
Cannabidiol and Nano-Selenium Increase Microvascularization and Reduce Degenerative Changes in Superficial Breast Muscle in C. perfringens-Infected Chickens
 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Results
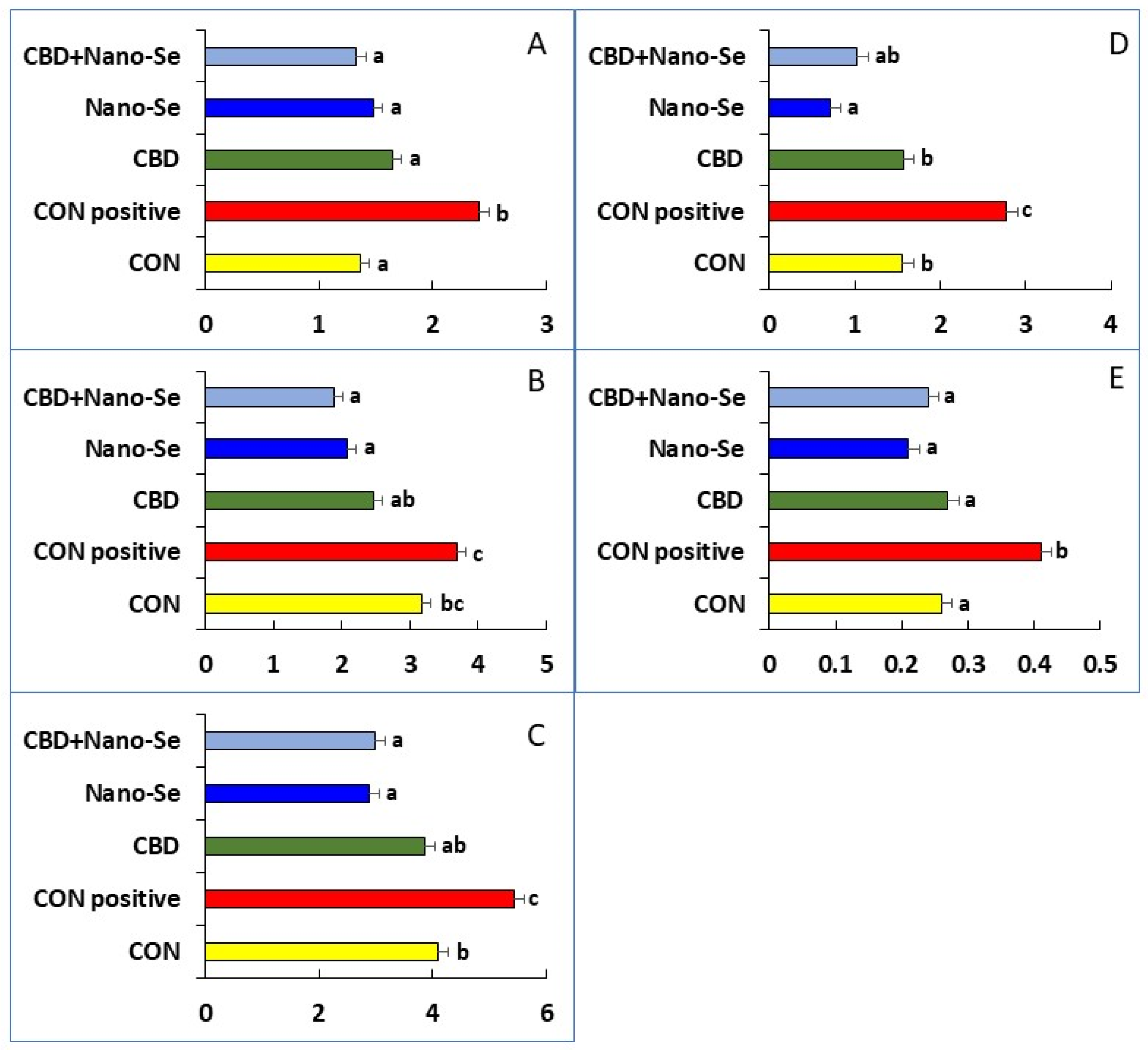
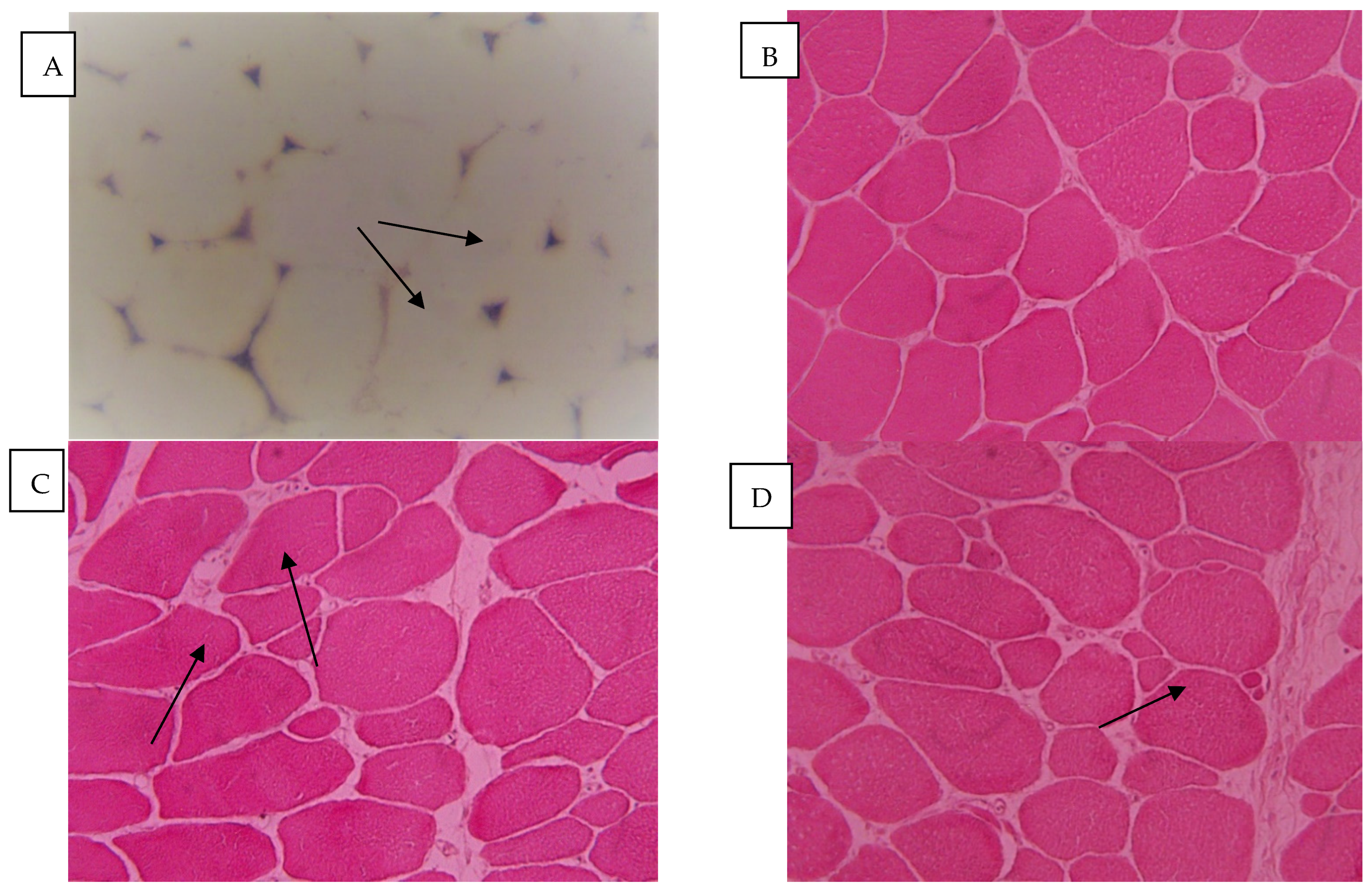
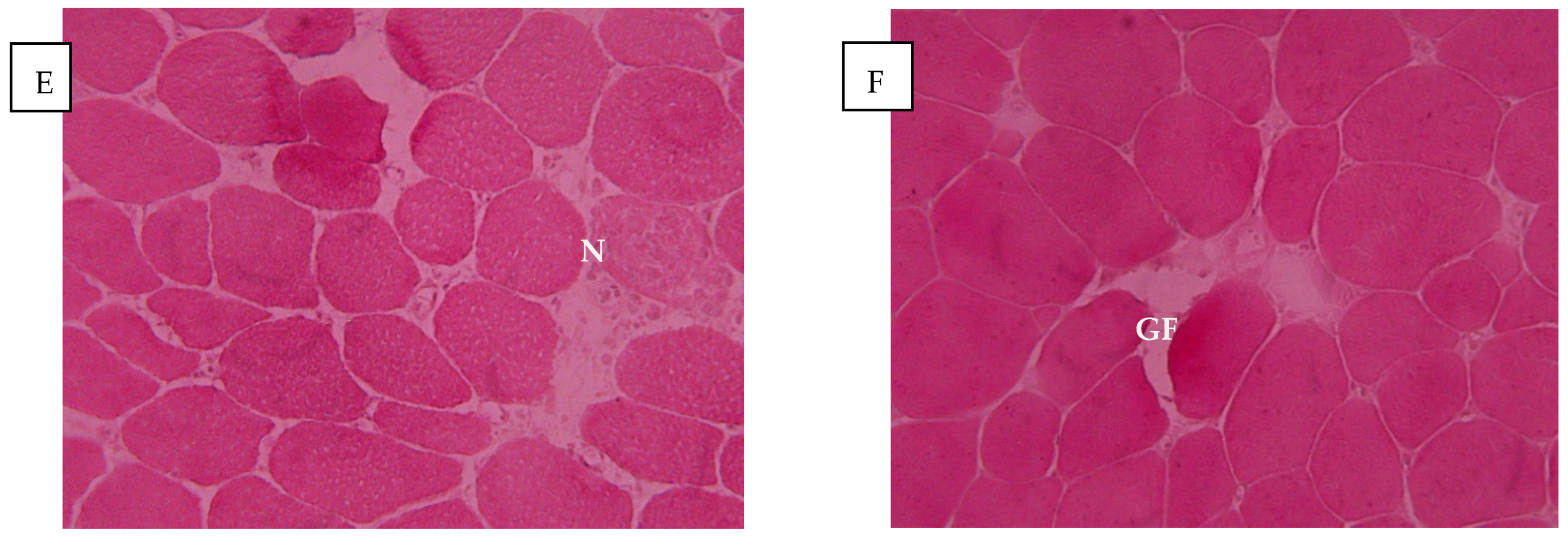
2.1. Breast Meat Microstructure in Response to Treatment
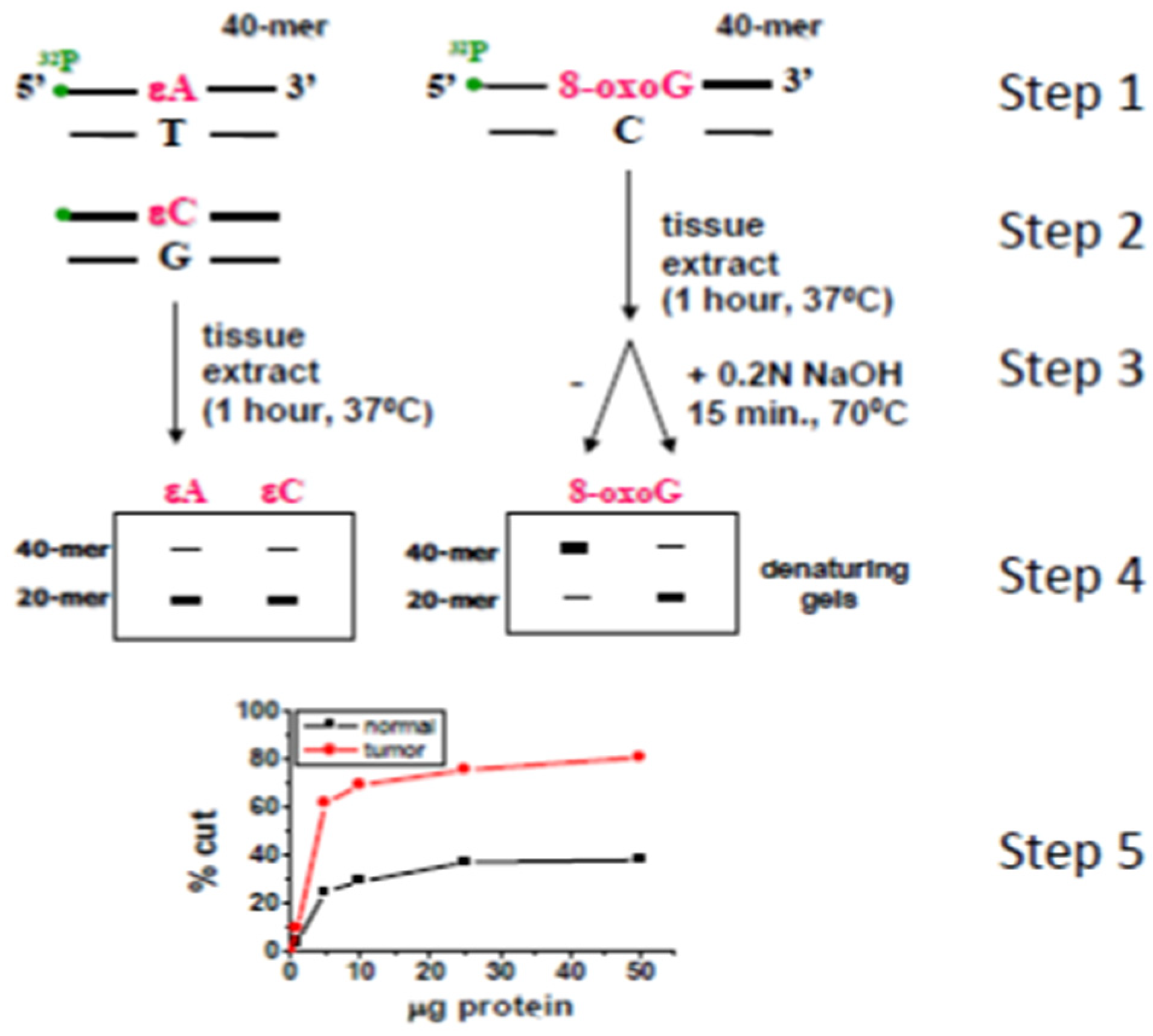
2.2. Gut Barrier Condition as Response to Treatment
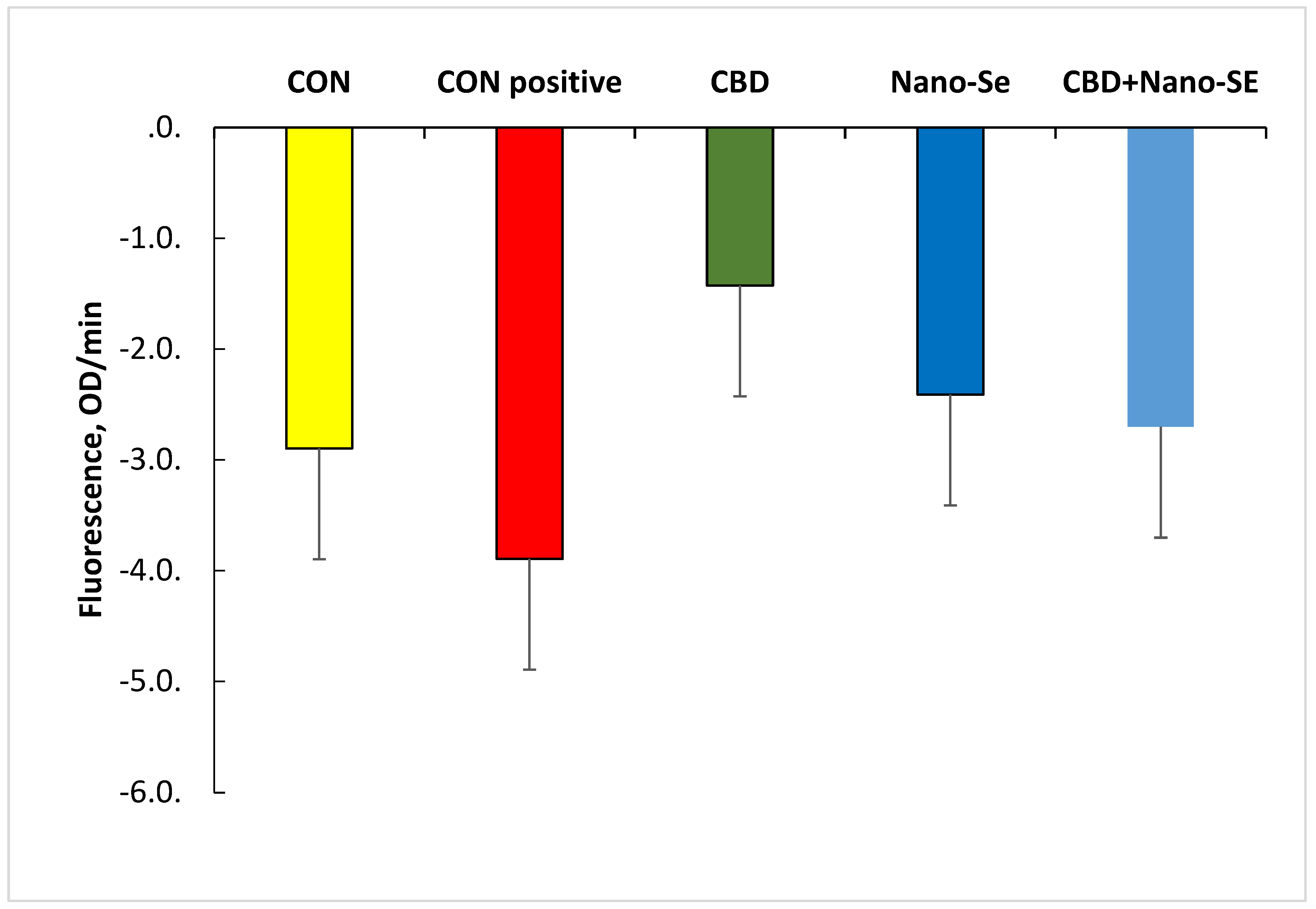
2.3. Oxygen Consumption Rate in Platelet Mitochondria as a Response to Treatments
3. Discussion
4. Materials and Methods
4.1. Chemical Composition of Hemp Extract and Nano-Selenium
4.2. Birds, Treatments and Challenge Model
4.3. Evaluation of the Microstructure of the Breast Muscles of Broiler Chickens
4.4. Assessment of the Activity of DNA Damage Repair Enzymes in the Ileum
4.5. Analysis of Mitochondrial Oxygen Metabolism Using the Oxygen Consumption Assay
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Skinner, J.T.; Bauer, S.; Young, V.; Pauling, G.; Wilson, J. An economic analysis of the impact of subclinical (Mild) necrotic enteritis in broiler chickens. Avian Dis. 2010, 54, 1237–1240. [Google Scholar] [CrossRef] [PubMed]
- Goossens, E.; Dierick, E.; Ducatelle, R.; Van Immerseel, F. Spotlight on avian pathology: Untangling contradictory disease descriptions of necrotic enteritis and necro-haemorrhagic enteritis in broilers. Avian Pathol. 2020, 49, 423–427. [Google Scholar] [CrossRef] [PubMed]
- Van Immerseel, F.; De Buck, J.; Pasmans, F.; Huyghebaert, G.; Haesebrouck, F.; Ducatelle, R. Clostridium perfringens in poultry: An emerging threat for animal and public health. Avian Pathol. 2004, 33, 537–549. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, Y.; Hosotani, M.; Saito, M.; Nagasawa, T.; Mori, Y.; Kawasaki, T.; Yamada, M.; Maeda, N.; Watanabe, T.; Iwasaki, T. Mitochondrial characteristics of chicken breast muscle affected by wooden breast. Comp. Biochem. Physiol.-Part A Mol. Integr. Physiol. 2022, 273, 111296. [Google Scholar] [CrossRef]
- Bogucka, J.; Dankowiakowska, A.; Stanek, M.; Stadnicka, K.; Kirkiłło-Stacewicz, K. Effect of synbiotics administered in ovo on microvascularization and histopathological changes in pectoral muscle and the biochemical profile of broiler chicken blood. Poult. Sci. 2022, 101, 101628. [Google Scholar] [CrossRef]
- Yekhtin, Z.; Khuja, I.; Meiri, D.; Or, R.; Almogi-Hazan, O. Differential Effects of D9 Tetrahydrocannabinol (THC)- and Cannabidiol (CBD)-Based Cannabinoid Treatments on Macrophage Immune Function In Vitro and on Gastrointestinal Inflammation in a Murine Model. Biomedicines 2022, 10, 1793. [Google Scholar] [CrossRef]
