
Outer Membrane Vesicles (OMVs) as Biomedical Tools and Their Relevance as Immune-Modulating Agents against H. pylori Infections: Current Status and Future Prospects


Abstract
:1. Introduction
2. OMVs
2.1. OMV Biogenesis
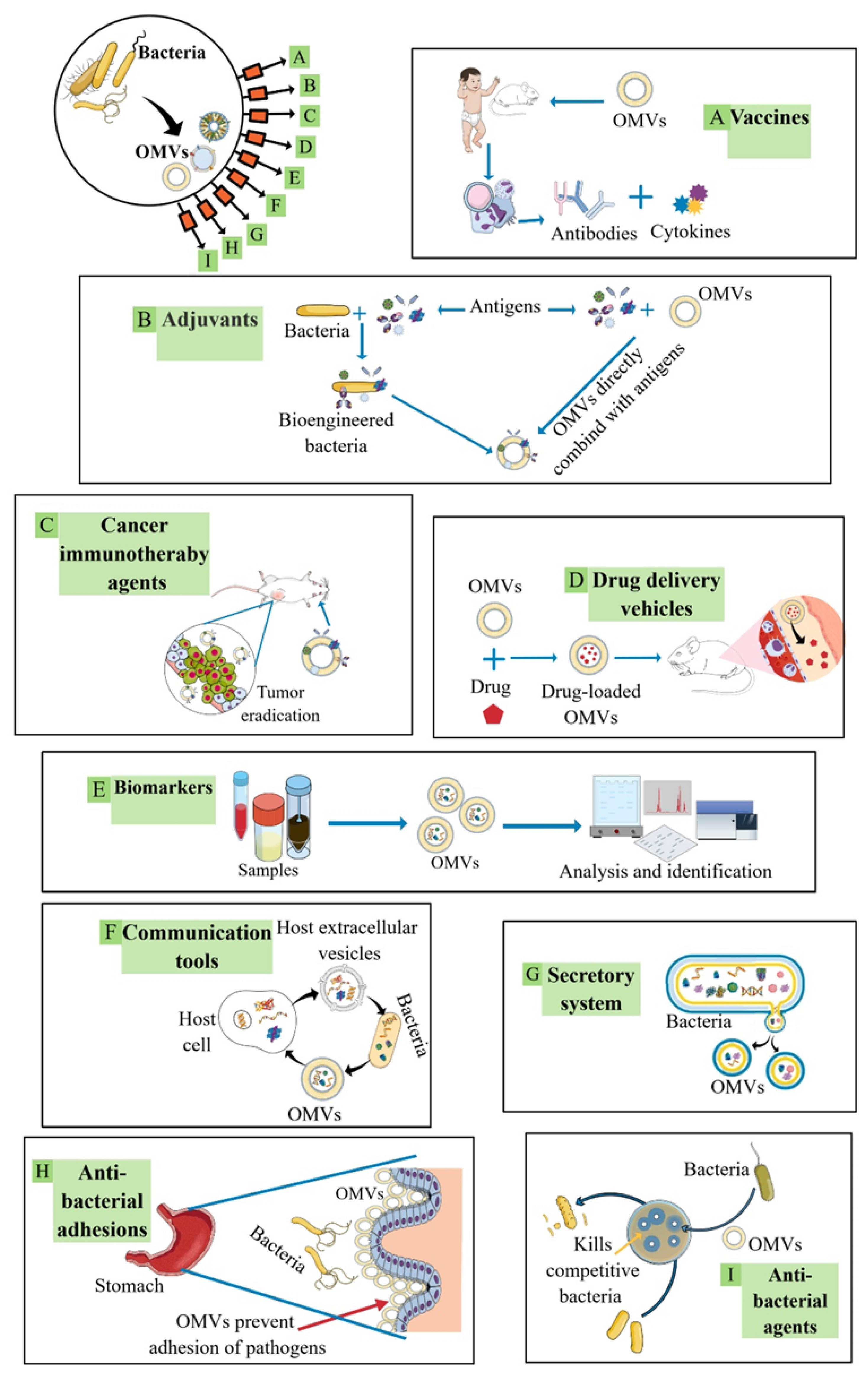
2.2. OMVs in Biomedical Applications
3. Helicobacter pylori (H. pylori)
H. pylori Components and Their Potential in Immune Modulation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| H. pylori Immunogenic Component | Immune Response | Refs. |
|---|---|---|
| Vacuolating cytotoxin A (VacA) |
| [154,155,156,157,158,159] |
| Cytotoxin-associated gene A (CagA) |
| [160,161,162,163,164,165] |
| Urease |
| [166,167,168,169] |
| Flagellum |
| [170,171,172] |
| Catalase |
| [173] |
| Superoxidase dismutase (Sod) |
| [174] |
| Lipopolysaccharide (LPS) |
| [175,176,177,178,179,180,181,182,183,184] |
| Blood group antigen-binding adhesin (BabA) |
| [185,186] |
| Sialic acid-binding adhesin (SabA) |
| [187] |
| Outer inflammatory protein A (OipA) |
| [188,189,190,191,192] |
| Duodenal ulcer promoting gene A (DupA) |
| [193,194,195,196] |
| Adherence-associated lipoprotein A and B (AlpA/AlpB) |
| [197] |
| Induced by contact with epithelium gene A (IceA) |
| [198,199,200] |
| Cholesteryl α-glucosyltransferase (αCgT) |
| [201,202,203,204,205] |
| γ-glutamyl-transpeptidase (Ggt) |
| [206,207,208] |
| Neutrophil-activating protein (Nap) |
| [209,210,211,212,213,214,215,216,217] |
| Heat shock protein 60 (Hsp60) |
| [218,219,220] |
4. OMVs as Immune-Modulating Agents against H. pylori Infections
4.1. Using OMVs Isolated from Standard or Manipulated Growth Conditions or from a Specific Growth Stage
4.2. Using OMVs from Bacterial Strains That Contain Nontoxigenic Virulence Factor Genotypes (e.g., CagA, VacA, DupA) or That Lack Certain Virulence Factors (e.g., CagA-Negative H. pylori Strains, DupA-Negative H. pylori Strains)
4.3. Using OMVs from Probiotic or Commensal Bacteria as Antigen Carriers for the Antigens of Interest
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Leitão, A.L.; Enguita, F.J. Non-Coding RNAs and Inter-Kingdom Communication; Springer: Cham, Switzerland, 2016; pp. 1–251. [Google Scholar]
- Chen, J.; Zhang, H.; Wang, S.; Du, Y.; Wei, B.; Wu, Q.; Wang, H. Inhibitors of bacterial extracellular vesicles. Front. Microbiol. 2022, 13, 319. [Google Scholar] [CrossRef] [PubMed]
- Schwechheimer, C.; Kuehn, M.J. Outer-membrane vesicles from Gram-negative bacteria: Biogenesis and functions. Nat. Rev. Microbiol. 2015, 13, 605–619. [Google Scholar] [CrossRef] [PubMed]
- Sarra, A.; Celluzzi, A.; Bruno, S.P.; Ricci, C.; Sennato, S.; Ortore, M.G.; Casciardi, S.; Del Chierico, F.; Postorino, P.; Bordi, F.; et al. Biophysical characterization of membrane phase transition profiles for the discrimination of outer membrane vesicles (OMVs) from Escherichia coli grown at different temperatures. Front. Microbiol. 2020, 11, 290. [Google Scholar] [CrossRef] [PubMed]
- Zavan, L.; Bitto, N.J.; Johnston, E.L.; Greening, D.W.; Kaparakis-Liaskos, M. Helicobacter pylori growth stage determines the size, protein composition, and preferential cargo packaging of outer membrane vesicles. Proteomics 2019, 19, 1800209. [Google Scholar] [CrossRef]
- Zavan, L.; Bitto, N.J.; Kaparakis-Liaskos, M. Introduction, history, and discovery of bacterial membrane vesicles. In Bacterial Membrane Vesicles; Springer: Berlin/Heidelberg, Germany, 2020; pp. 1–21. [Google Scholar]
- Cecil, J.D.; Sirisaengtaksin, N.; O’Brien-Simpson, N.M.; Krachler, A.M. Outer membrane vesicle-host cell interactions. Microbiol. Spectr. 2019, 7. [Google Scholar] [CrossRef]
- Jones, L.B.; Bell, C.R.; Bibb, K.E.; Gu, L.; Coats, M.T.; Matthews, Q.L. Pathogens and their effect on exosome biogenesis and composition. Biomedicines 2018, 6, 79. [Google Scholar] [CrossRef]
- Kikuchi, Y.; Obana, N.; To Yofuku, M.; Kodera, N.; Soma, T.; Ando, T.; Fukumori, Y.; Nomura, N.; Taoka, A. Diversity of physical properties of bacterial extracellular membrane vesicles revealed through atomic force microscopy phase imaging. Nanoscale 2020, 12, 7950–7959. [Google Scholar] [CrossRef]
- Hasegawa, Y.; Futamata, H.; Tashiro, Y. Complexities of cell-to-cell communication through membrane vesicles: Implications for selective interaction of membrane vesicles with microbial cells. Front. Microbiol. 2015, 6, 633. [Google Scholar] [CrossRef]
- Zhang, W.; Jiang, X.; Bao, J.; Wang, Y.; Liu, H.; Tang, L. Exosomes in pathogen infections: A bridge to deliver molecules and link functions. Front. Microbiol. 2018, 9, 90. [Google Scholar] [CrossRef]
- Jarzab, M.; Posselt, G.; Meisner-Kober, N.; Wessler, S. Helicobacter pylori-derived outer membrane vesicles (OMVs): Role in bacterial pathogenesis? Microorganisms 2020, 8, 1328. [Google Scholar] [CrossRef]
- Biller, S.; Muñoz Marin, M.; Lima, S.; Matinha-Cardoso, J.; Tamagnini, P.; Oliveira, P. Isolation and characterization of cyanobacterial extracellular vesicles. J. Vis. Exp. 2022, 180, e63481. [Google Scholar]
- DeVoe, I.W.; Gilchrist, J.E. Pili on meningococci from primary cultures of nasopharyngeal carriers and cerebrospinal fluid of patients with acute disease. J. Exp. Med. 1975, 141, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhou, H.; Yang, C.; Wu, Y.; Zhou, X.; Liu, H.; Wang, Y. Bacterial outer membrane vesicles as a platform for biomedical applications: An update. J. Control. Release 2020, 323, 253–268. [Google Scholar] [CrossRef] [PubMed]
- Qiang, L.; Hu, J.; Tian, M.; Li, Y.; Ren, C.; Deng, Y.; Jiang, Y. Extracellular vesicles from Helicobacter pylori-infected cells and Helicobacter pylori outer membrane vesicles in atherosclerosis. Helicobacter 2022, 27, e12877. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.A.Q.; Qi, F.; Zheng, R.; Xiao, L.; Abdalla, A.M.E.; Mao, L.; Bakadia, B.M.; Liu, L.; Atta, O.M.; Li, X.; et al. The impact of ExHp-CD (outer membrane vesicles) released from Helicobacter pylori SS1 on macrophage RAW 264.7 cells and their immunogenic potential. Life Sci. 2021, 279, 119644. [Google Scholar] [CrossRef]
- Liu, Q.; Li, X.; Zhang, Y.; Song, Z.; Li, R.; Ruan, H.; Huang, X. Orally-administered outer-membrane vesicles from Helicobacter pylori reduce H. pylori infection via Th2-biased immune responses in mice. Pathog. Dis. 2019, 77, ftz050. [Google Scholar] [CrossRef]
- Furuyama, N.; Sircili, M.P. Outer membrane vesicles (OMVs) produced by Gram-negative bacteria: Structure, functions, biogenesis, and vaccine application. BioMed Res. Int. 2021, 2021, 1490732. [Google Scholar] [CrossRef]
- Wang, S.; Guo, J.; Bai, Y.; Sun, C.; Wu, Y.; Liu, Z.; Liu, X.; Wang, Y.; Wang, Z.; Zhang, Y.; et al. Bacterial outer membrane vesicles as a candidate tumor vaccine platform. Front. immunol. 2022, 13, 5204. [Google Scholar] [CrossRef]
- Kadurugamuwa, J.L.; Beveridge, T.J. Bacteriolytic effect of membrane vesicles from Pseudomonas aeruginosa on other bacteria including pathogens: Conceptually new antibiotics. J. Bacteriol. 1996, 178, 2767–2774. [Google Scholar] [CrossRef]
- Michel, L.V.; Gaborski, T. Outer membrane vesicles as molecular biomarkers for Gram-negative sepsis: Taking advantage of nature’s perfect packages. J. Biol. Chem. 2022, 298, 102483. [Google Scholar] [CrossRef]
- Mashburn, L.M.; Whiteley, M. Membrane vesicles traffic signals and facilitate group activities in a prokaryote. Nature 2005, 437, 422–425. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, Y.; Lo, C.; Zhuang, J.; Angsantikul, P.; Zhang, Q.; Wei, X.; Zhou, Z.; Obonyo, M.; Fang, R.H.; et al. Inhibition of pathogen adhesion by bacterial outer membrane-coated nanoparticles. Angew. Chem. Int. Ed. Engl. 2019, 58, 11404–11408. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Meng, L.; Chen, Y.; Dong, Z.; Peng, Q. Bacterial outer membrane vesicles as potential biological nanomaterials for antibacterial therapy. Acta Biomater. 2022, 140, 102–115. [Google Scholar] [CrossRef]
- Schulz, E.; Goes, A.; Garcia, R.; Panter, F.; Koch, M.; Muller, R.; Fuhrmann, K.; Fuhrmann, G. Biocompatible bacteria-derived vesicles show inherent antimicrobial activity. J. Control. Release 2018, 290, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Turnbull, L.; Toyofuku, M.; Hynen, A.L.; Kurosawa, M.; Pessi, G.; Petty, N.K.; Osvath, S.R.; Cárcamo-Oyarce, G.; Gloag, E.S.; Shimoni, R.; et al. Explosive cell lysis as a mechanism for the biogenesis of bacterial membrane vesicles and biofilms. Nat. Commun. 2016, 7, 11220. [Google Scholar] [CrossRef] [PubMed]
