

Iron Dyshomeostasis in COVID-19: Biomarkers Reveal a Functional Link to 5-Lipoxygenase Activation
 , , , , ,
, , , , ,  , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
2.1. Clinical Characteristics of COVID-19 and Long-COVID Patients
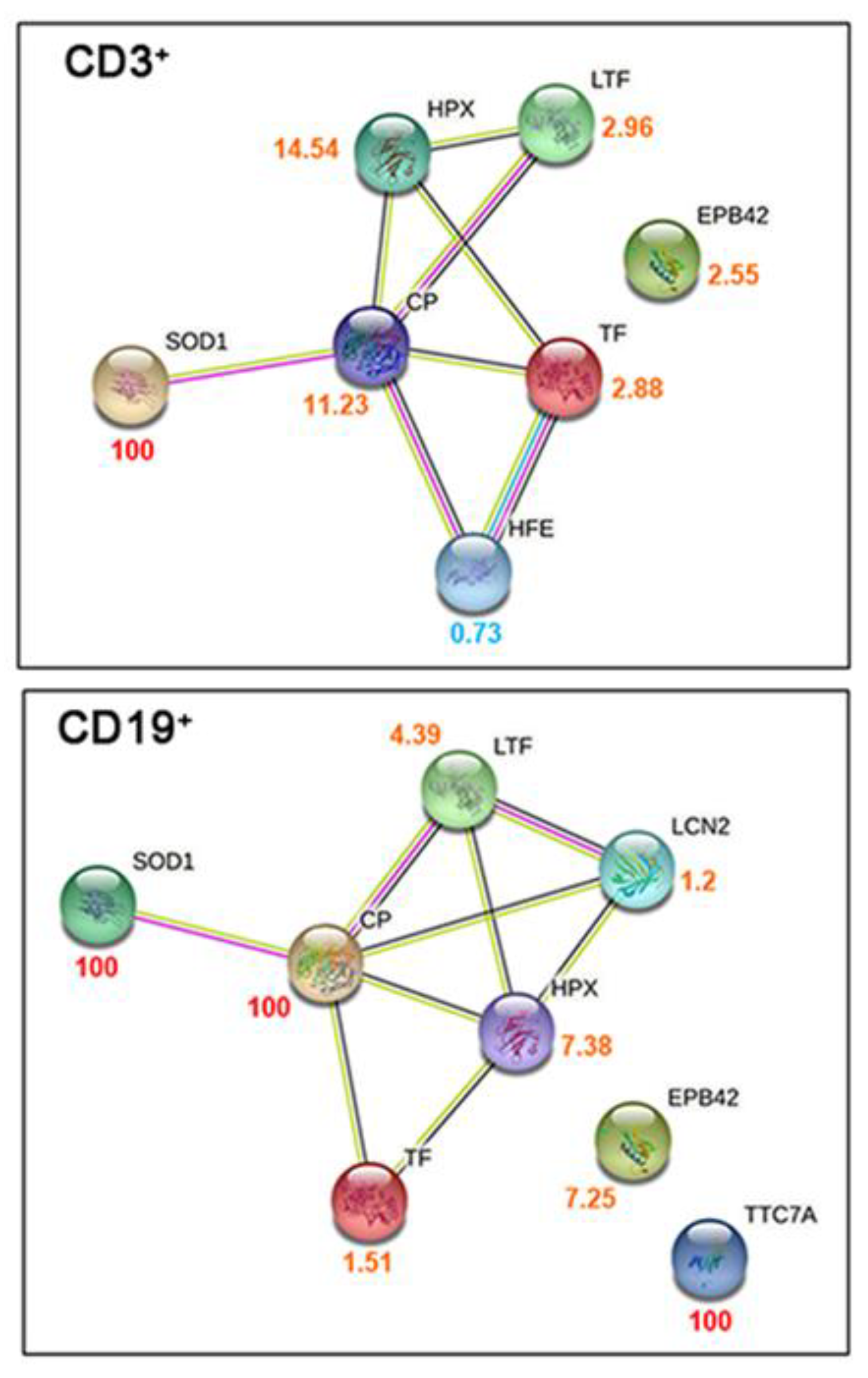
2.2. Proteomic and Metabolomic Changes in COVID-19 Lymphocytes
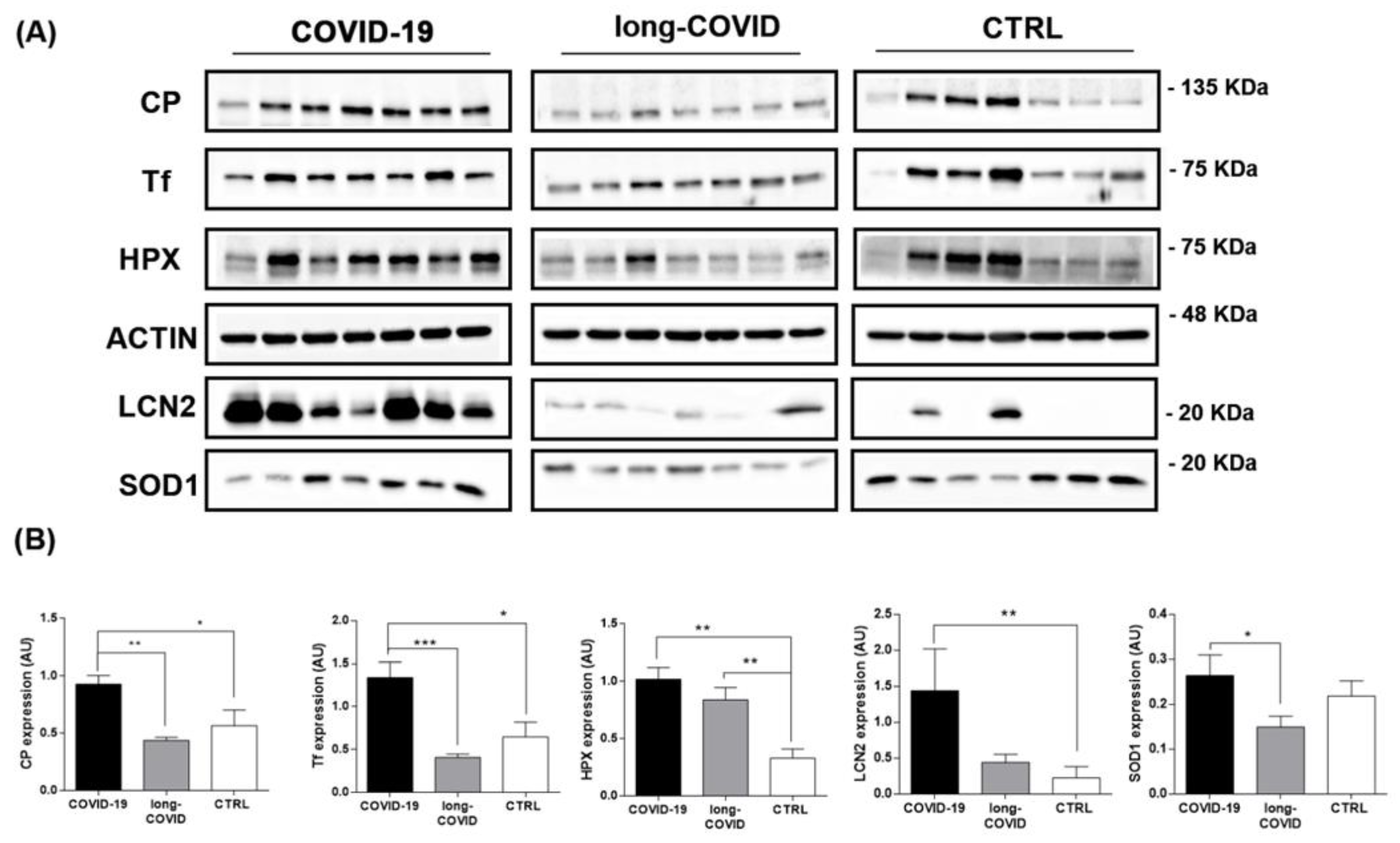
2.3. Iron-Related Biomarker Proteins Are Dysregulated in COVID-19 and Long-COVID Patients
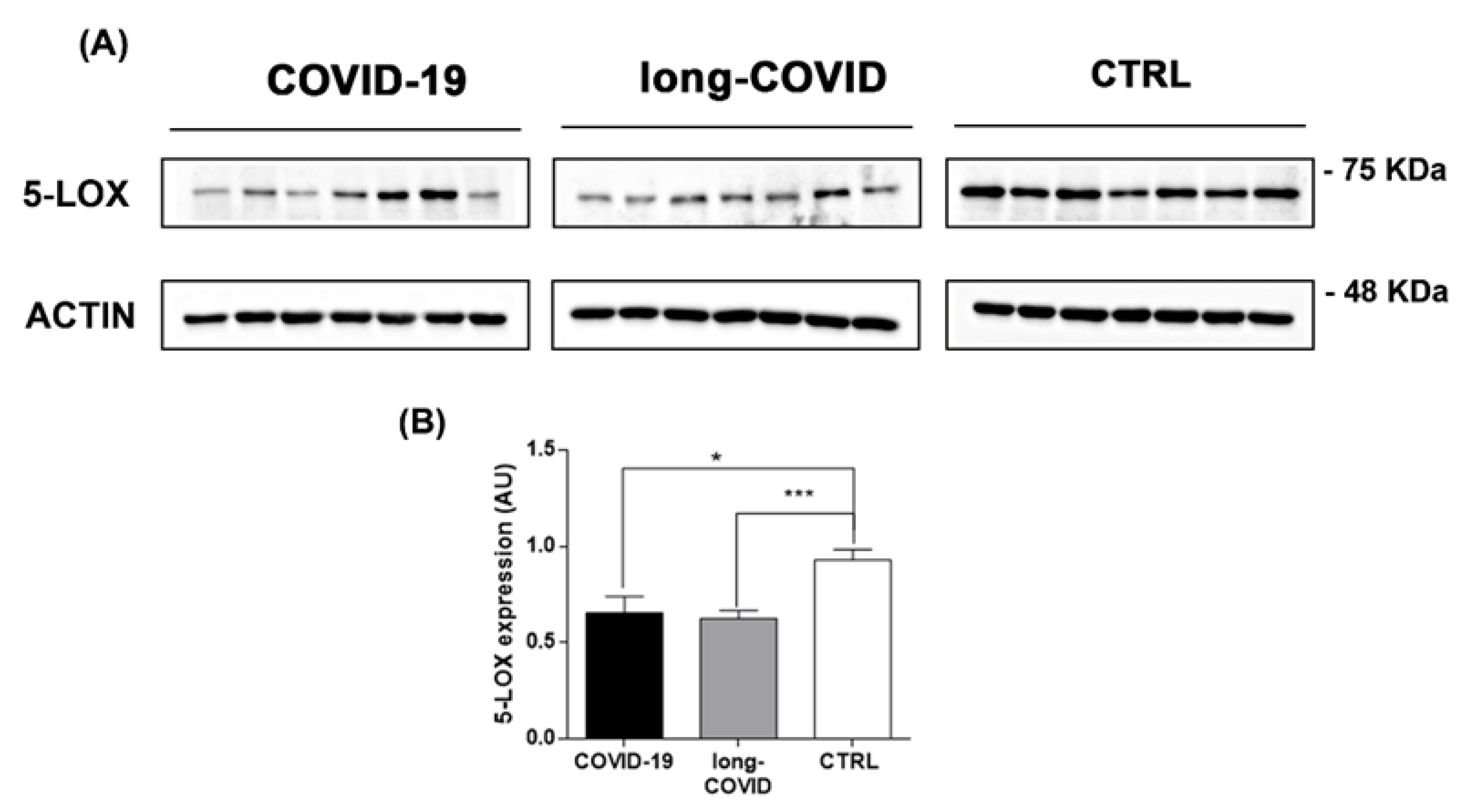
2.4. 5-LOX Expression Is Modulated in COVID-19 and Long-COVID Patients
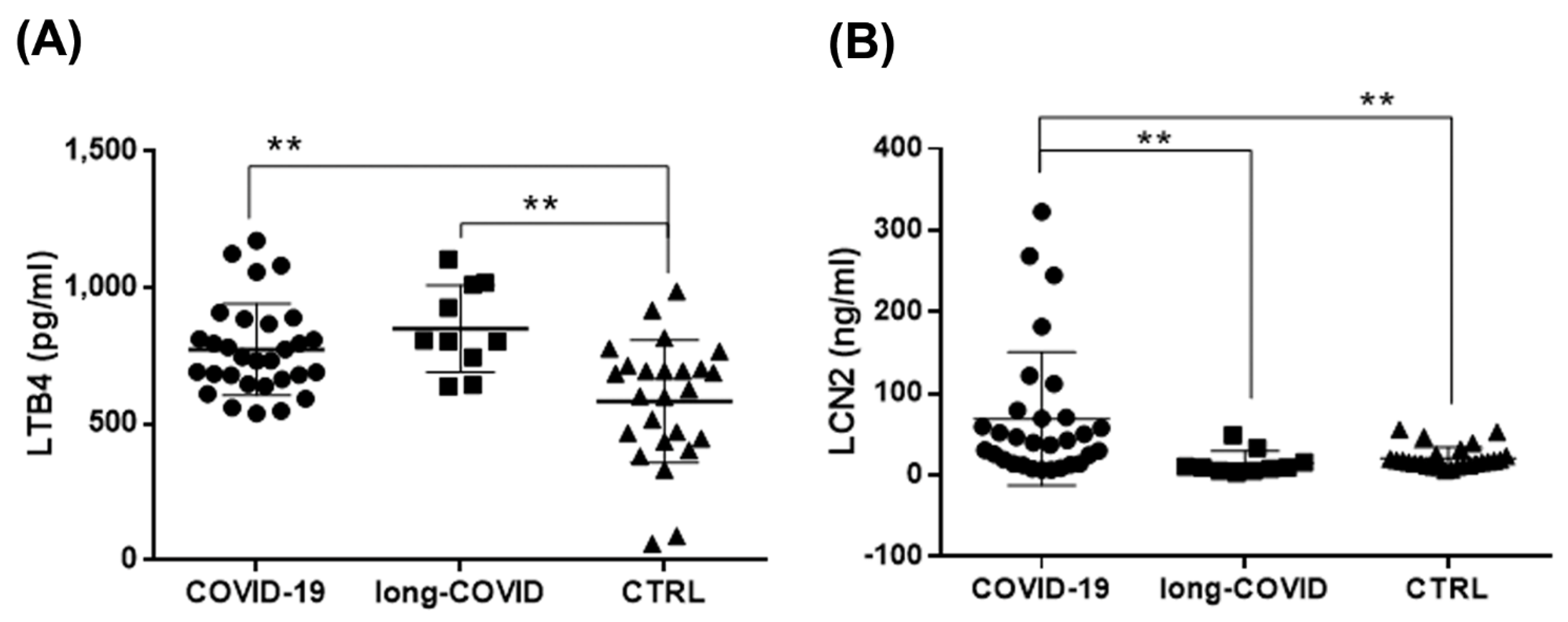
2.5. Increased LTB4 and LCN2 Plasma Levels in COVID-19 and Long-COVID-19 Patients
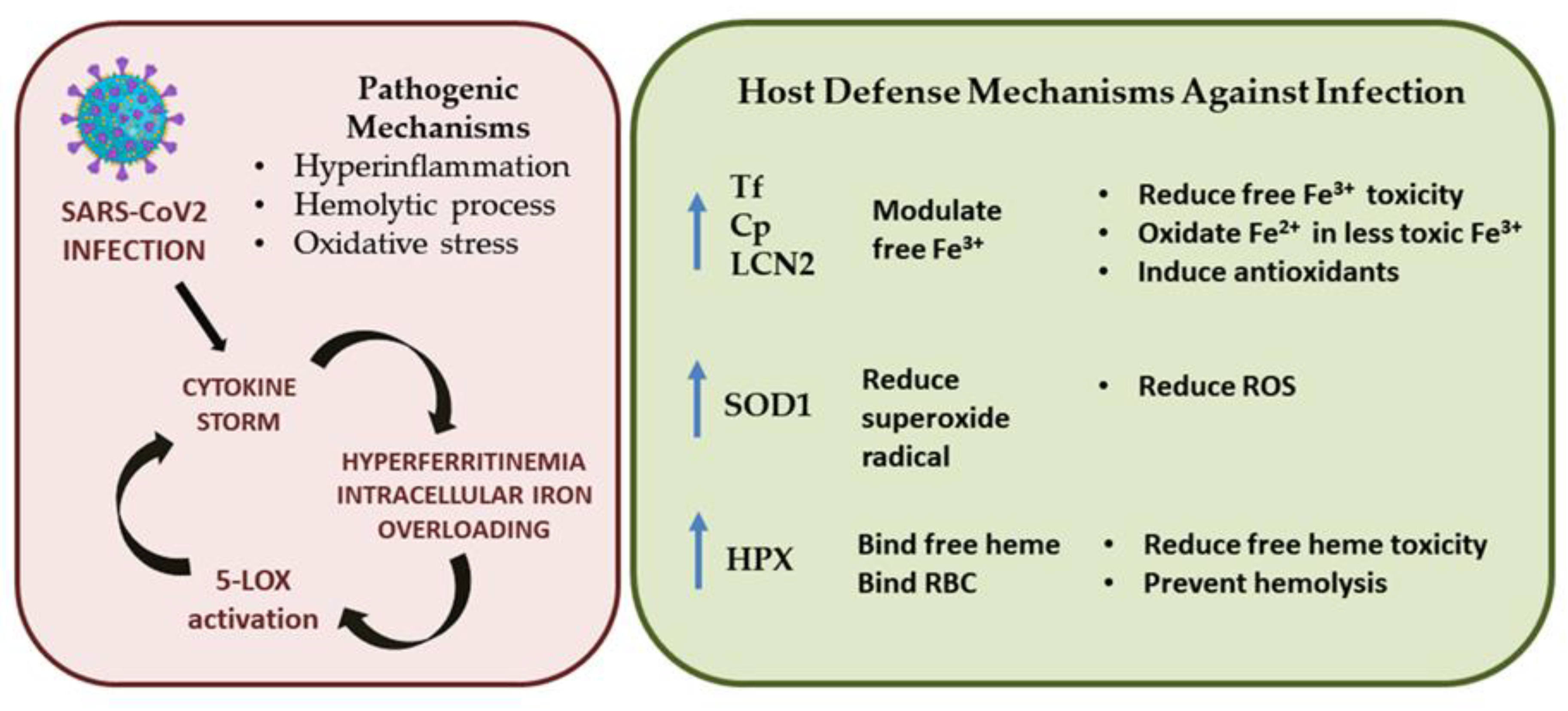
3. Discussion
4. Materials and Methods
4.1. Patients and Sample Information
4.2. Peripheral Immune Cell (PBMC) Isolation by Fluorescence-Activated Cell Sorting
4.3. Proteomics Analysis and Data Processing
4.4. Western Blot Analysis
4.5. Quantitative Real-Time Reverse Transcription–Polymerase Chain Reaction (qRT-PCR) Analysis
4.6. Enzyme-Linked Immunosorbent Assays
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organisation (WHO). COVID-19 Weekly Epidemiological Update; WHO: Geneva, Switzerland, 2022; pp. 1–26. [Google Scholar]
- Caramaschi, S.; Kapp, M.E.; Miller, S.E.; Eisenberg, R.; Johnson, J.; Epperly, G.; Maiorana, A.; Silvestri, G.; Giannico, G.A. Histopathological Findings and Clinicopathologic Correlation in COVID-19: A Systematic Review. Mod. Pathol. 2021, 34, 1614–1633. [Google Scholar] [CrossRef] [PubMed]
- Di Carlo, P.; Falasca, K.; Ucciferri, C.; Sinjari, B.; Aruffo, E.; Antonucci, I.; Di Serafino, A.; Pompilio, A.; Damiani, V.; Mandatori, D.; et al. Normal Breathing Releases SARS-CoV-2 into the Air. J. Med. Microbiol. 2021, 70, 001328. [Google Scholar] [CrossRef] [PubMed]
- Meyerowitz, E.A.; Richterman, A.; Gandhi, R.T.; Sax, P.E. Transmission of SARS-CoV-2: A Review of Viral, Host, and Environmental Factors. Ann. Intern. Med. 2021, 174, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical Features of Patients Infected with 2019 Novel Coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gandhi, R.T.; Lynch, J.B.; del Rio, C. Mild or Moderate COVID-19. N. Engl. J. Med. 2020, 383, 1757–1766. [Google Scholar] [CrossRef]
- Meng, X.; Deng, Y.; Dai, Z.; Meng, Z. COVID-19 and Anosmia: A Review Based on up-to-Date Knowledge. Am. J. Otolaryngol. 2020, 41, 102581. [Google Scholar] [CrossRef]
- Vaira, L.A.; Salzano, G.; Deiana, G.; De Riu, G. Anosmia and Ageusia: Common Findings in COVID-19 Patients. Laryngoscope 2020, 130, 1787. [Google Scholar] [CrossRef] [Green Version]
- Chen, N.; Zhou, M.; Dong, X.; Qu, J.; Gong, F.; Han, Y.; Qiu, Y.; Wang, J.; Liu, Y.; Wei, Y.; et al. Epidemiological and Clinical Characteristics of 99 Cases of 2019 Novel Coronavirus Pneumonia in Wuhan, China: A Descriptive Study. Lancet 2020, 395, 507–513. [Google Scholar] [CrossRef] [Green Version]
- Mo, P.; Xing, Y.; Xiao, Y.; Deng, L.; Zhao, Q.; Wang, H.; Xiong, Y.; Cheng, Z.; Gao, S.; Liang, K.; et al. Clinical Characteristics of Refractory COVID-19 Pneumonia in Wuhan, China. Clin. Infect. Dis. 2020, 73, e4208–e4213. [Google Scholar] [CrossRef] [Green Version]
- Meyer, N.J.; Gattinoni, L.; Calfee, C.S. Acute Respiratory Distress Syndrome. Lancet 2021, 398, 622–637. [Google Scholar] [CrossRef] [PubMed]
- Gorman, E.A.; O’Kane, C.M.; McAuley, D.F. Acute Respiratory Distress Syndrome in Adults: Diagnosis, Outcomes, Long-Term Sequelae, and Management. Lancet 2022, 400, 1157–1170. [Google Scholar] [CrossRef]
- Zhou, F.; Yu, T.; Du, R.; Fan, G.; Liu, Y.; Liu, Z.; Xiang, J.; Wang, Y.; Song, B.; Gu, X.; et al. Clinical Course and Risk Factors for Mortality of Adult Inpatients with COVID-19 in Wuhan, China: A Retrospective Cohort Study. Lancet 2020, 395, 1054–1062. [Google Scholar] [CrossRef] [PubMed]
- Deer, R.R.; Rock, M.A.; Vasilevsky, N.; Carmody, L.; Rando, H.; Anzalone, A.J.; Basson, M.D.; Bennett, T.D.; Bergquist, T.; Boudreau, E.A.; et al. Characterizing Long COVID: Deep Phenotype of a Complex Condition. EBioMedicine 2021, 74, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Nirantharakumar, K.; Hughes, S.; Myles, P.; Williams, T.; Gokhale, K.M.; Taverner, T.; Chandan, J.S.; Brown, K.; Simms-Williams, N.; et al. Symptoms and Risk Factors for Long COVID in Non-Hospitalized Adults. Nat. Med. 2022, 28, 1706–1714. [Google Scholar] [CrossRef] [PubMed]
