
Proportion of Concentrate in the Diet of Early Lactation Dairy Cows Has Contrasting Effects on Circulating Leukocyte Global Transcriptomic Profiles, Health and Fertility According to Parity
 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Results
2.1. Effect of Diets on Dry Matter Intakes, Milk Yield, Energy Balance and Blood Metabolites
2.2. Effect of Diets on Inflammatory Parameters
2.3. Fertility Data
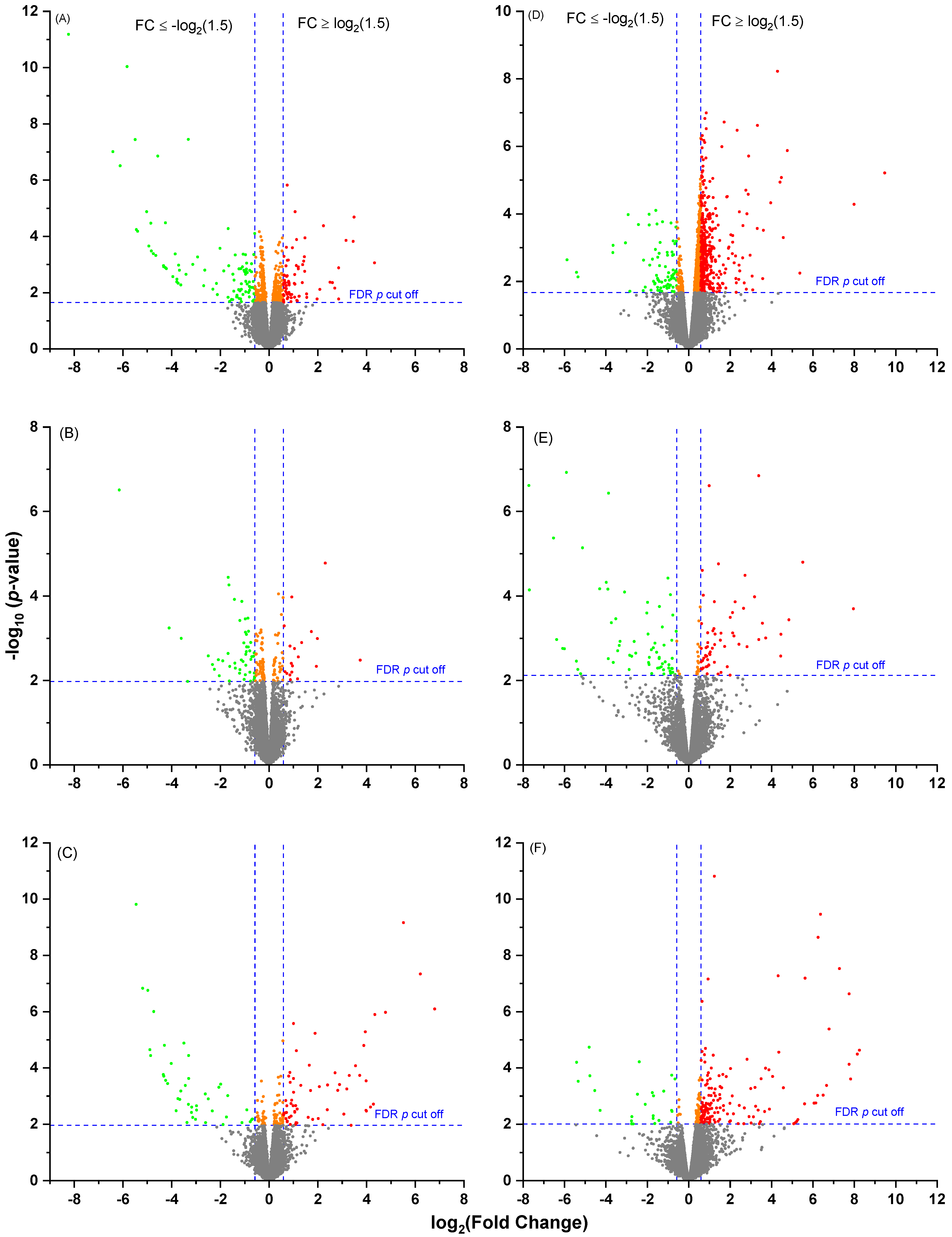
2.4. Leukocyte Transcriptomic Profiles in Cows Offered Different Proportions of Concentrate
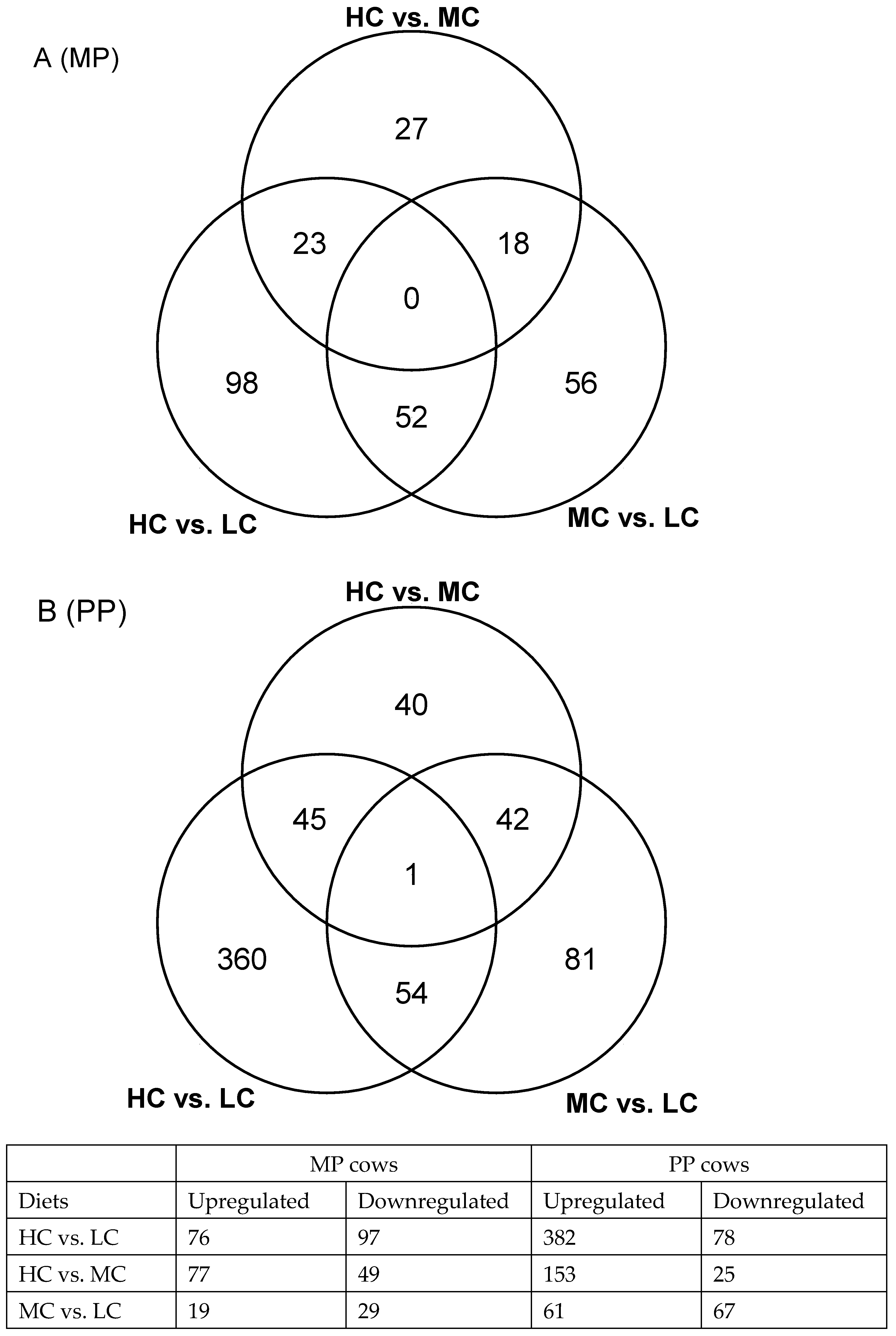
2.5. Comparison of Leukocyte Gene Expression Profiles between Multiparous Cows Receiving High or Low Concentrate Diets
2.6. Comparison of Leukocyte Gene Expression Patterns between Primiparous Cows Receiving High or Low Concentrate Diets
3. Discussion
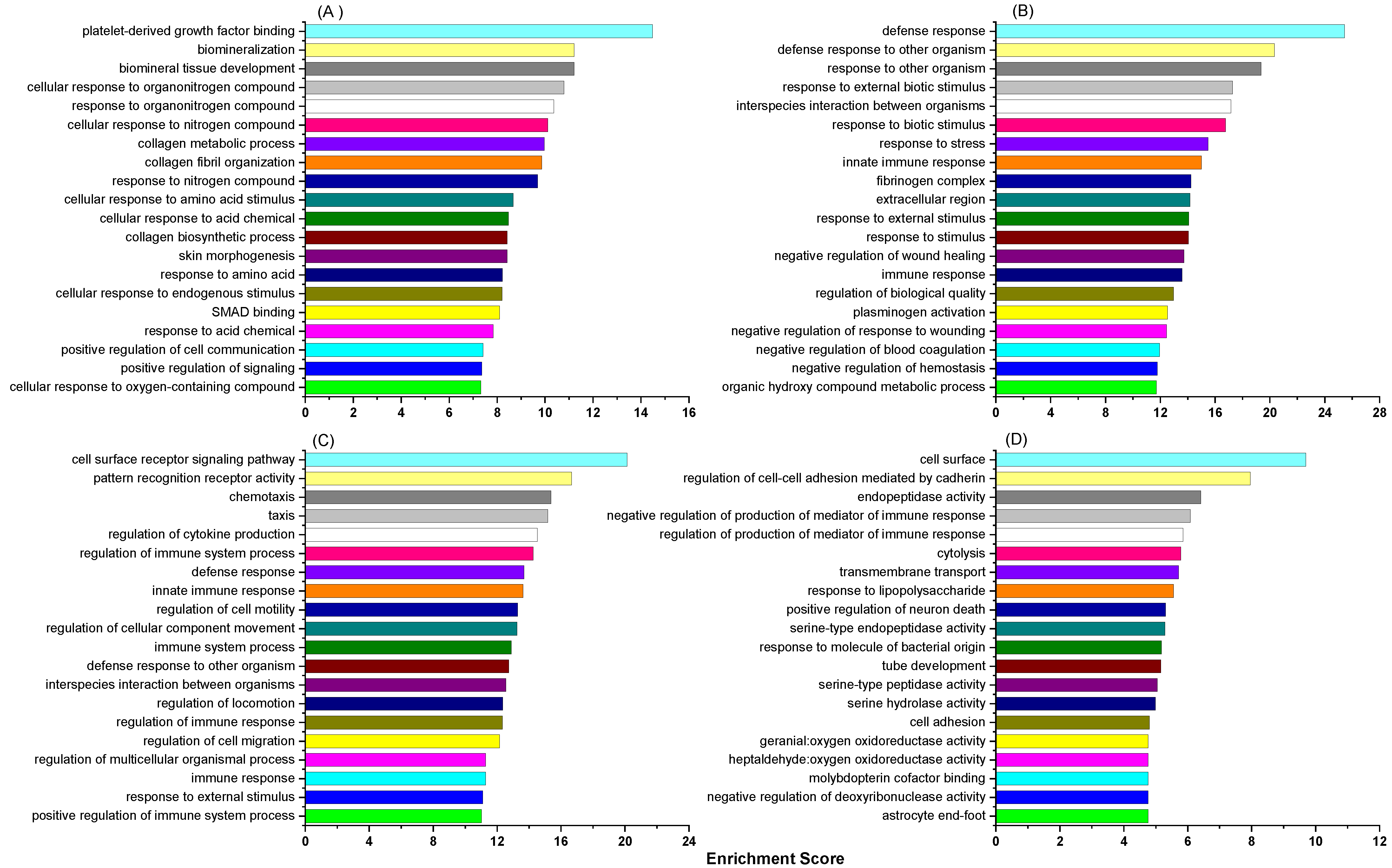
3.1. Comparison of the Effects of the High and Low Concentrate Diets in the Multiparous Cows
3.2. Comparison of the Effects of the High and Low Concentrate Diets in the Primiparous Cows
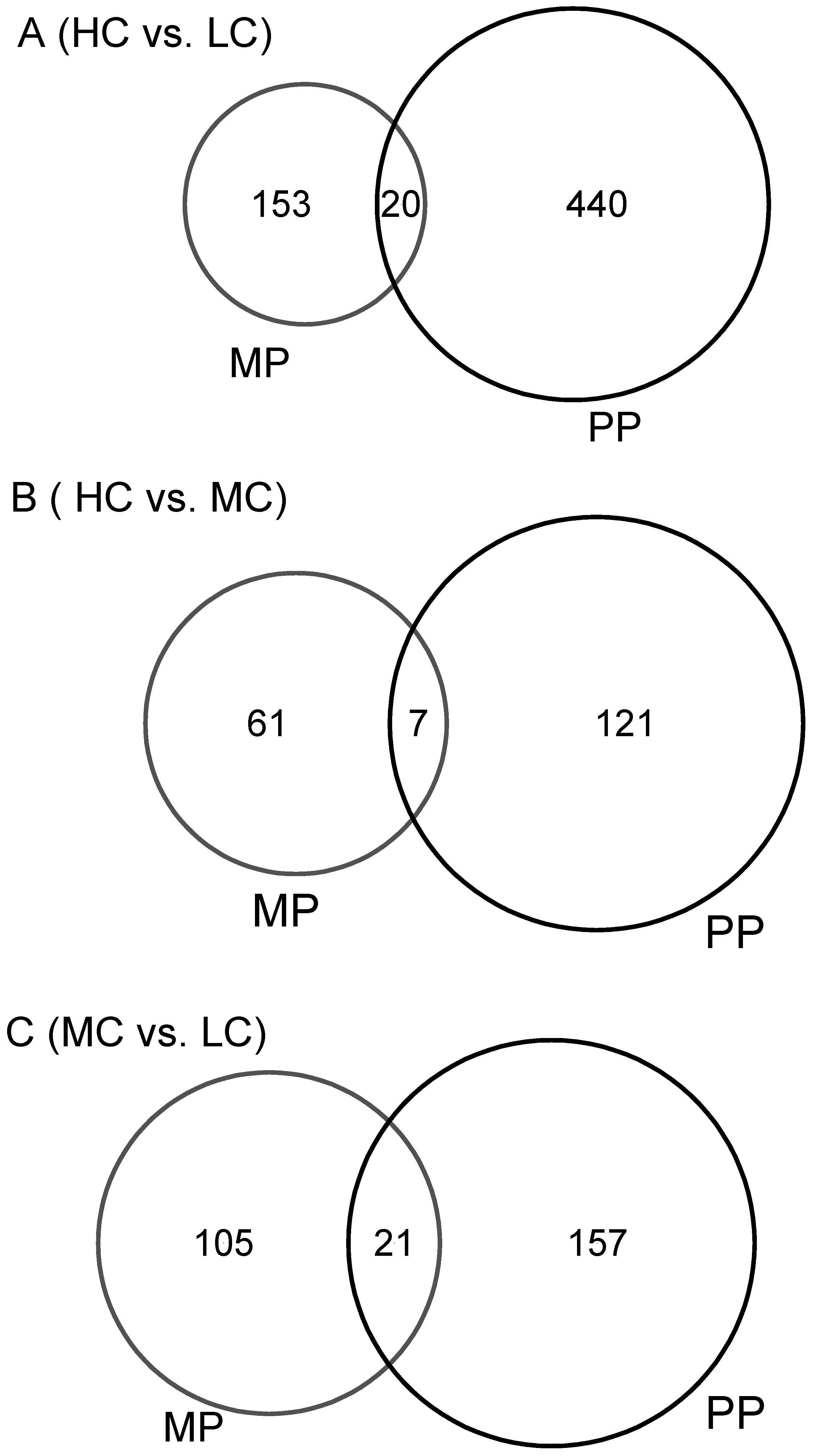
3.3. Similarities in Response to Diet by Multiparous and Primiparous Cows
3.4. Implications for Fertility
3.5. Limitations of the Study
4. Materials and Methods
4.1. Animals and Diets
4.2. Cow Phenotype Data Collection
4.3. Uterine Cytology Analysis
4.4. Analysis of Circulating Metabolites and IGF-1
4.5. Blood RNA Extraction
4.6. RNA-Sequencing, Mapping and Quantification
4.7. Analysis of Differential Gene Expression between the Dietary Groups
4.8. Gene Ontology (GO) Enrichment Analysis
4.9. Statistical Analysis of Phenotype Data
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mallard, B.A.; Dekkers, J.C.; Ireland, M.J.; Leslie, K.E.; Sharif, S.; Vankampen, C.L.; Wagter, L.; Wilkie, B.N. Alteration in immune responsiveness during the peripartum period and its ramification on dairy cow and calf health. J. Dairy Sci. 1998, 81, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Wathes, D.C.; Cheng, Z.; Chowdhury, W.; Fenwick, M.A.; Fitzpatrick, R.; Morris, D.G.; Patton, J.; Murphy, J.J. Negative energy balance alters global gene expression and immune responses in the uterus of postpartum dairy cows. Physiol. Genom. 2009, 39, 1–13. [Google Scholar] [CrossRef]
- Ingvartsen, K.L.; Moyes, K. Nutrition, immune function and health of dairy cattle. Animal 2013, 7 (Suppl. 1), 112–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kehrli, M.E., Jr.; Nonnecke, B.J.; Roth, J.A. Alterations in bovine neutrophil function during the periparturient period. Am. J. Vet. Res. 1989, 50, 207–214. [Google Scholar] [PubMed]
- Ster, C.; Loiselle, M.C.; Lacasse, P. Effect of postcalving serum nonesterified fatty acids concentration on the functionality of bovine immune cells. J. Dairy Sci. 2012, 95, 708–717. [Google Scholar] [CrossRef] [PubMed]
- Lacetera, N.; Scalia, D.; Bernabucci, U.; Ronchi, B.; Pirazzi, D.; Nardone, A. Lymphocyte functions in overconditioned cows around parturition. J. Dairy Sci. 2005, 88, 2010–2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nonnecke, B.J.; Kimura, K.; Goff, J.P.; Kehrli, M.E., Jr. Effects of the mammary gland on functional capacities of blood mononuclear leukocyte populations from periparturient cows. J. Dairy Sci. 2003, 86, 2359–2368. [Google Scholar] [CrossRef] [Green Version]
- Habel, J.; Sundrum, A. Mismatch of glucose allocation between different life functions in the transition period of dairy cows. Animals 2020, 10, 1028. [Google Scholar] [CrossRef]
- Horst, E.A.; Kvidera, S.K.; Baumgard, L.H. Invited review: The influence of immune activation on transition cow health and performance-A critical evaluation of traditional dogmas. J. Dairy Sci. 2021, 104, 8380–8410. [Google Scholar] [CrossRef]
- Wathes, D.C.; Becker, F.; Buggiotti, L.; Crowe, M.A.; Ferris, C.; Foldager, L.; Grelet, C.; Hostens, M.; Ingvartsen, K.L.; Marchitelli, C.; et al. Associations between circulating IGF-1 concentrations, disease status and the leukocyte transcriptome in early lactation dairy cows. Ruminants 2021, 1, 147–177. [Google Scholar] [CrossRef]
- Dimeloe, S.; Burgener, A.V.; Grahlert, J.; Hess, C. T-cell metabolism governing activation, proliferation and differentiation; a modular view. Immunology 2017, 150, 35–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loftus, R.M.; Finlay, D.K. Immunometabolism: Cellular metabolism turns immune regulator. J. Biol. Chem. 2016, 291, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauman, D.E.; Currie, W.B. Partitioning of nutrients during pregnancy and lactation: A review of mechanisms involving homeostasis and homeorhesis. J. Dairy Sci. 1980, 63, 1514–1529. [Google Scholar] [CrossRef] [PubMed]
- Drackley, J.K. ADSA Foundation Scholar Award. Biology of dairy cows during the transition period: The final frontier? J. Dairy Sci. 1999, 82, 2259–2273. [Google Scholar] [CrossRef] [PubMed]
- Jabbour, H.N.; Sales, K.J.; Catalano, R.D.; Norman, J.E. Inflammatory pathways in female reproductive health and disease. Reproduction 2009, 138, 903–919. [Google Scholar] [CrossRef]
