
Interplay of Cellular mRNA, miRNA and Viral miRNA during Infection of a Cell

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. General Scheme
2.2. Coarse-Grained Steps
2.3. Mean-Field Kinetic Equations
2.4. Stochasticity
3. Results and Discussion
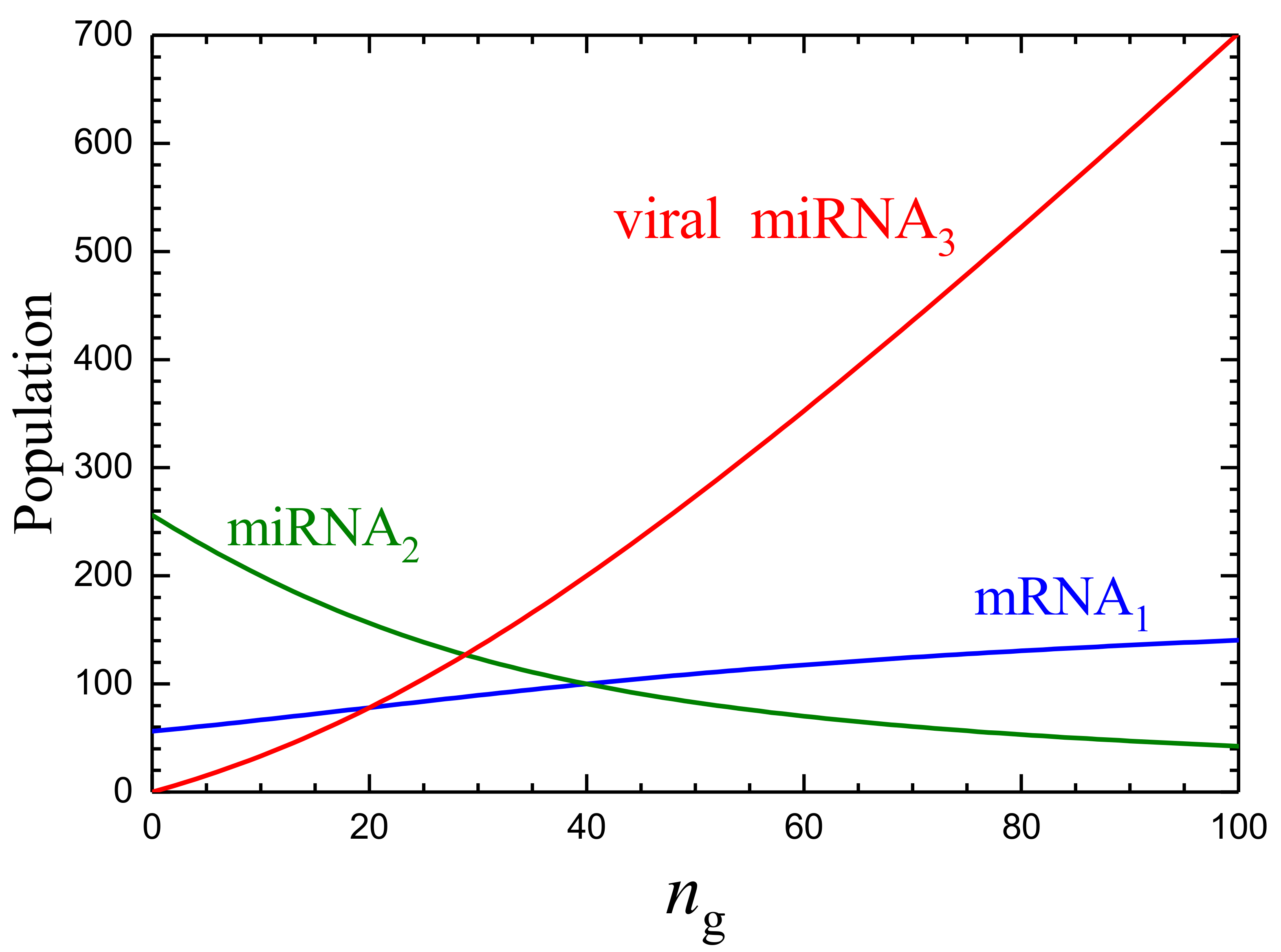
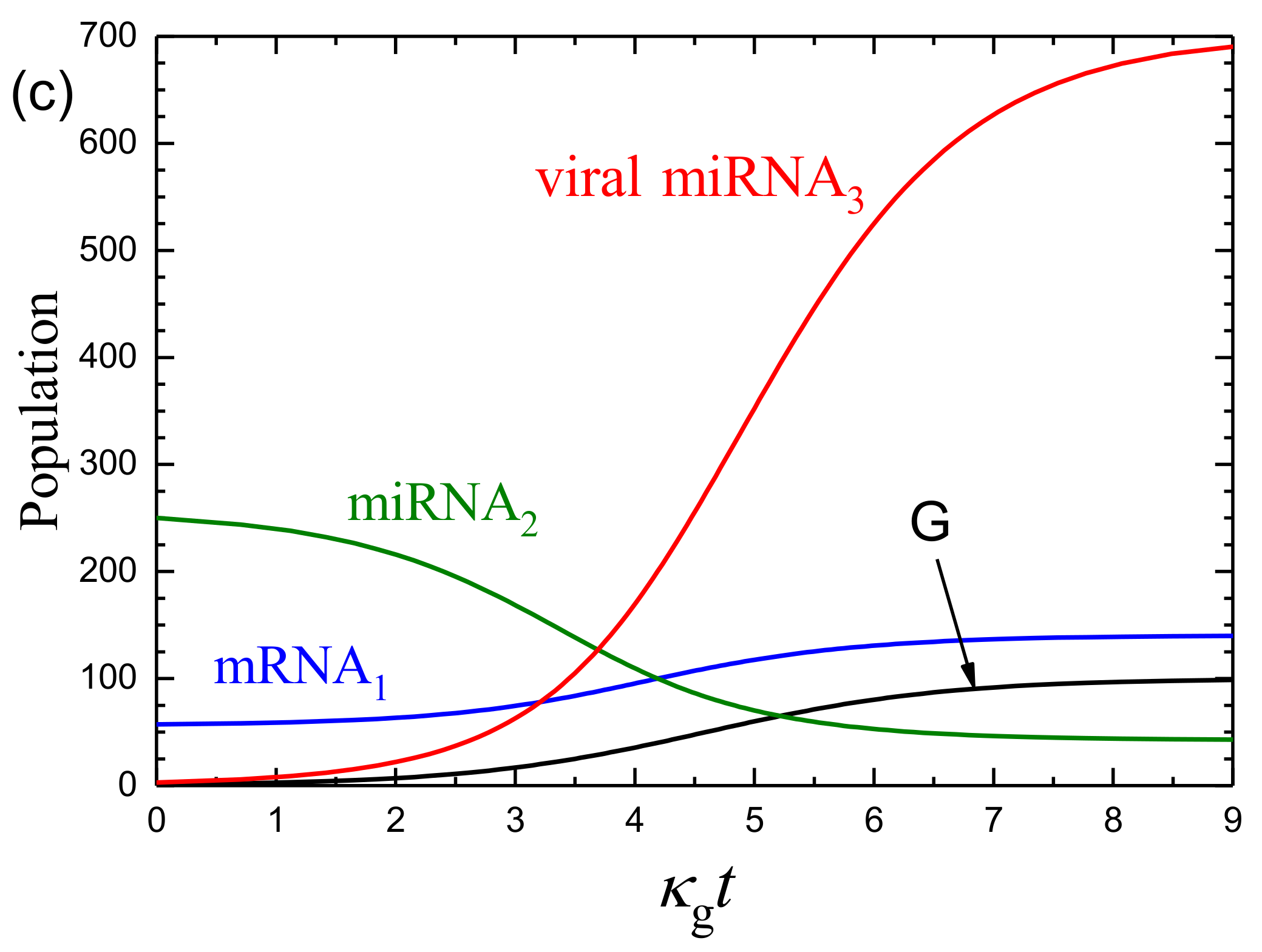
3.1. Mean-Field Kinetics
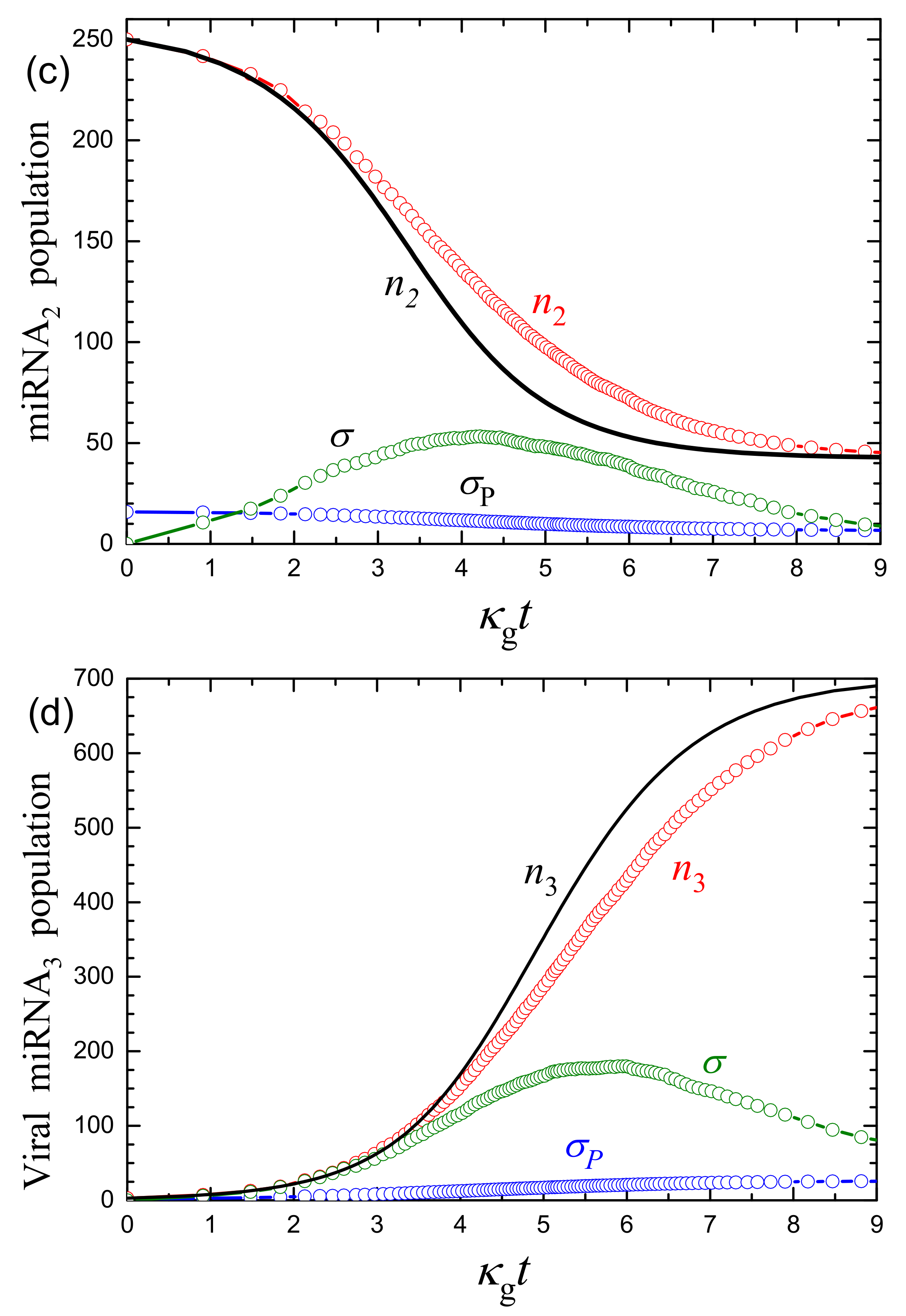
3.2. Monte Carlo Simulations
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Orphanides, G.; Reinber, D. A unified theory of gene expression. Cell 2002, 108, 439–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaern, M.; Elston, T.C.; Blake, W.J.; Collins, J.J. Stochasticity in gene expression: From theories to phenotypes. Nat. Rev. Genet. 2005, 6, 451–464. [Google Scholar] [CrossRef] [PubMed]
- Paulsson, J. Models of stochastic gene expression. Phys. Life Sci. 2005, 2, 157–175. [Google Scholar] [CrossRef]
- Karlebach, G.; Shamir, R. Modelling and analysis of gene regulatory networks. Nat. Rev. Molec. Cell Biol. 2008, 9, 771–780. [Google Scholar] [CrossRef] [PubMed]
- Vilar, J.M.G.; Saiz, L. Systems biophysics of gene expression. Biophys. J. 2013, 104, 2574–2585. [Google Scholar] [CrossRef] [Green Version]
- Zhdanov, V.P. Kinetic models of gene expression including non-coding RNAs. Phys. Rep. 2011, 500, 1–42. [Google Scholar] [CrossRef]
- Lin, J.; Amir, A. Disentangling intrinsic and extrinsic gene expression noise in growing cells. Phys. Rev. Lett. 2021, 126, 078101. [Google Scholar] [CrossRef]
- Matsushita, Y.; Hatakeyama, T.S.; Kaneko, K. Dynamical systems theory of cellular reprogramming. Phys. Rev. Res. 2022, 4, L022008. [Google Scholar] [CrossRef]
