development [9,14].
The previous concepts important for the carcino-evo-devo theory are the “embryonal rest” or “embryonal remnants” theory of cancer [48,49,50]; the morphological laws of the evolution of ontogenesis [51]; and the concept of neoplasia as a disease of differentiation [52].
Several previous ideas may be considered historical background for our theory, such as the idea of the role of “hopeful monsters” in evolution [53,54,55,56], and the idea of the positive role of viruses in evolution [57].
The carcino-evo-devo theory has been developing concurrently with Darwinian medicine [58], evolutionary epidemiology [59], and several branches of evolutionary oncology (e.g., the somatic evolution of tumor cells and selection in tumor cell populations [60,61,62], cancer selection [63], and the ecological hypothesis [64]). Comparative oncology has been in place since the 19th century (reviewed in [14]).
So, the carcino-evo-devo theory is deeply rooted in experimental oncology, developmental and evolutionary biology, and evolutionary medicine.
3. Tumor Features That Suggest the Role of Tumors in the Evolution of Organisms
There are features of tumors that suggest their evolutionary role. Many of them have been discussed in the author’s book [14]. Here, the author will briefly discuss this topic with the addition of new data and references, which were not cited in the book or appeared after its publication.
3.1. Hereditary Tumors, like Any Hereditary Trait, Could Be Used in Evolution
Many tumors are hereditary. The high incidence of certain tumors in different strains of laboratory animals is a well-known phenomenon [14,65,66]. Cellular oncogenes and tumor suppressor genes are inherited as part of germline DNA and thus provide a genetic basis for tumor inheritance. The importance of germline mutations in cancer was recognized after the pioneering works of Alfred Knudson [67,68,69]. Twenty percent of cancer-associated mutations occur in the germline DNA, and ten percent are both somatic and germline mutations [70,71]. There is an inverse association between germline susceptibility risk and somatic genetic variation in human cancer [72,73]. Heritable epimutations associated with hereditary tumors (e.g., in Lynch syndrome) have also been described [74].
Tumor prevalence in wild and captive animals can reach a high percentage (e.g., >50% in Santa Catalina Island foxes) [75,76]. The highest cancer prevalence has been recorded in species or populations with low genetic diversity, which suggests that it may have a genetic basis (reviewed in [75]). In humans, increased cancer prevalence is also associated with reduced genetic diversity [77,78].
Hereditary cancer syndromes connected with inherited mutations in cancer predisposition genes, such as MMR or BRCA genes, account for a considerable proportion (up to 10%) of all cancers in humans [79,80,81,82]. The identification and clinical management of patients with such mutations became routine in modern oncology [83]. For the purpose of our discussion, it is important that hereditary cancer syndromes are much more frequent than noncancer hereditary syndromes, the classical genetic diseases (reviewed in [84]).
Any hereditary trait may be selected and may potentially be evolutionarily meaningful. The fact that many tumors are hereditary suggests that they might participate in the evolution of organisms. For example, germline mutations in BRCA1 and BRCA2 tumor suppressor genes increase the risk of breast and ovarian cancer in humans, and Brca1 and Brca2 genes participate in the embryonic development and differentiation of mammary glands in mice (reviewed in [10]).
When I talk about the evolutionary role of tumors, I mean hereditary tumors. More discussion on hereditary tumors may be found in [14].
3.2. The Widespread Occurrence of Tumors in Multicellular Organisms
Tumors are widespread throughout the phylogenetic tree [14]. New reviews on this topic, which appeared after the publication of the author’s book, support the broad distribution of tumors across the tree of life [85,86].
Cellular oncogenes and tumor suppressor genes are also widespread throughout the phylogenetic tree, and many of them are ancient (oncogenes were already present in sponges). As we have shown in our recent paper [35], cellular oncogenes and tumor suppressor genes are the oldest gene classes, except for housekeeping genes. This means that tumors could have been present during the evolution of multicellular organisms since the very early stages.
The paper of interest to our discussion describes naturally occurring tumors in the pre-bilaterian cnidarian Hydra, with properties similar to cancer in vertebrates [87].
The widespread occurrence of tumors throughout the phylogenetic tree suggests that they may be evolutionarily meaningful.
3.3. A Considerable Portion of Tumors Never Kill Their Hosts
Many (maybe most) tumors never kill their hosts. Benign tumors are widespread in nature. They may represent more than half of all tumors [14]. New publications support this view: up to 80% of mammalian tumors are benign [76,88]. For example, in humans, benign lesions in the nasopharynx accounted for 75% of cases, with malignancies accounting for 25% [89].
At earlier stages of progression, tumors do not kill their hosts. Moreover, the overall direction of tumors in the early stages is toward regression [90]. Tumors can be selected for new functions at the early stages of progression [91].
The list of tumors that do not kill their hosts and could be used in evolution includes fetal, neonatal, and infantile tumors; carcinomas in situ and pseudodiseases; tumors that spontaneously regress; and sustainable tumor masses or extra cell masses in dynamic equilibrium with the organism [14].
When I talk about the evolutionary role of hereditary tumors, I mean benign tumors or tumors at the early stages of progression, or the kinds of tumors mentioned above, but not malignant tumors at the final stages of progression, which kill their hosts.
3.4. Tumors Have Many Features That Could Be Used in Evolution
Tumors are excessive cell masses that are not functionally necessary for the organism. Many unusual genes, which are not expressed in normal tissues, are expressed in tumors. There are also many unusual gene combinations expressed in tumors. Tumor cells can differentiate with the loss of malignancy, and tumors have morphogenetic potential.
All these features are much in demand in evolving multicellular organisms. However, they are not widely appreciated or discussed because the scientific community is more interested in tumors that kill the organism (i.e., malignant tumors, and the medical aspects of malignancy). On the contrary, in this paper, we are interested in tumors that do not kill their hosts, are inherited, and could be used in evolution.
The data exist that tumors were indeed used in evolution (e.g., the positive selection of tumor-associated genes in primate evolution (reviewed in [14])), and examples of organs that possibly originated from tumors [9,10,11,14,92]. The placenta [9,10,14], the mammary gland and prostate [9,10], and mammalian adipose [11] are a few examples of organs that might have developed from tumors. In the author’s book [14], further examples are covered, including the nitrogen-fixing root nodules of legumes, melanomatous cells and macromelanophores of Xiphophorus fish, hoods of Lionhead goldfish, and malignant papillomatosis and symbiovilli of voles.
3.5. “Nothing in Biology Makes Sense except in the Light of Evolution” [95]
Tumors are no exception to Dobzhansky’s maxima, and biologists have started thinking about the place that tumors occupy in evolution. Several monographs have been published over the last decade on tumors and evolution [14,63,96,97]. Until very recently, the role of tumors in evolution was considered completely negative. The “tumors and evolution” relationships were studied mainly in a comparative biological context, or in the context of the evolution of tumor cells in tumor cell populations. However, the hereditary nature of many tumors, the widespread occurrence of tumors, the fact that a considerable proportion of tumors never kill their hosts, and tumor features that could be used in the evolution of multicellular organisms are all features that suggest that hereditary tumors could play some positive role in the evolution of host organisms.
4. The Hypothesis of the Evolutionary Role of Hereditary Tumors (the Main Hypothesis)
The hypothesis of the evolutionary role of hereditary tumors was first formulated in [2], and it has since been developed in a series of articles [3,4,5,6,8].
In the book Evolution by Tumor Neofunctionalization (Chapter 10, Part 10.1), the hypothesis of the evolutionary role of hereditary tumors (or the “evolution by tumor neofunctionalization” hypothesis, or “main hypothesis”) was formulated as follows:
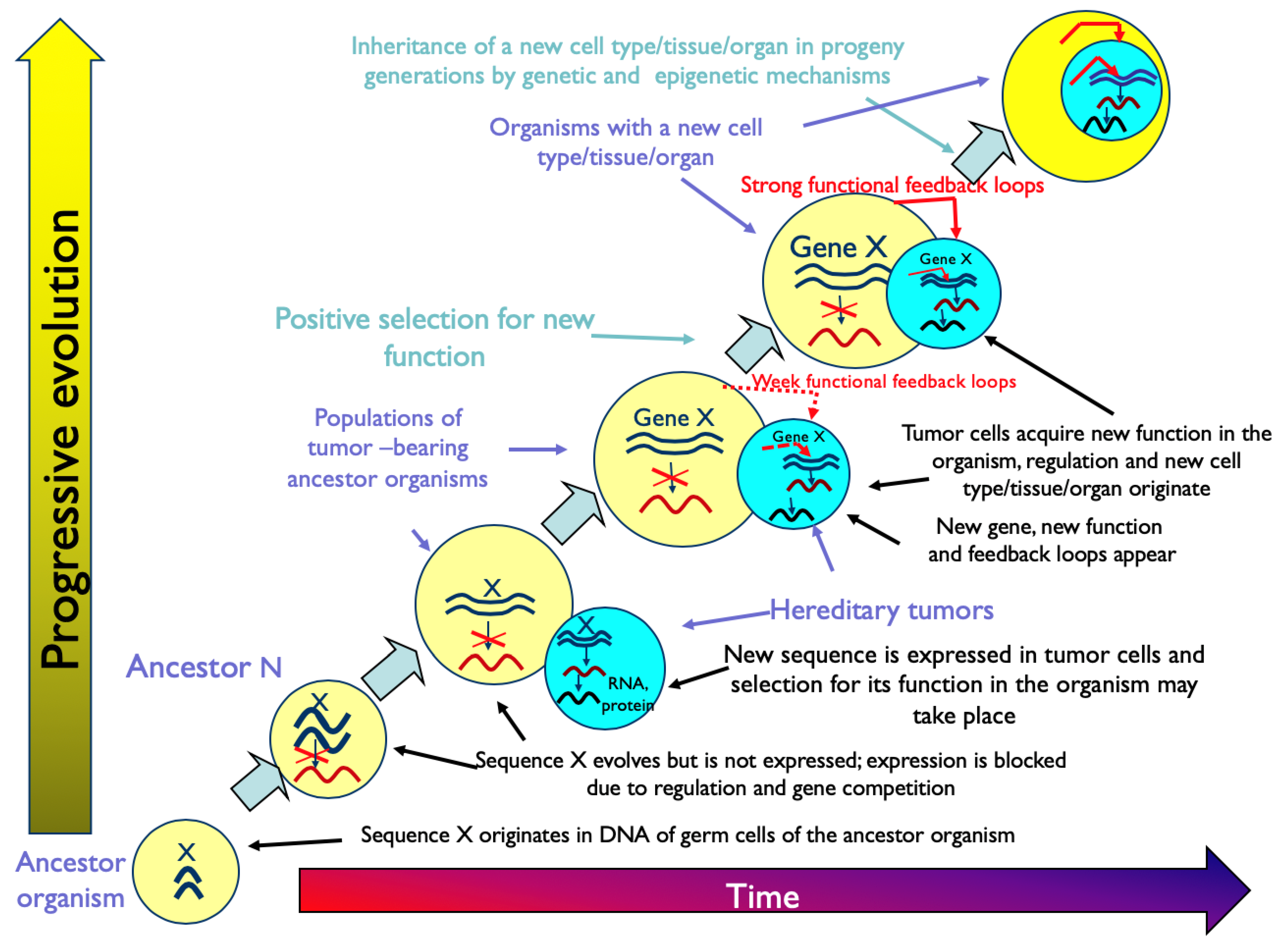
“Tumors are the source of extra cell masses, which may be used in the evolution of multicellular organisms for the expression of evolutionarily new genes, for the origin of new differentiated cell types with novel functions and for building new structures, which constitute evolutionary innovations and morphological novelties. Hereditary tumors may play an evolutionary role by providing conditions (space and resources) for the expression of genes newly evolving in the DNA of germ cells. As a result of the expression of novel genes, tumor cells may acquire new functions and differentiate in new directions, which may lead to the origin of new cell types, tissues and organs. The new cell type is inherited in progeny generations due to genetic and epigenetic mechanisms similar to those for pre-existing cell types. Tumors at the early stages of progression, benign tumors, pseudoneoplasms, and tumor-like processes, which provide evolving multicellular organisms with extra cell masses functionally unnecessary to the organism, are considered potentially evolutionarily meaningful. Malignant tumors at the late stages of progression, however, are not” [14].