- Pascottini, O.B.; LeBlanc, S.J. Modulation of immune function in the bovine uterus peripartum. Theriogenology 2020, 150, 193–200. [Google Scholar] [CrossRef]
- Sheldon, I.M.; Lewis, G.S.; LeBlanc, S.; Gilbert, R.O. Defining postpartum uterine disease in cattle. Theriogenology 2006, 65, 1516–1530. [Google Scholar] [CrossRef]
- Kuhla, B. Review: Pro-inflammatory cytokines and hypothalamic inflammation: Implications for insufficient feed intake of transition dairy cows. Animal 2020, 14, s65–s77. [Google Scholar] [CrossRef] [Green Version]
- Ingvartsen, K.L.; Moyes, K.M. Factors contributing to immunosuppression in the dairy cow during the periparturient period. Jpn. J. Vet. Res. 2015, 63 (Suppl. 1), S15–S24. [Google Scholar]
- Wathes, D.C.; Cheng, Z.; Salavati, M.; Buggiotti, L.; Takeda, H.; Tang, L.; Becker, F.; Ingvartsen, K.I.; Ferris, C.; Hostens, M.; et al. Relationships between metabolic profiles and gene expression in liver and leukocytes of dairy cows in early lactation. J. Dairy Sci. 2021, 104, 3596–3616. [Google Scholar] [CrossRef]
- Pinedo, P.; Melendez, P. Liver disorders associated with metabolic imbalances in dairy cows. Vet. Clin. N. Am. Food Anim. Pract. 2022, 38, 433–446. [Google Scholar] [CrossRef] [PubMed]
- Fenwick, M.A.; Fitzpatrick, R.; Kenny, D.A.; Diskin, M.G.; Patton, J.; Murphy, J.J.; Wathes, D.C. Interrelationships between negative energy balance (NEB) and IGF regulation in liver of lactating dairy cows. Domest. Anim. Endocrinol. 2008, 34, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, Y.; Boyd, C.K.; Bracken, C.J.; Lamberson, W.R.; Keisler, D.H.; Lucy, M.C. Reduced growth hormone receptor (GHR) messenger ribonucleic acid in liver of periparturient cattle is caused by a specific down-regulation of GHR 1A that is associated with decreased insulin-like growth factor I. Endocrinology 1999, 140, 3947–3954. [Google Scholar] [CrossRef]
- Kvidera, S.K.; Horst, E.A.; Abuajamieh, M.; Mayorga, E.J.; Fernandez, M.V.; Baumgard, L.H. Glucose requirements of an activated immune system in lactating Holstein cows. J. Dairy Sci. 2017, 100, 2360–2374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Z.; Wylie, A.; Ferris, C.; Ingvartsen, K.L.; Wathes, D.C.; Gplus, E.C. Effect of diet and nonesterified fatty acid levels on global transcriptomic profiles in circulating peripheral blood mononuclear cells in early lactation dairy cows. J. Dairy Sci. 2021, 104, 10059–10075. [Google Scholar] [CrossRef]
- Wathes, D.C.; Fenwick, M.; Cheng, Z.; Bourne, N.; Llewellyn, S.; Morris, D.G.; Kenny, D.; Murphy, J.; Fitzpatrick, R. Influence of negative energy balance on cyclicity and fertility in the high producing dairy cow. Theriogenology 2007, 68 (Suppl. 1), S232–S241. [Google Scholar] [CrossRef]
- Santos, J.E.; Bisinotto, R.S.; Ribeiro, E.S. Mechanisms underlying reduced fertility in anovular dairy cows. Theriogenology 2016, 86, 254–262. [Google Scholar] [CrossRef] [Green Version]
- Pascottini, O.B.; Leroy, J.; Opsomer, G. Maladaptation to the transition period and consequences on fertility of dairy cows. Reprod. Dom. Anim. 2022, 57 (Suppl. 4), 21–32. [Google Scholar] [CrossRef]
- Leroy, J.L.; Valckx, S.D.; Jordaens, L.; De Bie, J.; Desmet, K.L.; Van Hoeck, V.; Britt, J.H.; Marei, W.F.; Bols, P.E. Nutrition and maternal metabolic health in relation to oocyte and embryo quality: Critical views on what we learned from the dairy cow model. Reprod. Fertil. Dev. 2015, 27, 693–703. [Google Scholar] [CrossRef]
- Coffey, M.P.; Hickey, J.; Brotherstone, S. Genetic aspects of growth of Holstein-Friesian dairy cows from birth to maturity. J. Dairy Sci. 2006, 89, 322–329. [Google Scholar] [CrossRef] [Green Version]
- Wathes, D.C.; Cheng, Z.; Bourne, N.; Taylor, V.J.; Coffey, M.P.; Brotherstone, S. Differences between primiparous and multiparous dairy cows in the inter-relationships between metabolic traits, milk yield and body condition score in the periparturient period. Domest. Anim. Endocrinol. 2007, 33, 203–225. [Google Scholar] [CrossRef] [PubMed]
- Little, M.W.; Wylie, A.R.G.; O’Connell, N.E.; Welsh, M.D.; Grelet, C.; Bell, M.J.; Gordon, A.; Ferris, C.P. Immunological effects of altering the concentrate inclusion level in a grass silage-based diet for early lactation Holstein Friesian cows. Animal 2019, 13, 799–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buggiotti, L.; Cheng, Z.; Salavati, M.; Wathes, C.D.; Genotype plus Environment, C. Comparison of the transcriptome in circulating leukocytes in early lactation between primiparous and multiparous cows provides evidence for age-related changes. BMC Genom. 2021, 22, 693. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, F.C.; Kalscheur, K.F.; Drackley, J.K. Symposium review: Nutrition strategies for improved health, production, and fertility during the transition period. J. Dairy Sci. 2020, 103, 5684–5693. [Google Scholar] [CrossRef] [PubMed]
- Ingvartsen, K.L. Feeding- and management-related diseases in the transition cow. Anim. Feed Sci. Technol. 2006, 126, 175–213. [Google Scholar] [CrossRef]