- Roier, S.; Zingl, F.G.; Cakar, F.; Durakovic, S.; Kohl, P.; Eichmann, T.O.; Klug, L.; Gadermaier, B.; Weinzerl, K.; Prassl, R.; et al. A novel mechanism for the biogenesis of outer membrane vesicles in Gram-negative bacteria. Nat. Commun. 2016, 7, 10515. [Google Scholar] [CrossRef]
- Zingl, F.G.; Kohl, P.; Cakar, F.; Leitner, D.R.; Mitterer, F.; Bonnington, K.E.; Rechberger, G.N.; Kuehn, M.J.; Guan, Z.; Reidl, J. Outer membrane vesiculation facilitates surface exchange and in vivo adaptation of Vibrio cholerae. Cell Host Microbe 2020, 27, 225–237.e8. [Google Scholar] [CrossRef]
- Malinverni, J.C.; Silhavy, T.J. An ABC transport system that maintains lipid asymmetry in the gram-negative outer membrane. Proc. Natl. Acad. Sci. USA 2009, 106, 8009–8014. [Google Scholar] [CrossRef]
- Kulp, A.; Kuehn, M.J. Biological functions and biogenesis of secreted bacterial outer membrane vesicles. Annu. Rev. Microbiol. 2010, 64, 163–184. [Google Scholar] [CrossRef]
- Tashiro, Y.; Ichikawa, S.; Nakajima-Kambe, T.; Uchiyama, H.; Nomura, N. Pseudomonas quinolone signal affects membrane vesicle production in not only Gram-negative but also Gram-positive bacteria. Microbes Environ. 2010, 25, 120–125. [Google Scholar] [CrossRef]
- Mashburn-Warren, L.; Howe, J.; Garidel, P.; Richter, W.; Steiniger, F.; Roessle, M.; Brandenburg, K.; Whiteley, M. Interaction of quorum signals with outer membrane lipids: Insights into prokaryotic membrane vesicle formation. Mol. Microbiol. 2008, 69, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, C.; Jagannadham, M.V. Virulence factors are released in association with outer membrane vesicles of Pseudomonas syringae pv. tomato T1 during normal growth. Biochim. Biophys. Acta-Proteins Proteom. 2013, 1834, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Kadurugamuwa, J.L.; Beveridge, T.J. Virulence factors are released from Pseudomonas aeruginosa in association with membrane vesicles during normal growth and exposure to gentamicin: A novel mechanism of enzyme secretion. J. Bacteriol. 1995, 177, 3998–4008. [Google Scholar] [CrossRef] [PubMed]
- Sabra, W.; Lünsdorf, H.; Zeng, A.-P. Alterations in the formation of lipopolysaccharide and membrane vesicles on the surface of Pseudomonas aeruginosa PAO1 under oxygen stress conditions. Microbiology 2003, 149, 2789–2795. [Google Scholar] [CrossRef] [PubMed]
- Schwechheimer, C.; Kulp, A.; Kuehn, M.J. Modulation of bacterial outer membrane vesicle production by envelope structure and content. BMC Microbiol. 2014, 14, 324. [Google Scholar] [CrossRef] [PubMed]
- McBroom, A.J.; Kuehn, M.J. Release of outer membrane vesicles by Gram-negative bacteria is a novel envelope stress response. Mol. Microbiol. 2007, 63, 545–558. [Google Scholar] [CrossRef] [PubMed]
- Deatherage, B.L.; Cookson, B.T. Membrane vesicle release in bacteria, eukaryotes, and archaea: A conserved yet underappreciated aspect of microbial life. Infect. Immun. 2012, 80, 1948–1957. [Google Scholar] [CrossRef]
- Tashiro, Y.; Sakai, R.; Toyofuku, M.; Sawada, I.; Nakajima-Kambe, T.; Uchiyama, H.; Nomura, N. Outer membrane machinery and alginate synthesis regulators control membrane vesicle production in Pseudomonas aeruginosa. J. Bacteriol. 2009, 191, 7509–7519. [Google Scholar] [CrossRef]
- Wensink, J.; Witholt, B. Outer membrane vesicles released by normally growing Escherichia coli contain very little lipoprotein. Eur. J. Biochem. 1981, 116, 331–335. [Google Scholar] [CrossRef]
- Deatherage, B.L.; Lara, J.C.; Bergsbaken, T.; Rassoulian Barrett, S.L.; Lara, S.; Cookson, B.T. Biogenesis of bacterial membrane vesicles. Mol. Microbiol. 2009, 72, 1395–1407. [Google Scholar] [CrossRef]
- Toyofuku, M.; Cárcamo-Oyarce, G.; Yamamoto, T.; Eisenstein, F.; Hsiao, C.-C.; Kurosawa, M.; Gademann, K.; Pilhofer, M.; Nomura, N.; Eberl, L. Prophage-triggered membrane vesicle formation through peptidoglycan damage in Bacillus subtilis. Nat. Commun. 2017, 8, 481. [Google Scholar] [CrossRef]
- Fazal, S.; Lee, R. Biomimetic bacterial membrane vesicles for drug delivery applications. Pharmaceutics 2021, 13, 1430. [Google Scholar] [CrossRef] [PubMed]
- Schetters, S.T.; Jong, W.S.; Horrevorts, S.K.; Kruijssen, L.J.; Engels, S.; Stolk, D.; Daleke-Schermerhorn, M.H.; Garcia-Vallejo, J.; Houben, D.; Unger, W.W. Outer membrane vesicles engineered to express membrane-bound antigen program dendritic cells for cross-presentation to CD8+ T cells. Acta Biomater. 2019, 91, 248–257. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Q.; Xu, J.; Deng, D.; Chao, T.; Li, J.; Zhang, R.; Peng, R.; Liu, Z. Bacteria-derived membrane vesicles to advance targeted photothermal tumor ablation. Biomaterials 2021, 268, 120550. [Google Scholar] [CrossRef] [PubMed]
- Kuerban, K.; Gao, X.; Zhang, H.; Liu, J.; Dong, M.; Wu, L.; Ye, R.; Feng, M.; Ye, L. Doxorubicin-loaded bacterial outer-membrane vesicles exert enhanced anti-tumor efficacy in non-small-cell lung cancer. Acta Pharm. Sin. B 2020, 10, 1534–1548. [Google Scholar] [CrossRef]
- Pan, J.; Li, X.; Shao, B.; Xu, F.; Huang, X.; Guo, X.; Zhou, S. Self-blockade of PD-L1 with bacteria-derived outer-membrane vesicle for enhanced cancer immunotherapy. Adv. Mater. 2022, 34, 2106307. [Google Scholar] [CrossRef]
- Kashyap, D.; Panda, M.; Baral, B.; Varshney, N.; Bhandari, V.; Parmar, H.S.; Prasad, A.; Jha, H.C. Outer membrane vesicles: An emerging vaccine platform. Vaccines 2022, 10, 1578. [Google Scholar] [CrossRef]
- Ayed, Z.; Cuvillier, L.; Dobhal, G.; Goreham, R.V. Electroporation of outer membrane vesicles derived from Pseudomonas aeruginosa with gold nanoparticles. SN Appl. Sci. 2019, 1, 1600. [Google Scholar] [CrossRef]
- Gothelf, A.; Gehl, J. What you always needed to know about electroporation based DNA vaccines. Hum. Vaccines Immunother. 2012, 8, 1694–1702. [Google Scholar] [CrossRef]
- Young, J.L.; Dean, D.A. Chapter Three—Electroporation-mediated gene delivery. In Advances in Genetics; Huang, L., Liu, D., Wagner, E., Eds.; Academic Press: Cambridge, MA, USA, 2015; pp. 49–88. [Google Scholar]
- Podolak, I.; Galanty, A.; Sobolewska, D. Saponins as cytotoxic agents: A review. Phytochem. Rev. 2010, 9, 425–474. [Google Scholar] [CrossRef]
- Fuhrmann, G.; Serio, A.; Mazo, M.; Nair, R.; Stevens, M.M. Active loading into extracellular vesicles significantly improves the cellular uptake and photodynamic effect of porphyrins. J. Control. Release 2015, 205, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Kalani, A.; Chaturvedi, P.; Kamat, P.K.; Maldonado, C.; Bauer, P.; Joshua, I.G.; Tyagi, S.C.; Tyagi, N. Curcumin-loaded embryonic stem cell exosomes restored neurovascular unit following ischemia-reperfusion injury. Int. J. Biochem. Cell Biol. 2016, 79, 360–369. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.T.; Umezaki, K.; Sawada, S.; Mukai, S.-A.; Sasaki, Y.; Harada, N.; Shiku, H.; Akiyoshi, K. Engineering hybrid exosomes by membrane fusion with liposomes. Sci. Rep. 2016, 6, 21933. [Google Scholar] [CrossRef] [PubMed]
- Haney, M.J.; Klyachko, N.L.; Zhao, Y.; Gupta, R.; Plotnikova, E.G.; He, Z.; Patel, T.; Piroyan, A.; Sokolsky, M.; Kabanov, A.V. Exosomes as drug delivery vehicles for Parkinson’s disease therapy. J. Control. Release 2015, 207, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.; Pillai, J. Bacterial membrane vesicles as novel nanosystems for drug delivery. Int. J. Nanomed. 2017, 12, 6329–6341. [Google Scholar] [CrossRef]
- Xu, C.H.; Ye, P.J.; Zhou, Y.C.; He, D.X.; Wei, H.; Yu, C.Y. Cell membrane-camouflaged nanoparticles as drug carriers for cancer therapy. Acta Biomater. 2020, 105, 1–14. [Google Scholar] [CrossRef]
- Ben-Akiva, E.; Meyer, R.A.; Yu, H.; Smith, J.T.; Pardoll, D.M.; Green, J.J. Biomimetic anisotropic polymeric nanoparticles coated with red blood cell membranes for enhanced circulation and toxin removal. Sci. Adv. 2020, 6, eaay9035. [Google Scholar] [CrossRef]
- Zhang, Q.; Fang, R.H.; Gao, W.; Zhang, L. A biomimetic nanoparticle to “Lure and Kill” Phospholipase A2. Angew. Chem. Int. Ed. 2020, 59, 10461–10465. [Google Scholar] [CrossRef]
- Huang, W.; Zhang, Q.; Li, W.; Yuan, M.; Zhou, J.; Hua, L.; Chen, Y.; Ye, C.; Ma, Y. Development of novel nanoantibiotics using an outer membrane vesicle-based drug efflux mechanism. J. Control. Release 2020, 317, 1–22. [Google Scholar] [CrossRef]
- Huang, Y.; Beringhs, A.O.R.; Chen, Q.; Song, D.; Chen, W.; Lu, X.; Fan, T.-H.; Nieh, M.-P.; Lei, Y. Genetically Engineered Bacterial Outer Membrane Vesicles with Expressed Nanoluciferase Reporter for in Vivo Bioluminescence Kinetic Modeling through Noninvasive Imaging. ACS Appl. Bio Mater. 2019, 2, 5608–5615. [Google Scholar] [CrossRef]