- Shah, W.; Hillman, T.; Playford, E.D.; Hishmeh, L. Managing the Long Term Effects of COVID-19: Summary of NICE, SIGN, and RCGP Rapid Guideline. BMJ 2021, 372, 10–13. [Google Scholar] [CrossRef]
- Yong, S.J. Long COVID or Post-COVID-19 Syndrome: Putative Pathophysiology, Risk Factors, and Treatments. Infect. Dis. 2021, 53, 737–754. [Google Scholar] [CrossRef]
- Miyazato, Y.; Morioka, S.; Tsuzuki, S.; Akashi, M.; Osanai, Y.; Tanaka, K.; Terada, M.; Suzuki, M.; Kutsuna, S.; Saito, S.; et al. Prolonged and Late-Onset Symptoms of Coronavirus Disease 2019. Open Forum Infect. Dis. 2020, 7, 4–6. [Google Scholar] [CrossRef]
- Huang, C.; Huang, L.; Wang, Y.; Li, X.; Ren, L.; Gu, X.; Kang, L.; Guo, L.; Liu, M.; Zhou, X.; et al. 6-Month Consequences of COVID-19 in Patients Discharged from Hospital: A Cohort Study. Lancet 2021, 397, 220–232. [Google Scholar] [CrossRef]
- Greenhalgh, T.; Knight, M.; A’Court, C.; Buxton, M.; Husain, L. Management of Post-Acute COVID-19 in Primary Care. BMJ 2020, 370, m3026. [Google Scholar] [CrossRef]
- Davis, H.E.; Assaf, G.S.; McCorkell, L.; Wei, H.; Low, R.J.; Re’em, Y.; Redfield, S.; Austin, J.P.; Akrami, A. Characterizing Long COVID in an International Cohort: 7 Months of Symptoms and Their Impact. eClinicalMedicine 2021, 38, 2–19. [Google Scholar] [CrossRef]
- Crook, H.; Raza, S.; Nowell, J.; Young, M.; Edison, P. Long COVID—Mechanisms, Risk Factors, and Management. BMJ 2021, 374, n1648. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Zhou, L.; Hu, Z.; Zhang, S.; Yang, S.; Tao, Y.; Xie, C.; Ma, K.; Shang, K.; Wang, W.; et al. Dysregulation of Immune Response in Patients with Coronavirus 2019 (COVID-19) in Wuhan, China. Clin. Infect. Dis. 2020, 71, 762–768. [Google Scholar] [CrossRef] [PubMed]
- Diao, B.; Wang, C.; Tan, Y.; Chen, X.; Liu, Y.; Ning, L.; Chen, L.; Li, M.; Liu, Y.; Wang, G.; et al. Reduction and Functional Exhaustion of T Cells in Patients with Coronavirus Disease 2019 (COVID-19). Front. Immunol. 2020, 11, 827. [Google Scholar] [CrossRef]
- Mohiuddin Chowdhury, A.T.M.; Karim, M.R.; Ali, M.A.; Islam, J.; Li, Y.; He, S. Clinical Characteristics and the Long-Term Post-Recovery Manifestations of the COVID-19 Patients—A Prospective Multicenter Cross-Sectional Study. Front. Med. 2021, 8, 66367. [Google Scholar] [CrossRef] [PubMed]
- Hasan Ali, O.; Bomze, D.; Risch, L.; Brugger, S.D.; Paprotny, M.; Weber, M.; Thiel, S.; Kern, L.; Albrich, W.C.; Kohler, P.; et al. Severe Coronavirus Disease 2019 (COVID-19) Is Associated with Elevated Serum Immunoglobulin (Ig) A and Antiphospholipid IgA Antibodies. Clin. Infect. Dis. 2021, 73, E2869–E2874. [Google Scholar] [CrossRef] [PubMed]
- Heidepriem, J.; Dahlke, C.; Kobbe, R.; Santer, R.; Koch, T.; Fathi, A.; Seco, B.M.S.; Ly, M.L.; Schmiedel, S.; Schwinge, D.; et al. Longitudinal Development of Antibody Responses in COVID-19 Patients of Different Severity with Elisa, Peptide, and Glycan Arrays: An Immunological Case Series. Pathogens 2021, 10, 438. [Google Scholar] [CrossRef]
- Varghese, J.; Sandmann, S.; Ochs, K.; Schrempf, I.M.; Frömmel, C.; Dugas, M.; Schmidt, H.H.; Vollenberg, R.; Tepasse, P.R. Persistent Symptoms and Lab Abnormalities in Patients Who Recovered from COVID-19. Sci. Rep. 2021, 11, 12775. [Google Scholar] [CrossRef]
- Klein, J.; Wood, J.; Jaycox, J.; Lu, P.; Dhodapkar, R.M.; Gehlhausen, J.R.; Tabachnikova, A.; Tabacof, L.; Malik, A.A.; Kamath, K.; et al. Distinguishing Features of Long COVID Identified through Immune Profiling. medRxiv 2022, 1–13. [Google Scholar] [CrossRef]
- Amiri-Dashatan, N.; Koushki, M.; Parsamanesh, N.; Chiti, H. Serum Cortisol Concentration and COVID-19 Severity: A Systematic Review and Meta-Analysis. J. Investig. Med. 2022, 70, 766–772. [Google Scholar] [CrossRef]
- Ramezani, M.; Simani, L.; Karimialavijeh, E.; Rezaei, O.; Hajiesmaeili, M.; Pakdaman, H. Anxiety and Cortisol in Outcomes of Patients With COVID-19. Basic Clin. Neurosci. J. 2020, 11, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Crowe, W.E.; Maglova, L.M.; Ponka, P.; Russell, J.M. Human Cytomegalovirus-Induced Host Cell Enlargement Is Iron Dependent. Am. J. Physiol. Cell Physiol. 2004, 287, 1023–1030. [Google Scholar] [CrossRef] [PubMed]
