
Comparative Transcriptome Profiling of Cassava Tuberous Roots in Response to Postharvest Physiological Deterioration

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Comparison of the PPD Tolerance of SC8 and RYG1 Cassava Tuberous Roots during Storage
2.2. Microscopic Structure of SC8 and RYG1 Cassava
2.3. Assembly of Transcriptomes and Quality Assessments
2.4. Identification of Differentially Expressed Genes
2.5. Venn Analysis of the DEGs
2.6. GO Functional Enrichment Analysis of Co-DEGs
2.7. KEGG Pathway Analysis of co-DEGs
2.8. Differentially Expressed Genes Related to Photosynthesis
2.9. Differentially Expressed Genes in Protein Processing in the Endoplasmic Reticulum Pathway
2.10. Differentially Expressed Genes Related to Starch and Sucrose Metabolism and Galactose Metabolism
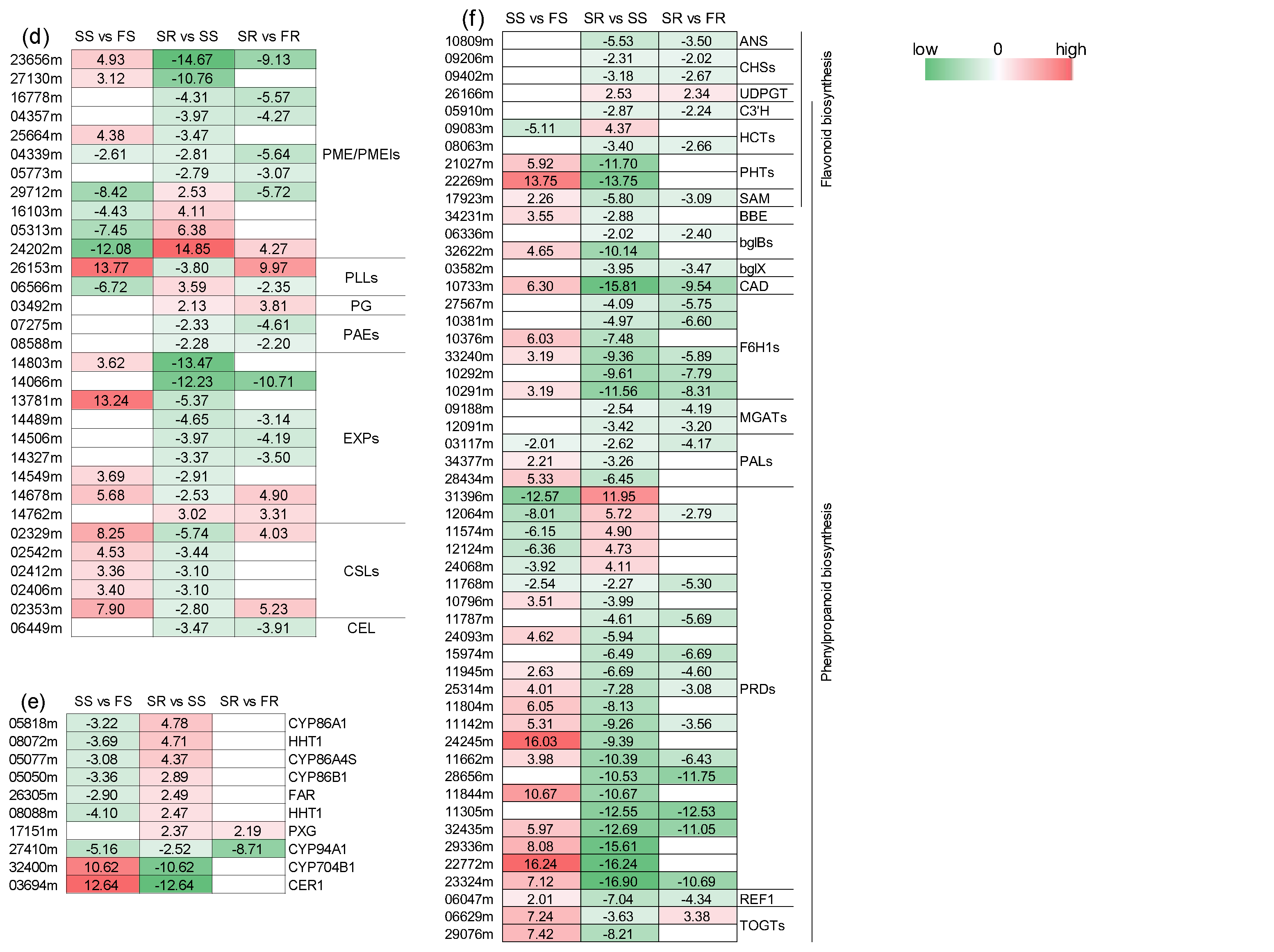
2.11. Differentially Expressed Genes Related to Cell Wall, Cutin, Suberin and Wax Biosynthesis, Phenylpropanoid Biosynthesis and Flavonoid Biosynthesis
2.12. Differentially Expressed Genes Related to Hormone Signaling
2.13. Differentially Expressed Genes Related to Transcriptional Regulation
2.14. Protein–Protein Interaction Network Construction and Hub Gene Identification
3. Discussion
4. Materials and Methods
4.1. Plant Treatment and Tissue Sampling
4.2. Transmission Electron Microscope Observations
4.3. RNA-Seq Transcriptome Analysis
4.4. Screening of Differentially Expressed Genes (DEGs) and Enrichment Analysis
4.5. Enrichment Analysis, PPI Interaction Network Construction of co-DEGs
4.6. Quantitative Real-Time PCR Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wilson, M.C.; Mutka, A.M.; Hummel, A.W.; Berry, J.; Chauhan, R.D.; Vijayaraghavan, A.; Taylor, N.J.; Voytas, D.F.; Chitwood, D.H.; Bart, R.S. Gene expression atlas for the food security crop cassava. N. Phytol. 2017, 213, 1632–1641. [Google Scholar] [CrossRef] [Green Version]
- Tize, I.; Fotso, A.K.; Nukenine, E.N.; Masso, C.; Ngome, F.A.; Suh, C.; Lendzemo, V.W.; Nchoutnji, I.; Manga, G.; Parkes, E.; et al. New cassava germplasm for food and nutritional security in Central Africa. Sci. Rep. 2021, 11, 7394. [Google Scholar] [CrossRef]
- Talsma, E.F.; Borgonjen-van den Berg, K.J.; Melse-Boonstra, A.; Mayer, E.V.; Verhoef, H.; Demir, A.Y.; Ferguson, E.L.; Kok, F.J.; Brouwer, I.D. The potential contribution of yellow cassava to dietary nutrient adequacy of primary-school children in Eastern Kenya; the use of linear programming. Public Health Nutr. 2018, 21, 365–376. [Google Scholar] [CrossRef] [Green Version]
- Sonnewald, U.; Fernie, A.R.; Gruissem, W.; Schläpfer, P.; Anjanappa, R.B.; Chang, S.-H.; Ludewig, F.; Rascher, U.; Muller, O.; van Doorn, A.M.; et al. The Cassava Source–Sink project: Opportunities and challenges for crop improvement by metabolic engineering. Plant J. 2020, 103, 1655–1665. [Google Scholar] [CrossRef]
- Bizzuti, B.E.; de Abreu Faria, L.; da Costa, W.S.; Lima, P.M.T.; Ovani, V.S.; Kruger, A.M.; Louvandini, H.; Abdalla, A.L. Potential use of cassava by-product as ruminant feed. Trop. Anim. Health Prod. 2021, 53, 108. [Google Scholar] [CrossRef]
- Bakare, A.G.; Zindove, T.J.; Iji, P.A.; Stamatopoulos, K.; Cowieson, A.J. A review of limitations to using cassava meal in poultry diets and the potential role of exogenous microbial enzymes. Trop. Anim. Health Prod. 2021, 53, 426. [Google Scholar] [CrossRef]