- Chen, Q.; Rozovsky, S.; Chen, W. Engineering multi-functional bacterial outer membrane vesicles as modular nanodevices for biosensing and bioimaging. Chem. Commun. 2017, 53, 7569–7572. [Google Scholar] [CrossRef]
- Alves, N.J.; Turner, K.B.; Daniele, M.A.; Oh, E.; Medintz, I.L.; Walper, S.A. Bacterial Nanobioreactors--Directing Enzyme Packaging into Bacterial Outer Membrane Vesicles. ACS Appl. Mater. Interfaces 2015, 7, 24963–24972. [Google Scholar] [CrossRef] [PubMed]
- Gujrati, V.; Kim, S.; Kim, S.-H.; Min, J.J.; Choy, H.E.; Kim, S.C.; Jon, S. Bioengineered bacterial outer membrane vesicles as cell-specific drug-delivery vehicles for cancer therapy. ACS Nano 2014, 8, 1525–1537. [Google Scholar] [CrossRef] [PubMed]
- Gujrati, V.; Prakash, J.; Malekzadeh-Najafabadi, J.; Stiel, A.; Klemm, U.; Mettenleiter, G.; Aichler, M.; Walch, A.; Ntziachristos, V. Bioengineered bacterial vesicles as biological nano-heaters for optoacoustic imaging. Nat. Commun. 2019, 10, 1114. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhao, R.; Cheng, K.; Zhang, K.; Wang, Y.; Zhang, Y.; Li, Y.; Liu, G.; Xu, J.; Xu, J.; et al. Bacterial outer membrane vesicles presenting programmed death 1 for improved cancer immunotherapy via immune activation and checkpoint inhibition. ACS Nano 2020, 14, 16698–16711. [Google Scholar] [CrossRef] [PubMed]
- van de Waterbeemd, B.; Streefland, M.; van der Ley, P.; Zomer, B.; van Dijken, H.; Martens, D.; Wijffels, R.; van der Pol, L. Improved OMV vaccine against Neisseria meningitidis using genetically engineered strains and a detergent-free purification process. Vaccine 2010, 28, 4810–4816. [Google Scholar] [CrossRef]
- Bolandi, Z.; Mokhberian, N.; Eftekhary, M.; Sharifi, K.; Soudi, S.; Ghanbarian, H.; Hashemi, S.M. Adipose derived mesenchymal stem cell exosomes loaded with miR-10a promote the differentiation of Th17 and Treg from naive CD4(+) T cell. Life Sci. 2020, 259, 118218. [Google Scholar] [CrossRef]
- Li, Y.; Li, Q.; Li, D.; Gu, J.; Qian, D.; Qin, X.; Chen, Y. Exosome carrying PSGR promotes stemness and epithelial-mesenchymal transition of low aggressive prostate cancer cells. Life Sci. 2020, 264, 118638. [Google Scholar] [CrossRef]
- Yao, M.; Cui, B.; Zhang, W.; Ma, W.; Zhao, G.; Xing, L. Exosomal miR-21 secreted by IL-1beta-primed-mesenchymal stem cells induces macrophage M2 polarization and ameliorates sepsis. Life Sci. 2020, 264, 118658. [Google Scholar] [CrossRef]
- Chen, T.; Zhang, G.; Kong, L.; Xu, S.; Wang, Y.; Dong, M. Leukemia-derived exosomes induced IL-8 production in bone marrow stromal cells to protect the leukemia cells against chemotherapy. Life Sci. 2019, 221, 187–195. [Google Scholar] [CrossRef]
- Shao, J.; Li, S.; Liu, Y.; Zheng, M. Extracellular vesicles participate in macrophage-involved immune responses under liver diseases. Life Sci. 2020, 240, 117094. [Google Scholar] [CrossRef]
- Bagheri, E.; Abnous, K.; Farzad, S.A.; Taghdisi, S.M.; Ramezani, M.; Alibolandi, M. Targeted doxorubicin-loaded mesenchymal stem cells-derived exosomes as a versatile platform for fighting against colorectal cancer. Life Sci. 2020, 261, 118369. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Luo, J.; Chen, X.; Liu, W.; Chen, T. Cell membrane coating technology: A promising strategy for biomedical applications. Nano-Micro Lett. 2019, 11, 100. [Google Scholar] [CrossRef] [PubMed]
- Dash, P.; Piras, A.M.; Dash, M. Cell membrane coated nanocarriers—An efficient biomimetic platform for targeted therapy. J. Control. Release 2020, 327, 546–570. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.J.; Dawan, J.; Jeon, G.; Yu, T.; He, X.; Ahn, J. The role of bacterial membrane vesicles in the dissemination of antibiotic resistance and as promising carriers for therapeutic agent delivery. Microorganisms 2020, 8, 670. [Google Scholar] [CrossRef] [PubMed]
- Bjune, G.; Høiby, E.; Grønnesby, J.; Arnesen, Ø.; Fredriksen, J.H.; Lindbak, A.; Nøkleby, H.; Rosenqvist, E.; Solberg, L.; Closs, O. Effect of outer membrane vesicle vaccine against group B meningococcal disease in Norway. Lancet 1991, 338, 1093–1096. [Google Scholar] [CrossRef] [PubMed]
- Rosenqvist, E.; Høiby, E.A.; Wedege, E.; Bryn, K.; Kolberg, J.; Klem, A.; Rønnild, E.; Bjune, G.; Nøkleby, H. Human antibody responses to meningococcal outer membrane antigens after three doses of the Norwegian group B meningococcal vaccine. Infect. Immun. 1995, 63, 4642–4652. [Google Scholar] [CrossRef]
- Vernikos, G.; Medini, D. Bexsero® chronicle. Pathog. Glob. Health 2014, 108, 305–316. [Google Scholar] [CrossRef]
- Arnold, R.; Galloway, Y.; McNicholas, A.; O’Hallahan, J. Effectiveness of a vaccination programme for an epidemic of meningococcal B in New Zealand. Vaccine 2011, 29, 7100–7106. [Google Scholar] [CrossRef]
- Nieves, W.; Asakrah, S.; Qazi, O.; Brown, K.A.; Kurtz, J.; AuCoin, D.P.; McLachlan, J.B.; Roy, C.J.; Morici, L.A. A naturally derived outer-membrane vesicle vaccine protects against lethal pulmonary Burkholderia pseudomallei infection. Vaccine 2011, 29, 8381–8389. [Google Scholar] [CrossRef]
- Chen, D.J.; Osterrieder, N.; Metzger, S.M.; Buckles, E.; Doody, A.M.; DeLisa, M.P.; Putnam, D. Delivery of foreign antigens by engineered outer membrane vesicle vaccines. Proc. Natl. Acad. Sci. USA 2010, 107, 3099–3104. [Google Scholar] [CrossRef] [PubMed]
- Tavano, R.; Franzoso, S.; Cecchini, P.; Cartocci, E.; Capecchi, B.; Arico, B.; Papini, E. Self-adjuvant and immune-stimulating activity of the anti-meningococcus B vaccine candidate Neisseria Meningitidis adhesin A as a soluble recombinant antigen (NadAΔ351–405) or as part of bacterial outer membrane vesicles. New Biotechnol. 2009, 25S, S6. [Google Scholar] [CrossRef]
- Rommasi, F. Bacterial-based methods for cancer treatment: What we know and where we are. Oncol. Ther. 2021, 10, 23–54. [Google Scholar] [CrossRef]
- Haiyan, C.; Mengyuan, Z.; Yuteng, Z.; Ziyan, L.; Pan, W.; Han, L. Recent advances on biomedical applications of bacterial outer membrane vesicles. J. Mater. Chem. B 2022, 10, 7384–7396. [Google Scholar]
- Paoli, C.J.; Reynolds, M.A.; Sinha, M.; Gitlin, M.; Crouser, E. Epidemiology and costs of sepsis in the United States—An analysis based on timing of diagnosis and severity level. Crit. Care Med. 2018, 46, 1889–1897. [Google Scholar] [CrossRef]
- Stranieri, I.; Kanunfre, K.A.; Rodrigues, J.C.; Yamamoto, L.; Nadaf, M.I.V.; Palmeira, P.; Okay, T.S. Assessment and comparison of bacterial load levels determined by quantitative amplifications in blood culture-positive and negative neonatal sepsis. Rev. Inst. Med. Trop. Sao Paulo 2018, 60, 1–10. [Google Scholar] [CrossRef]
- Evans, L.; Rhodes, A.; Alhazzani, W.; Antonelli, M.; Coopersmith, C.M.; French, C.; Machado, F.R.; Mcintyre, L.; Ostermann, M.; Prescott, H.C.; et al. Surviving sepsis campaign: International guidelines for management of sepsis and septic Shock 2021. Crit. Care Med. 2021, 47, 1181–1247. [Google Scholar]
- Pilecky, M.; Schildberger, A.; Knabl, L.; Orth-Höller, D.; Weber, V. Influence of antibiotic treatment on the detection of S. aureus in whole blood following pathogen enrichment. BMC Microbiol. 2019, 19, 180. [Google Scholar] [CrossRef]
- Cheng, M.P.; Stenstrom, R.; Paquette, K.; Stabler, S.N.; Akhter, M.; Davidson, A.C.; Gavric, M.; Lawandi, A.; Jinah, R.; Saeed, Z.; et al. Blood culture results before and after antimicrobial administration in patients with severe manifestations of sepsis: A diagnostic study. Ann. Intern. Med. 2019, 171, 547–554. [Google Scholar] [CrossRef]
- Ho, S.-A.; Hoyle, J.A.; Lewis, F.A.; Secker, A.D.; Cross, D.; Mapstone, N.P.; Dixon, M.F.; Wyatt, J.; Tompkins, D.S.; Taylor, G.R. Direct polymerase chain reaction test for detection of Helicobacter pylori in humans and animals. J. Clin. Microbiol. 1991, 29, 2543–2549. [Google Scholar] [CrossRef]
- Clayton, C.; Kleanthous, H.; Coates, P.; Morgan, D.; Tabaqchali, S. Sensitive detection of Helicobacter pylori by using polymerase chain reaction. J. Clin. Microbiol. 1992, 30, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.S.; Golding, G.B. Evolution of HSP70 gene and its implications regarding relationships between archaebacteria, eubacteria, and eukaryotes. J. Mol. Evol. 1993, 37, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Mishra, S.; Rao, G.R.; Jain, A.K.; Dixit, V.K.; Gulati, A.K.; Mahajan, D.; McClelland, M.; Nath, G. Evaluation of nested PCR in detection of Helicobacter pylori targeting a highly conserved gene: HSP60. Helicobacter 2008, 13, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Paul, B.; Kavia Raj, K.; Murali, T.S.; Satyamoorthy, K. Species-specific genomic sequences for classification of bacteria. Comput. Biol. Med. 2020, 123, 103874. [Google Scholar] [CrossRef]