- Drakesmith, H.; Prentice, A. Viral Infection and Iron Metabolism. Nat. Rev. Microbiol. 2008, 6, 541–552. [Google Scholar] [CrossRef] [PubMed]
- Ganz, T. Systemic Iron Homeostasis. Physiol. Rev. 2013, 93, 1721–1741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogdan, A.R.; Miyazawa, M.; Hashimoto, K.; Tsuji, Y. Regulators of Iron Homeostasis: New Players in Metabolism, Cell Death, and Disease. Trends Biochem. Sci. 2016, 41, 274–286. [Google Scholar] [CrossRef] [Green Version]
- Des, R.; Richardson, P.P. The Molecular Mechanisms of Iron and Ferritin Metabolism in Normal and Neoplastic Cells. Philosophy 1996, 1331, 1–40. [Google Scholar] [CrossRef]
- Ward, D.M.; Kaplan, J. Ferroportin-Mediated Iron Transport: Expression and Regulation. Biochim. Biophys. Acta-Mol. Cell Res. 2012, 1823, 1426–1433. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Ren, Z.; Gao, S.; Shen, J.; Wang, L.; Xu, Z.; Yu, Y.; Bachina, P.; Zhang, H.; Fan, X.; et al. Structural Basis of Ion Transport and Inhibition in Ferroportin. Nat. Commun. 2020, 11, 5686. [Google Scholar] [CrossRef]
- Camaschella, C.; Nai, A.; Silvestri, L. Iron Metabolism and Iron Disorders Revisited in the Hepcidin Era. Haematologica 2020, 105, 260–272. [Google Scholar] [CrossRef] [Green Version]
- Winter, W.E.; Bazydlo, L.A.L.; Harris, N.S. The Molecular Biology of Human Iron Metabolism. Lab. Med. 2014, 45, 92–102. [Google Scholar] [CrossRef] [Green Version]
- Drakesmith, H.; Prentice, A.M. Hepcidin and the Iron-Infection Axis. Science 2012, 338, 768–772. [Google Scholar] [CrossRef] [PubMed]
- Pacht, E.R.; Davis, W.B. Role of Transferrin and Ceruloplasmin in Antioxidant Activity of Lung Epithelial Lining Fluid. J. Appl. Physiol. 1988, 64, 2092–2099. [Google Scholar] [CrossRef] [PubMed]
- Ponka, P.; Lok, C.N. The Transferrin Receptor: Role in Health and Disease. Int. J. Biochem. Cell Biol. 1999, 31, 1111–1137. [Google Scholar] [CrossRef] [PubMed]
- Daniels, T.R.; Delgado, T.; Rodriguez, J.A.; Helguera, G.; Penichet, M.L. The Transferrin Receptor Part I: Biology and Targeting with Cytotoxic Antibodies for the Treatment of Cancer. Clin. Immunol. 2006, 121, 144–158. [Google Scholar] [CrossRef]
- Xiao, X.; Yeoh, B.S.; Vijay-Kumar, M. Lipocalin 2: An Emerging Player in Iron Homeostasis and Inflammation. Annu. Rev. Nutr. 2017, 37, 103–130. [Google Scholar] [CrossRef]
- Devireddy, L.R.; Gazin, C.; Zhu, X.; Green, M.R. A Cell-Surface Receptor for Lipocalin 24p3 Selectively Mediates Apoptosis and Iron Uptake. Cell 2005, 123, 1293–1305. [Google Scholar] [CrossRef] [Green Version]
- Holmes, M.A.; Paulsene, W.; Jide, X.; Ratledge, C.; Strong, R.K. Siderocalin (Lcn 2) Also Binds Carboxymycobactins, Potentially Defending against Mycobacterial Infections through Iron Sequestration. Structure 2005, 13, 29–41. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.; McCulloh, R.J. Hemopexin and Haptoglobin: Allies against Heme Toxicity from Hemoglobin Not Contenders. Front. Physiol. 2015, 6, 187. [Google Scholar] [CrossRef]
- Eskew, J.D.; Vanacore, R.M.; Sung, L.M.; Morales, P.J.; Smith, A. Cellular Protection Mechanisms against Extracellular Heme: Heme- Hemopexin, but Not Free Heme, Activates the N-Terminal c-Jun Kinase. J. Biol. Chem. 1999, 274, 638–648. [Google Scholar] [CrossRef] [Green Version]
- Yamada, Y.; Miyamoto, T.; Kashima, H.; Kobara, H.; Asaka, R.; Ando, H.; Higuchi, S.; Ida, K.; Shiozawa, T. Lipocalin 2 Attenuates Iron-Related Oxidative Stress and Prolongs the Survival of Ovarian Clear Cell Carcinoma Cells by up-Regulating the CD44 Variant. Free Radic. Res. 2016, 50, 414–425. [Google Scholar] [CrossRef] [Green Version]
- Ighodaro, O.M.; Akinloye, O.A. First Line Defence Antioxidants-Superoxide Dismutase (SOD), Catalase (CAT) and Glutathione Peroxidase (GPX): Their Fundamental Role in the Entire Antioxidant Defence Grid. Alexandria J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef]
- Schmidt, P.J.; Toran, P.T.; Giannetti, A.M.; Bjorkman, P.J.; Andrews, N.C. The Transferrin Receptor Modulates Hfe-Dependent Regulation of Hepcidin Expression. Cell Metab. 2008, 7, 205–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thursz, M. Iron, Haemochromatosis and Thalassaemia as Risk Factors for Fibrosis in Hepatitis C Virus Infection. Gut 2007, 56, 613–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, M.L.; Hu, J.H.; Yen, C.H.; Chen, K.H.; Kuo, C.J.; Lin, M.S.; Lee, C.H.; Chen, S.C.; Chien, R.N. Evolution of Ferritin Levels in Hepatitis C Patients Treated with Antivirals. Sci. Rep. 2020, 10, 19744. [Google Scholar] [CrossRef] [PubMed]