- Klein, T.W.; Cabral, G.A. Cannabinoid-induced immune suppression and modulation of antigen-presenting cells. J. Neuroimmune Pharmacol. 2006, 1, 50–64. [Google Scholar] [CrossRef]
- Konieczka, P.; Szkopek, D.; Kinsner, M.; Fotschki, B.; Juśkiewicz, J.; Banach, J. Cannabis-derived cannabidiol and nanoselenium improve gut barrier function and affect bacterial enzyme activity in chickens subjected to C. perfringens challenge. Vet. Res. 2020, 51, 141. [Google Scholar] [CrossRef]
- Varga, Z.V.; Ferdinandy, P.; Liaudet, L.; Pacher, P. Drug-induced mitochondrial dysfunction and cardiotoxicity. Am. J. Physiol.-Heart Circ. Physiol. 2015, 309, H1453–H1467. [Google Scholar] [CrossRef] [Green Version]
- Fraguas-Sánchez, A.I.; Fernández-Carballido, A.; Delie, F.; Cohen, M.; Martin-Sabroso, C.; Mezzanzanica, D.; Figini, M.; Satta, A.; Torres-Suárez, A.I. Enhancing ovarian cancer conventional chemotherapy through the combination with cannabidiol loaded microparticles. Eur. J. Pharm. Biopharm. 2020, 154, 246–258. [Google Scholar] [CrossRef]
- Huestis, M.A.; Solimini, R.; Pichini, S.; Pacifici, R.; Carlier, J.; Busardò, F.P. Cannabidiol Adverse Effects and Toxicity. Curr. Neuropharmacol. 2019, 17, 974–989. [Google Scholar] [CrossRef]
- Chan, J.Z.; Duncan, R.E. Regulatory effects of cannabidiol on mitochondrial functions: A review. Cells 2021, 10, 1251. [Google Scholar] [CrossRef] [PubMed]
- Yuan, D.; Zhan, X.A.; Wang, Y.X. Effect of selenium sources on the expression of cellular glutathione peroxidase and cytoplasmic thioredoxin reductase in the liver and kidney of broiler breeders and their offspring. Poult. Sci. 2012, 91, 936–942. [Google Scholar] [CrossRef]
- Perić, L.; Milošević, N.; Žikić, D.; Kanački, Z.; Džinić, N.; Nollet, L.; Spring, P. Effect of selenium sources on performance and meat characteristics of broiler chickens. J. Appl. Poult. Res. 2009, 18, 403–409. [Google Scholar] [CrossRef]
- Surai, P.F.; Kochish, I.I.; Fisinin, V.I.; Velichko, O.A. Selenium in poultry nutrition: From sodium selenite to organic selenium sources. J. Poult. Sci. 2018, 55, 79–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selim, N.A.; Radwan, N.L.; Youssef, S.F.; Salah Eldin, T.A.; Abo Elwafa, S. Effect of inclusion inorganic, organic or Nano selenium forms in Broiler diets on: 1-Growth Performance, carcass and Meat Characteristics. Int. J. Poult. Sci. 2015, 14, 135–143. [Google Scholar] [CrossRef] [Green Version]