- Jiang, D.; Hao, M.; Fu, J.; Tian, G.; Ding, F. Estimating the potential of energy saving and carbon emission mitigation of cassava-based fuel ethanol using life cycle assessment coupled with a biogeochemical process model. Int. J. Biometeorol. 2019, 63, 701–710. [Google Scholar] [CrossRef]
- Lin, Z.; Liu, H.; Wu, J.; Patakova, P.; Branska, B.; Zhang, J. Effective continuous acetone-butanol-ethanol production with full utilization of cassava by immobilized symbiotic TSH06. Biotechnol. Biofuels 2019, 12, 219. [Google Scholar] [CrossRef]
- Nwose, E.U.; Onodu, B.C.; Anyasodor, A.E.; Sedowo, M.O.; Okuzor, J.N.; Culas, R.J. Ethnopharmacological values of cassava and its potential for diabetes and dyslipidemia management: Knowledge survey and critical review of report. J. Intercult. Ethnopharmacol. 2017, 6, 260–266. [Google Scholar] [CrossRef]
- Djabou, A.S.M.; Carvalho, L.; Li, Q.X.; Niemenak, N.; Chen, S. Cassava postharvest physiological deterioration: A complex phenomenon involving calcium signaling, reactive oxygen species and programmed cell death. Acta Physiol. Plant 2017, 39, 91. [Google Scholar] [CrossRef]
- Naziri, D.; Quaye, W.; Siwoku, B.; Wanlapatit, S.; Viet, P.; Bennett, C. JJoA, Tropics RDit, Subtropics: The diversity of postharvest losses in cassava value chains in selected developing countries. J. Agric. Rural. Dev. Trop. Subtrop. 2014, 115, 111–123. [Google Scholar]
- García, J.A.; Sánchez, T.; Ceballos, H.; Alonso, L. Non-destructive sampling procedure for biochemical or gene expression studies on post-harvest physiological deterioration of cassava roots. Postharvest Biol. Tec. 2013, 86, 529–535. [Google Scholar] [CrossRef]
- Zainuddin, I.M.; Fathoni, A.; Sudarmonowati, E.; Beeching, J.R.; Gruissem, W.; Vanderschuren, H. Cassava post-harvest physiological deterioration: From triggers to symptoms. Postharvest Biol. Tec. 2018, 142, 115–123. [Google Scholar] [CrossRef]
- Njoku, D.N.; Amadi, C.O.; Mbe, J.; Amanze, N.J. Strategies to overcome post-harvest physiological deterioration in cassava (Manihot esculenta) root: A review. Niger. Agric. J. 2014, 45, 126568023. [Google Scholar]
- Cortes, D.F.; Reilly, K.; Okogbenin, E.; Beeching, J.R.; Iglesias, C.; Tohme, J. Mapping wound-response genes involved in post-harvest physiological deterioration (PPD) of cassava (Manihot esculenta Crantz). Euphytica 2002, 128, 47–53. [Google Scholar] [CrossRef]
- Brito, A.C.; Oliveira, S.A.S.; Oliveira, E.J. Genome-wide association study for resistance to cassava root rot. J. Agric. Sci. 2017, 155, 1424–1441. [Google Scholar] [CrossRef]
- de Andrade, L.R.B.; Sousa, M.B.E.; Oliveira, E.J.; de Resende, M.D.V.; Azevedo, C.F. Cassava yield traits predicted by genomic selection methods. PLoS ONE 2019, 14, e0224920. [Google Scholar] [CrossRef] [Green Version]
- Torres, L.G.; de Resende, M.D.V.; Azevedo, C.F.; Silva, F.F.E.; de Oliveira, E.J. Genomic selection for productive traits in biparental cassava breeding populations. PLoS ONE 2019, 14, e0220245. [Google Scholar] [CrossRef] [Green Version]