- Silverthorne, T.; Oh, E.S.; Stinchcombe, A.R. Promoter methylation in a mixed feedback loop circadian clock model. Phys. Rev. E 2022, 105, 034411. [Google Scholar] [CrossRef]
- Chen, M.; Luo, S.; Cao, M.; Guo, C.; Zhou, T.; Zhang, J. Exact distributions for stochastic gene expression models with arbitrary promoter architecture and translational bursting. Phys. Rev. E 2022, 105, 014405. [Google Scholar] [CrossRef]
- Szavits-Nossan, J.; Grima, R. Mean-field theory accurately captures the variation of copy number distributions across the mRNA life cycle. Phys. Rev. E 2022, 105, 014410. [Google Scholar] [CrossRef] [PubMed]
- Handel, A.; Liao, L.E.; Beauchemin, C.A.A. Progress and trends in mathematical modelling of influenza A virus infections. Curr. Opin. Syst. Biol. 2018, 12, 30–36. [Google Scholar] [CrossRef]
- Yin, J.; Redovich, J. Kinetic modeling of virus growth in cells. Microbiol. Mol. Biol. Rev. 2018, 82, e00066-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goyal, A.; Liao, E.; Perelson, A.S. Within-host mathematical models of hepatitis B virus infection: Past, present, and future. Curr. Opin. Syst. Biol. 2019, 18, 27–35. [Google Scholar] [CrossRef]
- Srivastava, R.; You, L.; Summers, J.; Yin, J. Stochastic vs. deterministic modeling of intracellular viral kinetics. J. Theor. Biol. 2002, 218, 309–324. [Google Scholar] [CrossRef] [Green Version]
- Sidorenko, Y.; Reichl, U. Structured model of influenza virus replication in MDCK cells. Biotechn. Bioing. 2004, 88, 1–14. [Google Scholar] [CrossRef]
- Sidorenko, Y.; Voigt, A.; Schulze-Horsel, J.; Reichl, U.; Kienle, A. Stochastic population balance modeling of influenza virus replication in vaccine production processes. II. Detailed description of the replication mechanism. Chem. Eng. Sci. 2008, 63, 2299–2304. [Google Scholar] [CrossRef]