Figure 1 represents the graphical description of the hypothesis of evolution by tumor neofunctionalization.
Chapter 10 of the book [14] contains twelve more parts that discuss different aspects of the main hypothesis, including those represented in Figure 1. Evidently, the carcino-evo-devo theory was elaborated around the main hypothesis.
In Part 10.13 of the book, the author wrote:
“Tumors could be regarded as unstable transitory search organs for innovations and expression of evolutionarily novel genes and new combinations of expressed genes, which are not possible in established organs…With the origin of new functions, atypical organs could be stabilized by functional feedback, accumulate a larger proportion of differentiated cells, and become new organs” [14].
Thus, in the book [14], the evolutionary role of hereditary tumors was considered as providing conditions for the expression of evolutionarily new genes and gene combinations. By doing so, we reconciled the genetic theory of morphological evolution, which emphasizes the combinatorial changes in the expression of pre-existing genes [98,99,100,101,102,103,104], and the theory of evolution by gene duplication [105].
5. Nontrivial Predictions of the Carcino-Evo-Devo Theory
An important demand for any new theory in experimental science is the necessity to formulate nontrivial predictions that may be experimentally confirmed. The carcino-evo-devo theory has formulated such predictions.
The author’s previous article [38] was entirely devoted to nontrivial predictions of the carcino-evo-devo theory. The author examined several predictions of the theory and their confirmation in detail. That is why, here, we will discuss them only briefly.
5.1. The Number of Cellular Oncogenes should Correspond to the Number of Cell Types in the Organism [4,6]
This prediction was first formulated when only a few oncogenes had been described, and the anticipated number of oncogenes was estimated to be around twenty. At the same time, about 200 cell types had already been identified. So, the prediction of an order of magnitude larger number of oncogenes was quite bold.
Since the prediction was made, the number of oncogenes described has been continuously increasing. In our 2019 paper [35], we summarized the discovered numbers of oncogenes and cell types. In humans, the described oncogene number was between 245 (TAG database) and 380 [106], and the number of cell types was between 240 and 411, according to different authors (reviewed in [35]). A similar relationship was observed in other multicellular organisms [35].
So, as a result of the efforts of many laboratories, the prediction about the large number of oncogenes and its correspondence to the number of cell types was confirmed.
5.2. The Evolution of Oncogenes, Tumor Suppressors, and Differentiation Gene Classes Should Proceed Concurrently [14]
In [35], we showed that the gene age distribution curves of oncogenes, tumor suppressor genes, and differentiation genes overlap and form a cluster supported by hierarchical cluster analysis with perfect bootstrap reliability. The fact that the gene age distribution curves overlap means that oncogenes, tumor suppressor genes, and differentiation genes evolve concurrently, as predicted. Our results have been confirmed by other authors [71].
5.3. Evolutionarily New and Evolving Genes Should Be Specifically (or Predominantly) Expressed in Tumors (TSEEN Genes)
The laboratory of the author has devoted considerable effort to confirm this prediction. We have published many papers on this issue [12,17,18,19,20,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39]. We have described individual genes, such as PBOV1 [25,27,29] and ELFN1-AS1 [23,26], and whole classes of tumor-specifically expressed, evolutionarily novel (TSEEN) genes, such as CT antigen genes [28] and ncRNA genes [32,35]. Many of our findings have been confirmed by other authors with references to our priorities [107,108,109,110,111,112,113,114]. In [38], the author suggested considering TSEEN genes as a new superclass of novel and evolving genes with tumor-specific or predominant expression, with several classes and families of TSEEN genes, which includes TSEEN genes of various phyla of organisms. A more detailed discussion of TSEEN genes may be found in a recent review by the author [38].
5.4. Human Orthologs of Fish TSEEN Genes Should Acquire Progressive Functions That Are Nonexistent in Fish
We studied this prediction using the KRAS transgenic inducible tumor model in fish [36]. The tumor after regression is a proxy for a new organ evolving from a tumor. The Gene Ontology method was used to investigate human orthologs of fish TSEEN genes that are expressed in tumors following regression. We found many orthologous human genes involved in the development of progressive traits in humans, such as the lung, mammary gland, placenta, ventricular septum, etc., that are nonexistent in fish [36]. The author suggested calling genes that originated from ancestral TSEEN genes and acquired progressive functions “carcino-evo-devo” genes. The discovery of carcino-evo-devo genes is a direct confirmation of the main hypothesis.
5.5. Selection of Tumors for New Functions in the Organism Is Possible
As a result of selection by Chinese breeders, a new organ, the hood, originated on the heads of certain varieties of goldfish. We have shown that such hoods are benign tumors [92]. Goldfish hoods have features of both normal organs (location, symmetrical shape, benign character, and appearance at certain stages of development) and tumors (the capability for unlimited growth and histological peculiarities). The author suggested calling such organs “tumor-like organs” [92].
Examples of the natural selection of tumors for new organismal functions are also known: symbiovilli in voles are the result of selection at the early stages of papillomatosis [91], and macromelanophores in swordtails are the result of sexual selection [115]. Many tumor-related genes in the primate lineage are positively selected (reviewed in [14]).
5.6. Evolutionarily Novel Organs Should Recapitulate Tumor Features in Their Development
The placenta, mammary gland, and prostate are evolutionarily novel to mammals. In [9,10,14], the author discusses many tumor features characteristic of these organs and classifies them as “tumor-like organs”. The term “tumor-like organ” has also been used for the placenta by other authors [116,117].
The ependymal region of the adult human spinal cord is different from that of rodents and primates and may be considered evolutionarily novel to humans. It shows ependymoma-like features morphologically and by gene expression profiling [118]. This example can be added to the list of mammalian tumor-like organs described in [10,11]. The discovery of D. Garcia-Ovejero and co-authors [118] means that, in humans, new cell types and tissues are being evolved, with the participation of tumor-like processes.
Unexpected results have been obtained with mammalian adipose. Mammalian adipose is a complex multidepot organ that consists of several adipose tissues and controls energy metabolism [119,120]. The adipose organ is evolutionarily novel to mammals because of its involvement in thermoregulation [121,122,123,124,125]. The author’s study of the tumor-like features of the mammalian adipose organ discovered many such features, including those belonging to the so-called “hallmarks of cancer” [11]. The most impressive similarities include the capability for unlimited expansion caused by cell hypertrophy and hyperplasia, chronic inflammation, and the infiltration of other organs, which are characteristic of obese adipose. This means that obesity is a tumor-like process, a conclusion that may be important for obesity prevention and treatment in the future. The author further studied whether the carcino-evo-devo genes described in [36] play any role in mammalian adipose, and discovered seven genes that participate in adipose development and form a gene network with mutual influences: LEP, SPRY1, PPARG, ID2, CIDEA, NOTCH1, and ZAG genes. Each of these genes, depending on context, also has tumor-promoting or tumor-suppressing functions [11,12,38]. Such an antagonistic bifunctionality may have fundamental importance for cancer and obesity problems.
5.7. Biological Examples of Cell Types, Tissues, and Organs Originated from Tumors
There are more biological examples of cell types, tissues, and organs that possibly originated from tumors: the nitrogen-fixing root nodules of legumes; the melanomatous cells and macromelanophores of Xyphophorus fishes; malignant papillomatosis and symbiovilli in the stomachs of voles (reviewed in [14]). The human brain recapitulates many features of tumors, especially during childhood and infancy [10,14].
Less characterized but interesting from the point of view of the main hypothesis are the set-aside cells of sea urchins and imaginal discs of insects; the ancestral neural crest and its derivatives; the anterior embryonic field that forms the right ventricle of the vertebrate heart; and morphological novelties and outgrowths in different species (e.g., the prepupal horn primordia of the Onthophagus species resembling papillomas (reviewed in [14])).
Some biological examples were suggested by the audience during the author’s presentations. This means that our concept captured the minds of biologists from different fields and did not contradict their professional experiences.
Malignant papillomatosis and symbiovilli in the stomachs of voles, described by N. Vorontsov [91], were suggested by the audience as examples during the author’s lecture at St. Petersburg University.
During the author’s presentations at Stazione Zoologia Anton Dorn, Naples, and the Zhirmunsky Institute of Marine Biology, Vladivostok, Italian and Russian zoologists independently suggested the following example and provided a reference: adults of sea urchins develop from asymmetrical tumor-like structures located at the left side (left coelomic pouch) of the larvae [126].
An outstanding example of the ependymal region of the adult human spinal cord [118] was advised by Dr. V. Korzh from Singapore University.
Biological examples are very important. In our case, it is difficult to find biological examples because tumor-bearing organisms that played a role in evolution are transient forms. The characterization of unstable elementary particles in physics demonstrates similar difficulties.
5.8. Conclusion about Nontrivial Predictions
The carcino-evo-devo theory has formulated several nontrivial predictions, which have been successfully confirmed. Thus, the carcino-evo-devo theory has predictive power, which is a fundamental requirement for a new theory.