- Lima, L.P.; Azevedo, C.F.; de Resende, M.D.V.; Silva, F.F.E.; Viana, J.M.S.; de Oliveira, E.J. Triple categorical regression for genomic selection: Application to cassava breeding. Sci. Agric. 2019, 76, 368–375. [Google Scholar] [CrossRef] [Green Version]
- Esuma, W.; Ozimati, A.; Kulakow, P.; Gore, M.A.; Wolfe, M.D.; Nuwamanya, E.; Egesi, C.; Kawuki, R.S. Effectiveness of genomic selection for improving provitamin A carotenoid content and associated traits in cassava. G3 Genes Genomes Genet. 2021, 11, jkab160. [Google Scholar] [CrossRef]
- Ozimati, A.; Kawuki, R.; Esuma, W.; Kayondo, S.I.; Pariyo, A.; Wolfe, M.; Jannink, J.L. Genetic Variation and Trait Correlations in an East African Cassava Breeding Population for Genomic Selection. Crop. Sci. 2019, 59, 460–473. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Mao, L.; Lu, W.; Ying, T.; Luo, Z. Transcriptome profiling of postharvest strawberry fruit in response to exogenous auxin and abscisic acid. Planta 2016, 243, 183–197. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Zhang, J.; Jiao, C.; Yin, X.; Fei, Z.; Wu, Q.; Chen, K. Transcriptome analysis provides insights into the regulation of metabolic processes during postharvest cold storage of loquat (Eriobotrya japonica) fruit. Hortic. Res. 2019, 6, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.Y.; Dou, G.X.; Sun, X.X.; Chen, L.; Zheng, Y.; Xiao, H.M.; Wang, Y.P.; Li, H.Y.; Guo, J.H.; Jiang, C.H. Transcriptome and Biochemical Analysis Jointly Reveal the Effects of Bacillus cereus AR156 on Postharvest Strawberry Gray Mold and Fruit Quality. Front. Plant. Sci. 2021, 12, 700446. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.X.; Zou, Y.; Yu, Y.H.; Chen, B.X.; Zheng, Q.W.; Ye, Z.W.; Wei, T.; Ye, S.Q.; Guo, L.Q.; Lin, J.F. Comparative transcriptome and proteome provide new insights into the regulatory mechanisms of the postharvest deterioration of Pleurotus tuoliensis fruitbodies during storage. Food Res. Int. 2021, 147, 110540. [Google Scholar] [CrossRef]
- Ding, Y.; Chang, J.; Ma, Q.; Chen, L.; Liu, S.; Jin, S.; Han, J.; Xu, R.; Zhu, A.; Guo, J.; et al. Network analysis of postharvest senescence process in citrus fruits revealed by transcriptomic and metabolomic profiling. Plant Physiol. 2015, 168, 357–376. [Google Scholar] [CrossRef]
- Li, T.; Wu, Q.; Duan, X.; Yun, Z.; Jiang, Y. Proteomic and transcriptomic analysis to unravel the influence of high temperature on banana fruit during postharvest storage. Funct. Integr. Genomics 2019, 19, 467–486. [Google Scholar] [CrossRef]
- Lin, Q.; Xie, Y.; Guan, W.; Duan, Y.; Wang, Z.; Sun, C. Combined transcriptomic and proteomic analysis of cold stress induced sugar accumulation and heat shock proteins expression during postharvest potato tuber storage. Food Chem. 2019, 297, 124991. [Google Scholar] [CrossRef]
- Huang, J.; Bachem, C.; Jacobsen, E.; Visser, R.G.F. Molecular analysis of differentially expressed genes during postharvest deterioration in cassava (Manihot esculenta Crantz) tuberous roots. Euphytica 2001, 120, 85–93. [Google Scholar] [CrossRef]
- Reilly, K.; Bernal, D.; Cortes, D.F.; Gomez-Vasquez, R.; Tohme, J.; Beeching, J.R. Towards identifying the full set of genes expressed during cassava post-harvest physiological deterioration. Plant Mol. Biol. 2007, 64, 187–203. [Google Scholar] [CrossRef]