- Haider, B.A.; Spiegelman, D.; Hertzmark, E.; Sando, D.; Duggan, C.; Makubi, A.; Sudfeld, C.; Aris, E.; Chalamilla, G.E.; Fawzi, W.W. Anemia, Iron Deficiency, and Iron Supplementation in Relation to Mortality among HIV-Infected Patients Receiving Highly Active Antiretroviral Therapy in Tanzania. Am. J. Trop. Med. Hyg. 2019, 100, 1512–1520. [Google Scholar] [CrossRef] [PubMed]
- Costagliola, D.G.; De Montalembert, M.; Lefrere, J.J.; Briand, C.; Rebulla, P.; Baruchel, S.; Dessi, C.; Fondu, P.; Karagiorga, M.; Perrimond, H. Dose of Desferrioxamine and Evolution of HIV-1 Infection in Thalassaemic Patients. Br. J. Haematol. 1994, 87, 849–852. [Google Scholar] [CrossRef]
- Georgiou, N.A.; Van Der Bruggen, T.; Oudshoorn, M.; Nottet, H.S.L.M.; Marx, J.J.M.; van Asbeck, B.S. Inhibition of Human Immunodeficiency Virus Type 1 Replication in Human Mononuclear Blood Cells by the Iron Chelators Deferoxamine, Deferiprone, and Bleomycin. J. Infect. Dis. 2000, 181, 484–490. [Google Scholar] [CrossRef]
- Gwamaka, M.; Kurtis, J.D.; Sorensen, B.E.; Holte, S.; Morrison, R.; Mutabingwa, T.K.; Fried, M.; Duffy, P.E. Iron Deficiency Protects against Severe Plasmodium Falciparum Malaria and Death in Young Children. Clin. Infect. Dis. 2012, 54, 1137–1144. [Google Scholar] [CrossRef] [Green Version]
- Spottiswoode, N.; Duffy, P.E.; Drakesmith, H. Iron, Anemia and Hepcidin in Malaria. Front. Pharmacol. 2014, 5, 125. [Google Scholar] [CrossRef] [Green Version]
- Cassat, J.E.; Skaar, E.P. Iron in Infection and Immunity. Cell Host Microbe 2013, 13, 509–519. [Google Scholar] [CrossRef] [Green Version]
- Dufrusine, B.; Di Francesco, A.; Oddi, S.; Scipioni, L.; Angelucci, C.B.; D’Addario, C.; Serafini, M.; Häfner, A.K.; Steinhilber, D.; Maccarrone, M.; et al. Iron-Dependent Trafficking of 5-Lipoxygenase and Impact on Human Macrophage Activation. Front. Immunol. 2019, 10, 01347. [Google Scholar] [CrossRef] [PubMed]
- Torti, S.V.; Torti, F.M. Iron and Cancer: More Ore to Be Mined. Nat. Rev. Cancer 2013, 13, 342–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Yu, L.; Ding, J.; Chen, Y. Iron Metabolism in Cancer. Int. J. Mol. Sci. 2019, 20, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahroum, N.; Alghory, A.; Kiyak, Z.; Alwani, A.; Seida, R. Ferritin—from Iron, through Inflammation and Autoimmunity, to COVID-19. J. Autoimmun. 2020, 126, 102778. [Google Scholar] [CrossRef]
- Weiss, G.; Ganz, T.; Goodnough, L.T. Anemia of in Fl Ammation. Blood 2019, 133, 40–50. [Google Scholar] [CrossRef] [Green Version]
- Nikseresht, S.; Bush, A.I.; Ayton, S. Treating Alzheimer’s Disease by Targeting Iron. Br. J. Pharmacol. 2019, 176, 3622–3635. [Google Scholar] [CrossRef] [PubMed]
- Gong, N.J.; Dibb, R.; Bulk, M.; van der Weerd, L.; Liu, C. Imaging Beta Amyloid Aggregation and Iron Accumulation in Alzheimer’s Disease Using Quantitative Susceptibility Mapping MRI. Neuroimage 2019, 191, 176–185. [Google Scholar] [CrossRef]
- Lai, C.C.; Wang, Y.H.; Chen, K.H.; Chen, C.H.; Wang, C.Y. The Clinical Efficacy and Safety of Anti-Viral Agents for Non-Hospitalized Patients with COVID-19: A Systematic Review and Network Meta-Analysis of Randomized Controlled Trials. Viruses 2022, 14, 1706. [Google Scholar] [CrossRef]
- Ucciferri, C.; Barone, M.; Vecchiet, J.; Falasca, K. Pidotimod in Paucisymptomatic SARS-CoV2 Infected Patients. Mediterr. J. Hematol. Infect. Dis. 2020, 12, 11–13. [Google Scholar] [CrossRef]
- D’ardes, D.; Pontolillo, M.; Esposito, L.; Masciarelli, M.; Boccatonda, A.; Rossi, I.; Bucci, M.; Guagnano, M.T.; Ucciferri, C.; Santilli, F.; et al. Duration of COVID-19: Data from an Italian Cohort and Potential Role for Steroids. Microorganisms 2020, 8, 1327. [Google Scholar] [CrossRef]
- Ucciferri, C.; Vecchiet, J.; Falsca, K. Role of monoclonal antibody drugs in the treatment of COVID-19. World J. Clin. Cases 2020, 8, 4280–4285. [Google Scholar] [CrossRef] [PubMed]
- World Health Organisation. A Clinical Case Definition of Post COVID-19 Condition by a Delphi Consensus. 2021. Available online: https://WHO/2019-nCoV/Post_COVID-19_condition/Clinical_case_definition/2021.1 (accessed on 23 December 2021).