- Michalczuk, M.; Batorska, M.; Sikorska, U.; Bień, D.; Urban, J.; Capecka, K.; Konieczka, P. Selenium and the health status, production results, and product quality in poultry. Anim. Sci. J. 2021, 92, e13662. [Google Scholar] [CrossRef] [PubMed]
- Hosnedlova, B.; Kepinska, M.; Skalickova, S.; Fernandez, C.; Ruttkay-Nedecky, B.; Donald Malevu, T.; Sochor, J.; Baron, M.; Melcova, M.; Zidkova, J.; et al. A summary of new findings on the biological effects of selenium in selected animal species—A critical review. Int. J. Mol. Sci. 2017, 18, 2209. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, S.; Liu, C.; Cai, Y. The protection of selenium on ROS mediated-apoptosis by mitochondria dysfunction in cadmium-induced LLC-PK1 cells. Toxicol. Vitr. 2009, 23, 288–294. [Google Scholar] [CrossRef]
- Taskin, E.; Dursun, N. Recovery of adriamycin induced mitochondrial dysfunction in liver by selenium. Cytotechnology 2015, 67, 977–986. [Google Scholar] [CrossRef]
- Liu, H.J.; Qin, Y.; Zhao, Z.H.; Zhang, Y.; Yang, J.H.; Zhai, D.H.; Cui, F.; Luo, C.; Lu, M.X.; Liu, P.P.; et al. Lentinan-functionalized selenium nanoparticles target tumor cell mitochondria via TLR4/TRAF3/MFN1 pathway. Theranostics 2020, 10, 9083–9099. [Google Scholar] [CrossRef] [PubMed]
- Hosnedlova, B.; Kepinska, M.; Skalickova, S.; Fernandez, C.; Ruttkay-Nedecky, B.; Peng, Q.; Baron, M.; Melcova, M.; Opatrilova, R.; Zidkova, J.; et al. Nano-selenium and its nanomedicine applications: A critical review. Int. J. Nanomed. 2018, 13, 2107–2128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kogut, M.H.; Yin, X.; Yuan, J.; Broom, L. Gut health in poultry. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2017, 12, 1–7. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, J.; Zhong, H.; Li, N.; Xu, H.; Zhu, Q.; Liu, Y. Effect of probiotics on the meat flavour and gut microbiota of chicken. Sci. Rep. 2017, 7, 6400. [Google Scholar] [CrossRef] [Green Version]
- Castro, F.L.D.S.; Kim, W.K. Secondary functions of arginine and sulfur amino acids in poultry health: Review. Animals 2020, 10, 2106. [Google Scholar] [CrossRef]
- Bortoluzzi, C.; Rochell, S.J.; Applegate, T.J. Threonine, arginine, and glutamine: Influences on intestinal physiology, immunology, and microbiology in broilers. Poult. Sci. 2018, 97, 937–945. [Google Scholar] [CrossRef]
- Celi, P.; Cowieson, A.J.; Fru-Nji, F.; Steinert, R.E.; Kluenter, A.M.; Verlhac, V. Gastrointestinal functionality in animal nutrition and health: New opportunities for sustainable animal production. Anim. Feed Sci. Technol. 2017, 234, 88–100. [Google Scholar] [CrossRef]