- Sardanyes, J.; Sole, R.V.; Elena, S.F. Replication mode and landscape topology differentially affect RNA virus mutational load and robustness. J. Virol. 2009, 83, 12579–12589. [Google Scholar] [CrossRef] [Green Version]
- Rodrigo, G.; Carrera, J.; Jaramillo, A.; Elena, S.F. Optimal viral strategies for bypassing RNA silencing. J. Roy. Soc. Interface 2011, 8, 257–268. [Google Scholar] [CrossRef] [Green Version]
- Sardanyes, J.; Martinez, F.; Daros, J.-A.; Elena, S.F. Dynamics of alternative modes of RNA replication for positive-sense RNA viruses. J. Roy. Soc. Interface 2012, 9, 768–776. [Google Scholar] [CrossRef]
- Andreu-Moreno, I.; Bou, J.-V.; Sanjuan, R. Cooperative nature of viral replication. Sci. Adv. 2020, 6, eabd4942. [Google Scholar] [CrossRef] [PubMed]
- Lopacinski, A.B.; Sweatt, A.J.; Smolko, C.M.; Gray-Gaillard, E.; Borgman, C.A.; Shah, M.; Janes, K.A. Modeling the complete kinetics of coxsackievirus B3 reveals human determinants of host-cell feedback. Cell Syst. 2021, 12, 304–323.e13. [Google Scholar] [CrossRef] [PubMed]
- Chhajer, H.; Rizvi, V.A.; Roy, R. Life cycle process dependencies of positive-sense RNA viruses suggest strategies for inhibiting productive cellular infection. J. Roy. Soc. Interface 2021, 18, 20210401. [Google Scholar] [CrossRef] [PubMed]
- Zhdanov, V.P. Stochastic kinetics of reproduction of virions inside a cell. BioSystems 2004, 77, 143–150. [Google Scholar] [CrossRef]
- Zhdanov, V.P. Monte Carlo simulation of bifurcation in the intracellular viral kinetics. Phys. Biol. 2005, 2, 46–50. [Google Scholar] [CrossRef]