- Rushton, D.H.; Barth, J.H. What Is the Evidence for Gender Differences in Ferritin and Haemoglobin? Crit. Rev. Oncol. Hematol. 2010, 73, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Pasini, E.; Corsetti, G.; Romano, C.; Scarabelli, T.M.; Chen-Scarabelli, C.; Saravolatz, L.; Dioguardi, F.S. Serum Metabolic Profile in Patients with Long-COVID (PASC) Syndrome: Clinical Implications. Front. Med. 2021, 8, 714426. [Google Scholar] [CrossRef] [PubMed]
- Faul, F.; Erdfelder, E.; Lang, A.-G.; Buchner, A. G*Power 3: A Flexible Statistical Power Analysis Program for the Social, Behavioral, and Biomedical Sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Zecca, L.; Youdim, M.B.H.; Riederer, P.; Connor, J.R.; Crichton, R.R. Iron, Brain Ageing and Neurodegenerative Disorders. Nat. Rev. Neurosci. 2004, 5, 863–873. [Google Scholar] [CrossRef]
- Ali, M.K.; Kim, R.Y.; Brown, A.C.; Donovan, C.; Vanka, K.S.; Mayall, J.R.; Liu, G.; Pillar, A.L.; Jones-Freeman, B.; Xenaki, D.; et al. Critical Role for Iron Accumulation in the Pathogenesis of Fibrotic Lung Disease. J. Pathol. 2020, 251, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhang, H.; Chang, J.; Li, D.; Cao, P. Iron Overload and Mitochondrial Dysfunction Orchestrate Pulmonary Fibrosis. Eur. J. Pharmacol. 2021, 912, 174613. [Google Scholar] [CrossRef]
- Sun, Y.; Chen, P.; Zhai, B.; Zhang, M.; Xiang, Y.; Fang, J.; Xu, S.; Gao, Y.; Chen, X.; Sui, X.; et al. The Emerging Role of Ferroptosis in Inflammation. Biomed. Pharmacother. 2020, 127, 110108. [Google Scholar] [CrossRef]
- Regenboog, M.; van Kuilenburg, A.B.P.; Verheij, J.; Swinkels, D.W.; Hollak, C.E.M. Hyperferritinemia and Iron Metabolism in Gaucher Disease: Potential Pathophysiological Implications. Blood Rev. 2016, 30, 431–437. [Google Scholar] [CrossRef]
- Camaschella, C.; Poggiali, E. Towards Explaining “Unexplained Hyperferrltlnemla”. Haematologica 2009, 94, 307–309. [Google Scholar] [CrossRef] [Green Version]
- Schulert, G.S.; Canna, S.W. Convergent Pathways of the Hyperferritinemic Syndromes. Int. Immunol. 2018, 30, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Rosário, C.; Zandman-Goddard, G.; Meyron-Holtz, E.G.; D’Cruz, D.P.; Shoenfeld, Y. The Hyperferritinemic Syndrome: Macrophage Activation Syndrome, Still’s Disease, Septic Shock and Catastrophic Antiphospholipid Syndrome. BMC Med. 2013, 11, 185–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, W.; Knovich, M.A.; Coffman, L.; Torti, F.; Torti, S.V. Serum Ferritin: Past, Present and Future Wei. Bone 2011, 23, 760–769. [Google Scholar] [CrossRef]
- Lalueza, A.; Ayuso, B.; Arrieta, E.; Trujillo, H.; Folgueira, D.; Cueto, C.; Serrano, A.; Laureiro, J.; Arévalo-Cañas, C.; Castillo, C.; et al. Elevation of Serum Ferritin Levels for Predicting a Poor Outcome in Hospitalized Patients with Influenza Infection. Clin. Microbiol. Infect. 2020, 26, 1557.e9–1557.e15. [Google Scholar] [CrossRef] [PubMed]
- Barut, S.; Dincer, F.; Sahin, I.; Ozyurt, H.; Akkus, M.; Erkorkmaz, U. Increased Serum Ferritin Levels in Patients with Crimean-Congo Hemorrhagic Fever: Can It Be a New Severity Criterion? Int. J. Infect. Dis. 2010, 14, 50–54. [Google Scholar] [CrossRef] [Green Version]
- Mehta, P.; McAuley, D.F.; Brown, M.; Sanchez, E.; Tattersall, R.S.; Manson, J.J. COVID-19: Consider Cytokine Storm Syndromes and Immunosuppression. Lancet 2020, 395, 1033–1034. [Google Scholar] [CrossRef]
- Colafrancesco, S.; Alessandri, C.; Conti, F.; Priori, R. COVID-19 Gone Bad: A New Character in the Spectrum of the Hyperferritinemic Syndrome? Autoimmun. Rev. 2020, 19, 102573–102578. [Google Scholar] [CrossRef]
- Perricone, C.; Bartoloni, E.; Bursi, R.; Cafaro, G.; Guidelli, G.M.; Shoenfeld, Y.; Gerli, R. COVID-19 as Part of the Hyperferritinemic Syndromes: The Role of Iron Depletion Therapy. Immunol. Res. 2020, 68, 213–224. [Google Scholar] [CrossRef]
- Suriawinata, E.; Mehta, K.J. Iron and Iron-Related Proteins in COVID-19. Clin. Exp. Med. 2022, 1–23. [Google Scholar] [CrossRef]
- Sonnweber, T.; Boehm, A.; Sahanic, S.; Pizzini, A.; Aichner, M.; Sonnweber, B.; Kurz, K.; Koppelstätter, S.; Haschka, D.; Petzer, V.; et al. Persisting Alterations of Iron Homeostasis in COVID-19 Are Associated with Non-Resolving Lung Pathologies and Poor Patients’ Performance: A Prospective Observational Cohort Study. Respir. Res. 2020, 21, 276–285. [Google Scholar] [CrossRef]
- Habib, H.M.; Ibrahim, S.; Zaim, A.; Ibrahim, W.H. The Role of Iron in the Pathogenesis of COVID-19 and Possible Treatment with Lactoferrin and Other Iron Chelators. Biomed. Pharmacother. 2021, 136, 111228–111238. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.J.; Sugiyama, M.; Shimokawa, F.; Murakami, M.; Hashimoto, O.; Matsui, T.; Funaba, M. Response to Iron Overload in Cultured Hepatocytes. Sci. Rep. 2020, 10, 21184. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, M.F.; Goldberg, M.F.; Cerejo, R.; Tayal, A.H. Cerebrovascular Disease in COVID-19. Am. J. Neuroradiol. 2020, 41, 1170–1172. [Google Scholar] [CrossRef] [PubMed]
- Edeas, M.; Saleh, J.; Peyssonnaux, C. Iron: Innocent Bystander or Vicious Culprit in COVID-19 Pathogenesis? Int. J. Infect. Dis. 2020, 97, 303–305. [Google Scholar] [CrossRef] [PubMed]
- Cavezzi, A.; Troiani, E.; Corrao, S. COVID-19: Hemoglobin, Iron, and Hypoxia beyond Inflammation. A Narrative Review. Clin. Pract. 2020, 10, 24–30. [Google Scholar] [CrossRef]
- Hippchen, T.; Altamura, S.; Muckenthaler, M.U.; Merle, U. Hypoferremia Is Associated with Increased Hospitalization and Oxygen Demand in COVID-19 Patients. HemaSphere 2020, 4, e492. [Google Scholar] [CrossRef]
- Roeser, H.P.; Lee, G.R.; Nacht, S.; Cartwright, G.E. The Role of Ceruloplasmin in Iron Metabolism. J. Clin. Investig. 1970, 49, 2408–2417. [Google Scholar] [CrossRef]
- Granick, S. Ferritin: Its Properties and Significance for Iron Metabolism. Chem. Rev. 1945, 38, 379–403. [Google Scholar] [CrossRef]
- Sabatucci, A.; Vachette, P.; Vasilyev, V.B.; Beltramini, M.; Sokolov, A.; Pulina, M.; Salvato, B.; Angelucci, C.B.; Maccarrone, M.; Cozzani, I.; et al. Structural Characterization of the Ceruloplasmin: Lactoferrin Complex in Solution. J. Mol. Biol. 2007, 371, 1038–1046. [Google Scholar] [CrossRef]
- Gupta, Y.; Maciorowski, D.; Medernach, B.; Becker, D.P.; Durvasula, R.; Libertin, C.R.; Kempaiah, P. Iron Dysregulation in COVID-19 and Reciprocal Evolution of SARS-CoV-2: Natura Nihil Frustra Facit. J. Cell. Biochem. 2022, 123, 601–619. [Google Scholar] [CrossRef]
- Eid, R.; Arab, N.T.T.; Greenwood, M.T. Iron Mediated Toxicity and Programmed Cell Death: A Review and a Re-Examination of Existing Paradigms. Biochim. Biophys. Acta Mol. Cell Res. 2017, 1864, 399–430. [Google Scholar] [CrossRef] [PubMed]
- Winterbourn, C.C. Toxicity of Iron and Hydrogen Peroxide: The Fenton Reaction. Toxicol. Lett. 1995, 82–83, 969–974. [Google Scholar] [CrossRef] [PubMed]
- Kehrer, J.P. The Haber-Weiss Reaction and Mechanisms of Toxicity. Toxicology 2000, 149, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Xu, Y.; Zhang, K.; Shen, L.; Deng, M. Ferroptosis in COVID-19-Related Liver Injury: A Potential Mechanism and Therapeutic Target. Front. Cell. Infect. Microbiol. 2022, 12, 922511. [Google Scholar] [CrossRef] [PubMed]