- Fallahi, S.; Bobak, Ł.; Opaliński, S. Hemp in Animal Diets—Cannabidiol. Animals 2022, 12, 2541. [Google Scholar] [CrossRef]
- Wang, Q.; Zhan, X.; Wang, B.; Wang, F.; Zhou, Y.; Xu, S.; Li, X.; Tang, L.; Jin, Q.; Li, W.; et al. Modified Montmorillonite Improved Growth Performance of Broilers by Modulating Intestinal Microbiota and Enhancing Intestinal Barriers, Anti-Inflammatory Response, and Antioxidative Capacity. Antioxidants 2022, 11, 1799. [Google Scholar] [CrossRef]
- Lu, M.; Yuan, B.; Yan, X.; Sun, Z.; Lillehoj, H.S.; Lee, Y.; Baldwin-Bott, C.; Li, C. Clostridium perfringens-induced host-pathogen transcriptional changes in the small intestine of broiler chickens. Pathogens 2021, 10, 1607. [Google Scholar] [CrossRef]
- Konieczka, P.; Żelechowska, E.; Przybylski, W.; Jaworska, D.; Sałek, P.; Kinsner, M.; Jankowski, J. The sarcoplasmic protein profile of breast muscle in Turkeys in response to different dietary ratios of limiting amino acids and Clostridium perfringens-induced inflammation. Poult. Sci. 2022, 101, 102195. [Google Scholar] [CrossRef] [PubMed]
- Cai, K.; Shao, W.; Chen, X.; Campbell, Y.L.; Nair, M.N.; Suman, S.P.; Beach, C.M.; Guyton, M.C.; Schilling, M.W. Meat quality traits and proteome profile of woody broiler breast (pectoralis major) meat. Poult. Sci. 2018, 97, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Sihvo, H.K.; Airas, N.; Lindén, J.; Puolanne, E. Pectoral Vessel Density and Early Ultrastructural Changes in Broiler Chicken Wooden Breast Myopathy. J. Comp. Pathol. 2018, 161, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Gornatti-Churria, C.D.; Crispo, M.; Shivaprasad, H.L.; Uzal, F.A. Gangrenous dermatitis in chickens and turkeys. J. Vet. Diagn. Investig. 2018, 30, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Zúñiga-Pereira, A.M.; Santamaría, C.; Gutierrez, J.M.; Alape-Girón, A.; Flores-Díaz, M. Deficient skeletal muscle regeneration after injury induced by a clostridium perfringens strain associated with gas gangrene. Infect. Immun. 2019, 87, e00200-19. [Google Scholar] [CrossRef] [Green Version]
- Konieczka, P.; Wojtasik-Kalinowska, I.; Poltorak, A.; Kinsner, M.; Szkopek, D.; Fotschki, B.; Juśkiewicz, J.; Banach, J.; Michalczuk, M. Cannabidiol affects breast meat volatile compounds in chickens subjected to different infection models. Sci. Rep. 2022, 12, 18940. [Google Scholar] [CrossRef]
- Zhou, M.; Zeng, D.; Ni, X.; Tu, T.; Yin, Z.; Pan, K.; Jing, B. Effects of bacillus licheniformis on the growth performance and expression of lipid metabolism-related genes in broiler chickens challenged with clostridium perfringens-induced necrotic enteritis. Lipids Health Dis. 2016, 15, 48. [Google Scholar] [CrossRef] [Green Version]