- Koressaar, T.; Jõers, K.; Remm, M. Automatic identification of species-specific repetitive DNA sequences and their utilization for detecting microbial organisms. Bioinformatics 2009, 25, 1349–1355. [Google Scholar] [CrossRef]
- Koressaar, T.; Remm, M. Characterization of species-specific repeats in 613 prokaryotic species. DNA Res. 2012, 19, 219–230. [Google Scholar] [CrossRef]
- Brameyer, S.; Plener, L.; Müller, A.; Klingl, A.; Wanner, G.; Jung, K. Outer membrane vesicles facilitate trafficking of the hydrophobic signaling molecule CAI-1 between Vibrio harveyi cells. J. Bacteriol. 2018, 200, e00740-17. [Google Scholar] [CrossRef]
- Toyofuku, M.; Morinaga, K.; Hashimoto, Y.; Uhl, J.; Shimamura, H.; Inaba, H.; Schmitt-Kopplin, P.; Eberl, L.; Nomura, N. Membrane vesicle-mediated bacterial communication. ISME J. 2017, 11, 1504–1509. [Google Scholar] [CrossRef]
- Koeppen, K.; Hampton, T.H.; Jarek, M.; Scharfe, M.; Gerber, S.A.; Mielcarz, D.W.; Demers, E.G.; Dolben, E.L.; Hammond, J.H.; Hogan, D.A.; et al. A novel mechanism of host-pathogen interaction through sRNA in bacterial outer membrane vesicles. PLoS Pathog. 2016, 12, e1005672. [Google Scholar] [CrossRef]
- Blenkiron, C.; Simonov, D.; Muthukaruppan, A.; Tsai, P.; Dauros, P.; Green, S.; Hong, J.; Print, C.G.; Swift, S.; Phillips, A.R. Uropathogenic Escherichia coli releases extracellular vesicles that are associated with RNA. PLoS ONE 2016, 11, e0160440. [Google Scholar] [CrossRef]
- Tsatsaronis, J.A.; Franch-Arroyo, S.; Resch, U.; Charpentier, E. Extracellular vesicle RNA: A universal mediator of microbial communication? Trends Microbiol. 2018, 26, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Rueter, C.; Bielaszewska, M. Secretion and delivery of intestinal pathogenic Escherichia coli virulence factors via outer membrane vesicles. Front. Cell Infect. Microbiol. 2020, 10, 91. [Google Scholar] [CrossRef] [PubMed]
- Lima, S.; Matinha-Cardoso, J.; Tamagnini, P.; Oliveira, P. Extracellular vesicles: An overlooked secretion system in cyanobacteria. Life 2020, 10, 129. [Google Scholar] [CrossRef] [PubMed]
- Aytar Çelik, P.; Derkuş, B.; Erdoğan, K.; Barut, D.; Blaise Manga, E.; Yıldırım, Y.; Pecha, S.; Çabuk, A. Bacterial membrane vesicle functions, laboratory methods, and applications. Biotechnol. Adv. 2022, 54, 107869. [Google Scholar] [CrossRef]
- Collins, S.M.; Brown, A.C. Bacterial outer membrane vesicles as antibiotic delivery vehicles. Front. Immunol. 2021, 12, 3773. [Google Scholar] [CrossRef]
- Berne, C.; Ellison, C.K.; Ducret, A.; Brun, Y.V. Bacterial adhesion at the single-cell level. Nat. Rev. Microbiol. 2018, 16, 616–627. [Google Scholar] [CrossRef]
- Cusumano, C.K.; Pinkner, J.S.; Han, Z.; Greene, S.E.; Ford, B.A.; Crowley, J.R.; Henderson, J.P.; Janetka, J.W.; Hultgren, S.J. Treatment and prevention of urinary tract infection with orally active FimH inhibitors. Sci. Transl. Med. 2011, 3, 109ra115. [Google Scholar] [CrossRef]
- Turner, L.; Praszkier, J.; Hutton, M.L.; Steer, D.; Ramm, G.; Kaparakis-Liaskos, M.; Ferrero, R.L. Increased outer membrane vesicle formation in a Helicobacter pylori tolB mutant. Helicobacter 2015, 20, 269–283. [Google Scholar] [CrossRef]
- Lundqvist, M.; Stigler, J.; Elia, G.; Lynch, I.; Cedervall, T.; Dawson, K.A. Nanoparticle size and surface properties determine the protein corona with possible implications for biological impacts. Proc. Natl. Acad. Sci. USA 2008, 105, 14265–14270. [Google Scholar] [CrossRef]
- Knowles, M.R.; Boucher, R.C. Mucus clearance as a primary innate defense mechanism for mammalian airways. J. Clin. Investig. 2002, 109, 571–577. [Google Scholar] [CrossRef]
- Bobrovskyy, M.; Willing, S.E.; Schneewind, O.; Missiakas, D. EssH peptidoglycan hydrolase enables Staphylococcus aureus type VII secretion across the bacterial cell wall envelope. J. Bacteriol. 2018, 200, e00268-18. [Google Scholar] [CrossRef] [PubMed]
- Evans, A.G.; Davey, H.M.; Cookson, A.; Currinn, H.; Cooke-Fox, G.; Stanczyk, P.J.; Whitworth, D.E. Predatory activity of Myxococcus xanthus outer-membrane vesicles and properties of their hydrolase cargo. Microbiology 2012, 158, 2742–2752. [Google Scholar] [CrossRef] [PubMed]
- Vasilyeva, N.V.; Tsfasman, I.M.; Suzina, N.E.; Stepnaya, O.A.; Kulaev, I.S. Secretion of bacteriolytic endopeptidase L5 of Lysobacter sp. XL1 into the medium by means of outer membrane vesicles. FEBS Lett. 2008, 275, 3827–3835. [Google Scholar] [CrossRef] [PubMed]
- Gan, Y.; Li, C.; Peng, X.; Wu, S.; Li, Y.; Tan, J.P.K.; Yang, Y.Y.; Yuan, P.; Ding, X. Fight bacteria with bacteria: Bacterial membrane vesicles as vaccines and delivery nanocarriers against bacterial infections. Nanomed. Nanotechnol. Biol. Med. 2021, 35, 102398. [Google Scholar] [CrossRef]
- Tashiro, Y.; Hasegawa, Y.; Shintani, M.; Takaki, K.; Ohkuma, M.; Kimbara, K.; Futamata, H. Interaction of bacterial membrane vesicles with specific species and their potential for delivery to target cells. Front. Microbiol. 2017, 8, 571. [Google Scholar] [CrossRef]
- Borka Balas, R.; Meli, L.E.; Mărginean, C.O. Worldwide prevalence and risk factors of Helicobacter pylori infection in children. Children 2022, 9, 1359. [Google Scholar] [CrossRef]
- Elbehiry, A.; Marzouk, E.; Aldubaib, M.; Abalkhail, A.; Anagreyyah, S.; Anajirih, N.; Almuzaini, A.M.; Rawway, M.; Alfadhel, A.; Draz, A.; et al. Helicobacter pylori infection: Current status and future prospects on diagnostic, therapeutic and control challenges. Antibiotics 2023, 12, 191. [Google Scholar] [CrossRef]
- Shatila, M.; Thomas, A.S. Current and future perspectives in the diagnosis and management of Helicobacter pylori infection. J. Clin. Med. 2022, 11, 5086. [Google Scholar] [CrossRef]
- Kim, N. Prevalence and transmission routes of H. pylori. In Helicobacter pylori; Kim, N., Ed.; Springer: Singapore, 2016; pp. 3–19. [Google Scholar]
- Suerbaum, S.; Michetti, P. Helicobacter pylori infection. N. Engl. J. Med. 2002, 347, 1175–1186. [Google Scholar] [CrossRef]
- Bauer, B.; Meyer, T. The human gastric pathogen Helicobacter pylori and Its association with gastric cancer and ulcer disease. Ulcers 2011, 2011, 340157. [Google Scholar] [CrossRef]
- Akamatsu, T.; Tabata, K.; Hironga, M.; Kawakami, H.; Yyeda, M. Transmission of Helicobacter pylori infection via flexible fiberoptic endoscopy. Am. J. Infect. Control 1996, 24, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Favero, M.S.; Pugliese, G. Infections transmitted by endoscopy: An international problem. Am. J. Infect. Control 1996, 24, 343–345. [Google Scholar] [CrossRef]
- Brown, L.M. Helicobacter Pylori: Epidemiology and routes of transmission. Epidemiol. Rev. 2000, 22, 283–297. [Google Scholar] [CrossRef] [PubMed]
- Nürnberg, M.; Schulz, H.J.; Rüden, H.; Vogt, K. Do conventional cleaning and disinfection techniques avoid the risk of endoscopic Helicobacter pylori Transmission? Endoscopy 2003, 35, 295–299. [Google Scholar] [CrossRef] [PubMed]
- Amieva, M.R.; El-Omar, E.M. Host-bacterial interactions in Helicobacter pylori infection. Gastroenterology 2008, 134, 306–323. [Google Scholar] [CrossRef] [PubMed]
- Boltin, D.; Niv, Y.; Schutte, K.; Schulz, C. Review: Helicobacter pylori and non-malignant upper gastrointestinal diseases. Helicobacter 2019, 24 (Suppl. S1), e12738. [Google Scholar] [CrossRef]
- Venerito, M.; Vasapolli, R.; Rokkas, T.; Delchier, J.C.; Malfertheiner, P. Helicobacter pylori, gastric cancer and other gastrointestinal malignancies. Helicobacter 2017, 22 (Suppl. S1), e12413. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, F.; Covino, M.; Roubaud Baudron, C. Review: Helicobacter pylori and extragastric diseases. Helicobacter 2019, 24 (Suppl. S1), 3204–3221. [Google Scholar] [CrossRef]
- Feng, R.M.; Zong, Y.N.; Cao, S.M.; Xu, R.H. Current cancer situation in China: Good or bad news from the 2018 Global Cancer Statistics? Cancer Commun. 2019, 39, 1–12. [Google Scholar] [CrossRef]
- Hooi, J.K.Y.; Lai, W.Y.; Ng, W.K.; Suen, M.M.Y.; Underwood, F.E.; Tanyingoh, D.; Malfertheiner, P.; Graham, D.Y.; Wong, V.W.S.; Wu, J.C.Y.; et al. Global prevalence of Helicobacter pylori infection: Systematic review and meta-analysis. Gastroenterology 2017, 153, 420–429. [Google Scholar] [CrossRef]