6. Nontrivial Explanations of the Carcino-Evo-Devo Theory and Its Relationship to Other Biological Theories
The purpose of the theory is to explain unexplained (or not completely understood) phenomena, generate new knowledge, and create a scientific worldview. The creation of the new theory reflects the fact that previous theories do not completely resolve certain questions and thus do not satisfy the scientific community. The carcino-evo-devo theory provides a new explanation of the mechanisms of progressive evolution by taking into consideration hereditary tumors and tumor-like processes as active participants in complexity growth. For the first time, the carcino-evo-devo theory introduces the concept of relatively unstable transitional forms in evolution—tumor-bearing organisms—which explains the mechanisms of the origins of complex morphologies and multigene functions. The carcino-evo-devo theory adds the coevolution of neoplastic and normal development to the agenda of the evo-devo theory.
The carcino-evo-devo theory also explains several particular problems of the theory of progressive evolution, evo-devo, and other biological theories, from a new point of view. These explanations are discussed at length in [9,10,11,12,13,14] and are summarized in Table 1. As can be seen from Table 1, the carcino-evo-devo theory, in many cases, provides a new understanding of the existing problems, and it suggests experimentally testable mechanisms.
The carcino-evo-devo theory does not contradict Darwinism or the modern synthesis. In fact, the concept of selection is used throughout the author’s book [14] and in this paper (Section 5.5). However, the carcino-evo-devo theory is devoted to the origins of new cell types, tissues, and organs from hereditary tumors, which represent macroevolutionary events. Some authors believe that Darwinism and the modern synthesis, with their gradualism, do not explain macroevolution [54,55,56]. These authors suggest that different styles of genetic change, which include the rapid reorganization of the genome, are necessary [54,55,56]. Interestingly, the latter idea finds recent support in studies on the somatic evolution of cancer cells that demonstrate macroevolutionary catastrophic events, such as whole-genome doubling, chromosomal chromoplexy, and chromothripsis [141]. The genome architecture concept of H.H. Heng also considers cancer as emerging through macroevolutionary events, which is different from the somatic mutation theory of cancer [142,143].
The present author supposes that the carcino-evo-devo theory is complementary to Darwinism by providing a new mechanism of progressive evolution (i.e., evolution by tumor neofunctionalization). The carcino-evo-devo theory gravitates toward the concept of evolution determined by law, or nomogenesis, as described by Lev Berg [144].
Similarly, the carcino-evo-devo theory is complementary to the theory of evolution by gene duplication [105] in that it begins where the latter theory leaves off by failing to account for the origin of extra cell masses required for the expression of evolutionarily novel genes.
The carcino-evo-devo theory actively uses the ideas of the neutral theory of molecular evolution. Part 10.11 of the author’s book [14] is devoted to the nonadaptive origins of organismal complexity. The neutral theory of molecular evolution “regards protein and DNA polymorphisms as a transient phase of molecular evolution” [145]. In much the same way, the carcino-evo-devo theory regards hereditary tumors and tumor-bearing organisms as transient forms in progressive evolution. According to M. Kimura, “random genetic drift acting on selectively neutral mutations must have played some very important role in macroevolution, in particular in the evolution of gene duplicates” [146]. The carcino-evo-devo theory also deals with macroevolution. In [14], the author reviews the mechanisms of gene duplication and the neutral models of the evolution and maintenance of gene duplicates. Finally, random genetic drift acting on selectively neutral mutants may play a role in biological computational processes (see Section 8 below).
However, the carcino-evo-devo theory describes evolution at three structural levels of organization (molecular, cellular, and multicellular), as well as three main types of biological development (individual, evolutionary, and neoplastic), which sets it apart from the neutral theory, which is the theory of molecular evolution.
Many biological phenomena are explained by the theories that have been discussed above. The carcino-evo-devo theory makes synergistic links with the existing biological science branches by developing new explanations, and by providing a new mechanism of complexity growth (i.e., evolution by tumor neofunctionalization). The carcino-evo-devo theory fills in the spaces between and within the existing biological theories.
Representatives of numerous biological scientific areas largely agree with the carcino-evo-devo idea. As was already indicated, during the author’s presentations, the audience offered various biological examples.
Connections with many biological theories suggest the fundamental nature of the carcino-evo-devo theory. The carcino-evo-devo theory unites three main types of biological development—evolutionary, ontogenetic, and neoplastic development—in one consideration, and that is why it has the potential to become a unifying biological theory.
7. Carcino-Evo-Devo Diagrams
Carcino-evo-devo diagrams were introduced in [9], and more diagrams were added in [10]. At the moment, four diagrams have been published describing different aspects of the carcino-evo-devo theory [9,10].
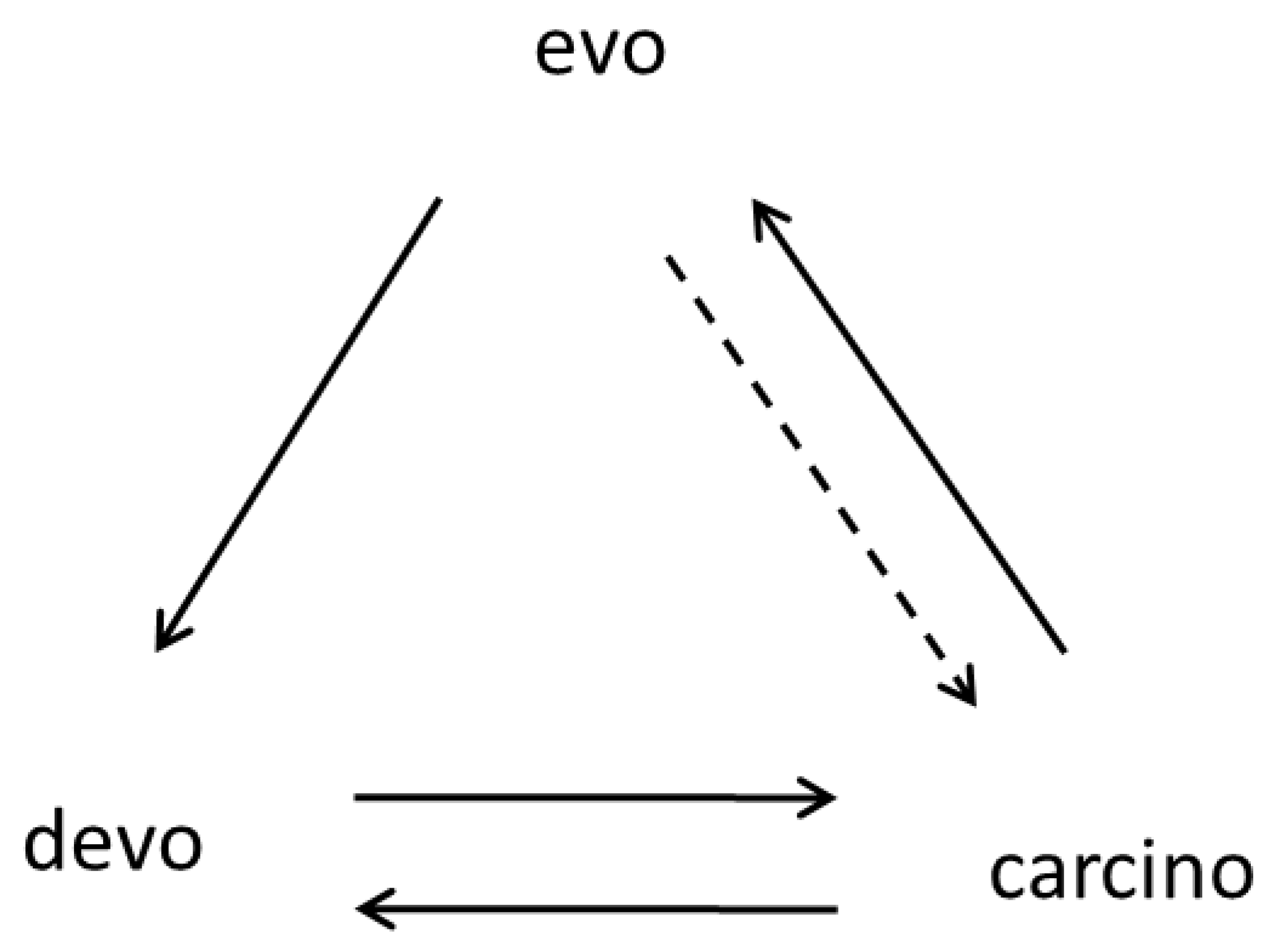
The diagram in Figure 2 shows the interrelationship between three main types of biological development: individual, neoplastic, and evolutionary. From this diagram, it follows that normal ontogenies can participate in progressive evolution only through the intermediate transitionary stage, carcino. The devo→evo arrow is absent, which means that this particular process is prohibited because any deviation from normal development may be lethal (a developmental constraint). This is the fundamental prohibition—a prohibition of the saltatory origins of complex evolutionary innovations and morphological novelties, in which a multitude of genes are involved, from normal ontogenesis. Despite a strong resemblance to the central dogma of molecular biology, the first carcino-evo-devo diagram was built on an independent basis. However, the resemblance is striking and deserves a separate examination.