- Bień, D.; Michalczuk, M.; Szkopek, D.; Kinsner, M.; Konieczka, P. Changes in lipids metabolism indices as a result of different form of selenium supplementation in chickens. Sci. Rep. 2022, 12, 13817. [Google Scholar] [CrossRef]
- Ullah Khan, R.; Durrani, F.-E.-R.; Chand, N.; Anwar, H.; Naz, S.; Farooqi, A.; Manzoor, M.N. Effect of cannabis sativa fortified feed on muscle growth and visceral organs in broiler chicks. Int. J. Biol. Biotechnol. 2009, 6, 179–182. [Google Scholar]
- Skřivan, M.; Englmaierová, M.; Taubner, T.; Skřivanová, E. Effects of dietary hemp seed and flaxseed on growth performance, meat fatty acid compositions, liver tocopherol concentration and bone strength of cockerels. Animals 2020, 10, 458. [Google Scholar] [CrossRef] [Green Version]
- Šťastník, O.; Jůzl, M.; Karásek, F.; Fernandová, D.; Mrkvicová, E.; Pavlata, L.; Nedomová, Š.; Vyhnánek, T.; Trojan, V.; Doležal, P. The effect of hempseed expellers on selected quality indicators of broiler chicken’s meat. Acta Vet. Brno 2019, 88, 121–128. [Google Scholar] [CrossRef]
- Cai, S.J.; Wu, C.X.; Gong, L.M.; Song, T.; Wu, H.; Zhang, L.Y. Effects of nano-selenium on performance, meat quality, immune function, oxidation resistance, and tissue selenium content in broilers. Poult. Sci. 2012, 91, 2532–2539. [Google Scholar] [CrossRef] [PubMed]
- Li, J.L.; Zhang, L.; Yang, Z.Y.; Zhang, Z.Y.; Jiang, Y.; Gao, F.; Zhou, G.H. Effects of Different Selenium Sources on Growth Performance, Antioxidant Capacity and Meat Quality of Local Chinese Subei Chickens. Biol. Trace Elem. Res. 2018, 181, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Langer, H.T.; Mossakowski, A.A.; Pathak, S.; Mascal, M.; Baar, K. Cannabidiol does not impair anabolic signaling following eccentric contractions in rats. Int. J. Sport Nutr. Exerc. Metab. 2021, 31, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Bogucka, J.; Ribeiro, D.M.; Bogusławska-Tryk, M.; Dankowiakowska, A.; da Costa, R.P.R.; Bednarczyk, M. Microstructure of the small intestine in broiler chickens fed a diet with probiotic or synbiotic supplementation. J. Anim. Physiol. Anim. Nutr. 2019, 103, 1785–1791. [Google Scholar] [CrossRef] [PubMed]
- Dankowiakowska, A.; Bogucka, J.; Sobolewska, A.; Tavaniello, S.; Maiorano, G.; Bednarczyk, M. Effects of in ovo injection of prebiotics and synbiotics on the productive performance and microstructural features of the superficial pectoral muscle in broiler chickens. Poult. Sci. 2019, 98, 5157–5165. [Google Scholar] [CrossRef]