- Lazarian, G.; Quinquenel, A.; Bellal, M.; Siavellis, J.; Jacquy, C.; Re, D.; Merabet, F.; Mekinian, A.; Braun, T.; Damaj, G.; et al. Autoimmune Haemolytic Anaemia Associated with COVID-19 Infection. Br. J. Haematol. 2020, 190, 29–31. [Google Scholar] [CrossRef]
- Gadotti, A.C.; Lipinski, A.L.; Vasconcellos, F.T.; Marqueze, L.F.; Cunha, E.B.; Campos, A.C.; Oliveira, C.F.; Amaral, A.N.; Baena, C.P.; Telles, J.P.; et al. Susceptibility of the Patients Infected with SARS-CoV2 to Oxidative Stress and Possible Interplay with Severity of the Disease. Free Radic. Biol. Med. 2021, 165, 184–190. [Google Scholar] [CrossRef]
- Tiwari, N.R.; Redewad, N.K.; Ketkar, A.A.; Sharma, V.R. Elevated Plasma Hemoglobin in COVID-19–Related Illnesses: A Critical Appraisal. Ann. Hematol. 2021, 100, 2125–2126. [Google Scholar] [CrossRef]
- Terpos, E.; Ntanasis-Stathopoulos, I.; Elalamy, I.; Kastritis, E.; Sergentanis, T.N.; Politou, M.; Psaltopoulou, T.; Gerotziafas, G.; Dimopoulos, M.A. Hematological Findings and Complications of COVID-19. Am. J. Hematol. 2020, 95, 834–847. [Google Scholar] [CrossRef] [Green Version]
- Aggarwal, S.; Ahmad, I.; Lam, A.; Carlisle, M.A.; Li, C.; Wells, J.M.; Raju, S.V.; Athar, M.; Rowe, S.M.; Dransfield, M.T.; et al. Heme Scavenging Reduces Pulmonary Endoplasmic Reticulum Stress, Fibrosis, and Emphysema. JCI Insight 2018, 3, e120694. [Google Scholar] [CrossRef] [Green Version]
- Aggarwal, S.; Lam, A.; Bolisetty, S.; Carlisle, M.A.; Traylor, A.; Agarwal, A.; Matalon, S. Heme Attenuation Ameliorates Irritant Gas Inhalation-Induced Acute Lung Injury. Antioxidants Redox Signal. 2016, 24, 99–112. [Google Scholar] [CrossRef] [Green Version]
- Poggi, C.; Dani, C. Antioxidant Strategies and Respiratory Disease of the Preterm Newborn: An Update. Oxid. Med. Cell. Longev. 2014, 2014, 721043. [Google Scholar] [CrossRef] [PubMed]
- Martín-Fernández, M.; Aller, R.; Heredia-Rodríguez, M.; Gómez-Sánchez, E.; Martínez-Paz, P.; Gonzalo-Benito, H.; Sánchez-de Prada, L.; Gorgojo, Ó.; Carnicero-Frutos, I.; Tamayo, E.; et al. Lipid Peroxidation as a Hallmark of Severity in COVID-19 Patients. Redox Biol. 2021, 48, 102181–102188. [Google Scholar] [CrossRef] [PubMed]
- Mehri, F.; Rahbar, A.H.; Ghane, E.T.; Souri, B.; Esfahani, M. Changes in Oxidative Markers in COVID-19 Patients. Arch. Med. Res. 2021, 52, 843–849. [Google Scholar] [CrossRef] [PubMed]
- Tamagawa, K.; Taooka, Y.; Maeda, A.; Hiyama, K.; Ishioka, S.; Yamakido, M. Inhibitory Effects of a Lecithinized Superoxide Dismutase on Bleomycin- Induced Pulmonary Fibrosis in Mice. Am. J. Respir. Crit. Care Med. 2000, 161, 1279–1284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, K.I.; Ishihara, T.; Azuma, A.; Kudoh, S.; Ebina, M.; Nukiwa, T.; Sugiyama, Y.; Tasaka, Y.; Namba, T.; Ishihara, T.; et al. Therapeutic Effect of Lecithinized Superoxide Dismutase on Bleomycin-Induced Pulmonary Fibrosis. Am. J. Physiol. Lung Cell. Mol. Physiol. 2010, 298, 348–360. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.I.; Tamura, F.; Sugizaki, T.; Kawahara, M.; Kuba, K.; Imai, Y.; Mizushima, T. Evaluation of Lecithinized Superoxide Dismutase for the Prevention of Acute Respiratory Distress Syndrome in Animal Models. Am. J. Respir. Cell Mol. Biol. 2017, 56, 179–190. [Google Scholar] [CrossRef]
- Yaghoubi, N.; Youssefi, M.; Jabbari Azad, F.; Farzad, F.; Yavari, Z.; Zahedi Avval, F. Total Antioxidant Capacity as a Marker of Severity of COVID-19 Infection: Possible Prognostic and Therapeutic Clinical Application. J. Med. Virol. 2022, 94, 1558–1565. [Google Scholar] [CrossRef]
- Abouhashem, A.S.; Singh, K.; Azzazy, H.M.E.; Sen, C.K. Is Low Alveolar Type II Cell SOD3 in the Lungs of Elderly Linked to the Observed Severity of COVID-19? Antioxidants Redox Signal. 2020, 33, 59–65. [Google Scholar] [CrossRef]
- Keles, E.S. Mild SARS-CoV-2 Infections in Children Might Be Based on Evolutionary Biology and Linked with Host Reactive Oxidative Stress and Antioxidant Capabilities. New Microbes New Infect. 2020, 36, 100723. [Google Scholar] [CrossRef]
- Maes, M. An Intriguing and Hitherto Unexplained Co-Occurrence: Depression and Chronic Fatigue Syndrome Are Manifestations of Shared Inflammatory, Oxidative and Nitrosative (IO&NS) Pathways. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2011, 35, 784–794. [Google Scholar] [CrossRef]
- Liu, T.; Zhong, S.; Liao, X.; Chen, J.; He, T.; Lai, S.; Jia, Y. A Meta-Analysis of Oxidative Stress Markers in Depression. PLoS ONE 2015, 10, e0138904. [Google Scholar] [CrossRef] [PubMed]
- Maes, M.; Rachayon, M.; Jirakran, K.; Sodsai, P.; Klinchanhom, S.; Gałecki, P.; Sughondhabirom, A.; Basta-Kaim, A. The Immune Profile of Major Dysmood Disorder: Proof of Concept and Mechanism Using the Precision Nomothetic Psychiatry Approach. Cells 2022, 11, 1183. [Google Scholar] [CrossRef] [PubMed]
- Al-Hakeim, H.K.; Al-Rubaye, H.T.; Al-Hadrawi, D.S.; Almulla, A.F.; Maes, M. Long-COVID Post-Viral Chronic Fatigue Syndrome and Affective Symptoms Are Associated with Oxidative Damage, Lowered Antioxidant Defenses and Inflammation: A Proof of Concept and Mechanism Study. Mol Psychiatry 2022, 24, 1–15. [Google Scholar] [CrossRef]
- Paul, B.D.; Lemle, M.D.; Komaroff, A.L.; Snyder, S.H. Redox Imbalance Links COVID-19 and Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. Proc. Natl. Acad. Sci. USA 2021, 118, e2024358118. [Google Scholar] [CrossRef]
- Fox, P.L.; Mazumder, B.; Ehrenwald, E.; Mukhopadhyay, C.K. Ceruloplasmin and Cardiovascular Disease. Free Radic. Biol. Med. 2000, 28, 1735–1744. [Google Scholar] [CrossRef]
- Lage, S.L.; Amaral, E.P.; Hilligan, K.L.; Laidlaw, E.; Rupert, A.; Namasivayan, S.; Rocco, J.; Galindo, F.; Kellogg, A.; Kumar, P.; et al. Persistent Oxidative Stress and Inflammasome Activation in CD14highCD16− Monocytes From COVID-19 Patients. Front. Immunol. 2022, 12, 799558. [Google Scholar] [CrossRef]
- Vollbracht, C.; Kraft, K. Oxidative Stress and Hyper-Inflammation as Major Drivers of Severe COVID-19 and Long COVID: Implications for the Benefit of High-Dose Intravenous Vitamin C. Front. Pharmacol. 2022, 13, e2024358118. [Google Scholar] [CrossRef]
- Menshawey, R.; Menshawey, E.; Alserr, A.H.K.; Abdelmassih, A.F. Low Iron Mitigates Viral Survival: Insights from Evolution, Genetics, and Pandemics—A Review of Current Hypothesis. Egypt. J. Med. Hum. Genet. 2020, 21, 2–14. [Google Scholar] [CrossRef]
- Chang, H.C.; Bayeva, M.; Taiwo, B.; Palella, F.J.; Hope, T.J.; Ardehali, H. Short Communication: High Cellular Iron Levels Are Associated with Increased HIV Infection and Replication. AIDS Res. Hum. Retroviruses 2015, 31, 305–312. [Google Scholar] [CrossRef]
- Kinsella, M.; Monk, C. Evaluation of Iron Deficiency As a Nutritional Adaptation to Infectious Disease: An Evolutionary Medicine Perspective. Am. J. Hum. Biol. 2012, 23, 172–179. [Google Scholar] [CrossRef]