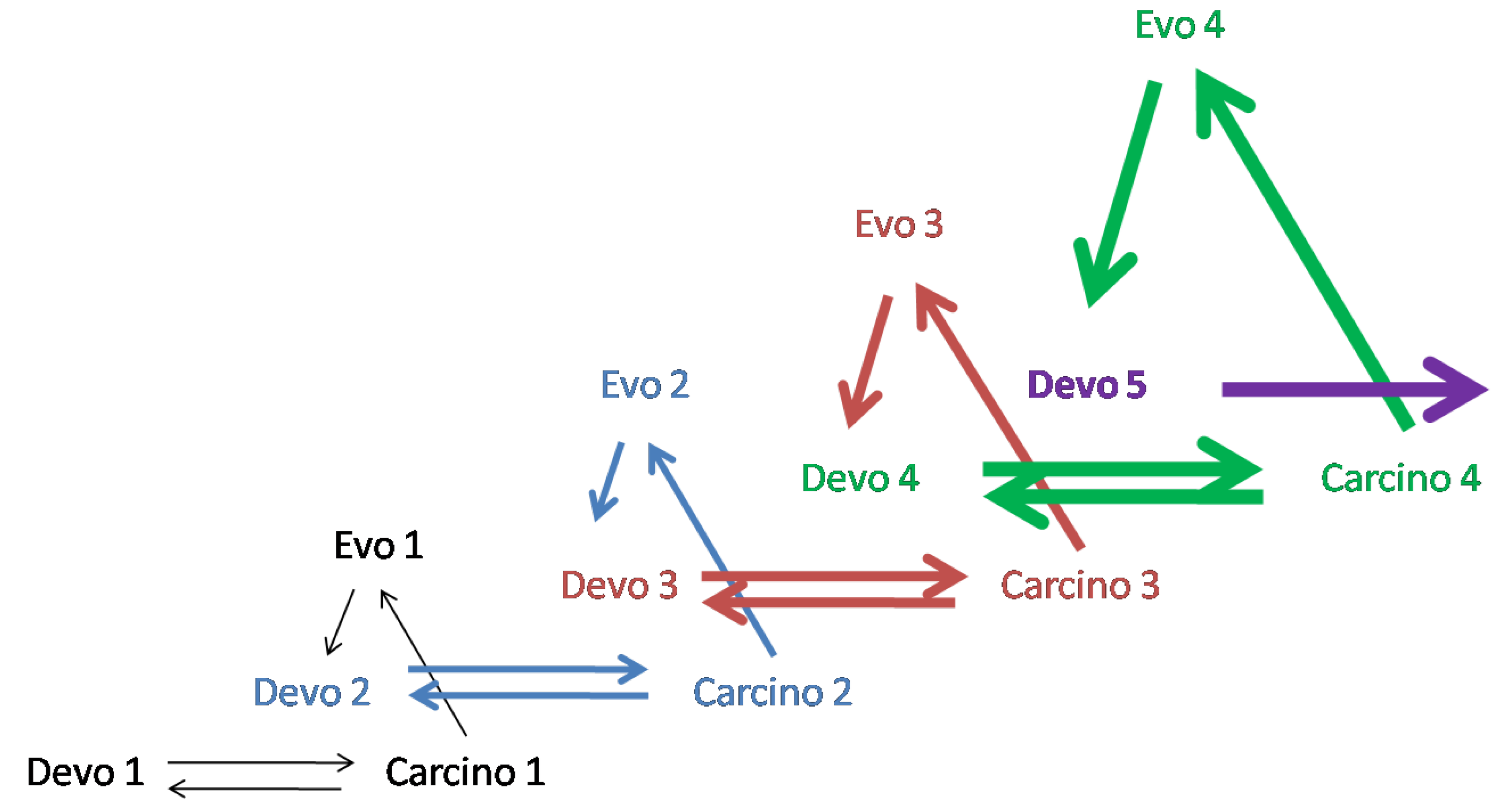
Figure 3 shows four successive steps in the evolution of ontogenesis. From this diagram, it follows that four different tumor processes are used at each of the successive steps of evolution. A more detailed description of the diagrams in Figure 2 and Figure 3 can be found in [9].
The diagrams published in [10] describe the role of tumor-like organs in the evolution of development and the origins of the placenta, mammary gland, and prostate in Eutherian ancestors [10].
The diagram of the central dogma of molecular biology and diagrams describing metabolic, immunological, and signaling pathways already exist and are widely accepted. Carcino-evo-devo diagrams may significantly add to this impressive diagrammatic description of fundamental biological connections. Mathematical category theory, in which objects are linked by arrows, may, in time, produce new generalizations in theoretical biology.
8. Tumors as Search Engines in the Space of Biological Possibilities and Biological Computation
The description of tumors as a “caricature” [147] or “parody” [148] of normal development, as well as the abnormal combination of normal cell features in tumors [52,149], have both sparked the idea that tumors can act as search engines. Tumors as search engines for novel molecular combinations are the focus of a specific section (Part 10.13) in [14].
In [13], the search engine nature of tumors was studied using the concept of biological computational processes [150,151,152,153,154] and Karl Popper’s concept of the possibility space [155].
As shown in [13], the search for new entities in the space of unrealized biological possibilities starts with the DNA computation of new genetic information. An evolutionarily novel gene may be considered a primary DNA computation. An evolutionarily novel gene that has not yet acquired a function may be considered Popper’s “reality in the making”.
The immunoglobulin locus of vertebrates demonstrates an example of DNA computation. Due to V(D)J recombination and subsequent mutations, a variety of immunoglobulins is created that corresponds to the variety of all possible antigens that vertebrates can meet [156,157].
The author suggested that, in much the same way, the evolving vertebrate genome can compute the whole space of possibilities for the evolution of vertebrate morphological structures and their adaptations to different environments [13].
However, DNA computation is not enough to generate the necessary diversity of immunoglobulins. As discussed in [9,13], additional cell masses are necessary (e.g., in amphibians). Additional cell masses can be provided by hereditary tumors [2,9,14].
Hereditary tumors also play the role of search engines in the possibility space of genetic information, which is the next step of biological computation [13]. Tumors search for new gene expression combinations. Many unusual genes and gene combinations, including TSEEN genes, are expressed in tumor cells due to their epigenetic peculiarities. Gene expression patterns that satisfy the compatibility rules described in [2] are frozen by functional feedbacks and natural selection [14].
9. The Increase in Complexity Principle
Earlier, the author discussed the fundamental nature of complexity’s increase at different levels of structural organization [2]. In the author’s book, this is further discussed in relation to the role of tumors in evolution [14]. The increase in complexity in progressive evolution has been discussed in terms of the expression of evolutionarily new genes and gene combinations in the extra cell masses of hereditary tumors [2,14]. In [13], the author discusses complexity growth from the point of view of computational biology.
The author suggests a biocomputational formulation of an old principle of the increase in complexity in living nature:
“The complexity increase in progressive evolution is realized through biological computation of the maximum number of compatible structural entities in evolving lineages of multicellular organisms. Biological computation of complexity increase involves DNA computation in the space of unrealized possibilities; stochastic gene expression and gene competition; compatibility search and incompatibility neutralization at different levels of organization; autonomous search engines and unfolding possibility spaces; unstable transitionary forms; and “freezing” of biologically meaningful constellations of entities, compatible within the ontogeny of multicellular organisms. The complexity of progressively evolving organisms tends to increase to a maximum and can be measured as the number of structural entities from the biological possibility space realized within the ontogeny of the multicellular organism” [13].
Briefly, in living nature, there is a tendency to realize the maximum number of compatible structural entities from the biological possibility space. This tendency is realized with the help of biological computational processes and autonomous search engines. The result is the increase in complexity of multicellular organisms in progressive evolution.
Thus, the concept of biological computational processes explains how complex multigenic functions and organs could originate, which otherwise would be difficult to imagine.
In [13], the author also formulated a new nontrivial prediction:
“After the origin of the genetic code, the evolution of DNA determined the evolution of living matter. That is why the existing informational network of DNA is not only for the storage of information about previous periods of evolutionary development. It also contains information about the future pathways of evolutionary development, which can be studied by bioinformatic approaches with the help of supercomputers. We can perform in silico computing of future functions of evolutionarily novel genes (in silico evolution), which are in the stage of their origin (Popper’s reality in the making) [149]. We plan to use machine learning algorithms to analyze the trajectory of TSEEN gene evolution between fish and humans. The obtained pattern of gene evolution will be used to predict the future functions of human TSEEN genes. Such computing will be similar to the calculation of unoccupied electron orbitals in physics” [13].
We can see that the carcino-evo-devo theory is actively developing, and its biocomputational part formulates new nontrivial explanations and predictions.
10. Conclusions
Carcino-evo-devo is a new biological theory that has predictive power and explains many unexplained phenomena. It suggests new ways of research and stimulates further thinking. As far as three major types of biological development—evolutionary, individual, and neoplastic development—are integrated in carcino-evo-devo consideration, this theory has the potential to become a unifying biological theory. This theory satisfies the criteria of beauty (carcino-evo-devo diagrams) and paradoxicality.
Carcino-evo-devo is also a new oncological theory that claims that hereditary tumors played a positive role in evolution, participating in the origins of new cell types, tissues, and organs. This is why oncological diseases may be considered a recapitulation of ancestral tumors, especially in evolutionarily novel tumor-like organs, such as the mammary gland and prostate. Using what we know about the co-evolution of tumors and normal development, we may understand how to stabilize tumors instead of trying to kill them by all possible means.
The carcino-evo-devo theory predicts the possibility of tumor stabilization by the enforcement of the functional feedbacks in the networks of TSEEN genes.
Funding
This research was funded by the Ministry of Science and Higher Education of the Russian Federation under the strategic academic leadership program ‘Priority 2030′ (Agreement 075-15-2021-1333 dated 30 September 2021).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The author declares no conflict of interest.