- Ferris, C.P.; Gordon, F.J.; Patterson, D.C.; Mayne, C.S.; McCoy, M.A. A shortterm comparison of the performance of four grassland-based systems of milk production for autumn-calving dairy cows. Grass Forage Sci. 2003, 58, 8. [Google Scholar] [CrossRef]
- Taylor, V.J.; Cheng, Z.; Pushpakumara, P.G.; Beever, D.E.; Wathes, D.C. Relationships between the plasma concentrations of insulin-like growth factor-I in dairy cows and their fertility and milk yield. Vet. Rec. 2004, 155, 583–588. [Google Scholar] [CrossRef]
- LeBlanc, S.J. Interactions of metabolism, inflammation, and reproductive tract health in the postpartum period in dairy cattle. Reprod. Domc. Anim. 2012, 47 (Suppl. 5), 18–30. [Google Scholar] [CrossRef]
- Vailati-Riboni, M.; Kanwal, M.; Bulgari, O.; Meier, S.; Priest, N.V.; Burke, C.R.; Kay, J.K.; McDougall, S.; Mitchell, M.D.; Walker, C.G.; et al. Body condition score and plane of nutrition prepartum affect adipose tissue transcriptome regulators of metabolism and inflammation in grazing dairy cows during the transition period. J. Dairy Sci. 2016, 99, 758–770. [Google Scholar] [CrossRef] [Green Version]
- Trevisi, E.; Minuti, A. Assessment of the innate immune response in the periparturient cow. Res. Vet. Sci. 2018, 116, 47–54. [Google Scholar] [CrossRef]
- Druker, S.A.; Sicsic, R.; van Straten, M.; Goshen, T.; Kedmi, M.; Raz, T. Cytological endometritis diagnosis in primiparous versus multiparous dairy cows. J. Dairy Sci. 2022, 105, 665–683. [Google Scholar] [CrossRef] [PubMed]
- Hamzeh-Cognasse, H.; Laradi, S.; Osselaer, J.C.; Cognasse, F.; Garraud, O. Amotosalen-HCl-UVA pathogen reduction does not alter poststorage metabolism of soluble CD40 ligand, Ox40 ligand and interkeukin-27, the cytokines that generally associate with serious adverse events. Vox Sang. 2015, 108, 205–207. [Google Scholar] [CrossRef] [PubMed]
- de Greeff, A.; Zadoks, R.; Ruuls, L.; Toussaint, M.; Nguyen, T.K.; Downing, A.; Rebel, J.; Stockhofe-Zurwieden, N.; Smith, H. Early host response in the mammary gland after experimental Streptococcus uberis challenge in heifers. J. Dairy Sci. 2013, 96, 3723–3736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Z.; Chauhan, L.; Barry, A.T.; Abudureyimu, A.; Oguejiofor, C.F.; Chen, X.; Wathes, D.C. Acute bovine viral diarrhea virus infection inhibits expression of interferon tau-stimulated genes in bovine endometrium. Biol. Reprod. 2017, 96, 1142–1153. [Google Scholar] [CrossRef]
- Schoggins, J.W. Interferon-Stimulated Genes: What Do They All Do? Annu. Rev. Virol. 2019, 6, 567–584. [Google Scholar] [CrossRef] [PubMed]
- Ohto, U. Activation and regulation mechanisms of NOD-like receptors based on structural biology. Front. Immunol. 2022, 13, 953530. [Google Scholar] [CrossRef]
- Cheng, Z.; Palma-Vera, S.; Buggiotti, L.; Salavati, M.; Becker, F.; Werling, D.; Wathes, D.C.; Gplus, E.C. Transcriptomic analysis of circulating leukocytes obtained during the recovery from clinical mastitis caused by Escherichia coli in Holstein dairy cows. Animals 2022, 12, 2146. [Google Scholar] [CrossRef]
- Loria, V.; Dato, I.; Graziani, F.; Biasucci, L.M. Myeloperoxidase: A new biomarker of inflammation in ischemic heart disease and acute coronary syndromes. Mediat. Inflamm. 2008, 2008, 135625. [Google Scholar] [CrossRef] [Green Version]
- Olivera, A.; Beaven, M.A.; Metcalfe, D.D. Mast cells signal their importance in health and disease. J. Allergy Clin. Immunol. 2018, 142, 381–393. [Google Scholar] [CrossRef] [Green Version]
- Dickinson, S.E.; Griffin, B.A.; Elmore, M.F.; Kriese-Anderson, L.; Elmore, J.B.; Dyce, P.W.; Rodning, S.P.; Biase, F.H. Transcriptome profiles in peripheral white blood cells at the time of artificial insemination discriminate beef heifers with different fertility potential. BMC Genom. 2018, 19, 129. [Google Scholar] [CrossRef] [Green Version]
- Van Meulder, F.; Van Coppernolle, S.; Borloo, J.; Rinaldi, M.; Li, R.W.; Chiers, K.; Van den Broeck, W.; Vercruysse, J.; Claerebout, E.; Geldhof, P. Granule exocytosis of granulysin and granzyme B as a potential key mechanism in vaccine-induced immunity in cattle against the nematode Ostertagia ostertagi. Infect. Immun. 2013, 81, 1798–1809. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.L.; Brown, B.L. Structural basis for dysregulation of aminolevulinic acid synthase in human disease. J. Biol. Chem. 2022, 298, 101643. [Google Scholar] [CrossRef] [PubMed]
- Trapani, J.A.; Sutton, V.R. Granzyme B: Pro-apoptotic, antiviral and antitumor functions. Curr. Opin. Immunol. 2003, 15, 533–543. [Google Scholar] [CrossRef] [PubMed]
- Campanile, G.; Baruselli, P.S.; Limone, A.; D’Occhio, M.J. Local action of cytokines and immune cells in communication between the conceptus and uterus during the critical period of early embryo development, attachment and implantation—Implications for embryo survival in cattle: A review. Theriogenology 2021, 167, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Bougarne, N.; Weyers, B.; Desmet, S.J.; Deckers, J.; Ray, D.W.; Staels, B.; De Bosscher, K. Molecular Actions of PPARalpha in lipid metabolism and inflammation. Endocr. Rev. 2018, 39, 760–802. [Google Scholar] [CrossRef] [Green Version]