- Chakurkar, V.; Rajapurkar, M.; Lele, S.; Mukhopadhyay, B.; Lobo, V.; Injarapu, R.; Sheikh, M.; Dholu, B.; Ghosh, A.; Jha, V. Increased Serum Catalytic Iron May Mediate Tissue Injury and Death in Patients with COVID-19. Sci. Rep. 2021, 11, 19618. [Google Scholar] [CrossRef] [PubMed]
- Mohus, R.M.; Flatby, H.; Liyanarachi, K.V.; DeWan, A.T.; Solligård, E.; Damås, J.K.; Åsvold, B.O.; Gustad, L.T.; Rogne , T. Iron Status and the Risk of Sepsis and Severe COVID-19: A Two-Sample Mendelian Randomization Study. Sci. Rep. 2022, 12, 16157. [Google Scholar] [CrossRef] [PubMed]
- Hanauske-Abel, H.M.; Saxena, D.; Palumbo, P.E.; Hanauske, A.R.; Luchessi, A.D.; Cambiaghi, T.D.; Hoque, M.; Spino, M.; Gandolfi, D.D.A.; Heller, D.S.; et al. Drug-Induced Reactivation of Apoptosis Abrogates HIV-1 Infection. PLoS ONE 2013, 8, e74414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chhabra, R.; Saha, A.; Chamani, A.; Schneider, N.; Nanjundan, M.; Shah, R. Iron pathways and iron chelation approaches in viral, microbial, and fungal infections. Pharmaceuticals 2020, 13, 275. [Google Scholar] [CrossRef]
- Abobaker, A. Reply: Iron Chelation May Harm Patients with COVID-19. Eur. J. Clin. Pharmacol. 2021, 77, 267–268. [Google Scholar] [CrossRef]
- Bonyek-Silva, I.; Machado, A.F.A.; Cerqueira-Silva, T.; Nunes, S.; Cruz, M.R.S.; Silva, J.; Santos, R.L.; Barral, A.; Oliveira, P.R.S.; Khouri, R.; et al. LTB4-Driven Inflammation and Increased Expression of ALOX5/ACE2 during Severe COVID-19 in Individuals with Diabetes. Diabetes 2021, 70, 2120–2130. [Google Scholar] [CrossRef]
- Dugas, B.; Paul-Eugene, N.; Cairns, J.; Gordon, J.; Calenda, A.; Mencia-Huerta, J.M.; Braquet, P. LeukotrieneB4 Potentiates the Expression and Release of FceRII/CD23, and Proliferation and Differentiation of Human B Lymphocytes Induced by IL-4. J. Immunol. 1990, 145, 3406–3411. [Google Scholar]
- Lone, A.M.; Taskén, K. Proinflammatory and Immunoregulatory Roles of Eicosanoids in T Cells. Front. Immunol. 2013, 4, 00130. [Google Scholar] [CrossRef] [Green Version]
- Spanbroek, R.; Hildner, M.; Köhler, A.; Müller, A.; Zintl, F.; Kühn, H.; Rådmark, O.; Samuelsson, B.; Habenicht, A.J.R. IL-4 Determines Eicosanoid Formation in Dendritic Cells by down-Regulation of 5-Lipoxygenase and up-Regulation of 15-Lipoxygenase 1 Expression. Proc. Natl. Acad. Sci. USA 2001, 98, 5152–5157. [Google Scholar] [CrossRef] [Green Version]
- Vaz de Paula, C.B.; de Azevedo, M.L.V.; Nagashima, S.; Martins, A.P.C.; Malaquias, M.A.S.; Miggiolaro, A.F.R.d.S.; da Silva Motta Júnior, J.; Avelino, G.; do Carmo, L.A.P.; Carstens, L.B.; et al. IL-4/IL-13 Remodeling Pathway of COVID-19 Lung Injury. Sci. Rep. 2020, 10, 4–11. [Google Scholar] [CrossRef]
- Afonso, P.V.; Janka-Junttila, M.; Lee, Y.J.; McCann, C.P.; Oliver, C.M.; Aamer, K.A.; Losert, W.; Cicerone, M.T.; Parent, C.A. LTB4 Is a Signal-Relay Molecule during Neutrophil Chemotaxis. Dev. Cell 2012, 22, 1079–1091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Funk, C.D.; Ardakani, A. A Novel Strategy to Mitigate the Hyperin Fl Ammatory Response to COVID-19 by Targeting Leukotrienes. Front. Pharmacol. 2020, 11, 01214. [Google Scholar] [CrossRef] [PubMed]
- Miyahara, N.; Miyahara, S.; Takeda, K.; Gelfand, E.W. Role of the LTB4/BLT1 Pathway in Allergen-Induced Airway Hyperresponsiveness and Inflammation. Allergol. Int. 2006, 55, 91–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Bel, M.; Brunet, A.; Gosselin, J. Leukotriene B4, an Endogenous Stimulator of the Innate Immune Response against Pathogens. J. Innate Immun. 2014, 6, 159–168. [Google Scholar] [CrossRef]
- Widegren, H.; Andersson, M.; Borgeat, P.; Flamand, L.; Johnston, S.; Greiff, L. LTB4 Increases Nasal Neutrophil Activity and Conditions Neutrophils to Exert Antiviral Effects. Respir. Med. 2011, 105, 997–1006. [Google Scholar] [CrossRef] [Green Version]
- He, R.; Chen, Y.; Cai, Q. The Role of the LTB4-BLT1 Axis in Health and Disease. Pharmacol. Res. 2020, 158, 104857. [Google Scholar] [CrossRef]
- Brandt, S.L.; Wang, S.; Dejani, N.N.; Klopfenstein, N.; Winfree, S.; Filgueiras, L.; McCarthy, B.P.; Territo, P.R.; Serezani, C.H. Excessive Localized Leukotriene B4 Levels Dictate Poor Skin Host Defense in Diabetic Mice. JCI Insight 2018, 3, e120220. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Zhao, A.; Wong, F.; Ayala, J.M.; Struthers, M.; Ujjainwalla, F.; Wright, S.D.; Springer, M.S.; Evans, J.; Cui, J. Leukotriene B4 Strongly Increases Monocyte Chemoattractant Protein-1 in Human Monocytes. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 1783–1788. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, F.; Yokomizo, T. The Leukotriene Receptors as Therapeutic Targets of Inflammatory Diseases. Int. Immunol. 2019, 31, 607–615. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Lam, B.K.; Kanaoka, Y.; Nigrovic, P.A.; Audoly, L.P.; Austen, K.F.; Lee, D.M. Neutrophil-Derived Leukotriene B4 Is Required for Inflammatory Arthritis. J. Exp. Med. 2006, 203, 837–842. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Clemente, M.; Clària, J.; Titos, E. The 5-Lipoxygenase/Leukotriene Pathway in Obesity, Insulin Resistance, and Fatty Liver Disease. Curr. Opin. Clin. Nutr. Metab. Care 2011, 14, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Maamar, M.; Artime, A.; Pariente, E.; Fierro, P.; Ruiz, Y.; Gutiérrez, S.; Tobalina, M.; Díaz-Salazar, S.; Ramos, C.; Olmos, J.M.; et al. Post-COVID-19 Syndrome, Low-Grade Inflammation and Inflammatory Markers: A Cross-Sectional Study. Curr. Med. Res. Opin. 2022, 38, 901–909. [Google Scholar] [CrossRef]
- Nalbandian, A.; Sehgal, K.; Gupta, A.; Madhavan, M.V.; McGroder, C.; Stevens, J.S.; Cook, J.R.; Nordvig, A.S.; Shalev, D.; Sehrawat, T.S.; et al. Post-Acute COVID-19 Syndrome. Nat. Med. 2021, 27, 601–615. [Google Scholar] [CrossRef] [PubMed]
- Furman, D.; Campisi, J.; Verdin, E.; Carrera-Bastos, P.; Targ, S.; Franceschi, C.; Ferrucci, L.; Gilroy, D.W.; Fasano, A.; Miller, G.W.; et al. Chronic Inflammation in the Etiology of Disease across the Life Span. Nat. Med. 2019, 25, 1822–1832. [Google Scholar] [CrossRef] [PubMed]
- Bohnacker, S.; Hartung, F.; Henkel, F.; Quaranta, A.; Kolmert, J.; Priller, A.; Ud-dean, M. Mild COVID-19 Imprints a Long-Term in Fl Ammatory Eicosanoid- and Chemokine Memory in Monocyte-Derived Macrophages. Mucosal Immunol. 2022, 15, 515–524. [Google Scholar] [CrossRef] [PubMed]
- Goetz, D.H.; Holmes, M.A.; Borregaard, N.; Bluhm, M.E.; Raymond, K.N.; Strong, R.K. The Neutrophil Lipocalin NGAL Is a Bacteriostatic Agent That Interferes with Siderophore-Mediated Iron Acquisition. Mol. Cell 2002, 10, 1033–1043. [Google Scholar] [CrossRef]
- Flo, T.; Smith, K.D.; Sato, S.; David, R.J.; Holmes, M.; Strong, R.K.; Aderem, A. Lipocalin 2 Mediates an Innate Immune Response to Bacterial Infection by Sequestering Iron. Nature 2004, 432, 913–917. [Google Scholar] [CrossRef]
- Akgül, B.; Bauer, B.; Zigrino, P.; Storey, A.; Mauch, C.; Pfister, H. Upregulation of Lipocalin-2 in Human Papillomavirus-Positive Keratinocytes and Cutaneous Squamous Cell Carcinomas. J. Gen. Virol. 2011, 92, 395–401. [Google Scholar] [CrossRef]