Abbreviations
| TSEEN genes | Tumor-specifically expressed, evolutionarily novel genes |
| devo | Normal ontogenies |
| carcino | Ontogenies with neoplastic development |
| evo | Progressive evolution of ontogenies |
References
- Kozlov, A.P. Regulatory Mechanisms as an Expression and the Result of Evolution of Competitive Relations between the Genes. In Salinity Adaptations of Water Organisms; Nauka: Leningrad, Russia, 1976; pp. 237–345. [Google Scholar]
- Kozlov, A.P. Evolution of Living Organisms as a Multilevel Process. J. Theor. Biol. 1979, 81, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Kozlov, A.P. The principles of multilevel development of organisms. In Problems of Analysis of Biological Systems; Maximov, V.N., Ed.; Moscow University Press: Moscow, Russia, 1983; pp. 48–62. [Google Scholar]
- Kozlov, A.P. Gene competition and the possible evolutionary role of tumors and cellular oncogenes. In Theoretical and Mathematical Aspects of Morphogenesis; Presnov, E.V., Maresin, V.M., Zotin, A.I., Eds.; Nauka: Moscow, Russia, 1987; pp. 136–140. [Google Scholar]
- Kozlov, A.P. Conservation principles in the system of molecular-biological laws. Trans. Leningr. Soc. Nat. Sci. 1988, 87, 4–21. [Google Scholar]
- Kozlov, A.P. Gene competition and the possible evolutionary role of tumors. Med. Hypotheses 1996, 46, 81–84. [Google Scholar] [CrossRef] [PubMed]
- Kozlov, A.P. Tumors and evolution. Vopr. Oncol. 2008, 54, 695–705. [Google Scholar]
- Kozlov, A.P. The Possible Evolutionary Role of Tumors in the Origin of New Cell Types. Med. Hypotheses 2010, 74, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Kozlov, A.P. The role of heritable tumors in evolution of development: A new theory of carcino-evo-devo. Acta Nat. 2019, 11, 65–72. [Google Scholar] [CrossRef]
- Kozlov, A.P. Mammalian tumor-like organs.1. The role of tumor-like normal organs and atypical tumor organs in the evolution of development (carcino-evo-devo). Infect. Agents Cancer 2022, 17, 2. [Google Scholar] [CrossRef]
- Kozlov, A.P. Mammalian tumor-like organs. 2. Mammalian adipose has many tumor features and obesity is a tumor-like process. Infect. Agents Cancer 2022, 17, 15. [Google Scholar] [CrossRef]
- Kozlov, A.P. Mammalian adipose has many tumor features, which suggests the possible way of its evolutionary origin. Proceedings of the American Association of Cancer Research Annual Meeting, New Orleans, LA, USA, 8–13 April 2022. Cancer Res. 2022, 82 (Suppl. S12), 6077. [Google Scholar] [CrossRef]
- Kozlov, A.P. Biological computation and compatibility search in the possibility space as the mechanism of complexity increase during progressive evolution. Evol. Bioinform. 2022, 18, 1–5. [Google Scholar] [CrossRef]
- Kozlov, A.P. Evolution by Tumor Neofunctionalization. The Role of Tumors in the Origin of New Cell Types, Tissues and Organs; Elsevier/Academic Press: Amsterdam, Boston, Heidelberg, London, New York, Oxford, Paris, San Diego, San Francisco, Singapore, Sydney, Tokyo, 2014; pp. 1–248. [Google Scholar]
- Kozlov, A.P. Evolution by Tumor Neofunctionalization: Neoplasms as the Factor of Progressive Evolution; Publishing House of Peter the Great Polytechnic University: St. Petersburg, Russia, 2016; pp. 1–263. [Google Scholar]
- Kozlov, A.P. Evolution by Tumor Neofunctionalization: The Role of Tumors in the Origin of New Cell Types, Tissues and Organs; China Science Publishing & Media Ltd. (Science Press): Beijing, China, 2019; pp. 1–200. [Google Scholar]
- Evtushenko, V.I.; Hanson, K.P.; Barabitskaya, O.V.; Emelyanov, A.V.; Reshetnikov, V.L.; Kozlov, A.P. Determination of the upper limit of rat genome expression. Mol. Biol. 1989, 23, 663–67518. [Google Scholar]
- Baranova, A.V.; Lobashev, A.V.; Ivanov, D.V.; Krukovskaya, L.L.; Yankovsky, N.K.; Kozlov, A.P. In silico screening for tumor-specific expressed sequences in human genome. FEBS Lett. 2001, 508, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Krukovskaya, L.L.; Baranova, A.; Tyezelova, T.; Polev, D.; Kozlov, A.P. Experimental study of human expressed sequences newly identified in silico as tumor specific. Tumor Biol. 2005, 26, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Kozlov, A.P.; Galachyants, Y.P.; Dukhovlinov, I.V.; Samusik, N.A.; Baranova, A.V.; Polev, D.E.; Krukovskaya, L.L. Evolutionarily new sequences expressed in tumors. Infect. Agents Cancer 2006, 1, 8. [Google Scholar] [CrossRef] [PubMed]
- Palena, C.; Polev, D.E.; Tsang, K.Y.; Fernando, R.I.; Litzinger, M.; Krukovskaya, L.L.; Baranova, A.V.; Kozlov, A.P.; Schlom, J. The human T-box mesodermal transcription factor Brachyury is a candidate target for T-cell-mediated cancer immunotherapy. Clin. Cancer Res. 2007, 13, 2471–2478. [Google Scholar] [CrossRef] [PubMed]
- Galachyants, Y.; Kozlov, A.P. CDD as a tool for discovery of specifically-expressed transcripts. Russ. J. AIDS Cancer Public Health 2009, 13, 60–61. [Google Scholar]
- Polev, D.; Nosova, J.; Krukovskaya, L.; Baranova, A.; Kozlov, A. Expression of transcripts related to the cluster HS.633957 in human tissues. Mol. Biol. 2009, 43, 97–102. [Google Scholar] [CrossRef]
- Krukovskaia, L.L.; Polev, D.E.; Nosova, I.K.; Baranova, A.V.; Koliubaeva, S.N.; Kozlov, A.P. Investigation of transcription factor Brachyury (T) expression in human normal and tumor tissues. Vopr. Onkol. 2008, 54, 739–743. [Google Scholar]
- Krukovskaja, L.L.; Samusik, N.D.; Shilov, Y.S.; Polev, D.Y.; Kozlov, A.P. Tumor-specific expression of the newly-discovered PBOV1 gene. Vopr. Onkol. 2010, 56, 327–332. [Google Scholar]
- Polev, D.E.; Krukovskaya, L.L.; Kozlov, A.P. Expression of the locus Hs.633957 in human digestive system and tumors. Vopr. Oncol. 2011, 57, 48–49. [Google Scholar]
- Samusik, N.A.; Galachyants, Y.P.; Kozlov, A.P. Analysis of evolutionary novelty of tumor-specifically expressed sequences. Russ. J. Genet. Appl. Res. 2011, 1, 138–148. [Google Scholar] [CrossRef]
- Dobrynin, P.; Matyunina, E.; Malov, S.V.; Kozlov, A.P. The novelty of human cancer/testis antigen encoding genes in evolution. Int. J. Genom. 2013, 2013, e105108. [Google Scholar] [CrossRef] [PubMed]
- Samusik, N.; Krukovskaya, L.; Meln, I.; Shilov, E.; Kozlov, A.P. PBOV1 is a human de novo gene with tumor-specific expression that is associated with a positive clinical outcome of cancer. PLoS ONE 2013, 8, e56162. [Google Scholar] [CrossRef]
- Polev, D.E.; Karnaukhova, J.K.; Krukovskaya, L.L.; Kozlov, A.P. ELFN1-AS1, a novel primate gene with possible microRNA function expressed predominantly in tumors. BioMed Res. Int. 2014, 2014, e398097. [Google Scholar] [CrossRef] [PubMed]
- Matyunina, E.; Emelyanov, A.; Kozlov, A. Evolutionarily novel genes expressed in fish tumors determine progressive evolutionary characters. Cancer Res. 2015, 75 (Suppl. S15), 1927. [Google Scholar] [CrossRef]
- Kozlov, A.P. Expression of evolutionarily novel genes in tumors. Infect. Agents Cancer 2016, 11, 34. [Google Scholar] [CrossRef]
- Krukovskaya, L.L.; Polev, D.E.; Kurbatova, T.V.; Karnaukhova, Y.K.; Kozlov, A.P. The study of the tumor specificity of expression of some evolutionarily novel genes. Vopr. Oncol. 2016, 62, 495–500. [Google Scholar]
- Karnaukhova, Y.K.; Polev, D.E.; Krukovskaya, L.L.; Kozlov, A.P. Study of the expression of gene Orthopedia homeobox in various tumor and normal human tissues. Vopr. Oncol. 2017, 63, 128–134. [Google Scholar]
- Makashov, A.; Malov, S.V.; Kozlov, A.P. Oncogenes, tumor suppressor and differentiation genes represent the oldest human gene classes and evolve concurrently. Sci. Rep. 2019, 9, 16410. [Google Scholar] [CrossRef]
- Matyunina, E.A.; Emelyanov, A.V.; Kurbatova, T.V.; Makashov, A.A.; Mizgirev, I.V.; Kozlov, A.P. Evolutionary novel genes are expressed in transgenic fish tumors and their orthologs are involved in development of progressive traits in humans. Infect. Agents Cancer 2019, 14, 46. [Google Scholar] [CrossRef]
- Kozlov, A.P.; Matyunina, E.A.; Makashov, A.A. The Biomedical Center TSEEN Genes Database. Certificate about Federal Registration of the Database №2021621840. St.Petersburg, 2021. Available online: https://tseendb.org/#/ (accessed on 7 October 2022).
- Kozlov, A.P. The theory of carcino-evo-devo and its non-trivial predictions. Genes 2022, 13, 2347. [Google Scholar] [CrossRef] [PubMed]
- Karnaukhova, I.K.; Polev, D.E.; Krukovskaya, L.L.; Makashov, A.A.; Masharsky, A.E.; Nazarenko, O.V.; Poverennaya, I.V.; Makeev, V.J.; Kozlov, A.P. A new cancer/testis long noncoding RNA, the OTP-AS1 RNA. Sci. Rep. 2022; in press. [Google Scholar]
- Abelev, G.I.; Perova, S.; Kramkova, N.I.; Postnikova, Z.; Irlin, I. Embryonal serum alpha-globulin and its synthesis by transplantable mouse hepatomas. Transplant. Bull. 1963, 1, 174–180. [Google Scholar]
- Tatarinov, Y.S. Presence of embryonic alpha-globulin in the serum of patient with primary hepatocellular carcinoma. Vopr. Med. Khim. 1964, 10, 90–91. [Google Scholar] [PubMed]
- Gold, P.; Freedman, S.O. Demonstration of tumor-specific antigens in human colonic carcinomata by immunological tolerance and absorption techniques. J. Exp. Med. 1965, 121, 439–462. [Google Scholar] [CrossRef] [PubMed]
- Topczewska, J.M.; Postovit, L.M.; Margaryan, N.V.; Sam, A.; Hess, A.R.; Wheaton, W.W.; Nickoloff, B.J.; Topczewski, J.; Hendrix, M.J. Embryonic and tumorigenic pathways converge via Nodal signaling: Role in melanoma aggressiveness. Nat. Med. 2006, 12, 925–932. [Google Scholar] [CrossRef] [PubMed]
- Hendrix, M.J.C.; Lurie, R.H. The convergence of embryonic and cancer signaling pathways: Role in tumor cell plasticity. FASEB Exp. Biol. Part 1 2007, 21, A32. [Google Scholar] [CrossRef]
- Hendrix, M.J.C.; Seftor, E.A.; Seftor, R.E.B.; Kasemeier-Kulesa, J.; Kulesa, P.M.; Postovit, L.M. Reprogramming metastatic tumor cells with embryonic microenvironment. Nat. Rev. Cancer 2007, 7, 246–255. [Google Scholar] [CrossRef]
- Hniz, D.; Schuijers, J.; Lin, C.Y.; Weintraub, A.S.; Abraham, B.J.; Lee, T.I.; Bradner, J.E.; Young, R.A. Covergence of developmental and oncogenic signaling pathways at transcriptional super-enhancers. Mol. Cell 2015, 58, 1–9. [Google Scholar] [CrossRef]
- Hall, B.K. Evo-Devo: Evolutionary developmental mechanisms. Int. J. Dev. Biol. 2003, 47, 491–495. [Google Scholar]
- Durante, F. Nessofisio-patologicotra la strutturadeineimaterni e la genesi di alcunitumorimaligni. Arch. Memo. Observ. Chir. Prat. 1874, 11, 217. [Google Scholar]
- Cohnheim, J. Vorlesungen uber Allgemein Pathologie; Hirschwald: Berlin, Germany, 1877. [Google Scholar]
- Cohnheim, J. Lectures on General Pathology: A Handbook for Practitioners and Students; The New Sydenham Society: London, UK, 1889; Volume 2.