- Sobolev, V.V.; Tchepourina, E.; Korsunskaya, I.M.; Geppe, N.A.; Chebysheva, S.N.; Soboleva, A.G.; Mezentsev, A. The Role of transcription factor PPAR-gamma in the pathogenesis of psoriasis, skin Cells, and immune cells. Int. J. Mol. Sci. 2022, 23, 9708. [Google Scholar] [CrossRef]
- Song, S.; Attia, R.R.; Connaughton, S.; Niesen, M.I.; Ness, G.C.; Elam, M.B.; Hori, R.T.; Cook, G.A.; Park, E.A. Peroxisome proliferator activated receptor alpha (PPARalpha) and PPAR gamma coactivator (PGC-1alpha) induce carnitine palmitoyltransferase IA (CPT-1A) via independent gene elements. Mol. Cell. Endocrinol. 2010, 325, 54–63. [Google Scholar] [CrossRef] [Green Version]
- Ziouzenkova, O.; Plutzky, J. Retinoid metabolism and nuclear receptor responses: New insights into coordinated regulation of the PPAR-RXR complex. FEBS Lett. 2008, 582, 32–38. [Google Scholar] [CrossRef] [Green Version]
- Espenshade, P.J.; Hughes, A.L. Regulation of sterol synthesis in eukaryotes. Annu. Rev. Genet. 2007, 41, 401–427. [Google Scholar] [CrossRef]
- Van Meer, G.; Voelker, D.R.; Feigenson, G.W. Membrane lipids: Where they are and how they behave. Nat. Rev. Mol. Cell. Biol. 2008, 9, 112–124. [Google Scholar] [CrossRef]
- Aguilar-Ballester, M.; Herrero-Cervera, A.; Vinue, A.; Martinez-Hervas, S.; Gonzalez-Navarro, H. Impact of cholesterol metabolism in immune cell function and atherosclerosis. Nutrients 2020, 12, 2021. [Google Scholar] [CrossRef] [PubMed]
- Mamedova, L.K.; Yuan, K.; Laudick, A.N.; Fleming, S.D.; Mashek, D.G.; Bradford, B.J. Toll-like receptor 4 signaling is required for induction of gluconeogenic gene expression by palmitate in human hepatic carcinoma cells. J. Nutr. Biochem. 2013, 24, 1499–1507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soto-Heredero, G.; Gomez de Las Heras, M.M.; Gabande-Rodriguez, E.; Oller, J.; Mittelbrunn, M. Glycolysis—A key player in the inflammatory response. FEBS J. 2020, 287, 3350–3369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kominsky, D.J.; Campbell, E.L.; Colgan, S.P. Metabolic shifts in immunity and inflammation. J. Immunol. 2010, 184, 4062–4068. [Google Scholar] [CrossRef] [Green Version]
- Afacan, N.J.; Yeung, A.T.; Pena, O.M.; Hancock, R.E. Therapeutic potential of host defense peptides in antibiotic-resistant infections. Curr. Pharm. Des. 2012, 18, 807–819. [Google Scholar] [CrossRef] [Green Version]
- Auvynet, C.; Rosenstein, Y. Multifunctional host defense peptides: Antimicrobial peptides, the small yet big players in innate and adaptive immunity. FEBS J. 2009, 276, 6497–6508. [Google Scholar] [CrossRef]
- Hilchie, A.L.; Wuerth, K.; Hancock, R.E. Immune modulation by multifaceted cationic host defense (antimicrobial) peptides. Nat. Chem. Biol. 2013, 9, 761–768. [Google Scholar] [CrossRef]
- Cheng, Z.; Buggiotti, L.; Salavati, M.; Marchitelli, C.; Palma-Vera, S.; Wylie, A.; Takeda, H.; Tang, L.; Crowe, M.A.; Wathes, D.C.; et al. Global transcriptomic profiles of circulating leucocytes in early lactation cows with clinical or subclinical mastitis. Mol. Biol. Rep. 2021, 48, 4611–4623. [Google Scholar] [CrossRef]
- Foley, C.; Chapwanya, A.; Callanan, J.J.; Whiston, R.; Miranda-CasoLuengo, R.; Lu, J.; Meijer, W.G.; Lynn, D.J.; O’Farrelly, C.; Meade, K.G. Integrated analysis of the local and systemic changes preceding the development of post-partum cytological endometritis. BMC Genom. 2015, 16, 811. [Google Scholar] [CrossRef] [Green Version]
- Machado, V.S.; Silva, T.H. Adaptive immunity in the postpartum uterus: Potential use of vaccines to control metritis. Theriogenology 2020, 150, 201–209. [Google Scholar] [CrossRef]
- Ley, K.; Hoffman, H.M.; Kubes, P.; Cassatella, M.A.; Zychlinsky, A.; Hedrick, C.C.; Catz, S.D. Neutrophils: New insights and open questions. Sci. Immunol. 2018, 3, eaat4579. [Google Scholar] [CrossRef] [PubMed]
- Wahl, S.M.; Feldman, G.M.; McCarthy, J.B. Regulation of leukocyte adhesion and signaling in inflammation and disease. J. Leukoc. Biol. 1996, 59, 789–796. [Google Scholar] [CrossRef] [PubMed]
- Schubert, K.; Polte, T.; Bonisch, U.; Schader, S.; Holtappels, R.; Hildebrandt, G.; Lehmann, J.; Simon, J.C.; Anderegg, U.; Saalbach, A. Thy-1 (CD90) regulates the extravasation of leukocytes during inflammation. Eur. J. Immunol. 2011, 41, 645–656. [Google Scholar] [CrossRef] [PubMed]
- Dreymueller, D.; Pruessmeyer, J.; Schumacher, J.; Fellendorf, S.; Hess, F.M.; Seifert, A.; Babendreyer, A.; Bartsch, J.W.; Ludwig, A. The metalloproteinase ADAM8 promotes leukocyte recruitment in vitro and in acute lung inflammation. Am. J. Physiol. Lung Cell. Mol. Physiol. 2017, 313, L602–L614. [Google Scholar] [CrossRef] [Green Version]
- Kostelnik, K.B.; Barker, A.; Schultz, C.; Mitchell, T.P.; Rajeeve, V.; White, I.J.; Aurrand-Lions, M.; Nourshargh, S.; Cutillas, P.; Nightingale, T.D. Dynamic trafficking and turnover of JAM-C is essential for endothelial cell migration. PLoS Biol. 2019, 17, e3000554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stillie, R.; Farooq, S.M.; Gordon, J.R.; Stadnyk, A.W. The functional significance behind expressing two IL-8 receptor types on PMN. J. Leukoc. Biol. 2009, 86, 529–543. [Google Scholar] [CrossRef] [Green Version]