- Savoldi, A.; Carrara, E.; Graham, D.Y.; Conti, M.; Tacconelli, E. Prevalence of antibiotic resistance in Helicobacter pylori: A systematic review and meta-analysis in world health organization regions. Gastroenterology 2018, 155, 1372–1382. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Zhu, Y.; Lu, N.H. Recent progress in Helicobacter pylori treatment. Chin. Med. J. 2020, 133, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Mégraud, F. Synopsis of Antimicrobial Resistance. In Helicobacter pylori; Kim, N., Ed.; Springer: Singapore, 2016; pp. 371–378. [Google Scholar]
- Graham, D.Y.; Mohammadi, M. Synopsis of Antibiotic Treatment. In Helicobacter pylori; Kim, N., Ed.; Springer: Singapore, 2016; pp. 417–426. [Google Scholar]
- Lee, J.Y. Triple Therapy. In Helicobacter pylori; Kim, N., Ed.; Springer: Singapore, 2016; pp. 427–436. [Google Scholar]
- Lee, J.Y. Quadruple Therapy. In Helicobacter pylori; Kim, N., Ed.; Springer: Singapore, 2016; pp. 437–445. [Google Scholar]
- Yoon, H. Sequential Therapy. In Helicobacter pylori; Kim, N., Ed.; Springer: Singapore, 2016; pp. 447–451. [Google Scholar]
- Hwang, S.W. Probiotics. In Helicobacter pylori; Kim, N., Ed.; Springer: Singapore, 2016; pp. 479–485. [Google Scholar]
- Mestre, A.; Sathiya Narayanan, R.; Rivas, D.; John, J.; Abdulqader, M.A.; Khanna, T.; Chakinala, R.C.; Gupta, S. Role of Probiotics in the Management of Helicobacter pylori. Cureus 2022, 14, e26463. [Google Scholar] [CrossRef]
- Eslami, M.; Yousefi, B.; Kokhaei, P.; Jazayeri Moghadas, A.; Sadighi Moghadam, B.; Arabkari, V.; Niazi, Z. Are probiotics useful for therapy of Helicobacter pylori diseases? Comp. Immunol. Microbiol. Infect. Dis. 2019, 64, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, T.; Naito, Y. The role of neutrophils and inflammation in gastric mucosal injury. Free Radic. Res. 2000, 33, 785–794. [Google Scholar] [CrossRef]
- Crabtree, J.E.; Mahony, M.J.; Taylor, J.D.; Heatley, R.V.; Littlewood, J.M.; Tompkins, D.S. Immune responses to Helicobacter pylori in children with recurrent abdominal pain. J. Clin. Pathol. 1991, 44, 768–771. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. The roles of TLRs, RLRs and NLRs in pathogen recognition. Int. Immunol. 2009, 21, 317–337. [Google Scholar] [CrossRef]
- Suarez, G.; Reyes, V.E.; Beswick, E.J. Immune response to H. pylori. World J. Gastroenterol. 2006, 12, 5593–5598. [Google Scholar] [CrossRef]
- Wilson, K.T.; Crabtree, J.E. Immunology of Helicobacter pylori: Insights into the failure of the immune response and perspectives on vaccine studies. Gastroenterology 2007, 133, 288–308. [Google Scholar] [CrossRef]
- Smith, S.M. Role of Toll-like receptors in Helicobacter pylori infection and immunity. World J. Gastrointest. Pathophysiol. 2014, 5, 133–146. [Google Scholar] [CrossRef]
- Meliț, L.E.; Mărginean, C.O.; Mărginean, C.D.; Mărginean, M.O. The relationship between Toll-like receptors and Helicobacter pylori-related gastropathies: Still a controversial topic. J. Immunol. Res. 2019, 2019, 8197048. [Google Scholar] [CrossRef]
- Cadamuro, A.C.T.; Rossi, A.F.T.; Maniezzo, N.M.; Silva, A.E. Helicobacter pylori infection: Host immune response, implications on gene expression and microRNAs. World J. Gastroenterol. 2014, 20, 1424–1437. [Google Scholar] [CrossRef]
- Schmausser, B.; Josenhans, C.; Endrich, S.; Suerbaum, S.; Sitaru, C.; Andrulis, M.; Brändlein, S.; Rieckmann, P.; Müller-Hermelink, H.K.; Eck, M. Downregulation of CXCR1 and CXCR2 expression on human neutrophils by Helicobacter pylori: A new pathomechanism in H. pylori infection? Infect. Immun. 2004, 72, 6773–6779. [Google Scholar] [CrossRef]
- Sukhan, D.S.; Vernygorodskyi, S.V.; Haidukov, N.V.; Ludkevich, H.P. Molecular and genetic aspects of Helicobacter pylori interaction with cells of gastric mucosa. Cytol. Genet. 2020, 54, 147–153. [Google Scholar] [CrossRef]
- Baj, J.; Forma, A.; Sitarz, M.; Portincasa, P.; Garruti, G.; Krasowska, D.; Maciejewski, R. Helicobacter pylori virulence factors—Mechanisms of bacterial pathogenicity in the gastric microenvironment. Cells 2021, 10, 27. [Google Scholar] [CrossRef]
- Sundrud, M.S.; Torres, V.J.; Unutmaz, D.; Cover, T.L. Inhibition of primary human T cell proliferation by Helicobacter pylori vacuolating toxin (VacA) is independent of VacA effects on IL-2 secretion. Proc. Natl. Acad. Sci. USA 2004, 101, 7727–7732. [Google Scholar] [CrossRef]
- Palframan, S.L.; Kwok, T.; Gabriel, K. Vacuolating cytotoxin A (VacA), a key toxin for Helicobacter pylori pathogenesis. Front. Cell Infect. Microbiol. 2012, 2, 92. [Google Scholar] [CrossRef]
- Holland, R.L.; Bosi, K.D.; Harpring, G.H.; Luo, J.; Wallig, M.; Phillips, H.; Blanke, S.R. Chronic in vivo exposure to Helicobacter pylori VacA: Assessing the efficacy of automated and long-term intragastric toxin infusion. Sci. Rep. 2020, 10, 9307. [Google Scholar] [CrossRef]
- Supajatura, V.; Ushio, H.; Wada, A.; Yahiro, K.; Okumura, K.; Ogawa, H.; Hirayama, T.; Ra, C. Cutting edge: VacA, a vacuolating cytotoxin of Helicobacter pylori, directly activates mast cells for migration and production of proinflammatory cytokines. J. Immunol. 2002, 168, 2603–2607. [Google Scholar] [CrossRef]
- Dela Pena-Ponce, M.G.; Jimenez, M.T.; Hansen, L.M.; Solnick, J.V.; Miller, L.A. The Helicobacter pylori type IV secretion system promotes IL-8 synthesis in a model of pediatric airway epithelium via p38 MAP kinase. PLoS ONE 2017, 12, e0183324. [Google Scholar] [CrossRef]
- Jang, S.; Su, H.; Blum, F.C.; Bae, S.; Choi, Y.H.; Kim, A.; Hong, Y.A.; Kim, J.; Kim, J.H.; Gunawardhana, N.; et al. Dynamic expansion and contraction of CagA copy number in Helicobacter pylori impact development of gastric disease. mBio 2017, 8, e01779-16. [Google Scholar] [CrossRef]
- Kido, M.; Watanabe, N.; Aoki, N.; Iwamoto, S.; Nishiura, H.; Maruoka, R.; Ikeda, A.; Azuma, T.; Chiba, T. Dual roles of CagA protein in Helicobacter pylori-induced chronic gastritis in mice. Biochem. Biophys. Res. Commun. 2011, 412, 266–272. [Google Scholar] [CrossRef]
- Eskandari-Nasab, E.; Sepanjnia, A.; Moghadampour, M.; Hadadi-Fishani, M.; Rezaeifar, A.; Asadi-Saghandi, A.; Sadeghi-Kalani, B.; Manshadi, M.D.; Pourrajab, F.; Pourmasoumi, H. Circulating levels of interleukin (IL)-12 and IL-13 in Helicobacter pylori-infected patients, and their associations with bacterial CagA and VacA virulence factors. Scand. J. Infect. Dis. 2013, 45, 342–349. [Google Scholar] [CrossRef]
- Zhang, F.; Chen, C.; Hu, J.; Su, R.; Zhang, J.; Han, Z.; Chen, H.; Li, Y. Molecular mechanism of Helicobacter pylori-induced autophagy in gastric cancer. Oncol. Lett. 2019, 18, 6221–6227. [Google Scholar]
- Bartchewsky, W., Jr.; Martini, M.R.; Masiero, M.; Squassoni, A.C.; Alvarez, M.C.; Ladeira, M.S.; Salvatore, D.; Trevisan, M.; Pedrazzoli, J., Jr.; Ribeiro, M.L. Effect of Helicobacter pylori infection on IL-8, IL-1beta and COX-2 expression in patients with chronic gastritis and gastric cancer. Scand. J. Gastroenterol. 2009, 44, 153–161. [Google Scholar] [CrossRef]
- Sharma, S.A.; Tummuru, M.; Miller, G.G.; Blaser, M.J. Interleukin-8 response of gastric epithelial cell lines to Helicobacter pylori stimulation in vitro. Infect. Immun. 1995, 63, 1681–1687. [Google Scholar] [CrossRef]
- Tanahashi, T.; Kita, M.; Kodama, T.; Yamaoka, Y.; Sawai, N.; Ohno, T.; Mitsufuji, S.; Wei, Y.-P.; Kashima, K.; Imanishi, J. Cytokine expression and production by purified Helicobacter pylori urease in human gastric epithelial cells. Infect. Immun. 2000, 68, 664–671. [Google Scholar] [CrossRef]
- Schmalstig, A.A.; Benoit, S.L.; Misra, S.K.; Sharp, J.S.; Maier, R.J. Noncatalytic antioxidant role for Helicobacter pylori urease. J. Bacteriol. 2018, 200, e00124-18. [Google Scholar]
- Saldinger, P.F.; Porta, N.; Launois, P.; Louis, J.A.; Waanders, G.A.; Bouzourène, H.; Michetti, P.; Blum, A.L.; Corthésy, I.E. Immunization of BALB/c mice with Helicobacter urease B induces a T helper 2 response absent in Helicobacter infection. Gastroenterology 1998, 115, 891–897. [Google Scholar] [CrossRef]
- Hathroubi, S.; Zerebinski, J.; Ottemann, K.M. Helicobacter pylori biofilm involves a multigene stress-biased response, including a structural role for flagella. mBio 2018, 9, e01973-18. [Google Scholar]
- Tang, R.-X.; Luo, D.-J.; Sun, A.-H.; Yan, J. Diversity of Helicobacter pylori isolates in expression of antigens and induction of antibodies. World J. Gastroenterol. 2008, 14, 4816–4822. [Google Scholar] [CrossRef]
- Gu, H. Role of flagella in the pathogenesis of Helicobacter pylori. Curr. Microbiol. 2017, 74, 863–869. [Google Scholar] [CrossRef]