- Zhdanov, V.P. Intracellular viral kinetics limited by the supply of amino acids for synthesis of viral proteins. BioSystems 2009, 97, 117–120. [Google Scholar] [CrossRef]
- Zhdanov, V.P. Interplay of viral miRNAs and host mRNAs and proteins. Cent. Eur. J. Phys. 2011, 9, 1366–1371. [Google Scholar] [CrossRef]
- Zhdanov, V.P. Initial phase of replication of plus-stranded RNA viruses. Biophys. Rev. Lett. 2018, 13, 93–108. [Google Scholar] [CrossRef]
- Bruscella, P.; Bottini, S.; Baudesson, C.; Pawlotsky, J.M.; Feray, C.; Trabucchi, M. Viruses and miRNAs: More friends than foes. Front. Microbiol. 2017, 8, 824. [Google Scholar] [CrossRef] [Green Version]
- Geraylow, K.R.; Hemmati, R.; Kadkhoda, S.; Ghafouri-Fard, S. miRNA expression in COVID-19. Gene Rep. 2022, 28, 101641. [Google Scholar] [CrossRef]
- Khan, M.A.-A.-K.; Sany, M.R.U.; Islam, M.S.; Islam, A.B.M.M.K. Epigenetic regulator miRNA pattern differences among SARS-CoV, SARS-CoV-2, and SARS-CoV-2 world-wide isolates delineated the mystery behind the epic pathogenicity and distinct clinical characteristics of pandemic COVID-19. Front. Genet. 2020, 11, 765. [Google Scholar] [CrossRef] [PubMed]
- Pfeffer, S.; Zavolan, M.; Grasser, F.A.; Chien, M.; Russo, J.J.; Ju, J.; John, B.; Enright, A.J.; Marks, D.; Sander, C.; et al. Identification of virus-encoded microRNAs. Science 2004, 304, 734–736. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Lu, S.; Zhang, Z.; Gonzalez, C.M.; Damania, B.; Cullen, B.R. Kaposi’s sarcoma-associated herpesvirus expresses an array of viral microRNAs in latently infected cells. Proc. Natl. Acad. Sci. USA 2005, 102, 5570–5575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hennig, T.; Prusty, A.B.; Kaufer, B.B.; Whisnant, A.W.; Lodha, M.; Enders, A.; Thomas, J.; Kasimir, F.; Grothey, A.; Klein, T.; et al. Selective inhibition of miRNA processing by a herpesvirus-encoded miRNA. Nature 2022, 605, 539. [Google Scholar] [CrossRef]