- Maiorano, G.; Stadnicka, K.; Tavaniello, S.; Abiuso, C.; Bogucka, J.; Bednarczyk, M. In ovo validation model to assess the efficacy of commercial prebiotics on broiler performance and oxidative stability of meat. Poult. Sci. 2017, 96, 511–518. [Google Scholar] [CrossRef]
- Pelyhe, C.; Mézes, M. Myths and Facts About the Effects of Nano- Selenium in Farm Animals—Mini-Review. Eur. Chem. Bull. 2013, 2, 1049–1052. [Google Scholar]
- Bilgili, S.F.; Hess, J.B. Green Muscle Disease in broilers increasing. World Poult. 2002, 18, 42–43. [Google Scholar]
- Velleman, S.G. Relationship of Skeletal Muscle Development and Growth to Breast Muscle Myopathies: A Review. Avian Dis. 2015, 59, 525–531. [Google Scholar] [CrossRef]
- Kaszyńska, A. Possibilities of therapeutic action of cannabinoids in neurodegenerative diseases. Pharmacother. Psychiatry Neurol. 2022, 38, 49–58. [Google Scholar] [CrossRef]
- MacRae, V.E.; Mahon, M.; Gilpin, S.; Sandercock, D.A.; Mitchell, M.A. Skeletal muscle fibre growth and growth associated myopathy in the domestic chicken (Gallus domesticus). Br. Poult. Sci. 2006, 47, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Olmon, E.D.; Delaney, S. Differential Ability of Five DNA Glycosylases to Recognize and Repair Damage on Nucleosomal DNA. ACS Chem. Biol. 2017, 12, 692–701. [Google Scholar] [CrossRef] [PubMed]
- Winczura, A.; Czubaty, A.; Winczura, K.; Masłowska, K.; Nałecz, M.; Dudzińska, D.A.; Saparbaev, M.; Staroń, K.; Tudek, B. Lipid peroxidation product 4-hydroxy-2-nonenal modulates base excision repair in human cells. DNA Repair 2014, 22, 1–11. [Google Scholar] [CrossRef]
- Kowalczyk, P.; Jaworek, J.; Kot, M.; Sokolowska, B.; Bielen, A.; Janowska, B.; Ciesla, J.M.; Szparecki, G.; Sados, B.; Tudek, B. Inflammation increases oxidative DNA damage repair and stimulates preneoplastic changes in colons of newborn rats. J. Physiol. Pharmacol. 2016, 67, 277–286. [Google Scholar]
- Soglia, F.; Petracci, M.; Davoli, R.; Zappaterra, M. A critical review of the mechanisms involved in the occurrence of growth-related abnormalities affecting broiler chicken breast muscles. Poult. Sci. 2021, 100, 101180. [Google Scholar] [CrossRef]
- Pampouille, E.; Hennequet-Antier, C.; Praud, C.; Juanchich, A.; Brionne, A.; Godet, E.; Bordeau, T.; Fagnoul, F.; Le Bihan-Duval, E.; Berri, C. Differential expression and co-expression gene network analyses reveal molecular mechanisms and candidate biomarkers involved in breast muscle myopathies in chicken. Sci. Rep. 2019, 9, 14905. [Google Scholar] [CrossRef] [Green Version]