- Noçon, A.L.; Ip, J.P.K.; Terry, R.; Lim, S.L.; Getts, D.R.; Müller, M.; Hofer, M.J.; King, N.J.C.; Campbell, I.L. The Bacteriostatic Protein Lipocalin 2 Is Induced in the Central Nervous System of Mice with West Nile Virus Encephalitis. J. Virol. 2014, 88, 679–689. [Google Scholar] [CrossRef] [Green Version]
- Al Jaberi, S.; Cohen, A.; D’Souza, C.; Abdulrazzaq, Y.M.; Ojha, S.; Bastaki, S.; Adeghate, E.A. Lipocalin-2: Structure, Function, Distribution and Role in Metabolic Disorders. Biomed. Pharmacother. 2021, 142, 112002. [Google Scholar] [CrossRef]
- Flower, D.R.; North, A.C.T.; Sansom, C.E. The Lipocalin Protein Family: Structural and Sequence Overview. Biochim. Biophys. Acta-Protein Struct. Mol. Enzymol. 2000, 1482, 9–24. [Google Scholar] [CrossRef] [PubMed]
- Fasshauer, M.; Blu, M. Adipokines in Health and Disease. Rev. Trends Pharmacol. Sci. 2015, 36, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Rubinstein, T.; Pitashny, M.; Putterman, C. The Novel Role of Neutrophil Gelatinase-B Associated Lipocalin (NGAL)/Lipocalin-2 as a Biomarker for Lupus Nephritis. Autoimmun. Rev. 2008, 7, 229–234. [Google Scholar] [CrossRef]
- Mike, E.V.; Makinde, H.M.; Gulinello, M.; Vanarsa, K.; Herlitz, L.; Gadhvi, G.; Winter, D.R.; Mohan, C.; Hanly, J.G.; Mok, C.C.; et al. Lipocalin-2 Is a Pathogenic Determinant and Biomarker of Neuropsychiatric Lupus. J. Autoimmun. 2019, 96, 59–73. [Google Scholar] [CrossRef] [PubMed]
- Warszawska, J.M.; Gawish, R.; Sharif, O.; Sigel, S.; Doninger, B.; Lakovits, K.; Mesteri, I.; Nairz, M.; Boon, L.; Spiel, A.; et al. Lipocalin 2 Deactivates Macrophages and Worsens Pneumococcal Pneumonia Outcomes. J. Clin. Investig. 2013, 123, 3363–3372. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Li, S.; Tang, X.; Liang, L.; Wang, F.; Du, H. Lipocalin 2 Protects Against Escherichia Coli Infection by Modulating Neutrophil and Macrophage Function. Front. Immunol. 2019, 10, 02594. [Google Scholar] [CrossRef] [Green Version]
- Hu, L.; Hittelman, W.; Lu, T.; Ji, P.; Arlinghaus, R.; Shmulevich, I.; Hamilton, S.R.; Zhang, W. NGAL Decreases E-Cadherin-Mediated Cellcell Adhesion and Increases Cell Motility and Invasion through Rac1 in Colon Carcinoma Cells. Lab. Investig. 2009, 89, 531–548. [Google Scholar] [CrossRef] [Green Version]
- Xiao, X.; Yeoh, B.S.; Saha, P.; Olvera, R.A.; Singh, V.; Vijay-Kumar, M. Lipocalin 2 Alleviates Iron Toxicity by Facilitating Hypoferremia of Inflammation and Limiting Catalytic Iron Generation. BioMetals 2016, 29, 451–465. [Google Scholar] [CrossRef] [Green Version]
- Bao, G.; Clifton, M.; Hoette, T.M.; Mori, K.; Deng, S.; Qiu, A.; Viltard, M.; Williams, D.; Paragas, N.; Leete, T.; et al. Iron Traffics in Circulation Bound to a Siderocalin (Ngal)- Catechol Complex. Nat Chem Biol. 2010, 6, 602–609. [Google Scholar] [CrossRef]
- Barasch, J.; Hollmen, M.; Deng, R.; Hod, E.A.; Rupert, P.B.; Abergel, R.J.; Allred, B.E.; Xu, K.; Darrah, S.F.; Tekabe, Y.; et al. Disposal of Iron by a Mutant Form of Lipocalin 2. Nat. Commun. 2016, 7, 12973. [Google Scholar] [CrossRef]
- Bao, G.; Road, C.; Xu, J.; Road, C.; Hu, F.; Wan, X.; Road, C.; Deng, S.; Barasch, J. EGCG Inhibit Chemical Reactivity of Iron through Forming an Ngal–EGCG–Iron Complex. Biometals 2015, 26, 1041–1050. [Google Scholar] [CrossRef]
- Wang, G.; Liu, X.; Meng, L.; Liu, S.; Wang, L.; Li, J.; Cui, C.; Meng, J.; Hu, S.; Wei, Y. Up-Regulated Lipocalin-2 in Pulmonary Hypertension Involving in Pulmonary Artery SMC Resistance to Apoptosis. Int. J. Biol. Sci. 2014, 10, 798–806. [Google Scholar] [CrossRef] [PubMed]
- Halabian, R.; Tehrani, H.A.; Jahanian-Najafabadi, A.; Habibi Roudkenar, M. Lipocalin-2-Mediated Upregulation of Various Antioxidants and Growth Factors Protects Bone Marrow-Derived Mesenchymal Stem Cells against Unfavorable Microenvironments. Cell Stress Chaperones 2013, 18, 785–800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callard, F.; Perego, E. How and Why Patients Made Long COVID. Soc. Sci. Med. 2021, 268, 2–5. [Google Scholar] [CrossRef] [PubMed]
- Marchisio, M.; Simeone, P.; Bologna, G.; Ercolino, E.; Pierdomenico, L.; Pieragostino, D.; Ventrella, A.; Antonini, F.; Del Zotto, G.; Vergara, D.; et al. Flow Cytometry Analysis of Circulating Extracellular Vesicle Subtypes from Fresh Peripheral Blood Samples. Int. J. Mol. Sci. 2021, 22, 48. [Google Scholar] [CrossRef] [PubMed]
- Cela, I.; Cufaro, M.C.; Fucito, M.; Pieragostino, D.; Lanuti, P.; Sallese, M.; Del Boccio, P.; Di Matteo, A.; Allocati, N.; De Laurenzi, V.; et al. Proteomic Investigation of the Role of Nucleostemin in Nucleophosmin-Mutated OCI-AML 3 Cell Line. Int. J. Mol. Sci. 2022, 23, 7655. [Google Scholar] [CrossRef]
- Dufrusine, B.; Damiani, V.; Capone, E.; Pieragostino, D.; Dainese, E.; De Marco, M.; Reppucci, F.; Turco, M.C.; Rosati, A.; Marzullo, L.; et al. BAG3 Induces Fibroblasts to Release Key Cytokines Involved in Pancreatic Cell Migration. J. Cell. Biochem. 2022, 123, 65–76. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Value in COVID-19 (n = 30) | Value in Long-COVID (n = 10) | p-Value |
|---|---|---|---|
| Gender (Female), n (%) | 14 (46.7%) | 5 (50%) | n.s. |
| Age, mean ± SD | 67.5 ± 14.8 | 62.7 ± 13.2 | n.s |
| Symptoms, n (%) | |||
| Fever | 20 (66.7%) | 3 (30%) | n.s. |
| Cough | 8 (26.7%) | 3 (30%) | n.s. |
| Dyspnea | 13 (43.3%) | 10 (100%) | 0.002 ** |
| Diarrhea | 2 (6.7%) | - | n.a. |
| Asthenia | 4 (13.3%) | - | n.a. |
| Comorbid Conditions, n (%) | |||
| Any | - | 3 (30%) | n.a. |
| Hypertension | 15 (50%) | 3 (30%) | n.s. |
| Diabetes Mellitus II | 5 (16.7%) | 2 (20%) | n.s. |
| Hypothyroidism | 1 (3.3%) | 1 (10%) | n.s. |
| Cancer | 5 (16.7%) | - | n.a. |
| Chronic kidney disease | 3 (10%) | - | n.a. |
| Obesity | 2 (6.7%) | - | n.a. |
| IL-6 (pg/mL) | |||
| Median value (min.–max.) | 128.6 (3.6–278.9) | 64.8 (11.1–194) | n.a. |
| Cut-off < 6.4 pg/mL | |||
| Ferritin (ng/mL) | |||
| Median value (min.–max.) | 991.8 (37.2–4265) | 1608.5 (205–4638) | n.a. |
| Cut-off 22–274 ng/mL |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dufrusine, B.; Valentinuzzi, S.; Bibbò, S.; Damiani, V.; Lanuti, P.; Pieragostino, D.; Del Boccio, P.; D’Alessandro, E.; Rabottini, A.; Berghella, A.; et al. Iron Dyshomeostasis in COVID-19: Biomarkers Reveal a Functional Link to 5-Lipoxygenase Activation. Int. J. Mol. Sci. 2023, 24, 15. https://doi.org/10.3390/ijms24010015
Dufrusine B, Valentinuzzi S, Bibbò S, Damiani V, Lanuti P, Pieragostino D, Del Boccio P, D’Alessandro E, Rabottini A, Berghella A, et al. Iron Dyshomeostasis in COVID-19: Biomarkers Reveal a Functional Link to 5-Lipoxygenase Activation. International Journal of Molecular Sciences. 2023; 24(1):15. https://doi.org/10.3390/ijms24010015
Chicago/Turabian StyleDufrusine, Beatrice, Silvia Valentinuzzi, Sandra Bibbò, Verena Damiani, Paola Lanuti, Damiana Pieragostino, Piero Del Boccio, Ersilia D’Alessandro, Alberto Rabottini, Alessandro Berghella, and et al. 2023. "Iron Dyshomeostasis in COVID-19: Biomarkers Reveal a Functional Link to 5-Lipoxygenase Activation" International Journal of Molecular Sciences 24, no. 1: 15. https://doi.org/10.3390/ijms24010015