- Hu, W.; Kong, H.; Guo, Y.; Zhang, Y.; Ding, Z.; Tie, W.; Yan, Y.; Huang, Q.; Peng, M.; Shi, H.; et al. Comparative Physiological and Transcriptomic Analyses Reveal the Actions of Melatonin in the Delay of Postharvest Physiological Deterioration of Cassava. Front. Plant. Sci. 2016, 7, 736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Q.; Zhang, T.; Zhang, P.; Wang, Z.Y. Melatonin attenuates postharvest physiological deterioration of cassava storage roots. J. Pineal Res. 2016, 60, 424–434. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Zhao, S.; Ding, Z.; Tie, W.; Hu, W. Comparative Transcriptomic Analysis of Storage Roots in Cassava during Postharvest Physiological Deterioration. Plant Mol. Biol. Rep. 2021, 39, 607–616. [Google Scholar] [CrossRef]
- Uarrota, V.G.; Nunes Eda, C.; Peruch, L.A.; Neubert Ede, O.; Coelho, B.; Moresco, R.; Dominguez, M.G.; Sanchez, T.; Melendez, J.L.; Dufour, D.; et al. Toward better understanding of postharvest deterioration: Biochemical changes in stored cassava (Manihot esculenta Crantz) roots. Food Sci. Nutr. 2016, 4, 409–422. [Google Scholar] [CrossRef] [Green Version]
- Yuan, S.; Li, R.; Zhou, Y.; Qiu, T.; Yao, Y.; Liu, J.; Duan, R.; Fu, S.; Hu, X.; Guo, J. Comparative Analysis of Agronomic Traits and Leaf Physiological Indexes of Two Cassava Germplasms in Field. Mol. Plant Breed. 2018, 16, 2316–2321. [Google Scholar]
- Owiti, J.; Grossmann, J.; Gehrig, P.; Dessimoz, C.; Laloi, C.; Hansen, M.B.; Gruissem, W.; Vanderschuren, H. iTRAQ-based analysis of changes in the cassava root proteome reveals pathways associated with post-harvest physiological deterioration. Plant J. 2011, 67, 145–156. [Google Scholar] [CrossRef]
- Eum, H.L.; Han, S.H.; Lee, E.J. High-CO2 Treatment Prolongs the Postharvest Shelf Life of Strawberry Fruits by Reducing Decay and Cell Wall Degradation. Foods 2021, 10, 1649. [Google Scholar] [CrossRef]
- Zhu, Y.; Wang, K.; Wu, C.; Zhao, Y.; Yin, X.; Zhang, B.; Grierson, D.; Chen, K.; Xu, C. Effect of Ethylene on Cell Wall and Lipid Metabolism during Alleviation of Postharvest Chilling Injury in Peach. Cells 2019, 8, 1612. [Google Scholar] [CrossRef] [Green Version]
- Shi, Z.; Yang, H.; Jiao, J.; Wang, F.; Lu, Y.; Deng, J. Effects of graft copolymer of chitosan and salicylic acid on reducing rot of postharvest fruit and retarding cell wall degradation in grapefruit during storage. Food Chem. 2019, 283, 92–100. [Google Scholar] [CrossRef]
- An, F.; Baker, M.R.; Qin, Y.; Chen, S.; Li, Q.X. Relevance of Class I alpha-Mannosidases to Cassava Postharvest Physiological Deterioration. ACS Omega 2019, 4, 8739–8746. [Google Scholar] [CrossRef] [Green Version]
- Michailidis, M.; Karagiannis, E.; Tanou, G.; Sarrou, E.; Stavridou, E.; Ganopoulos, I.; Karamanoli, K.; Madesis, P.; Martens, S.; Molassiotis, A. An integrated metabolomic and gene expression analysis identifies heat and calcium metabolic networks underlying postharvest sweet cherry fruit senescence. Planta 2019, 250, 2009–2022. [Google Scholar] [CrossRef] [PubMed]
- Wen, S.Y.; Neuhaus, H.E.; Cheng, J.T.; Bie, Z.L. Contributions of sugar transporters to crop yield and fruit quality. J. Exp. Bot. 2022, 73, 2275–2289. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.P.; Zhang, Z.J.; Wang, X.C.; Yuan, X.; Wu, Q.; Chen, S.Y.; Zou, Y.J.; Ma, F.W.; Li, C. Exogenous dopamine improves apple fruit quality via increasing flavonoids and soluble sugar contents. Sci. Hortic. 2021, 280, 109903. [Google Scholar] [CrossRef]