- Bouton, A.H.; Riggins, R.B.; Bruce-Staskal, P.J. Functions of the adapter protein Cas: Signal convergence and the determination of cellular responses. Oncogene 2001, 20, 6448–6458. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Cheng, Z.; Zhang, S.; Werling, D.; Wathes, D.C. Combining genome wide association studies and differential gene expression data analyses identifies candidate genes affecting mastitis caused by two different pathogens in the dairy cow. Open J. Anim. Sci. 2015, 5, 358–393. [Google Scholar] [CrossRef] [Green Version]
- Chang, G.W.; Hsiao, C.C.; Peng, Y.M.; Vieira Braga, F.A.; Kragten, N.A.; Remmerswaal, E.B.; van de Garde, M.D.; Straussberg, R.; Konig, G.M.; Kostenis, E.; et al. The adhesion G protein-coupled receptor GPR56/ADGRG1 is an inhibitory receptor on human NK cells. Cell. Rep. 2016, 15, 1757–1770. [Google Scholar] [CrossRef] [Green Version]
- Bi, J.; Tian, Z. NK cell dysfunction and checkpoint immunotherapy. Front. Immunol. 2019, 10, 1999. [Google Scholar] [CrossRef]
- Minuti, A.; Gallo, A.; Lopreiato, V.; Bruschi, S.; Piccioli-Cappelli, F.; Uboldi, O.; Trevisi, E. Effect of litter size on prepartum metabolic and amino acidic profile in rabbit does. Animal 2020, 14, 2109–2115. [Google Scholar] [CrossRef] [PubMed]
- Crookenden, M.A.; Moyes, K.M.; Kuhn-Sherlock, B.; Lehnert, K.; Walker, C.G.; Loor, J.J.; Mitchell, M.D.; Murray, A.; Dukkipati, V.S.R.; Vailati-Riboni, M.; et al. Transcriptomic analysis of circulating neutrophils in metabolically stressed peripartal grazing dairy cows. J. Dairy Sci. 2019, 102, 7408–7420. [Google Scholar] [CrossRef] [PubMed]
- Sharma, J.N.; Al-Omran, A.; Parvathy, S.S. Role of nitric oxide in inflammatory diseases. Inflammopharmacology 2007, 15, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Martino, M.R.; Gutierrez-Aguilar, M.; Yiew, N.K.H.; Lutkewitte, A.J.; Singer, J.M.; McCommis, K.S.; Ferguson, D.; Liss, K.H.H.; Yoshino, J.; Renkemeyer, M.K.; et al. Silencing alanine transaminase 2 in diabetic liver attenuates hyperglycemia by reducing gluconeogenesis from amino acids. Cell Rep. 2022, 39, 110733. [Google Scholar] [CrossRef] [PubMed]
- Tall, A.R.; Yvan-Charvet, L. Cholesterol, inflammation and innate immunity. Nat. Rev. Immunol. 2015, 15, 104–116. [Google Scholar] [CrossRef] [Green Version]
- Wathes, D.; Cheng, Z.; Mareiy, W.; Fouladi-Nasht, A. Polyunsaturated fatty acids and fertility in female mammals: An update. CABI Rev. 2013, 8, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Alipour, A.; van Oostrom, A.J.; Izraeljan, A.; Verseyden, C.; Collins, J.M.; Frayn, K.N.; Plokker, T.W.; Elte, J.W.; Castro Cabezas, M. Leukocyte activation by triglyceride-rich lipoproteins. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 792–797. [Google Scholar] [CrossRef] [Green Version]
- Castoldi, A.; Monteiro, L.B.; van Teijlingen Bakker, N.; Sanin, D.E.; Rana, N.; Corrado, M.; Cameron, A.M.; Hassler, F.; Matsushita, M.; Caputa, G.; et al. Triacylglycerol synthesis enhances macrophage inflammatory function. Nat. Commun. 2020, 11, 4107. [Google Scholar] [CrossRef]
- Bell, A.W. Regulation of organic nutrient metabolism during transition from late pregnancy to early lactation. J. Anim. Sci. 1995, 73, 2804–2819. [Google Scholar] [CrossRef]
- Moore, D.A.P.S.U.; Varga, G. BUN and MUN: Urea nitrogen testing in dairy cattle. Compendium 1996, 18, 712–720. [Google Scholar]
- Wathes, D.C.; Bourne, N.; Cheng, Z.; Mann, G.E.; Taylor, V.J.; Coffey, M.P. Multiple correlation analyses of metabolic and endocrine profiles with fertility in primiparous and multiparous cows. J. Dairy Sci. 2007, 90, 1310–1325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abe, R.; Donnelly, S.C.; Peng, T.; Bucala, R.; Metz, C.N. Peripheral blood fibrocytes: Differentiation pathway and migration to wound sites. J. Immunol. 2001, 166, 7556–7562. [Google Scholar] [CrossRef] [PubMed]
- Schreier, S.; Triampo, W. The blood circulating rare cell population. What is it and What is it Good For? Cells 2020, 9, 790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynn, A.K.; Yannas, I.V.; Bonfield, W. Antigenicity and immunogenicity of collagen. J. Biomed. Mater. Res. 2004, 71, 343–354. [Google Scholar] [CrossRef]
- Roche, J.R.; Burke, C.R.; Crookenden, M.A.; Heiser, A.; Loor, J.L.; Meier, S.; Mitchell, M.D.; Phyn, C.V.C.; Turner, S.A. Fertility and the transition dairy cow. Reprod. Fertil. Dev. 2017, 30, 85–100. [Google Scholar] [CrossRef]
- Galvao, K.N.; Flaminio, M.J.; Brittin, S.B.; Sper, R.; Fraga, M.; Caixeta, L.; Ricci, A.; Guard, C.L.; Butler, W.R.; Gilbert, R.O. Association between uterine disease and indicators of neutrophil and systemic energy status in lactating Holstein cows. J. Dairy Sci. 2010, 93, 2926–2937. [Google Scholar] [CrossRef] [Green Version]
- Dahl, M.O.; Maunsell, F.P.; De Vries, A.; Galvao, K.N.; Risco, C.A.; Hernandez, J.A. Evidence that mastitis can cause pregnancy loss in dairy cows: A systematic review of observational studies. J. Dairy Sci. 2017, 100, 8322–8329. [Google Scholar] [CrossRef] [Green Version]