- Li, Q.; Lowey, B.; Sodroski, C.; Krishnamurthy, S.; Alao, H.; Cha, H.; Chiu, S.; El-Diwany, R.; Ghany, M.G.; Liang, T.J. Cellular microRNA networks regulate host dependency of hepatitis C virus infection. Nat. Comm. 2017, 8, 1789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartel, D.P. Metazoan microRNAs. Cell 2018, 173, 20–51. [Google Scholar] [CrossRef] [Green Version]
- Levine, E.; Jacob, E.B.; Levine, H. Target-specific and global effectors in gene regulation by microRNA. Biophys. J. 2007, 93, L52–L54. [Google Scholar] [CrossRef] [Green Version]
- Shamir, M.; Bar-On, Y.; Phillips, R.; Milo, R. SnapShot: Timescales in cell biology. Cell 2016, 164, 1302. [Google Scholar] [CrossRef]
- Patra, T.; Bose, S.K.; Kwon, Y.-C.; Meyer, K.; Ray, R. Inhibition of p70 isoforms of S6K1 induces anoikis to prevent transformed human hepatocyte growth. Life Sci. 2021, 265, 118764. [Google Scholar] [CrossRef]
- Zhdanov, V.P. Virology from the perspective of theoretical colloid and interface science. Curr. Opin. Coll. Interf. Sci. 2021, 53, 101450. [Google Scholar] [CrossRef]
- Pasin, F.; Shan, H.; García, B.; Müller, M.; San León, D.; Ludman, M.; Fresno, D.H.; Fátyol, K.; Munné-Bosch, S.; Rodrigo, G.; et al. Abscisic acid connects phytohormone signaling with RNA metabolic pathways and promotes an antiviral response that is evaded by a self-controlled RNA virus. Plant Comm. 2020, 1, 100099. [Google Scholar] [CrossRef] [PubMed]
- Iyer-Biswas, S.; Crooks, G.E.; Scherer, N.F.; Dinner, A.R. Universality in stochastic exponential growth. Phys. Rev. Lett. 2014, 113, 028101. [Google Scholar] [CrossRef] [Green Version]