- Bechoff, A.; Tomlins, K.; Fliedel, G.; Lopez-Lavalle, L.A.B.; Westby, A.; Hershey, C.; Dufour, D. Cassava traits and end-user preference: Relating traits to consumer liking, sensory perception, and genetics. Crit. Rev. Food Sci. Nutr. 2018, 58, 547–567. [Google Scholar] [CrossRef]
- Saleh, H.; Mohammed, S.; Khamis, F.K.; Taib, H.J. On-farm testing of selected cassava clones. Afr. Crop. Sci. J. 2005, 12, 283–288. [Google Scholar] [CrossRef] [Green Version]
- de Araujo, F.D.B.; Moura, E.F.; Cunha, R.L.; Neto, J.T.D.; Silva, R.D. Chemical root traits differentiate ‘bitter’ and ‘sweet’ cassava accessions from the Amazon. Crop. Breed. Appl. Biotechnol. 2019, 19, 77–85. [Google Scholar] [CrossRef]
- Xu, W.J.; Wei, Y.Y.; Wang, X.X.; Han, P.P.; Chen, Y.; Xu, F.; Shao, X.F. Molecular cloning and expression analysis of hexokinase genes in peach fruit under postharvest disease stress. Postharvest Biol. Tec. 2021, 172, 111377. [Google Scholar] [CrossRef]
- Alves, H.L.S.; Matiolli, C.C.; Soares, R.C.; Almadanim, M.C.; Oliveira, M.M.; Abreu, I.A. Carbon/nitrogen metabolism and stress response networks—calcium-dependent protein kinases as the missing link? J. Exp. Bot. 2021, 72, 4190–4201. [Google Scholar] [CrossRef]
- Favre, N.; Bárcena, A.; Bahima, J.V.; Martínez, G.; Costa, L. Pulses of low intensity light as promising technology to delay postharvest senescence of broccoli. Postharvest Biol. Tec. 2018, 142, 107–114. [Google Scholar] [CrossRef]
- Yan, Z.; Zuo, J.; Zhou, F.; Shi, J.; Xu, D.; Hu, W.; Jiang, A.; Liu, Y.; Wang, Q. Integrated Analysis of Transcriptomic and Metabolomic Data Reveals the Mechanism by Which LED Light Irradiation Extends the Postharvest Quality of Pak-choi (Brassica campestris L. ssp. chinensis (L.) Makino var. communis Tsen et Lee). Biomolecules 2020, 10, 252. [Google Scholar] [CrossRef] [Green Version]
- Büchert, A.M.; Civello, P.M.; Martínez, G.A. Effect of hot air, UV-C, white light and modified atmosphere treatments on expression of chlorophyll degrading genes in postharvest broccoli (Brassica oleracea L.) florets. Sci. Hortic. 2011, 127, 214–219. [Google Scholar] [CrossRef]
- Zhang, Y.; Ma, Y.; Guo, Y.; Chen, Y.; Yang, M.; Fu, R.; Sun, Y. Physiological and iTRAQ-based proteomic analyses for yellowing of postharvest broccoli heads under elevated O2 controlled atmosphere. Sci. Hortic. 2022, 294, 110769. [Google Scholar] [CrossRef]
- Zhou, H.; Yu, Z.; Ye, Z. Key proteins associated to coloured compounds of peach peel using iTRAQ proteomic techniques during development and postharvest. Sci. Hortic. 2018, 239, 123–132. [Google Scholar] [CrossRef]
- Kasampalis, D.S.; Tsouvaltzis, P.; Siomos, A.S. Chlorophyll fluorescence, non-photochemical quenching and light harvesting complex as alternatives to color measurement, in classifying tomato fruit according to their maturity stage at harvest and in monitoring postharvest ripening during storage. Postharvest Biol. Tec. 2020, 161, 111036. [Google Scholar] [CrossRef]