- Abasht, B.; Mutryn, M.F.; Michalek, R.D.; Lee, W.R. Oxidative Stress and Metabolic Perturbations in Wooden Breast Disorder in Chickens. PLoS ONE 2016, 11, e0153750. [Google Scholar] [CrossRef] [Green Version]
- Gratta, F.; Fasolato, L.; Birolo, M.; Zomeño, C.; Novelli, E.; Petracci, M.; Pascual, A.; Xiccato, G.; Trocino, A. Effect of breast myopathies on quality and microbial shelf life of broiler meat. Poult. Sci. 2019, 98, 2641–2651. [Google Scholar] [CrossRef]
- Marchesi, J.A.P.; Ibelli, A.M.G.; Peixoto, J.O.; Cantão, M.E.; Pandolfi, J.R.C.; Marciano, C.M.M.; Zanella, R.; Settles, M.L.; Coutinho, L.L.; Ledur, M.C. Whole transcriptome analysis of the pectoralis major muscle reveals molecular mechanisms involved with white striping in broiler chickens. Poult. Sci. 2019, 98, 590–601. [Google Scholar] [CrossRef]
- Hynes, J.; Nadanaciva, S.; Swiss, R.; Carey, C.; Kirwan, S.; Will, Y. A high-throughput dual parameter assay for assessing drug-induced mitochondrial dysfunction provides additional predictivity over two established mitochondrial toxicity assays. Toxicol. Vitr. 2013, 27, 560–569. [Google Scholar] [CrossRef] [PubMed]
- De Paula Martins, R.; Glaser, V.; Da Luz Scheffer, D.; De Paula Ferreira, P.M.; Wannmacher, C.M.D.; Farina, M.; De Oliveira, P.A.; Prediger, R.D.; Latini, A. Platelet oxygen consumption as a peripheral blood marker of brain energetics in a mouse model of severe neurotoxicity. J. Bioenerg. Biomembr. 2013, 45, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Siewiera, K.; Kassassir, H.; Talar, M.; Wieteska, L.; Watala, C. Higher mitochondrial potential and elevated mitochondrial respiration are associated with excessive activation of blood platelets in diabetic rats. Life Sci. 2016, 148, 293–304. [Google Scholar] [CrossRef] [PubMed]
- Vuda, M.; Kamath, A. Drug induced mitochondrial dysfunction: Mechanisms and adverse clinical consequences. Mitochondrion 2016, 31, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Konieczka, P.; Barszcz, M.; Kowalczyk, P.; Szlis, M.; Jankowski, J. The potential of acetylsalicylic acid and vitamin e in modulating inflammatory cascades in chickens under lipopolysaccharide-induced inflammation. Vet. Res. 2019, 50, 65. [Google Scholar] [CrossRef] [Green Version]
- Ferdous, F.; Scott, T.R. A comparative examination of thrombocyte/platelet immunity. Immunol. Lett. 2015, 163, 32–39. [Google Scholar] [CrossRef]
- Wigley, P. Immunity to bacterial infection in the chicken. Dev. Comp. Immunol. 2013, 41, 413–417. [Google Scholar] [CrossRef]
- Aviagen Ross 308: Broiler Performance Objectives; Aviagen Inc.: Huntsville, AL, USA, 2019; pp. 1–15.