- Hansen, P.J.; Soto, P.; Natzke, R.P. Mastitis and fertility in cattle-possible involvement of inflammation or immune activation in embryonic mortality. Am. J. Reprod. Immunol. 2004, 51, 294–301. [Google Scholar] [CrossRef]
- Malinowski, E.; Gajewski, Z. Mastitis and fertility disorders in cows. Pol. J. Vet. Sci. 2010, 13, 555–560. [Google Scholar]
- Edmonson, A.J.; Lean, I.J.; Weaver, L.D.; Farver, T.; Webster, G.A. A body condition scoring chart for Holstein dairy cows. J. Dairy Sci. 1989, 72, 11. [Google Scholar] [CrossRef]
- Larsen, T.; Rontved, C.M.; Ingvartsen, K.L.; Vels, L.; Bjerring, M. Enzyme activity and acute phase proteins in milk utilized as indicators of acute clinical E. coli LPS-induced mastitis. Animal 2010, 4, 1672–1679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krogh, M.A.; Hostens, M.; Salavati, M.; Grelet, C.; Sorensen, M.T.; Wathes, D.C.; Ferris, C.P.; Marchitelli, C.; Signorelli, F.; Napolitano, F.; et al. Between- and within-herd variation in blood and milk biomarkers in Holstein cows in early lactation. Animal 2020, 14, 1067–1075. [Google Scholar] [CrossRef] [PubMed]
- Buggiotti, L.; Cheng, Z.; Wathes, D.C.; Gplus, E.C. Mining the unmapped reads in bovine RNA-Seq data reveals the prevalence of bovine herpes virus-6 in European dairy cows and the associated changes in their phenotype and leucocyte transcriptome. Viruses 2020, 12, 1451. [Google Scholar] [CrossRef] [PubMed]
- Beltman, M.E.; Forde, N.; Furney, P.; Carter, F.; Roche, J.F.; Lonergan, P.; Crowe, M.A. Characterisation of endometrial gene expression and metabolic parameters in beef heifers yielding viable or non-viable embryos on Day 7 after insemination. Reprod. Fertil. Dev. 2010, 22, 987–999. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Multiparous | Primiparous | |||||
|---|---|---|---|---|---|---|
| Parameters 3 | LC | MC | HC | LC | MC | HC |
| n | 14 | 15 | 15 | 6 | 6 | 6 |
| Total DMI (kg/d) | 16.3 ± 0.8 c | 20.4 ± 0.9 b | 23.6 ± 0.9 a | 12.9 ± 0.6 b | 15.8 ± 0.52 a | 17.1 ± 0.6 a |
| Body weight (kg) | 656 ± 21 | 643 ± 14 | 669 ± 11 | 534 ± 16 | 536 ± 13 | 525 ± 17 |
| Milk yield (kg/d) | 33.4 ± 1.4 b | 35.9 ± 1.5 a,b | 37.9 ± 1.1 a | 20.7 ± 1.1 | 20.5 ± 2.0 | 23.3 ± 1.1 |
| ECM (kg/d) | 34.8 ± 1.6 b | 36.4 ± 1.5 a,b | 40.4 ± 1.4 a | 21.0 ± 1.2 | 22.0 ± 2.1 | 24.2 ± 1.2 |
| EBAL (MJ/d) | −11.0 ± 1.5 b | −4.6 ± 1.4 a | −1.3 ± 1.3 a | −4.5 ± 1.7 b | 0.1 ± 1.4 a,b | 1.6 ± 1.6 a |
| BCS | 2.64 ± 0.11 | 2.38 ± 0.08 | 2.53 ± 0.07 | 2.92 ± 0.11 | 2.88 ± 0.09 | 2.96 ± 0.15 |
| Glucose (mmol/L) | 3.30 ± 0.08 b | 3.34 ± 0.07 b | 3.61 ± 0.06 a | 3.82 ± 0.14 | 3.92 ± 0.10 | 3.98 ± 0.06 |
| Urea (mmol/L) | 3.47 ± 0.17 a | 3.01 ± 0.23 b | 2.54 ± 0.11 b | 4.38 ± 0.26 c | 2.70 ± 0.15 b | 2.17 ± 0.15 a |
| BHB (mmol/L) | 0.64 ± 0.08 a | 0.60 ± 0.06 a,b | 0.42 ± 0.05 b | 0.44 ± 0.03 | 0.40 ± 0.04 | 0.33 ± 0.04 |
| NEFAs (mmol/L) | 796 ± 104 | 656 ± 95 | 608 ± 81 | 502 ± 149 | 350 ± 99 | 462 ± 119 |
| IGF-1 (ng/mL) | 56 ± 7 b | 55 ± 7 b | 103 ± 10 a | 110 ± 22 | 160 ± 20 | 167 ± 29 |
| Multiparous | Primiparous | |||||
|---|---|---|---|---|---|---|
| Parameters 3 | LC | MC | HC | LC | MC | HC |
| n | 14 | 15 | 15 | 6 | 6 | 6 |
| Uterine PMNs/HPF | 7.86 ± 1.85 | 8.82 ± 1.58 | 10.74 ± 2.11 | 4.68 ± 1.79 | 9.90 ± 3.16 | 10.03 ± 2.45 |
| Uterine EC/HPF | 12.49 ± 2.12 | 10.22 ± 2.02 | 9.09 ± 1.81 | 14.07 ± 1.48 | 7.25 ± 2.08 | 8.42 ± 2.31 |
| Uterine PMNs:UEC | 3.09 ± 1.49 | 3.56 ± 1.70 | 4.56 ± 1.76 | 0.39 ± 0.15 b | 1.90 ± 0.64 a,b | 2.10 ± 0.72 a |
| Log10 SCC (1000 cells/mL) | 1.85± 0.80 a | 1.55 ± 0.33 b | 1.32 ± 0.24 b | 1.74 ± 0.28 | 1.88 ± 0.51 | 2.09 ± 0.70 |
| Cases of clinical mastitis 4 (n) | 4 | 0 | 0 | 0 | 2 | 1 |
| Cases of subclinical mastitis (n) | 0 | 1 | 0 | 1 | 0 | 2 |
| % clinical and subclinical mastitis | 28.6% | 6.7% | 0% | 16.7% | 33.3% | 50% |
| Milk Nagase (mU/mL) | 2.99 ± 1.18 | 1.81 ± 0.12 | 1.78 ± 0.13 | 2.15 ± 0.18 | 2.40 ± 0.51 | 2.56 ± 0.46 |
| Milk LDH (mU/mL) | 5.20 ± 1.65 | 3.03 ± 0.28 | 2.93 ± 0.34 | 3.92 ± 0.44 | 4.79 ± 0.77 | 5.18 ± 0.88 |
| Multiparous | Primiparous | |||||
|---|---|---|---|---|---|---|
| Parameters 3 | LC | MC | HC | LC | MC | HC |
| n | 14 | 15 | 15 | 6 | 6 | 6 |
| DFS | 80.4 ± 4.1 | 89.3 ± 6.7 | 91.1 ± 9.5 | 77.5 ± 6.4 | 76.8 ± 8.8 | 87.0 ± 8.4 |
| Days open | 117.0 ± 11.9 | 109.5 ± 8.0 | 108.7 ± 13.0 | 82.0 ± 5.1 b | 115.0 ± 22.7 a,b | 111.3 ± 1.3 a |
| S/C | 2.4 ± 0.4 | 1.8 ± 0.3 | 1.6 ± 0.2 | 1.0 ± 0.0 | 2.0 ± 0.5 | 2.0 ± 0.4 |
| Not served (n) | 2 | 3 | 4 | 0 | 0 | 2 |
| FTC (n) | 2 | 1 | 2 | 1 | 0 | 1 |
| Pregnant (n) | 10 | 11 | 9 | 5 | 6 | 3 |
| ICB score 4 | 2.2 ± 0.3 | 2.1 ± 0.3 | 2.5 ± 0.4 | 1.5 ± 0.5 b | 1.3 ± 0.2 b | 3.0 ± 0.4 a |
| Functions | Enrichment Score | Upregulated or Downregulated DEGs in the Function |
|---|---|---|
| Response to stimulus | 10.9 | Upregulated DEGs (12): ALAS2, APLP1, COL1A1, COL1A2, COL3A1, GZMB, HTR1B, LIF, MS4A2, OR56A1, P2RY12, SPP1 Downregulated DEGs (33): AHSG, ALB, ALOX5, APOA2, CELA2A, CFB, CYP1A2, FABP1, FGA, FGB, FGG, GATM, GLYAT, IFI6, IFNB1, ISG15, KNG1, MPO, MX2, NUPR1, OAS1Y, OAS1Z, OAS2, ORM1, PCK1, PRKCG, RBP4, RSAD2, SELENOM, TF, TRPC3, VTN |