- Flores, H.E.; Dai, Y.; Cuello, J.L.; Maldonado-Mendoza, I.E.; Loyola-Vargas, V.M. Green Roots: Photosynthesis and Photoautotrophy in an Underground Plant Organ. Plant Physiol. 1993, 101, 363–371. [Google Scholar] [CrossRef] [Green Version]
- McClure, P.R.; Coker, G.T.; Schubert, K.R. Carbon Dioxide Fixation in Roots and Nodules of Alnus glutinosa: I. Role of Phosphoenolpyruvate Carboxylase and Carbamyl Phosphate Synthetase in Dark CO(2) Fixation, Citrulline Synthesis, and N(2) Fixation. Plant Physiol. 1983, 71, 652–657. [Google Scholar] [CrossRef] [Green Version]
- Saithong, T.; Rongsirikul, O.; Kalapanulak, S.; Chiewchankaset, P.; Siriwat, W.; Netrphan, S.; Suksangpanomrung, M.; Meechai, A.; Cheevadhanarak, S. Starch biosynthesis in cassava: A genome-based pathway reconstruction and its exploitation in data integration. BMC Syst. Biol. 2013, 7, 75. [Google Scholar] [CrossRef] [Green Version]
- Basha, E.; Jones, C.; Wysocki, V.; Vierling, E. Mechanistic Differences between Two Conserved Classes of Small Heat Shock Proteins Found in the Plant Cytosol. J. Biol. Chem. 2010, 285, 11489–11497. [Google Scholar] [CrossRef] [Green Version]
- Yun, Z.; Gao, H.; Liu, P.; Liu, S.; Luo, T.; Jin, S.; Xu, Q.; Xu, J.; Cheng, Y.; Deng, X. Comparative proteomic and metabolomic profiling of citrus fruit with enhancement of disease resistance by postharvest heat treatment. BMC Plant Biol. 2013, 13, 44. [Google Scholar] [CrossRef] [Green Version]
- Ugolini, L.; Righetti, L.; Carbone, K.; Paris, R.; Malaguti, L.; Di Francesco, A.; Micheli, L.; Paliotta, M.; Mari, M.; Lazzeri, L. Postharvest application of brassica meal-derived allyl-isothiocyanate to kiwifruit: Effect on fruit quality, nutraceutical parameters and physiological response. J. Food Sci. Technol. 2017, 54, 751–760. [Google Scholar] [CrossRef] [Green Version]
- Vanderschuren, H.; Nyaboga, E.; Poon, J.S.; Baerenfaller, K.; Grossmann, J.; Hirsch-Hoffmann, M.; Kirchgessner, N.; Nanni, P.; Gruissem, W. Large-Scale Proteomics of the Cassava Storage Root and Identification of a Target Gene to Reduce Postharvest Deterioration. Plant Cell. 2014, 26, 1913–1924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.; Ji, S.; Wei, B.; Cheng, S.; Wang, Y.; Hao, J.; Wang, S.; Zhou, Q. Transcriptome analysis of postharvest blueberries (Vaccinium corymbosum ‘Duke’) in response to cold stress. BMC Plant Biol. 2020, 20, 80. [Google Scholar] [CrossRef] [Green Version]
- Busatto, N.; Farneti, B.; Tadiello, A.; Oberkofler, V.; Cellini, A.; Biasioli, F.; Delledonne, M.; Cestaro, A.; Noutsos, C.; Costa, F. Wide transcriptional investigation unravel novel insights of the on-tree maturation and postharvest ripening of ‘Abate Fetel’ pear fruit. Hortic. Res. 2019, 6, 32. [Google Scholar] [CrossRef] [PubMed]
- Folsche, V.; Grossmann, C.; Richter, A.S. Impact of Porphyrin Binding to GENOMES UNCOUPLED 4 on Tetrapyrrole Biosynthesis in planta. Front. Plant Sci. 2022, 13, 850504. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Liu, S.; Tian, S.; Ma, T.; Wang, W. Light regulates chlorophyll biosynthesis via ELIP1 during the storage of Chinese cabbage. Sci. Rep. 2022, 12, 11098. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Li, C.; Wang, Y.; Huang, R.; Sun, C.; Xu, Z.; Zhu, J.; Gao, X.; Deng, X.; Wang, P. Identification of a Geranylgeranyl reductase gene for chlorophyll synthesis in rice. Springerplus 2014, 3, 201. [Google Scholar] [CrossRef] [Green Version]