- Miyashita, M.; Joh, T.; Watanabe, K.; Todoroki, I.; Seno, K.; Ohara, H.; Nomura, T.; Miyata, M.; Kasugai, K.; Tochikubo, K.; et al. Immune responses in mice to intranasal and intracutaneous administration of a DNA vaccine encoding Helicobacter pylori-catalase. Vaccine 2002, 20, 2336–2342. [Google Scholar] [CrossRef]
- Stent, A.; Every, A.L.; Chionh, Y.T.; Ng, G.Z.; Sutton, P. Superoxide dismutase from Helicobacter pylori suppresses the production of pro-inflammatory cytokines during in vivo infection. Helicobacter 2018, 23, e12459. [Google Scholar] [CrossRef]
- Chmiela, M.; Miszczyk, E.; Rudnicka, K. Structural modifications of Helicobacter pylori lipopolysaccharide: An idea for how to live in peace. World J. Gastroenterol. 2014, 20, 9882–9897. [Google Scholar] [CrossRef] [PubMed]
- Gonciarz, W.; Krupa, A.; Hinc, K.; Obuchowski, M.; Moran, A.P.; Gajewski, A.; Chmiela, M. The effect of Helicobacter pylori infection and different H. pylori components on the proliferation and apoptosis of gastric epithelial cells and fibroblasts. PLoS ONE 2019, 14, e0220636. [Google Scholar] [CrossRef] [PubMed]
- Sijmons, D.; Guy, A.J.; Walduck, A.K.; Ramsland, P.A. Helicobacter pylori and the role of lipopolysaccharide variation in innate immune evasion. Front. Immunol. 2022, 13, 868225. [Google Scholar] [CrossRef] [PubMed]
- Semeraro, N.; Montemurro, P.; Piccoli, C.; Muoio, V.; Colucci, M.; Giuliani, G.; Fumarola, D.; Pece, S.; Moran, A.P. Effect of Helicobacter pylori lipopolysaccharide (LPS) and LPS derivatives on the production of tissue factor and plasminogen activator inhibitor type 2 by human blood mononuclear cells. J. Infect. Dis. 1996, 174, 1255–1260. [Google Scholar] [CrossRef]
- Hansen, P.; Petersen, S.B.; Varming, K.; Nielsen, H. Helicobacter pylori additive effects of Helicobacter pylori lipopolysaccharide and proteins in monocyte inflammatory responses. Scand. J. Gastroenterol. 2002, 37, 765–771. [Google Scholar] [CrossRef]
- Basak, C.; Pathak, S.K.; Bhattacharyya, A.; Mandal, D.; Pathak, S.; Kundu, M. NF-kappaB- and C/EBPbeta-driven interleukin-1beta gene expression and PAK1-mediated caspase-1 activation play essential roles in interleukin-1beta release from Helicobacter pylori lipopolysaccharide-stimulated macrophages. J. Biol. Chem. 2005, 280, 4279–4288. [Google Scholar] [CrossRef]
- Xu, L.; Gong, C.; Li, G.; Wei, J.; Wang, T.; Meng, W.; Shi, M.; Wang, Y. Ebselen suppresses inflammation induced by Helicobacter pylori lipopolysaccharide via the p38 mitogen-activated protein kinase signaling pathway. Mol. Med. Rep. 2018, 17, 6847–6851. [Google Scholar] [PubMed]
- Taylor, J.M.; Ziman, M.E.; Huff, J.L.; Moroski, N.M.; Vajdy, M.; Solnick, J.V. Helicobacter pylori lipopolysaccharide promotes a Th1 type immune response in immunized mice. Vaccine 2006, 24, 4987–4994. [Google Scholar] [CrossRef] [PubMed]
- Shimoyama, A.; Saeki, A.; Tanimura, N.; Tsutsui, H.; Miyake, K.; Suda, Y.; Fujimoto, Y.; Fukase, K. Chemical synthesis of Helicobacter pylori lipopolysaccharide partial structures and their selective proinflammatory responses. Chemistry 2011, 17, 14464–14474. [Google Scholar] [CrossRef] [PubMed]
- Bliss, C.M., Jr.; Golenbock, D.T.; Keates, S.; Linevsky, J.K.; Kelly, C.N.P. Helicobacter pylori lipopolysaccharide binds to CD14 and stimulates release of interleukin-8, epithelial neutrophil-activating peptide 78, and monocyte chemotactic protein 1 by human monocytes. Infect. Immun. 1998, 66, 5357–5363. [Google Scholar] [CrossRef]
- Shahi, H.; Reiisi, S.; Bahreini, R.; Bagheri, N.; Salimzadeh, L.; Shirzad, H. Association between Helicobacter pylori cagA, babA2 virulence factors and gastric mucosal Interleukin-33 mRNA expression and clinical outcomes in dyspeptic patients. Int. J. Mol. Cell Med. 2015, 4, 227–234. [Google Scholar]
- Rad, R.; Gerhard, M.; Lang, R.; Scho, M.; Rösch, T.; Schepp, W.; Becker, I.; Wagner, H.; Prinz, C. The Helicobacter pylori blood group antigen-binding adhesin facilitates bacterial colonization and augments a nonspecific immune response. J. Immunol. 2002, 168, 3033–3041. [Google Scholar] [CrossRef]
- Unemo, M.; Aspholm-Hurtig, M.; Ilver, D.; Bergström, J.; Borén, T.; Danielsson, D.; Teneberg, S. The sialic acid binding SabA adhesin of Helicobacter pylori is essential for nonopsonic activation of human neutrophils. J. Biol. Chem. 2005, 280, 15390–15397. [Google Scholar] [CrossRef]
- Yamaoka, Y. Pathogenesis of Helicobacter pylori-related gastroduodenal diseases from molecular epidemiological studies. Gastroenterol. Res. Pract. 2012, 2012, 371503. [Google Scholar] [CrossRef]
- Alm, R.A.; Ling, L.S.; Moir, D.T.; King, B.L.; Brown, E.D.; Doig, P.C.; Smith, D.R.; Noonan, B.; Guild, B.C.; de Jonge, B.L.; et al. Genomic-sequence comparison of two unrelated isolates of the human gastric pathogen Helicobacter pylori. Nature 1999, 397, 176–180. [Google Scholar] [CrossRef]
- Wroblewski, L.E.; Peek, R.M., Jr.; Wilson, K.T. Helicobacter pylori and gastric cancer: Factors that modulate disease risk. Clin. Microbiol. Rev. 2010, 23, 713–739. [Google Scholar] [CrossRef]
- Teymournejad, O.; Mobarez, A.M.; Hassan, Z.M.; Moazzeni, S.M.; Ahmadabad, H.N. In vitro suppression of dendritic cells by Helicobacter pylori OipA. Helicobacter 2014, 19, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Yamaoka, Y.; Kudo, T.; Lu, H.; Casola, A.; Brasier, A.R.; Graham, D.Y. Role of interferon-stimulated responsive element-like element in interleukin-8 promoter in Helicobacter pylori infection. Gastroenterology 2004, 126, 1030–1043. [Google Scholar] [CrossRef] [PubMed]
- Youssefi, M.; Ghazvini, K.; Farsiani, H.; Tafaghodi, M.; Keikha, M. A systematic review and meta-analysis of outcomes of infection with Helicobacter pylori dupA+ strains in Iranian patients. Gene Rep. 2020, 19, 100650. [Google Scholar] [CrossRef]
- Lu, H.; Hsu, P.I.; Graham, D.Y.; Yamaoka, Y. Duodenal ulcer promoting gene of Helicobacter pylori. Gastroenterology 2005, 128, 833–848. [Google Scholar] [CrossRef]
- Jung, S.W.; Sugimoto, M.; Shiota, S.; Graham, D.Y.; Yamaoka, Y. The intact dupA cluster is a more reliable Helicobacter pylori virulence marker than dupA alone. Infect. Immun. 2012, 80, 381–387. [Google Scholar] [CrossRef]
- Hussein, N.R.; Argent, R.H.; Marx, C.K.; Patel, S.R.; Robinson, K.; Atherton, J.C. Helicobacter pylori dupA is polymorphic, and its active form induces proinflammatory cytokine secretion by mononuclear cells. J. Infect. Dis. 2010, 202, 261–269. [Google Scholar] [CrossRef]
- Lu, H.; Wu, J.Y.; Beswick, E.J.; Ohno, T.; Odenbreit, S.; Haas, R.; Reyes, V.E.; Kita, M.; Graham, D.Y.; Yamaoka, Y. Functional and intracellular signaling differences associated with the Helicobacter pylori AlpAB adhesin from Western and East Asian strains. J. Biol. Chem. 2007, 282, 6242–6254. [Google Scholar] [CrossRef]
- Xu, Q.; Blaser, M.J. Promoters of the CATG-specific methyltransferase gene hpyIM differ between IceA1 and IceA2 Helicobacter pylori strains. J. Bacteriol. 2001, 183, 3875–3884. [Google Scholar] [CrossRef]
- Dabiri, H.; Jafari, F.; Baghaei, K.; Shokrzadeh, L.; Abdi, S.; Pourhoseingholi, M.A.; Mohammadzadeh, A. Prevalence of Helicobacter pylori VacA, CagA, CagE, OipA, IceA, BabA2 and BabB genotypes in Iranian dyspeptic patients. Microb. Pathog. 2017, 105, 226–230. [Google Scholar] [CrossRef]
- Chiurillo, M.A.; Moran, Y.; Cañas, M.; Valderrama, E.; Alvarez, A.; Armanie, E. Combination of Helicobacter pylori-iceA2 and proinflammatory interleukin-1 polymorphisms is associated with the severity of histological changes in Venezuelan chronic gastritis patients. FEMS Immunol. Med. Microbiol. 2010, 59, 170–176. [Google Scholar] [CrossRef]
- Beigier-Bompadre, M.; Moos, V.; Belogolova, E.; Allers, K.; Schneider, T.; Churin, Y.; Ignatius, R.; Meyer, T.F.; Aebischer, T. Modulation of the CD4+ T-cell response by Helicobacter pylori depends on known virulence factors and bacterial cholesterol and cholesterol α-glucoside content. J. Infect. Dis. 2011, 204, 1339–1348. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.J.; Cheng, W.C.; Cheng, H.H.; Lai, C.H.; Wang, W.C. Helicobacter pylori cholesteryl glucosides interfere with host membrane phase and affect type IV secretion system function during infection in AGS cells. Mol. Microbiol. 2012, 83, 67–84. [Google Scholar] [CrossRef] [PubMed]