- Pandey, P.P.; Singh, H.; Jain, S. Exponential trajectories, cell size fluctuations, and the adder property in bacteria follow from simple chemical dynamics and division control. Phys. Rev. E 2020, 101, 062406. [Google Scholar] [CrossRef] [PubMed]
- Beentjes, C.H.L.; Perez-Carrasco, R.; Grima, R. Exact solution of stochastic gene expression models with bursting, cell cycle and replication dynamics. Phys. Rev. E 2020, 101, 032403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Z.; Grima, R. Analytical distributions for detailed models of stochastic gene expression in eukaryotic cells. Proc. Natl. Acad. Sci. USA 2020, 117, 4682–4692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dessalles, R.; Fromion, V.; Robert, P. Models of protein production along the cell cycle: An investigation of possible sources of noise. PLoS ONE 2020, 15, e0226016. [Google Scholar] [CrossRef] [Green Version]
- Zhdanov, V.P. Conditions of appreciable influence of microRNA on a large number of target mRNAs. Molec. BioSyst. 2009, 5, 638–643. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhdanov, V.P. Interplay of Cellular mRNA, miRNA and Viral miRNA during Infection of a Cell. Int. J. Mol. Sci. 2023, 24, 122. https://doi.org/10.3390/ijms24010122
Zhdanov VP. Interplay of Cellular mRNA, miRNA and Viral miRNA during Infection of a Cell. International Journal of Molecular Sciences. 2023; 24(1):122. https://doi.org/10.3390/ijms24010122
Chicago/Turabian StyleZhdanov, Vladimir P. 2023. "Interplay of Cellular mRNA, miRNA and Viral miRNA during Infection of a Cell" International Journal of Molecular Sciences 24, no. 1: 122. https://doi.org/10.3390/ijms24010122