- Severtsov, A.N. Morphological Laws of Evolution. Collection of Works; Publishing House of the Academy of Sciences of the USSR: Moscow, Leningrad, 1949; Volume 5. [Google Scholar]
- Markert, C.L. Neoplasia: A disease of cell differentiation. Cancer Res. 1968, 28, 1908–1914. [Google Scholar] [PubMed]
- Shimkevich, V.M. Malformations and the Origin of Species; Volf Publishing House: St. Petersburg, Russia; Moscow, Russia, 1909. [Google Scholar]
- Goldschmidt, R. The Material Basis of Evolution; Yale University Press: New Haven, CT, USA, 1940. [Google Scholar]
- Gould, S.J. The return of hopeful monsters. Nat. Hist. 1977, 86, 22–30. [Google Scholar]
- Gould, S.J. Is a new and general theory of evolution emerging? Paleobiology 1980, 6, 119–130. [Google Scholar] [CrossRef]
- Anderson, N.G. Evolutionary significance of viral infection. Nature 1970, 227, 1346–1347. [Google Scholar] [CrossRef]
- Williams, G.C.; Nesse, R.M. The dawn of Darwinian medicine. Q. Rev. Biol. 1991, 66, 88–100. [Google Scholar] [CrossRef]
- Ewald, P.W. Evolution of Infectious Disease; Oxford University Press: New York, NY, USA, 1994. [Google Scholar]
- Cairns, J. Mutation, selection and the natural history of cancer. Nature 1975, 255, 197–200. [Google Scholar] [CrossRef]
- Nowell, P.C. The clonal evolution of tumor cell populations. Science 1976, 194, 23–28. [Google Scholar] [CrossRef]
- Nowell, P.C. Mechanisms of tumor progression. Cancer Res. 1986, 46, 2203–2207. [Google Scholar]
- Graham, J. Cancer Selection. The New Theory of Evolution; Aculeus Press Inc.: Lexington, KY, USA, 1992; pp. 1–226. [Google Scholar]
- Gatenby, R.A. A change of strategy in the war on cancer. Nature 2009, 459, 508–509. [Google Scholar] [CrossRef]
- Kuroiwa, Y.; Ando, R.; Kasahara, K.; Nagatani, M.; Yamakawa, S.; Okazaki, S. Transition of historical control data for high incidence tumors in F344 rats. J. Toxicol. Pathol. 2013, 26, 226–230. [Google Scholar] [CrossRef]
- Klaunig, J.E.; Dekant, W.; Plotzke, K.; Scialli, A.R. Biological relevance of decamethylcyclopentasiloxane (D5) induced rat uterine endometrial adenocarcinoma tumorigenesis: Mode of action and relevance to humans. Reg. Toxicol. Pharmacol. 2016, 74, S44–S56. [Google Scholar] [CrossRef] [PubMed]
- Knudson, A.G., Jr. Mutation and cancer: Statistical study of retinoblastoma. Proc. Natl. Acad. Sci. USA 1971, 68, 820–823. [Google Scholar] [CrossRef] [PubMed]
- Knudson, A.G., Jr. Mutagenesis and embryonal carcinogenesis. Natl. Cancer Inst. Monogr. 1979, 51, 19–24. [Google Scholar]
- Knudson, A.G., Jr. Epidemiology of genetically determined cancer. In Genetic Analysis of Tumour Suppression: Ciba Foundation Symposium 142; Bock, G., Marsh, J., Eds.; Wiley: Chichester, UK, 2007; pp. 3–19. [Google Scholar]
- Sondka, Z.; Bamford, S.; Cole, C.G.; Ward, S.A.; Dunham, I.; Forbes, S.A. The COSMIC cancer gene census: Describing genetic dysfunction across all human cancers. Nat. Rev. Cancer 2018, 18, 696–705. [Google Scholar] [CrossRef] [PubMed]
- Tollis, M.; Schneider-Utaka, A.K.; Maley, C.C. The evolution of cancer gene duplications across mammals. Mol. Biol. Evol. 2020, 37, 2875–2886. [Google Scholar] [CrossRef]
- Zhu, B.; Mukherjee, A.; Machiela, M.J.; Song, L.; Hua, X.; Shi, J.; Garcia-Closas, M.; Chanock, S.J.; Chatterjee, N. An investigation of genetic susceptibility risk with somatic mutation burden in breast cancer. Br. J. Cancer 2016, 115, 752–760. [Google Scholar] [CrossRef]
- Liu, Y.; Gusev, A.; Heng, Y.J.; Alexandrov, L.B.; Kraft, P. Somatic mutational profiles and germline polygenic risk scores in human cancer. Genome Med. 2022, 14, 14. [Google Scholar] [CrossRef]
- Leclerc, J.; Flament, C.; Lovecchio, T.; Delattre, L.; Ait Yahya, E.; Baert-Desurmont, S.; Burnichon, N.; Bronner, M.; Cabaret, O.; Lejeune, S.; et al. Diversity of genetic events associated with MLH1 promoter methylation in Lynch syndrome families with heritable constitutional epimutation. Genet. Med. 2018, 20, 1589–1599. [Google Scholar] [CrossRef]
- Madsen, T.; Arnal, A.; Vittecoq, M.; Bernex, F.; Abadie, J.; Labrut, S.; Garcia, D.; Faugere, D.; Lemberger, K.; Beckmann, C.; et al. Cancer prevalence and etiology in wild and captive animals. Ecol. Evol. Cancer 2017, 2017, 11–46. [Google Scholar] [CrossRef]
- Boddy, A.M.; Abegglen, L.M.; Pessier, A.P.; Aktipis, A.; Schiffman, J.D.; Maley, C.C.; Witte, C. Lifetime cancer prevalence and lifetime history traits in mammals. Evol. Med. Public Health 2020, 2020, 187–195. [Google Scholar] [CrossRef]
- Rudan, I.; Rudan, D.; Campbell, H.; Carothers, A.; Wright, A.; Smolej-Narancic, N.; Janicijevic, B.; Jin, L.; Chakraborty, R.; Deka, R.; et al. Inbreeding and risk of late onset complex disease. J. Med. Genet. 2003, 40, 925–932. [Google Scholar] [CrossRef] [PubMed]
- Assie, G.; LaFramboise, T.; Platzer, P.; Eng, C. Frequency of germline genomic homozygosity associated with cancer cases. JAMA 2008, 299, 1437–1445. [Google Scholar] [CrossRef] [PubMed]
- Garber, J.E.; Offit, K. Hereditary cancer predisposition syndromes. J. Clin. Oncol. 2005, 23, 276–292. [Google Scholar] [CrossRef]
- Rahner, N.; Steinke, V. Hereditary cancer syndromes. Dtsch. Arztebl. Int. 2008, 105, 706–714. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q. Cancer predisposition genes: Molecular mechanisms and clinical impact on personalized cancer care: Examples of Lynch and HBOC syndromes. Acta Pharmacol. Sin. 2016, 37, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Vietri, M.T.; D’Elia, G.; Caliendo, G.; Casamassimi, A.; Federico, A.; Passariello, L.; Cioffi, M.; Molinari, A.M. Prevalence of mutations in BRCA and MMR genes in patients affected with hereditary endometrial cancer. Med. Oncol. 2021, 38, 13. [Google Scholar] [CrossRef]
- Ryan, P.D. Genetics of hereditary breast cancer. In Principles of Clinical Cancer Genetics; Chung, D., Haber, D., Eds.; Springer: Boston, MA, USA, 2010; pp. 41–51. [Google Scholar]
- Sokolenko, A.P.; Imyanitov, E.N. Molecular diagnostics in clinical oncology. Front. Mol. Biosci. 2018, 5, 76. [Google Scholar] [CrossRef]
- Aktipis, C.A.; Boddy, A.M.; Jansen, G.; Hibner, U.; Hochberg, M.E.; Maley, C.C.; Wilkinson, G.S. Cancer across the tree of life: Cooperation and cheating in multicellularity. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140219. [Google Scholar] [CrossRef]
- Albuquerque, T.A.F.; Drummond do Val, L.; Doherty, A.; Pedro de Magalhaes, J. From humans to hydra: Patterns of cancer across the tree of life. Biol. Rev. 2018, 93, 1715–1734. [Google Scholar] [CrossRef]
- Domazet-Loso, T.; Klimovich, A.; Anokhin, B.; Anton-Erxleben, F.; Hamm, M.J.; Lange, C.; Bosch, T.C.G. Naturally occurring tumors in the basal metazoan Hydra. Nat. Commun. 2014, 5, 4222. [Google Scholar] [CrossRef]
- Boutry, J.; Tissot, S.; Ujvari, B.; Capp, J.P.; Giraudeau, M.; Nedelcu, A.M.; Thomas, F. The evolution and ecology of benign tumors. BBA—Rev. Cancer 2022, 1877, 188643. [Google Scholar] [CrossRef] [PubMed]
- Satarkar, R.N.; Srikanth, S. Tumors and tumor-like conditions of the nasal cavity, paranasal sinuses, and nasopharynx: A study of 206 cases. Indian J. Cancer 2016, 53, 478–482. [Google Scholar] [CrossRef] [PubMed]
- Clark, W.H. Tumour progression and the nature of cancer. Br. J. Cancer 1991, 64, 631–644. [Google Scholar] [CrossRef] [PubMed]
- Vorontsov, N.N. Macromutations and evolution: Fixation of Goldschmidt’s macromutations as species and genus characters. Papillomatosis and appearance of macrovilli in the rodent stomach. Russ. J. Genet. 2003, 39, 422–426. [Google Scholar]
- Kozlov, A.P.; Zabezhinski, M.A.; Popovich, I.G.; Polev, D.E.; Shilov, E.S.; Murashev, B.V. Hyperplastic skin growth on the head of goldfish--comparative oncology aspects. Vopr. Onkol. 2012, 58, 387–393. [Google Scholar]
- Voskarides, K. Combination of 247 genome-wide association studies reveals high cancer risk as a result of evolutionary adaptation. Mol. Biol. Evol. 2017, 35, 473–485. [Google Scholar] [CrossRef]
- Voskarides, K.; Dweep, H.; Chrysostomou, C. Evidence that DNA repair genes, a family of tumor suppressor genes, are associated with evolution rate and size of genomes. Hum. Genom. 2019, 13, 26. [Google Scholar] [CrossRef]
- Dobzhansky, T. Nothing in biology makes sense except in the light of evolution. Am. Biol. Teach. 1973, 35, 125–129. [Google Scholar] [CrossRef]
- Greaves, M. Cancer. The Evolutionary Legacy; Oxford University Press: Oxford, MS, USA, 2000; pp. 1–288. [Google Scholar]
- Aktipis, A. The Cheating Cell: How Evolution Helps Us Understand and Treat Cancer; Princeton University Press: Princeton, NJ, USA, 2020; pp. 1–256. [Google Scholar]
- King, M.C.; Wilson, A.C. Evolution at two levels in humans and chimpanzees. Science 1975, 188, 107–116. [Google Scholar] [CrossRef]
- Carrol, S.B. Homeotic genes and the evolution of arthropods and chordates. Nature 1995, 376, 479–485. [Google Scholar] [CrossRef]
- Carrol, S.B. Chance and necessity: The evolution of morphological complexity and diversity. Nature 2001, 409, 1102–1109. [Google Scholar] [CrossRef] [PubMed]
- Carrol, S.B. Evolution at two levels: On genes and form. PLoS Biol. 2005, 3, e245. [Google Scholar] [CrossRef] [PubMed]