- Morey, P.; Pfannkuch, L.; Pang, E.; Boccellato, F.; Sigal, M.; Imai-Matsushima, A.; Dyer, V.; Koch, M.; Mollenkopf, H.-J.; Schlaermann, P. Helicobacter pylori depletes cholesterol in gastric glands to prevent interferon gamma signaling and escape the inflammatory response. Gastroenterology 2018, 154, 1391–1404. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.H.; Huang, J.C.; Cheng, H.H.; Wu, M.C.; Huang, M.Z.; Hsu, H.Y.; Chen, Y.A.; Hsu, C.Y.; Pan, Y.J.; Chu, Y.T.; et al. Helicobacter pylori cholesterol glucosylation modulates autophagy for increasing intracellular survival in macrophages. Cell Microbiol. 2018, 20, e12947. [Google Scholar] [CrossRef] [PubMed]
- Du, S.Y.; Wang, H.J.; Cheng, H.H.; Chen, S.D.; Wang, L.H.; Wang, W.C. Cholesterol glucosylation by Helicobacter pylori delays internalization and arrests phagosome maturation in macrophages. J. Microbiol. Immunol. Infect. 2016, 49, 636–645. [Google Scholar] [CrossRef]
- Ricci, V.; Giannouli, M.; Romano, M.; Zarrilli, R. Helicobacter pylori gamma-glutamyl transpeptidase and its pathogenic role. World J. Gastroenterol. 2014, 20, 630–638. [Google Scholar] [CrossRef]
- Wüstner, S.; Anderl, F.; Wanisch, A.; Sachs, C.; Steiger, K.; Nerlich, A.; Vieth, M.; Mejías-Luque, R.; Gerhard, M. Helicobacter pylori γ-glutamyl transferase contributes to colonization and differential recruitment of T cells during persistence. Sci. Rep. 2017, 7, 13616. [Google Scholar] [CrossRef]
- Käbisch, R.; Semper, R.P.; Wüstner, S.; Gerhard, M.; Mejías-Luque, R. Helicobacter pylori γ-glutamyltranspeptidase induces tolerogenic human dendritic cells by activation of glutamate receptors. J. Immunol. 2016, 196, 4246–4252. [Google Scholar] [CrossRef]
- Montemurro, P.; Barbuti, G.; Dundon, W.G.; Del Giudice, G.; Rappuoli, R.; Colucci, M.; De Rinaldis, P.; Montecucco, C.; Semeraro, N.; Papini, N. Helicobacter pylori neutrophil-activating protein stimulates tissue factor and plasminogen activator inhibitor-2 production by human blood mononuclear cells. J. Infect. Dis. 2001, 183, 1055–1062. [Google Scholar] [CrossRef]
- Fu, H.W. Helicobacter pylori neutrophil-activating protein: From molecular pathogenesis to clinical applications. World J. Gastroenterol. 2014, 20, 5294–5301. [Google Scholar] [CrossRef]
- Polenghi, A.; Bossi, F.; Fischetti, F.; Durigutto, P.; Cabrelle, A.; Tamassia, N.; Cassatella, M.A.; Montecucco, C.; Tedesco, F.; de Bernard, M. The neutrophil-activating protein of Helicobacter pylori crosses endothelia to promote neutrophil adhesion in vivo. J. Immunol. 2007, 178, 1312–1320. [Google Scholar] [CrossRef] [PubMed]
- D’Elios, M.M.; Amedei, A.; Cappon, A.; Del Prete, G.; de Bernard, M. The neutrophil-activating protein of Helicobacter pylori (HP-NAP) as an immune modulating agent. FEMS Immunol. Med. Microbiol. 2007, 50, 157–164. [Google Scholar] [CrossRef] [PubMed]
- de Bernard, M.; D’Elios, M.M. The immune modulating activity of the Helicobacter pylori HP-NAP: Friend or foe? Toxicon 2010, 56, 1186–1192. [Google Scholar] [CrossRef] [PubMed]
- Codolo, G.; Fassan, M.; Munari, F.; Volpe, A.; Bassi, P.; Rugge, M.; Pagano, F.; D’Elios, M.M.; de Bernard, M. HP-NAP inhibits the growth of bladder cancer in mice by activating a cytotoxic Th1 response. Cancer Immunol. Immunother. 2012, 61, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Amedei, A.; Cappon, A.; Codolo, G.; Cabrelle, A.; Polenghi, A.; Benagiano, M.; Tasca, E.; Azzurri, A.; D’Elios, M.M.; Del Prete, G. The neutrophil-activating protein of Helicobacter pylori promotes Th1 immune responses. J. Clin. Investig. 2006, 116, 1092–1101. [Google Scholar] [CrossRef]
- Montecucco, C.; Rappuoli, R. Living dangerously: How Helicobacter pylori survives in the human stomach. Nat. Rev. Mol. Cell Biol. 2001, 2, 457–466. [Google Scholar] [CrossRef]
- Dundon, W.G.; Nishioka, H.; Polenghi, A.; Papinutto, E.; Zanotti, G.; Montemurro, P.; Del Giudice, G.; Rappuoli, R.; Montecucco, C. The neutrophil-activating protein of Helicobacter pylori. Int. J. Med. Microbiol. 2001, 291, 545–550. [Google Scholar] [CrossRef]
- Zhao, Y.; Yokota, K.; Ayada, K.; Yamamoto, Y.; Okada, T.; Shen, L.; Oguma, K. Helicobacter pylori heat-shock protein 60 induces interleukin-8 via a Toll-like receptor (TLR)2 and mitogen-activated protein (MAP) kinase pathway in human monocytes. J. Med. Microbiol. 2007, 56, 154–164. [Google Scholar] [CrossRef]
- Takenaka, R.; Yokota, K.; Ayada, K.; Mizuno, M.; Zhao, Y.; Fujinami, Y.; Lin, S.N.; Toyokawa, T.; Okada, H.; Shiratori, Y.; et al. Helicobacter pylori heat-shock protein 60 induces inflammatory responses through the Toll-like receptor-triggered pathway in cultured human gastric epithelial cells. Microbiology 2004, 150, 3913–3922. [Google Scholar] [CrossRef]
- Lin, C.Y.; Huang, Y.S.; Li, C.H.; Hsieh, Y.T.; Tsai, N.M.; He, P.J.; Hsu, W.T.; Yeh, Y.C.; Chiang, F.H.; Wu, M.S.; et al. Characterizing the polymeric status of Helicobacter pylori heat shock protein 60. Biochem. Biophys. Res. Commun. 2009, 388, 283–289. [Google Scholar] [CrossRef]
- Nurgalieva, Z.Z.; Conner, M.E.; Opekun, A.R.; Zheng, C.Q.; Elliott, S.N.; Ernst, P.B.; Osato, M.; Estes, M.K.; Graham, D.Y. B-cell and T-cell immune responses to experimental Helicobacter pylori infection in humans. Infect. Immun. 2005, 73, 2999–3006. [Google Scholar] [CrossRef]
- Kim, N. Immunological reactions on H. pylori infection. In Helicobacter pylori; Springer: Berlin/Heidelberg, Germany, 2016; pp. 35–52. [Google Scholar]
- O’Keeffe, J.; Moran, A.P. Conventional, regulatory, and unconventional T cells in the immunologic response to Helicobacter pylori. Helicobacter 2008, 13, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Kandulski, A.; Malfertheiner, P.; Wex, T. Role of regulatory T-cells in H. pylori-induced gastritis and gastric cancer. Anticancer Res. 2010, 30, 1093–1103. [Google Scholar] [PubMed]
- Pan, X.; Ke, H.; Niu, X.; Li, S.; Lv, J.; Pan, L. Protection against Helicobacter pylori infection in BALB/c mouse model by oral administration of multivalent epitope-based vaccine of cholera toxin B subunit-HUUC. Front. Immunol. 2018, 9, 1003. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Li, B.; Zhang, Y.; Li, R.; Ruan, H.; Wu, J.; Liu, Q. Outer membrane vesicles of Helicobacter pylori 7.13 as adjuvants promote protective efficacy against Helicobacter pylori infection. Front. Microbiol. 2020, 11, 1340. [Google Scholar] [CrossRef]
- D’Elios, M.M.; Manghetti, M.; Almerigogna, F.; Amedei, A.; Costa, F.; Burroni, D.; Baldari, C.T.; Romagnani, S.; Telford, J.L.; Del Prete, G. Different cytokine profile and antigen-specificity repertoire in Helicobacter pylori-specific T cell clones from the antrum of chronic gastritis patients with or without peptic ulcer. Eur. J. Immunol. 1997, 27, 1751–1755. [Google Scholar] [CrossRef]
- Berenson, L.S.; Ota, N.; Murphy, K.M. Issues in T-helper 1 development—Resolved and unresolved. Immunol. Rev. 2004, 202, 157–174. [Google Scholar] [CrossRef]
- Naidoo, J.; Page, D.B.; Wolchok, J.D. Immune modulation for cancer therapy. Br. J. Cancer. 2014, 111, 2214–2219. [Google Scholar] [CrossRef]
- Egan, A.J.F. Bacterial outer membrane constriction. Mol. Microbiol. 2018, 107, 676–687. [Google Scholar] [CrossRef]
- Waskito, L.A.; Salama, N.R.; Yamaoka, Y. Pathogenesis of Helicobacter pylori infection. Helicobacter 2018, 23 (Suppl. S1), 449–490. [Google Scholar] [CrossRef]
- Koebnik, R.; Locher, K.P.; Van Gelder, P. Structure and function of bacterial outer membrane proteins: Barrels in a nutshell. Mol. Microbiol. 2000, 37, 239–253. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.A.Q.; Zheng, R.; Abdalla, A.M.E.; Bakadia, B.M.; Qi, F.; Xiao, L.; Atta, O.M.; Mao, L.; Yang, G. Heterogeneous populations of outer membrane vesicles released from Helicobacter pylori SS1 with distinct biological properties. Eng. Sci. 2021, 15, 148–165. [Google Scholar] [CrossRef]
- Bauwens, A.; Kunsmann, L.; Marejková, M.; Zhang, W.; Karch, H.; Bielaszewska, M.; Mellmann, A. Intrahost milieu modulates production of outer membrane vesicles, vesicle-associated Shiga toxin 2a and cytotoxicity in Escherichia coli O157:H7 and O104:H4. Environ. Microbiol. Rep. 2017, 9, 626–634. [Google Scholar] [CrossRef] [PubMed]
- Sharif, E.; Eftekhari, Z.; Mohit, E. The effect of growth stage and isolation method on properties of ClearColi™ outer membrane vesicles (OMVs). Curr. Microbiol. 2021, 78, 1602–1614. [Google Scholar] [CrossRef]
- Winter, J.A.; Letley, D.P.; Cook, K.W.; Rhead, J.L.; Zaitoun, A.A.; Ingram, R.J.; Amilon, K.R.; Croxall, N.J.; Kaye, P.V.; Robinson, K. A role for the vacuolating cytotoxin, VacA, in colonization and Helicobacter pylori–induced metaplasia in the stomach. J. Infect. Dis. 2014, 210, 954–963. [Google Scholar] [CrossRef] [PubMed]