- Konieczka, P.; Sandvang, D.; Kinsner, M.; Szkopek, D.; Szyryńska, N.; Jankowski, J. Bacillus-based probiotics affect gut barrier integrity in different ways in chickens subjected to optimal or challenge conditions. Vet. Microbiol. 2022, 265, 109323. [Google Scholar] [CrossRef]
- Van Damme, L.; Cox, N.; Callens, C.; Haesebrouck, F.; Dargatz, M.; Ducatelle, R.; Van Immerseel, F.; Goossens, E.C. perfringens challenge reduces matrix metalloproteinase activity in the jejunal mucosa of Eimeria-infected broiler chickens. Vet. Res. 2020, 51, 100. [Google Scholar] [CrossRef]
- Olkowski, A.A.; Wojnarowicz, C.; Chirino-Trejo, M.; Drew, M.D. Responses of broiler chickens orally challenged with Clostridium perfringens isolated from field cases of necrotic enteritis. Res. Vet. Sci. 2006, 81, 99–108. [Google Scholar] [CrossRef]
- Timbermont, L.; Haesebrouck, F.; Ducatelle, R.; Van Immerseel, F. Necrotic enteritis in broilers: An updated review on the pathogenesis. Avian Pathol. 2011, 40, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Obtułowicz, T.; Winczura, A.; Speina, E.; Swoboda, M.; Janik, J.; Janowska, B.; Cieśla, J.M.; Kowalczyk, P.; Jawien, A.; Gackowski, D.; et al. Aberrant repair of etheno-DNA adducts in leukocytes and colon tissue of colon cancer patients. Free Radic. Biol. Med. 2010, 49, 1064–1071. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Group | ||||
|---|---|---|---|---|---|
| CON | CON Positive | CBD | Nano-Se | CBD + Nano-Se | |
| Number of capillaries per muscle fiber | 0.97 b ± 0.07 | 0.98 b ± 0.05 | 1.20 ab ± 0.10 | 1.11 ab ± 0.09 | 1.31 a ± 0.12 |
| Number of muscle fibers/1.5 mm2 | 165.38 ± 5.89 | 177.75 ± 12.70 | 165.13 ± 10.07 | 172.88 ± 7.82 | 178.13 ± 12.67 |
| Muscle fiber diameter (μm) | 48.56 ± 2.99 | 44.95 ± 5.62 | 47.59 ± 5.60 | 48.29 ± 3.10 | 45.66 ± 6.09 |
| Number of necrotic fibers/1.5 mm2 | 1.63 ab ± 0.32 | 2.38 a ± 0.53 | 1.50 ab ± 0.60 | 0.38 b ± 0.26 | 1.00 b ± 0.38 |
| Number of atrophic fibers/1.5 mm2 | 3.13 a ± 0.79 | 2.50 ab ± 0.53 | 1.13 b ± 0.30 | 2.25 ab ± 0.60 | 2.50 ab ± 0.42 |
| Number of split fibers /1.5 mm2 | 2.00 b ± 0.65 | 4.13 a ± 0.55 | 1.88 b ± 0.40 | 1.63 b ± 0.60 | 1.13 b ± 0.40 |
| Number of giant fibers/1.5 mm2 | 0.50 ± 0.27 | 0.63 ± 0.32 | 0.63 ± 0.32 | 0.75 ± 0.25 | 0.25 ± 0.16 |
| Treatment | CON | CON Positive | CBD | Nano-Se | CBD+Nano-Se |
|---|---|---|---|---|---|
| Commercial diet for broiler Ross 308 with no additives | + | + | + | + | + |
| C. perfringens Challenge | − | + | + | + | + |
| Cannabidiol | − | − | + | − | + |
| Nanosized selenium | − | − | − | + | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Konieczka, P.; Szkopek, D.; Kinsner, M.; Kowalczyk, P.; Michalczuk, M.; Bień, D.; Banach, J.; Matusevičius, P.; Bogucka, J. Cannabidiol and Nano-Selenium Increase Microvascularization and Reduce Degenerative Changes in Superficial Breast Muscle in C. perfringens-Infected Chickens. Int. J. Mol. Sci. 2023, 24, 237. https://doi.org/10.3390/ijms24010237
Konieczka P, Szkopek D, Kinsner M, Kowalczyk P, Michalczuk M, Bień D, Banach J, Matusevičius P, Bogucka J. Cannabidiol and Nano-Selenium Increase Microvascularization and Reduce Degenerative Changes in Superficial Breast Muscle in C. perfringens-Infected Chickens. International Journal of Molecular Sciences. 2023; 24(1):237. https://doi.org/10.3390/ijms24010237
Chicago/Turabian StyleKonieczka, Paweł, Dominika Szkopek, Misza Kinsner, Paweł Kowalczyk, Monika Michalczuk, Damian Bień, Joanna Banach, Paulius Matusevičius, and Joanna Bogucka. 2023. "Cannabidiol and Nano-Selenium Increase Microvascularization and Reduce Degenerative Changes in Superficial Breast Muscle in C. perfringens-Infected Chickens" International Journal of Molecular Sciences 24, no. 1: 237. https://doi.org/10.3390/ijms24010237