- Hou, Q.; Bartels, D. Comparative study of the aldehyde dehydrogenase (ALDH) gene superfamily in the glycophyte Arabidopsis thaliana and Eutrema halophytes. Ann. Bot. 2015, 115, 465–479. [Google Scholar] [CrossRef] [Green Version]
- Hao, J.; Petriacq, P.; de Bont, L.; Hodges, M.; Gakiere, B. Characterization of l-aspartate oxidase from Arabidopsis thaliana. Plant Sci. 2018, 271, 133–142. [Google Scholar] [CrossRef]
- Bi, F.; Barad, S.; Ment, D.; Luria, N.; Dubey, A.; Casado, V.; Glam, N.; Minguez, J.D.; Espeso, E.A.; Fluhr, R.; et al. Carbon regulation of environmental pH by secreted small molecules that modulate pathogenicity in phytopathogenic fungi. Mol. Plant Pathol. 2016, 17, 1178–1195. [Google Scholar] [CrossRef] [Green Version]
- Tian, J.; Pang, Y.; Zhao, Z. Drought, Salinity, and Low Nitrogen Differentially Affect the Growth and Nitrogen Metabolism of Sophora japonica (L.) in a Semi-Hydroponic Phenotyping Platform. Front. Plant Sci. 2021, 12, 715456. [Google Scholar] [CrossRef]
- Tsilikochrisos, G.; Tsaniklidis, G.; Delis, C.; Nikoloudakis, N.; Aivalakis, G. Glutamate dehydrogenase is differentially regulated in seeded and parthenocarpic tomato fruits during crop development and postharvest storage. Sci. Hortic. 2015, 181, 34–42. [Google Scholar] [CrossRef]
- Pratta, G.; Zorzoli, R.; Boggio, S.B.; Picardi, L.A.; Valle, E.M. Glutamine and glutamate levels and related metabolizing enzymes in tomato fruits with different shelf-life. J. Sci. Horticulturae. 2004, 100, 341–347. [Google Scholar] [CrossRef]
- Ferraro, G.; Bortolotti, S.; Mortera, P.; Schlereth, A.; Stitt, M.; Carrari, F.; Kamenetzky, L.; Valle, E.M. Novel glutamate dehydrogenase genes show increased transcript and protein abundances in mature tomato fruits. J. Plant Physiol. 2012, 169, 899–907. [Google Scholar] [CrossRef] [PubMed]
- Intarasit, S.; Faiyue, B.; Uthaibutra, J.; Saengnil, K. Amelioration of nitrosative stress to reduce pericarp browning of harvested longan by chlorine dioxide. Sci. Hortic. 2022, 293, 110678. [Google Scholar] [CrossRef]
- Sun, Q.; Zhang, N.; Wang, J.; Zhang, H.; Li, D.; Shi, J.; Li, R.; Weeda, S.; Zhao, B.; Ren, S. Melatonin promotes ripening and improves quality of tomato fruit during postharvest life. J. Exp. Botany 2015, 66, 657–668. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Li, R.; Lin, F.; Xiong, Y.; Wang, L.; Wang, B.; Guo, J.; Hu, C. Transcriptome Changes Induced by Different Potassium Levels in Banana Roots. Plants 2019, 9, 11. [Google Scholar] [CrossRef] [Green Version]
- Prochnik, S.; Marri, P.R.; Desany, B.; Rabinowicz, P.D.; Kodira, C.; Mohiuddin, M.; Rodriguez, F.; Fauquet, C.; Tohme, J.; Harkins, T.; et al. The Cassava Genome: Current Progress, Future Directions. Trop. Plant Biol. 2012, 5, 88–94. [Google Scholar] [CrossRef] [Green Version]
- Wesolowski, S.; Birtwistle, M.R.; Rempala, G.A. A Comparison of Methods for RNA-Seq Differential Expression Analysis and a New Empirical Bayes Approach. Biosensors 2013, 3, 238–258. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Chin, C.H.; Chen, S.H.; Wu, H.H.; Ho, C.W.; Ko, M.T.; Lin, C.Y. cytoHubba: Identifying hub objects and sub-networks from complex interactome. BMC Syst. Biol. 2014, 8, S11. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Classify | Pathway ID | Description | Gene Number | False Discovery Rate | |