- Carrol, S.B. Evo-devo and an expanding evolutionary synthesis: A genetic theory of morphological evolution. Cell 2008, 134, 25–36. [Google Scholar] [CrossRef]
- Davidson, E.H. The Regulatory Genome; Elsevier/Academic Press: Amsterdam, The Netherlands; Boston, MA, USA; Berlin/Heidelberg, Germany, 2006. [Google Scholar]
- Prud’homme, B.; Gompel, N.; Carrol, S.B. Emerging principles of regulatory evolution. Proc. Natl. Acad. Sci. USA 2007, 104, 8605–8612. [Google Scholar] [CrossRef] [PubMed]
- Ohno, S. Evolution by Gene Duplication; Springer: New York, NY, USA; Berlin/Heidelberg, Germany, 1970; pp. 1–175. [Google Scholar]
- Domaset-Loso, T.; Tautz, D. Phylostratigraphic tracking of cancer genes suggests a link to the emergence of multicellularity in metazoan. BMC Biol. 2010, 8, 66. [Google Scholar] [CrossRef]
- Zhang, Y.E.; Long, M. New genes contribute to genetic and phenotypic novelties in human evolution. Curr. Opin. Genet. Dev. 2014, 29, 90–96. [Google Scholar] [CrossRef]
- McLysaght, A.; Hurst, L.D. Open questions in the study of de novo genes: What, how and why? Nat. Rev. Genet. 2016, 17, 567. [Google Scholar] [CrossRef]
- Jie, Y.; Ye, L.; Chen, H.; Yu, X.; Cai, L.; He, W.; Fu, Y. ELFN1-AS1 accelerates cell proliferation, invasion and migration via regulating miR-497-3p/CLDN4 axis in ovarian cancer. Bioengineered 2020, 11, 872–882. [Google Scholar] [CrossRef]
- Du, Y.; Hou, Y.; Shi, Y.; Liu, J.; Li, T. Long Non-Coding RNA ELFN1-AS1 Promoted Colon Cancer Cell Growth and Migration via the miR-191-5p/Special AT-Rich Sequence-Binding Protein 1 Axis. Front. Oncol. 2021, 10, 588360. [Google Scholar] [CrossRef]
- Ma, G.; Li, G.; Gou, A.; Xiao, Z.; Xu, Y.; Song, S.; Guo, K.; Liu, Z. Long non-coding RNA ELFN1-AS1 in the pathogenesis of pancreatic cancer. Ann. Transl. Med. 2021, 9, 877. [Google Scholar] [CrossRef]
- Li, Y.; Gan, Y.; Liu, J.; Li, J.; Zhou, Z.; Tian, R.; Sun, R.; Liu, J.; Xiao, Q.; Li, Y.; et al. Downregulation of MEIS1 mediated by ELFN1-AS1/EZH2/DNMT3a axis promotes tumorigenesis and oxaliplatin resistance in colorectal cancer. Signal Transduct. Target. Ther. 2022, 7, 87. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Li, C.; Ma, H.; Yu, D.; Zhang, Y.; Zhang, D.; Su, T.; Wu, J.; Wang, X.; Zhang, L.; et al. Pan-cancer surveys indicate cell cycle-related roles of primate-specific genes in tumors and embryonic cerebrum. Genome Biol. 2022, 23, 251. [Google Scholar] [CrossRef]
- Weisman, C.M. The origins and functions of de novo genes: Against all odds? J. Mol. Evol. 2022, 90, 244–257. [Google Scholar] [CrossRef]
- Fernandez, A.A.; Morris, M.R. Mate choice for more melanin as a mechanism to maintain functional oncogene. Proc. Natl. Acad. Sci. USA 2008, 105, 13503–13507. [Google Scholar] [CrossRef] [PubMed]
- Lala, P.K.; Lee, B.P.; Xu, G.; Chakraborty, C. Human placental trophoblast as an in vitro model for tumor progression. Can. J. Physiol. Pharmacol. 2002, 80, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Lala, P.K.; Nandi, P.; Hadi, A.; Halari, C. A crossroad between placental and tumor biology: What have we learnt? Placenta 2021, 116, 12–30. [Google Scholar] [CrossRef]
- Garcia-Ovejero, D.; Arevalo-Martin, A.; Paniaga-Torija, B.; Florensa-Vila, J.; Ferrer, I.; Grassner, L.; Molina-Holgado, E. The ependymal region of the adult human spinal cord differs from other species and shows ependymoma-like features. Brain 2015, 138, 1583–1597. [Google Scholar] [CrossRef] [PubMed]
- Cinti, S. The Adipose Organ; Editrice Kurtis: Milan, Italy, 2001. [Google Scholar]
- Frayn, K.N.; Karpe, F.; Fielding, B.A.; Macdonald, I.A.; Coppack, S.W. Integrative physiology of human adipose tissue. Int. J. Obes. 2003, 27, 875–888. [Google Scholar] [CrossRef]
- Cinti, S. The Adipose Organ. In Adipose Tissue and Adipokines in Health and Disease; Nutrition and Health; Fantuzzi, G., Mazzone, T., Eds.; Humana Press Inc.: Totowa, NJ, USA, 2007; pp. 3–19. [Google Scholar]
- Pond, C.M. The Evolution of Mammalian Adipose Tissue. In Adipose Tissue Biology; Symonds, M., Ed.; Springer: New York, NY, USA, 2011; pp. 227–269. [Google Scholar]
- Cannon, B.; Nedergaard, J. Brown adipose tissue: Function and physiological significance. Physiol. Rev. 2004, 84, 277–359. [Google Scholar] [CrossRef]
- Gesta, S.; Tseng, Y.H.; Kahn, C.R. Developmental origin of fat: Tracking obesity to its source. Cell 2007, 131, 242–256. [Google Scholar] [CrossRef]
- Mezentseva, N.V.; Kumaratilake, J.S.; Newman, S.A. The brown adipocyte differentiation pathway in birds: An evolutionary road not taken. BMC Biol. 2008, 6, 17. [Google Scholar] [CrossRef]
- Warner, J.F.; Lyons, D.C.; McClay, D.R. Left-Right Asymmetry in the Sea Urchin Embryo: BMP and the Asymmetrical Origins of the Adult. PLoS Biol. 2012, 10, e1001404. [Google Scholar] [CrossRef] [PubMed]
- Severtsov, A.N. The Main Directions of the Evolutionary Process; Dumnov Publishing House: Moscow, Russia, 1925. [Google Scholar]
- Mead, L.S. Transforming over thinking about transitional forms. Evol. Educ. Outreach 2009, 2, 310–314. [Google Scholar] [CrossRef]
- Maynard Smith, J.; Burian, R.; Kauffman, S.; Alberch, P.; Campbell, J.; Goodwin, B.; Lande, R.; Raup, D.; Wolpert, L. Developmental constraints and evolution. Q. Rev. Biol. 1985, 60, 265–287. [Google Scholar]
- Galis, F.; Metz, J.A.J.; von Alphen, J.J.M. Development and evolutionary constraints in animals. Annu. Rev. Ecol. Evol. Syst. 2018, 49, 499–522. [Google Scholar] [CrossRef]
- Moczek, A.P.; Sultan, S.; Foster, S.; Ledón-Rettig, C.; Dworkin, I.; Nijhout, H.F.; Abouheif, E.; Pfennig, D.W. The role of developmental plasticity in evolutionary innovation. Proc. R. Soc. B 2011, 278, 2705–2713. [Google Scholar] [CrossRef] [PubMed]
- West-Eberhard, M.J. Developmental Plasticity and Evolution; Oxford University Press: New York, NY, USA, 2003; pp. 1–816. [Google Scholar]
- Uller, T.; Feiner, N.; Radesma, R.; Jackson, I.S.C.; Rago, F. Developmental plasticity and evolutionary explanations. Evol. Dev. 2020, 22, 47–55. [Google Scholar] [CrossRef]
- Ma, Y.; Zhang, P.; Wang, F.; Yang, J.; Yang, Z.; Qin, H. The relationship between early embryo development and tumorigenesis. J. Cell Mol. Med. 2010, 14, 2697–2701. [Google Scholar] [CrossRef]
- Micalizzi, D.S.; Farabaugh, S.M.; Ford, H.L. Epithelial-mesenchymal transition in cancer: Parallels between normal development and tumor progression. J. Mammary Gland Biol. Neoplasia 2010, 15, 117–134. [Google Scholar] [CrossRef]
- Aiello, N.M.; Stanger, B.Z. Echoes of the embryo: Using the developmental biology toolkit to study cancer. Dis. Model Mech. 2016, 9, 105–114. [Google Scholar] [CrossRef]
- Arendt, D. The evolution of cell types in animals: Emerging principles from molecular studies. Nat. Rev. Genet. 2008, 9, 868–882. [Google Scholar] [CrossRef] [PubMed]
- Arendt, D.; Musser, J.M.; Baker, C.V.H.; Bergman, A.; Cepko, C.; Erwin, D.H.; Pavlicev, M.; Schlosser, G.; Widder, S.; Laubichler, M.D.; et al. The origin and evolution of cell types. Nat. Rev. Genet. 2016, 17, 744–757. [Google Scholar] [CrossRef] [PubMed]
- Flajnik, M.F.; Kasahara, M. Origin and evolution of the adaptive immune system: Genetic events and selective pressures. Nat. Rev. Genet. 2010, 11, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Davies, J.A. Inverse correlation between an organ’s cancer rate and its evolutionary antiquity. Organogenesis 2004, 1, 60–63. [Google Scholar] [CrossRef]
- Vendramin, R.; Litchfield, K.; Swanton, C. Cancer evolution: Darwin and beyond. EMBO J. 2021, 40, e108389. [Google Scholar] [CrossRef]
- Heng, J.; Heng, H.H. Genome chaos, information creation, and cancer emergence: Searching for new framework on the 50th anniversary of the “war on cancer”. Genes 2021, 13, 101. [Google Scholar] [CrossRef]
- Furst, R. The importance of Henry H. Heng’s Genome Archit. Theory. Prog. Biophys. Mol. Diol. 2021, 165, 153–156. [Google Scholar] [CrossRef]
- Berg, L. Nomogenesis or, Evolution Determined by Law; The MIT Press Ltd.: Cambridge, MA, USA, 1969. [Google Scholar]
- Kimura, M. Neutral theory of molecular evolution and the world view of the neutralists. Genome 1989, 31, 24–31. [Google Scholar] [CrossRef]
- Kimura, M. Recent development of the neutral theory from the Wrightian tradition of theoretical population genetics. Proc. Natl. Acad. Sci. USA 1991, 88, 5969–5973. [Google Scholar] [CrossRef]
- Pierce, G.B. On the boundary between development and neoplasia. An interview with Professor G. Barry Pierce. Interview By Juan Arechaga. Int. J. Dev. Biol. 1993, 37, 5–16. [Google Scholar]
- Maenhaut, C.; Dumont, J.E.; Roger, P.P. Cancer stem cells: A reality, a myth, a fuzzy concept or a misnomer? An analysis. Carcinogenesis 2010, 31, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Coggin, J.H.; Anderson, N.G. Cancer, differentiation and embryonic antigens: Some central problems. Adv. Cancer Res. 1974, 19, 105–165. [Google Scholar] [CrossRef] [PubMed]
- Adleman, L.M. Molecular computation of solutions to combinatorial problems. Science 1994, 226, 1021–1024. [Google Scholar] [CrossRef] [PubMed]
- Emmeche, C. The computational notion of life. Theor. Segunda Epoca 1994, 9, 1–30. [Google Scholar]
- Paun, G.; Rozenberg, G.; Salomaa, A. DNA Computing. New Computing Paradigms; Springer: Berlin/Heidelberg, Germany, 1998; pp. 1–409. [Google Scholar]