- Ansari, S.; Yamaoka, Y. Role of vacuolating cytotoxin A in Helicobacter pylori infection and its impact on gastric pathogenesis. Expert. Rev. Anti. Infect. Ther. 2020, 18, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Saniee, P.; Jalili, S.; Ghadersoltani, P.; Daliri, L.; Siavoshi, F. Individual hosts carry H. pylori isolates with different CagA features—motifs and copy number. Infect. Genet. Evol. 2021, 93, 104961. [Google Scholar] [CrossRef]
- Liu, Q.; Yi, J.; Liang, K.; Zhang, X.; Liu, Q. Outer membrane vesicles derived from Salmonella Enteritidis protect against the virulent wild-type strain infection in a Mouse Model. J. Microbiol. Biotechnol. 2017, 27, 1519–1528. [Google Scholar] [CrossRef]
- Mattapallil, J.J.; Dandekar, S.; Canfield, D.R.; Solnick, J.V. A predominant Th1 type of immune response is induced early during acute Helicobacter pylori infection in rhesus macaques. Gastroenterology 2000, 118, 307–315. [Google Scholar] [CrossRef]
- Shi, Y.; Liu, X.F.; Zhuang, Y.; Zhang, J.Y.; Liu, T.; Yin, Z.; Wu, C.; Mao, X.H.; Jia, K.R.; Wang, F.J.; et al. Helicobacter pylori-induced Th17 responses modulate Th1 cell responses, benefit bacterial growth, and contribute to pathology in mice. J. Immunol. 2010, 184, 5121–5129. [Google Scholar]
- Mohammadi, M.; Nedrud, J.; Redline, R.; Lycke, N.; Czinn, S.J. Murine CD4 T-cell response to Helicobacter infection: TH1 cells enhance gastritis and TH2 cells reduce bacterial load. Gastroenterology 1997, 113, 1848–1857. [Google Scholar] [CrossRef]
- Ki, M.R.; Hong, I.H.; Park, J.K.; Hong, K.S.; Hwang, O.K.; Han, J.Y.; Ji, A.R.; Park, S.I.; Lee, S.K.; Yoo, S.E.; et al. Potent neutralization of vacuolating cytotoxin (VacA) of Helicobacter pylori by immunoglobulins against the soluble recombinant VacA. Anticancer Res. 2009, 29, 2393–2402. [Google Scholar] [PubMed]
- Fahimi, F.; Tohidkia, M.R.; Fouladi, M.; Aghabeygi, R.; Samadi, N.; Omidi, Y. Pleiotropic cytotoxicity of VacA toxin in host cells and its impact on immunotherapy. Bioimpacts 2017, 7, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Moyat, M.; Velin, D. Use of VacA as a Vaccine Antigen. Toxins 2016, 8, 181. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.P.; Pedersen, J.; Zhan, Y.; Lew, A.M.; Pearse, M.J.; Wijburg, O.L.; Strugnell, R.A. CD8+ T cells are associated with severe gastritis in Helicobacter pylori-infected mice in the absence of CD4+ T cells. Infect. Immun. 2008, 76, 1289–1297. [Google Scholar] [CrossRef] [PubMed]
- Hatakeyama, M. Structure and function of Helicobacter pylori CagA, the first-identified bacterial protein involved in human cancer. Proc. Jpn. Acad., Ser. B Phys. Biol. Sci. 2017, 93, 196–219. [Google Scholar] [CrossRef]
- Cheng, H.H.; Tseng, G.Y.; Yang, H.B.; Wang, H.J.; Lin, H.J.; Wang, W.C. Increased numbers of Foxp3-positive regulatory T cells in gastritis, peptic ulcer and gastric adenocarcinoma. World J. Gastroenterol. 2012, 18, 34–43. [Google Scholar] [CrossRef]
- Kaebisch, R.; Mejias-Luque, R.; Prinz, C.; Gerhard, M. Helicobacter pylori cytotoxin-associated gene A impairs human dendritic cell maturation and function through IL-10-mediated activation of STAT3. J. Immunol. 2014, 192, 316–323. [Google Scholar] [CrossRef]
- Denic, M.; Touati, E.; De Reuse, H. Review: Pathogenesis of Helicobacter pylori infection. Helicobacter 2020, 25 (Suppl. S1), e12638. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.K.; Zhu, H.F.; He, B.S.; Zhang, Z.Y.; Chen, Z.T.; Wang, Z.Z.; Wu, G.L. CagA+ H. pylori infection is associated with polarization of T helper cell immune responses in gastric carcinogenesis. World J. Gastroenterol. 2007, 13, 2923–2931. [Google Scholar] [CrossRef]
- Talebi Bezmin Abadi, A.; Perez-Perez, G. Role of dupA in virulence of Helicobacter pylori. World J. Gastroenterol. 2016, 22, 10118–10123. [Google Scholar] [CrossRef] [PubMed]
- Argent, R.H.; Burette, A.; Miendje Deyi, V.Y.; Atherton, J.C. The presence of dupA in Helicobacter pylori is not significantly associated with duodenal ulceration in Belgium, South Africa, China, or North America. Clin. Infect. Dis. 2007, 45, 1204–1206. [Google Scholar] [CrossRef] [PubMed]
- Paredes-Osses, E.; Sáez, K.; Sanhueza, E.; Hebel, S.; González, C.; Briceño, C.; García Cancino, A. Association between cagA, vacAi, and dupA genes of Helicobacter pylori and gastroduodenal pathologies in Chilean patients. Folia Microbiol. 2017, 62, 437–444. [Google Scholar] [CrossRef]
- Akbari, S.; Rezaeian, T.; Mohammadzadeh, R.; Meshkat, Z.; Namdar, A.B.; Aryan, E.; Youssefi, M.; Pishdadian, A.; Ahmadi, A.; Farsiani, H. Investigation of association between iceA, babA2, and oipA genotypes of Helicobacter pylori and IL-8-251 T>A polymorphism with clinical outcomes in Helicobacter pylori-infected Iranian patients. Gene Rep. 2021, 24, 101210. [Google Scholar] [CrossRef]
- Cañas, M.-A.; Giménez, R.; Fábrega, M.-J.; Toloza, L.; Baldomà, L.; Badia, J. Outer membrane vesicles from the probiotic Escherichia coli Nissle 1917 and the commensal ECOR12 enter intestinal epithelial cells via clathrin-dependent endocytosis and elicit differential effects on DNA damage. PLoS ONE 2016, 11, e0160374. [Google Scholar] [CrossRef]
- Sanders, M.E.; Merenstein, D.J.; Reid, G.; Gibson, G.R.; Rastall, R.A. Probiotics and prebiotics in intestinal health and disease: From biology to the clinic. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 605–616. [Google Scholar] [CrossRef]
- Hu, R.; Lin, H.; Li, J.; Zhao, Y.; Wang, M.; Sun, X.; Min, Y.; Gao, Y.; Yang, M. Probiotic Escherichia coli Nissle 1917-derived outer membrane vesicles enhance immunomodulation and antimicrobial activity in RAW264.7 macrophages. BMC Microbiol. 2020, 20, 268. [Google Scholar] [CrossRef]
- Cañas, M.-A.; Fábrega, M.-J.; Giménez, R.; Badia, J.; Baldomà, L. Outer membrane vesicles from probiotic and commensal Escherichia coli activate NOD1-mediated immune responses in intestinal epithelial cells. Front. Microbiol. 2018, 9, 498. [Google Scholar] [CrossRef]
- Tobias, J.; Lebens, M.; Wai, S.N.; Holmgren, J.; Svennerholm, A.M. Surface expression of Helicobacter pylori HpaA adhesion antigen on Vibrio cholerae, enhanced by co-expressed enterotoxigenic Escherichia coli fimbrial antigens. Microb. Pathog. 2017, 105, 177–184. [Google Scholar] [CrossRef]




| Method | Limitations | Refs. |
|---|---|---|
| Antibiotic treatment |
| [135,136,137,138] |
| Triple therapy: Treatment with proton-pump inhibitor (PPI), amoxicillin, and a third drug (e.g., levofloxacin or clarithromycin). |
| [138,139] |
| Quadruple therapy: Treatment with PPI, bismuth, tetracycline, and metronidazole. |
| [138,140] |
| Sequential therapy: Two treatment regimens are applied; the first one (consisting of PPI and amoxicillin) will be used in the first half of the treatment duration, and another regimen (consisting of PPI, clarithromycin, and one of the nitroimidazole family antibiotics) will be used for the second half of treatment duration. |
| [138,141] |
| Probiotics therapy: The use of bacteria that produce lactic acid such as Lactobacillus spp., etc., to eradicate H. pylori infection. |
| [142,143,144] |
| Strategies | Refs. |
|---|---|
| Using OMVs isolated from standard or manipulated growth conditions or from a specific growth stage | [17,18,233,234,235] |
| Using OMVs from bacterial strains that contain nontoxigenic virulence factor genotypes (e.g., CagA, VacA, DupA) or that lack certain virulence factors (e.g., CagA-negative H. pylori strains, DupA-negative H. pylori strains) | [155,157,194,196,236,237,238,239] |
| Using OMVs from probiotic or commensal bacteria as antigen carriers for the antigens of interest | [15] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed, A.A.Q.; Besio, R.; Xiao, L.; Forlino, A. Outer Membrane Vesicles (OMVs) as Biomedical Tools and Their Relevance as Immune-Modulating Agents against H. pylori Infections: Current Status and Future Prospects. Int. J. Mol. Sci. 2023, 24, 8542. https://doi.org/10.3390/ijms24108542
Ahmed AAQ, Besio R, Xiao L, Forlino A. Outer Membrane Vesicles (OMVs) as Biomedical Tools and Their Relevance as Immune-Modulating Agents against H. pylori Infections: Current Status and Future Prospects. International Journal of Molecular Sciences. 2023; 24(10):8542. https://doi.org/10.3390/ijms24108542
Chicago/Turabian StyleAhmed, Abeer Ahmed Qaed, Roberta Besio, Lin Xiao, and Antonella Forlino. 2023. "Outer Membrane Vesicles (OMVs) as Biomedical Tools and Their Relevance as Immune-Modulating Agents against H. pylori Infections: Current Status and Future Prospects" International Journal of Molecular Sciences 24, no. 10: 8542. https://doi.org/10.3390/ijms24108542