|---|---|---|---|---|---|
| upregulated | Energy metabolism | map00195 | Photosynthesis | 10 | 0.00024 |
| map00196 | Photosynthesis—antenna proteins | 6 | 0.00036 | ||
| Folding, sorting and degradation | map04141 | Protein processing in endoplasmic reticulum | 29 | 2.08 × 10−9 | |
| Global and overview maps | map01110 | Biosynthesis of secondary metabolites | 51 | 0.0252 | |
| Lipid metabolism | map00062 | Fatty acid elongation | 5 | 0.0418 | |
| map00073 | Cutin, suberine and wax biosynthesis | 7 | 0.0021 | ||
| Signal transduction | map04075 | Plant hormone signal transduction | 18 | 0.0156 | |
| downregulated | Amino acid metabolism | map00250 | Alanine, aspartate and glutamate metabolism | 9 | 0.0058 |
| map00270 | Cysteine and methionine metabolism | 23 | 9.50 × 10−7 | ||
| map00400 | Phenylalanine, tyrosine and tryptophan biosynthesis | 9 | 0.0061 | ||
| Biosynthesis of other secondary metabolites | map00940 | Phenylpropanoid biosynthesis | 42 | 8.91 × 10−14 | |
| map00941 | Flavonoid biosynthesis | 8 | 0.034 | ||
| map00945 | Stilbenoid, diarylheptanoid and gingerol biosynthesis | 8 | 0.0082 | ||
| Carbohydrate metabolism | map00052 | Galactose metabolism | 9 | 0.0153 | |
| map00500 | Starch and sucrose metabolism | 16 | 0.0136 | ||
| map00520 | Amino sugar and nucleotide sugar metabolism | 14 | 0.0315 | ||
| Global and overview maps | map01100 | Metabolic pathways | 211 | 2.41 × 10−21 | |
| map01110 | Biosynthesis of secondary metabolites | 149 | 2.07 × 10−22 | ||
| map01200 | Carbon metabolism | 27 | 0.0041 | ||
| map01230 | Biosynthesis of amino acids | 27 | 0.00043 | ||
| Metabolism of other amino acids | map00450 | Selenocompound metabolism | 5 | 0.0282 | |
| map00460 | Cyanoamino acid metabolism | 9 | 0.0035 | ||
| map00480 | Glutathione metabolism | 24 | 4.65 × 10−9 | ||
| Metabolism of terpenoids and polyketides | map00900 | Terpenoid backbone biosynthesis | 13 | 0.00043 | |
| map00904 | Diterpenoid biosynthesis | 6 | 0.03 | ||
| Signal transduction | map04016 | MAPK signaling pathway—plant | 24 | 2.89 × 10−5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, R.; Yuan, S.; Zhou, Y.; Wang, S.; Zhou, Q.; Ding, Z.; Wang, Y.; Yao, Y.; Liu, J.; Guo, J. Comparative Transcriptome Profiling of Cassava Tuberous Roots in Response to Postharvest Physiological Deterioration. Int. J. Mol. Sci. 2023, 24, 246. https://doi.org/10.3390/ijms24010246
Li R, Yuan S, Zhou Y, Wang S, Zhou Q, Ding Z, Wang Y, Yao Y, Liu J, Guo J. Comparative Transcriptome Profiling of Cassava Tuberous Roots in Response to Postharvest Physiological Deterioration. International Journal of Molecular Sciences. 2023; 24(1):246. https://doi.org/10.3390/ijms24010246
Chicago/Turabian StyleLi, Ruimei, Shuai Yuan, Yangjiao Zhou, Shijia Wang, Qin Zhou, Zhongping Ding, Yajie Wang, Yuan Yao, Jiao Liu, and Jianchun Guo. 2023. "Comparative Transcriptome Profiling of Cassava Tuberous Roots in Response to Postharvest Physiological Deterioration" International Journal of Molecular Sciences 24, no. 1: 246. https://doi.org/10.3390/ijms24010246