| Interspecies interaction between organisms | 10.9 | Upregulated DEGs (1): GZMB Downregulated DEGs (14): APOA2, CFB, FGA, FGB, IFI6, IFNB1, ISG15, MPO, MX2, OAS1Y, OAS1Z, OAS2, PCK1, RSAD2 |
| Biomineralisation | 7.1 | Upregulated DEGs (4): COL1A1, COL1A2, SPP1, TUFT1 Downregulated DEGs (0): |
| Immune system process | 6.5 | Upregulated DEGs (3): GZMB, LIF, MS4A2 Downregulated DEGs (11): ALOX5, CFB, FGA, FGB, IFI6, MPO, MX2, OAS1Y, OAS1Z, RSAD2, VTN |
| Detoxification | 5.9 | Upregulated DEGs (1): HBB Downregulated DEGs (3): FABP1, GSTA1, MPO |
| Metabolic process | 5.4 | Upregulated DEGs (16): ADAMDEC1, ADAMTS3, ALAS2, AOX1, APLP1, COL1A1, COL1A2, COL3A1, GZMB, IL1R2, LOC100139881, MAPK10, NRIP3, RNASE12, SPP1, ZNF215 Downregulated DEGs (31): ACAN, ALDOB, ALOX5, APOA2, APOC3, APOH, CELA2A, CFB, CYP1A2, CYP2E1, FGB, FGG, GATM, GC, GLYAT, GSTA1, HMGCS2, HPX, IFI6, ISG15, ITIH2, MPO, NUPR1, PCK1, PPP1R3C, PRKCG, PROC, PTPN3, RBP4, SELENOM, TTR |
| Multicellular organismal process | 5.2 | Upregulated DEGs (5): ADAMTS3, COL1A1, COL3A1, HTR1B, SPP1 Downregulated DEGs (16): AHSG, ALOX5, APOA2, APOC3, CLCN1, CYP1A2, FGB, FGG, GATM, KNG1, MPO, PRKCG, PROC, RBP4, SELENOM, SPP2 |
| Biological regulation | 3.85 | Upregulated DEGs (24): ADAMTS3, APLP1, CDKN1C, COL1A1, COL1A2, COL3A1, DAB2, GCSAML, GZMB, HTR1B, IL1R2, IL20RA, LIF, LOC281376, MAPK10, MEIS3, MS4A2, MYCL, OR56A1, P2RY12, PDE9A, PPP1R1B, RSPO1, SPP1 Downregulated DEGs (43): AHSG, ALB, ALDOB, ALOX5, AMBP, APOA2, APOC3, APOH, CELA2A, CFB, CLCN1, CYP1A2, DDX25, ELN, FABP1, FAM3B, FGB, FGG, GATM, HPX, IFI6, IFNB1, ISG15, ITIH2, KNG1, MX2, NUPR1, OAS2, OR12D2, ORM1, PCK1, PRKCG, PROC, PTPN3, RBP4, RHOBTB1, RSAD2, SELENOM, SERPINA3-7, TF, TRPC3, TTR, VTN |
| Pathways | Enrichment FDR p-Value | Number of DEGs |
|---|---|---|
| Complement and coagulation cascades | 9.972 × 10−5 | 8 |
| Drug metabolism—cytochrome P450 | 1.853 × 10−3 | 5 |
| Platelet activation | 1.853 × 10−3 | 7 |
| Protein digestion and absorption | 4.322 × 10−3 | 6 |
| ECM-receptor interaction | 6.542 × 10−3 | 5 |
| Metabolism of xenobiotics by cytochrome P450 | 1.200 × 10−2 | 4 |
| Hepatitis C | 1.200 × 10−2 | 6 |
| Focal adhesion | 1.207 × 10−2 | 7 |
| Chemical carcinogenesis | 1.377 × 10−2 | 4 |
| Amoebiasis | 1.377 × 10−2 | 5 |
| Influenza A | 1.377 × 10−2 | 7 |
| Fc epsilon RI signalling pathway | 2.039 × 10−2 | 4 |
| PPAR signalling pathway | 2.039 × 10−2 | 4 |
| NOD-like receptor signalling pathway | 2.433 × 10−2 | 6 |
| Cholesterol metabolism | 2.738 × 10−2 | 3 |
| Glycolysis/Gluconeogenesis | 4.341 × 10−2 | 3 |
| AGE-RAGE signalling pathway in diabetic complications | 4.341 × 10−2 | 4 |
| Measles | 4.341 × 10−2 | 5 |
| Herpes simplex infection | 4.341 × 10−2 | 6 |
| Retinol metabolism | 4.401 × 10−2 | 3 |
| Thyroid hormone synthesis | 4.401 × 10−2 | 3 |
| Drug metabolism—other enzymes | 4.942 × 10−2 | 3 |
| Sphingolipid signalling pathway | 4.942 × 10−2 | 4 |
| Functions | Enrichment Score | Upregulated and Downregulated DEGs in the Function |
|---|---|---|
| Immune system process | 17.0 | Upregulated DEGs (39): ACOD1, ADAM8, ADGRG3, ARG2, BCL6, C1RL, C5AR1, C5AR2, CD14, CD24, CFP, CLEC4A, CLEC4D, CLEC4E, CXCL13, FES, HCK, HLX, HP, IL1A, LST1, LTF, MMP9, MSRB1, NDRG1, NFIL3, NLRP1, PGLYRP1, PGLYRP4, S100A12, S100A8, S100A9, SEMA4A, SKAP2, SLC11A1, THY1, TLR4, TREM1, TYROBP Downregulated DEGs (5): BCAR1, CDH17, NKG7, PRF1, SRMS |
| Locomotion | 15.2 | Upregulated DEGs (16): ADAM8, C5AR1, C5AR2, CCL16, CXCL13, CXCR1, CXCR2, DEFB1, DEFB10, DEFB7, NRG1, NRP1, PROK2, PTAFR, S100A8, S100A9 Downregulated DEGs (2): BCAR1, WNT5A |
| Interspecies interaction between organisms | 10.3 | Upregulated DEGs (27): ARG2, C5AR1, CATHL6, CD14, CFP, CLEC4D, CLEC4E, CLEC5A, CXCL13, DEFB1, FN1, HMOX1, HP, LTF, MSRB1, NECTIN2, NLRP1, NRP1, PGLYRP1, PGLYRP4, S100A12, S100A8, S100A9, SCARB1, SLC11A1, TLR4, TREM1 Downregulated DEGs (3): GZMA, NKG7, PRF1 |
| Cellular process | 5.7 | Upregulated DEGs (210): ABCA6, ACSL6, ACVR1B, ADAM8, ADCY6, ADGRG3, ALDH1L2, ALOX5AP, ALPK1, ALPK3, AMPD3, ANGPTL3, ANKS4B, APH1B, ARAP3, ARG2, ARHGAP22, ARRDC4, ASS1, ATF3, BCL6, BMX, C5AR1, C5AR2, CAMKK1, CAPN3, CARMIL1, CATHL6, CCR1, CD14, CD24, CDS1, CHI3L1, CLEC4A, CLEC4D, CLEC4E, CLEC5A, COL1A1, COL1A2, COL3A1, CREB3L2, CRISPLD2, CTBP2, CXCL13, CXCR1, CXCR2, DACH1, DAGLB, DCK, DCN, DEFB1, DGAT2, DGKG, DIRAS3, DISC1, DLC1, DNER, DOCK4, DPYD, DUSP1, DYSF, EGR2, EIF4E3, EIF4EBP1, ELN, ENKUR, EREG, ETV5, FAT1, FES, FLT3, FMO2, FN1, FOS, FOSL2, GAB2, GADD45A, GGT5, GK, GPAT3, GPR27, GPR87, GPT2, GPX3, GSTO2, HAL, HCK, HEPACAM2, HK3, HLX, HMOX1, HP, HRH2, ICAM3, IFIT3, IL17RD, IL1A, IL1RAP, IL1RN, ISG20, KCNJ2, KREMEN1, LAMA3, LGR6, LOC100337213, LOC112447333, LOC514257, LOC521224, LOC527744, LOC538435, LTF, MAP3K6, MAPK13, MARCKSL1, MMP8, MMP9, MPV17L, MSRB1, MZB1, NAIP, NDRG1, NECTIN2, NFAM1, NFIL3, NLRP1, NRP1, OR52B2, OR52L1, OR52W1, OSCAR, PADI4, PCSK1, PDXK, PDZD3, PFKFB4, PGLYRP1, PGLYRP4, PIGR, PKD2, PLAUR, PLB1, PLIN3, PLXND1, PPP1R3B, PRKAA2, PRODH, PROK2, PTAFR, PTPN5, RAB20, RAB3D, RAB3IP, RBM47, RECK, RETN, RIDA, RND3, RPH3A, S100A12, S100A8, S100A9, SCARB1, SDS, SEMA4A, SEMA7A, SERINC2, SERPINE1, SH3PXD2B, SKAP2, SLC11A1, SLC13A3, SLC13A5, SLC16A3, SLC16A5, SLC28A3, SLC2A9, SLC30A4, SLC40A1, SLC5A9, SLC6A9, SLC8A1, SLCO4C1, SNX18, SNX24, SOCS3, SOCS6, SORT1, SPATA6, SRGN, STARD9, STEAP4, TARM1, TBXAS1, TCN1, TGFA, TGFBI, THY1, TJP2, TLR4, TNFAIP6, TNFRSF8, TYROBP, UGGT2, UPP1, USP35, VCAN, VLDLR, WDFY3, WIPI1, WLS Downregulated DEGs (28): ADGRG1, AOX1, ATP6V0A4, BACE1, BCAR1, CA8, CCR5, CD96, CDH17, CHN1, EPCAM, GRM8, GSDMA, GZMA, LOC784535, NKG7, PRF1, S1PR5, SAXO1, SCN11A, SLC22A23, SLC38A11, SLC6A15, SLC7A4, SRMS, SRPK3, TDRKH, WNT5A, ZWILCH |