- Feldman, D.P.; McTague, C.S.; Crutchfield, J.P. The organization of intrinsic computation: Complexity-entropy diagrams and the diversity of natural information processing. Chaos 2008, 18, 043106. [Google Scholar] [CrossRef] [PubMed]
- Grozinger, L.; Amos, M.; Gorochowski, T.E.; Carbonell, P.; Oyarzún, D.A.; Stoof, R.; Fellermann, H.; Zuliani, P.; Tas, H.; Goñi-Moreno, A. Pathways to cellular supremacy in biocomputing. Nat. Commun. 2019, 10, 5250. [Google Scholar] [CrossRef]
- Popper, K.R.A. World of Propensities: Two New Views of Causality; Thoemmes Antiquarian Books Ltd.: Bristol, UK, 1990; pp. 24–329. [Google Scholar]
- Janeway, C.A., Jr.; Travers, P.; Wolpert, M.; Shlomchik, M.J. Immunobiology: The Immune System in Health and Disease, 5th ed.; Garland Science: New York, NY, USA, 2001. [Google Scholar]
- Kovaltsuk, A.; Leem, J.; Keim, S.; Snowden, J.; Dean, C.M.; Krawczyk, K. Observed antibody space: A resource for data mining next-generation sequencing of antibody repertoires. J. Immunol. 2018, 201, 2502–2509. [Google Scholar] [CrossRef]
Figure 1.
Graphical description of the hypothesis of evolution by tumor neofunctionalization. From [14], with permission. Copyright Elsevier (2014).
Figure 1.
Graphical description of the hypothesis of evolution by tumor neofunctionalization. From [14], with permission. Copyright Elsevier (2014).

Figure 2.
Carcino-evo-devo diagram showing the interrelationship between three main types of biological development: individual, neoplastic, and evolutionary (devo: normal ontogenies; carcino: ontogenies with neoplastic development; evo: progressive evolution of ontogenies). Arrows indicate participation in the corresponding process or essential connections. The dotted arrow (evo-->carcino) represents the evolutionary influence on neoplastic development (e.g., anticancer selection, [63]). From [9], with permission.
Figure 2.
Carcino-evo-devo diagram showing the interrelationship between three main types of biological development: individual, neoplastic, and evolutionary (devo: normal ontogenies; carcino: ontogenies with neoplastic development; evo: progressive evolution of ontogenies). Arrows indicate participation in the corresponding process or essential connections. The dotted arrow (evo-->carcino) represents the evolutionary influence on neoplastic development (e.g., anticancer selection, [63]). From [9], with permission.

Figure 3.
Carcino-evo-devo diagram showing four successive steps in the progressive evolution of ontogenesis (marked by different-coloured arrows). Four different tumor processes are used at each of the successive steps of evolution. From [9], with permission.
Figure 3.
Carcino-evo-devo diagram showing four successive steps in the progressive evolution of ontogenesis (marked by different-coloured arrows). Four different tumor processes are used at each of the successive steps of evolution. From [9], with permission.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Nontrivial explanations of the carcino-evo-devo theory.
| What Is Explained | Carcino-Evo-Devo Theory Explanation | Other Existing Theories and Explanations |
|---|---|---|
| The nature of transitional forms in evolution, the origin of major morphological novelties, and complex evolutionary innovations in progressive evolution. | Populations of tumor-bearing organisms may represent transitional forms in progressive evolution. The morphological novelties and complex evolutionary innovations originate by tumor neofunctionalization [9,10,11,14]. | The paleontological record is incomplete because periods of complexity growth were rare and of short duration [127]. Transitional forms are missing, both fossil and imaginary functional intermediates. The new theory is necessary [55,56]. Paleontologists do not expect to find “missing links”, as this is a misconception [128]. |
| The possible way to overcome developmental constraints and developmental plasticity in evolution. | Developmental constraints are surmounted due to the increased plasticity of tumor cells and the transitory nature of tumor-bearing organisms and tumor-like organs [9,10,11,12,13,14]. | Morphological novelties originated in progressive evolution despite the existence of developmental constraints [129]. The breaking of constraints may be associated with relaxed selection [130]. The mechanisms of transitions are not understood, and how developmental plasticity promotes innovations is unclear [131], but evolutionary explanations include developmental plasticity [132,133]. |
| The common features of normal development and oncogenesis. The convergence of embryonic and neoplastic signaling pathways. | Common features of normal development and tumors, and convergence of embryonal and tumor signaling pathways, including carcinoembryonic antigens, follow from tumors’ participation in the evolution of development [9,14]. | These similarities are usually explained by “aberrant” reactivation or the deregulation of embryonal pathways in tumors [134,135,136]. |
| The source of extra cells for the origin of new cell types, and the origin of the neural crest determined cell types. | Our theory explains the origin of new cell types from tumor cells that are not functionally necessary to the organism [9,14]. The neural crest might originate from an ancestral embryonal tumor [14]. | The existing theories of new cell-type origin (e.g., “sister-cell-type model” [137] or “serial sister cell type” [138]) do not explain how additional cells for the origin of the new cell types appeared. |
| The origin of new multigenic functions and organs. | Tumors are considered biocomputational search engines in the space of biological possibilities [13,14]. Tumors search for new gene expression combinations, thereby participating in the origins of new multigenic functions and organs. | The existing theory of gene regulatory networks [103] explains the development of existing organs and functions, but not the origin of new multigenic functions and organs. |
| The evolutionary role of tumors and cellular oncogenes, the phenomenon of CT antigens, and TSEEN genes. | The evolutionary role of tumors and cellular oncogenes might consist in providing evolving multicellular organisms with extra cell masses for the expression of evolutionarily novel genes and gene combinations [4,6,14,35]. CT antigen genes are evolutionarily novel, which is why they are expressed in the testes and in tumors [28]. TSEEN genes have been predicted by the main hypothesis [2,6,14] and described in the laboratory of the author [32,38]. | To the best of the author’s knowledge, the carcino-evo-devo theory is the first theory that claims a positive evolutionary role for hereditary tumors. The author’s laboratory was the first to study the evolutionary novelty of CT antigen genes [28], later confirmed by other authors with reference to our priority [107]. TSEEN genes have also been confirmed by other authors with appropriate reference [113]. |
| The evolutionary origin of the adaptive immune system. | The evolutionary role of tumors helps to explain the origin of the clonal expansion and clonal selection of lymphocytes, as well as of different immune cell types and organs. The ancestral tumor as a search engine could participate in the origin of the combinatorial joining of V, D, and J elements [9,13]. | The existing theories of adaptive immunity’s evolutionary origins focus mainly on molecular events (e.g., the role of the RAG transposon and whole-genome duplications) [139]. |
| The source of extra cell masses where evolutionarily novel genes with progressive functions are expressed. | Hereditary tumors might be the source of extra cell masses where evolutionarily novel genes and gene combinations with progressive functions are expressed [14,32,36]. | The theory of evolution by gene duplication [105], as well as other theories of gene origin and genome evolution, do not address the cellular and multicellular aspects of the problem, focusing only on molecular events. |
| Peculiarities in structure and development of evolutionarily novel organs, and the higher incidence of tumors in such organs. | Tumor-like organs originated from ancestral hereditary tumors and recapitulate some of their features (the concept of tumor-like organs [10,11,12]). | J. Davis, who discovered the higher incidence of tumors in evolutionarily novel organs, such as the mammary gland and prostate, did not explain the phenomenon [140]. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kozlov, A.P. Carcino-Evo-Devo, A Theory of the Evolutionary Role of Hereditary Tumors. Int. J. Mol. Sci. 2023, 24, 8611. https://doi.org/10.3390/ijms24108611
AMA Style
Kozlov AP. Carcino-Evo-Devo, A Theory of the Evolutionary Role of Hereditary Tumors. International Journal of Molecular Sciences. 2023; 24(10):8611. https://doi.org/10.3390/ijms24108611
Chicago/Turabian StyleKozlov, Andrei P. 2023. "Carcino-Evo-Devo, A Theory of the Evolutionary Role of Hereditary Tumors" International Journal of Molecular Sciences 24, no. 10: 8611. https://doi.org/10.3390/ijms24108611
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.