| Detoxification | 4.5 | Upregulated DEGs (6): ALOX5AP, GPX3, GSTO2, HP, S100A8, S100A9 Downregulated DEGs (0) |
| Biological regulation | 3.5 | Upregulated DEGs (115): A2M, ACOD1, ACVR1B, ADAM8, ADGRG3, ALPK1, ANGPTL3, APH1B, ARG2, ARRDC4, ATF3, BASP1, BCL6, BST1, C1RL, C5AR1, C5AR2, CAPN3, CARMIL1, CCR1, CD101, CD14, CD24, CFP, CHI3L1, CLEC4A, CLEC4D, CLEC4E, CLEC5A, COL1A1, COL3A1, CREB3L2, CREB5, CTBP2, CXCL13, DACH1, DAGLB, DCN, DEFB1, DGKG, DISC1, DLC1, DOCK4, DUSP1, EGR2, EIF4EBP1, EREG, ETV5, FES, FMO2, FN1, FOS, FOSL2, GADD45A, GPAT3, GPR27, HCK, HLX, IL17RD, IL1A, IL1RAP, KREMEN1, LAMA3, LGR6, LOC100298356, LOC101903018, LOC515676, LRP1, LTF, MAPK13, MMP8, MMP9, MN1, MPV17L, MXD1, MZB1, NAIP, NECTIN2, NFAM1, NFIL3, NLRP1, NLRP12, NRG1, NRP1, PDZD3, PIGR, PLAUR, PPP1R3B, PRKAA2, RAP1GAP, RECK, RETN, RIDA, S100A8, S100A9, SCARB1, SEMA4A, SEMA7A, SLC11A1, SNX18, SOCS3, SOCS6, SORT1, TARM1, TGFA, THY1, TLR4, TNFAIP6, TNFRSF8, TYROBP, VLDLR, WIPI1, WLS, WWC2, ZNF114 Downregulated DEGs (10): ADGRG1, BCAR1, CD96, CHN1, EOMES, EPCAM, FCRL3, GZMA, NKG7, WNT5A |
| Signalling | 3.2 | Upregulated DEGs (6): CXCL13, EREG, IL1RAP, KCNJ2, PKD2, THY1 Downregulated DEGs (2): CCR5, WNT5A |
| Pathways | Enrichment FDR p-Value | Number of DEGs |
|---|---|---|
| Osteoclast differentiation | 1.660 × 10−4 | 11 |
| Relaxin signalling pathway | 3.384 × 10−4 | 10 |
| Amoebiasis | 4.880 × 10−4 | 9 |
| Fluid shear stress and atherosclerosis | 1.435 × 10−3 | 10 |
| Valine, leucine and isoleucine biosynthesis | 1.508 × 10−3 | 2 |
| Chemokine signalling pathway | 5.046 × 10−3 | 10 |
| ECM-receptor interaction | 5.677 × 10−3 | 6 |
| AGE-RAGE signalling pathway in diabetic complications | 8.527 × 10−3 | 7 |
| Vitamin B6 metabolism | 8.580 × 10−3 | 2 |
| Growth hormone synthesis, secretion and action | 9.80 × 10−3 | 7 |
| Arginine biosynthesis | 1.096 × 10−2 | 3 |
| Biosynthesis of amino acids | 1.124 × 10−2 | 5 |
| Platelet activation | 1.129 × 10−2 | 7 |
| Aldosterone synthesis and secretion | 1.326 × 10−2 | 6 |
| Cortisol synthesis and secretion | 1.355 × 10−2 | 5 |
| Drug metabolism—other enzymes | 1.549 × 10−2 | 5 |
| Cholesterol metabolism | 1.743 × 10−2 | 4 |
| Cholinergic synapse | 1.959 × 10−2 | 6 |
| Protein digestion and absorption | 2.162 × 10−2 | 6 |
| Glutathione metabolism | 2.364 × 10−2 | 4 |
| MAPK signalling pathway | 2.376 × 10−2 | 12 |
| Drug metabolism—cytochrome P450 | 2.479 × 10−2 | 4 |
| Arachidonic acid metabolism | 2.504 × 10−2 | 5 |
| TNF signalling pathway | 3.116 × 10−2 | 6 |
| PI3K-Akt signalling pathway | 3.183 × 10−2 | 14 |
| Salmonella infection | 3.656 × 10−2 | 5 |
| Pertussis | 3.656 × 10−2 | 5 |
| Longevity regulating pathway | 3.656 × 10−2 | 5 |
| Amphetamine addiction | 4.122 × 10−2 | 4 |
| Glycerolipid metabolism | 4.282 × 10−2 | 4 |
| Dopaminergic synapse | 4.310 × 10−2 | 6 |
| Phospholipase D signalling pathway | 4.439 × 10−2 | 7 |
| Complement and coagulation cascades | 4.534 × 10−2 | 5 |
| Thyroid hormone synthesis | 4.610 × 10−2 | 4 |
| Pantothenate and CoA biosynthesis | 4.805 × 10−2 | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, Z.; McLaughlin, D.L.; Little, M.W.; Ferris, C.; Salavati, M.; Ingvartsen, K.L.; Crowe, M.A.; Wathes, D.C.; the GplusE Consortium. Proportion of Concentrate in the Diet of Early Lactation Dairy Cows Has Contrasting Effects on Circulating Leukocyte Global Transcriptomic Profiles, Health and Fertility According to Parity. Int. J. Mol. Sci. 2023, 24, 39. https://doi.org/10.3390/ijms24010039
Cheng Z, McLaughlin DL, Little MW, Ferris C, Salavati M, Ingvartsen KL, Crowe MA, Wathes DC, the GplusE Consortium. Proportion of Concentrate in the Diet of Early Lactation Dairy Cows Has Contrasting Effects on Circulating Leukocyte Global Transcriptomic Profiles, Health and Fertility According to Parity. International Journal of Molecular Sciences. 2023; 24(1):39. https://doi.org/10.3390/ijms24010039
Chicago/Turabian StyleCheng, Zhangrui, Danielle L. McLaughlin, Mark W. Little, Conrad Ferris, Mazdak Salavati, Klaus L. Ingvartsen, Mark A. Crowe, D. Claire Wathes, and the GplusE Consortium. 2023. "Proportion of Concentrate in the Diet of Early Lactation Dairy Cows Has Contrasting Effects on Circulating Leukocyte Global Transcriptomic Profiles, Health and Fertility According to Parity" International Journal of Molecular Sciences 24, no. 1: 39. https://doi.org/10.3390/ijms24010039