
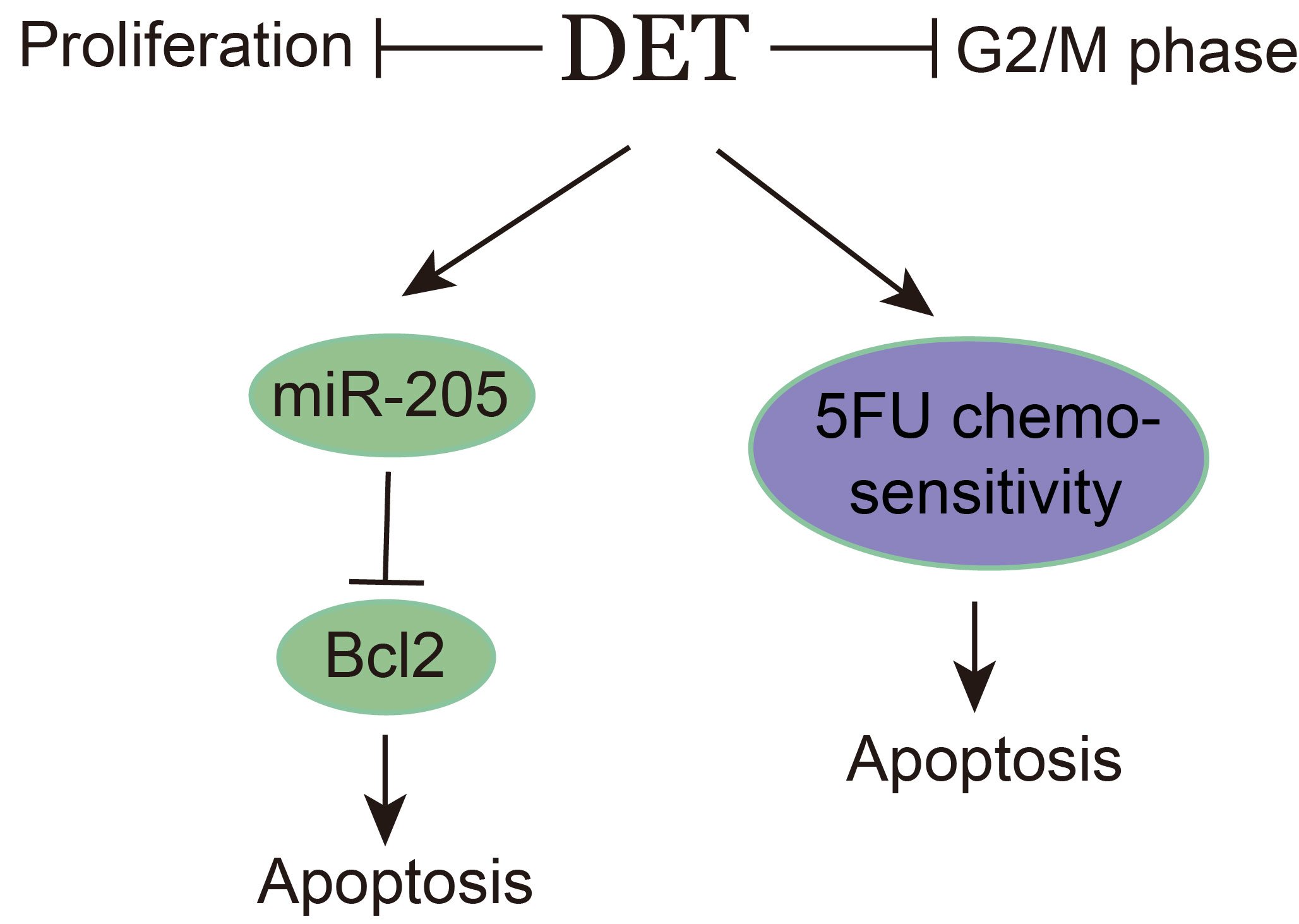
Deoxyelephantopin Induces Apoptosis and Enhances Chemosensitivity of Colon Cancer via miR-205/Bcl2 Axis

 , , ,
, , ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
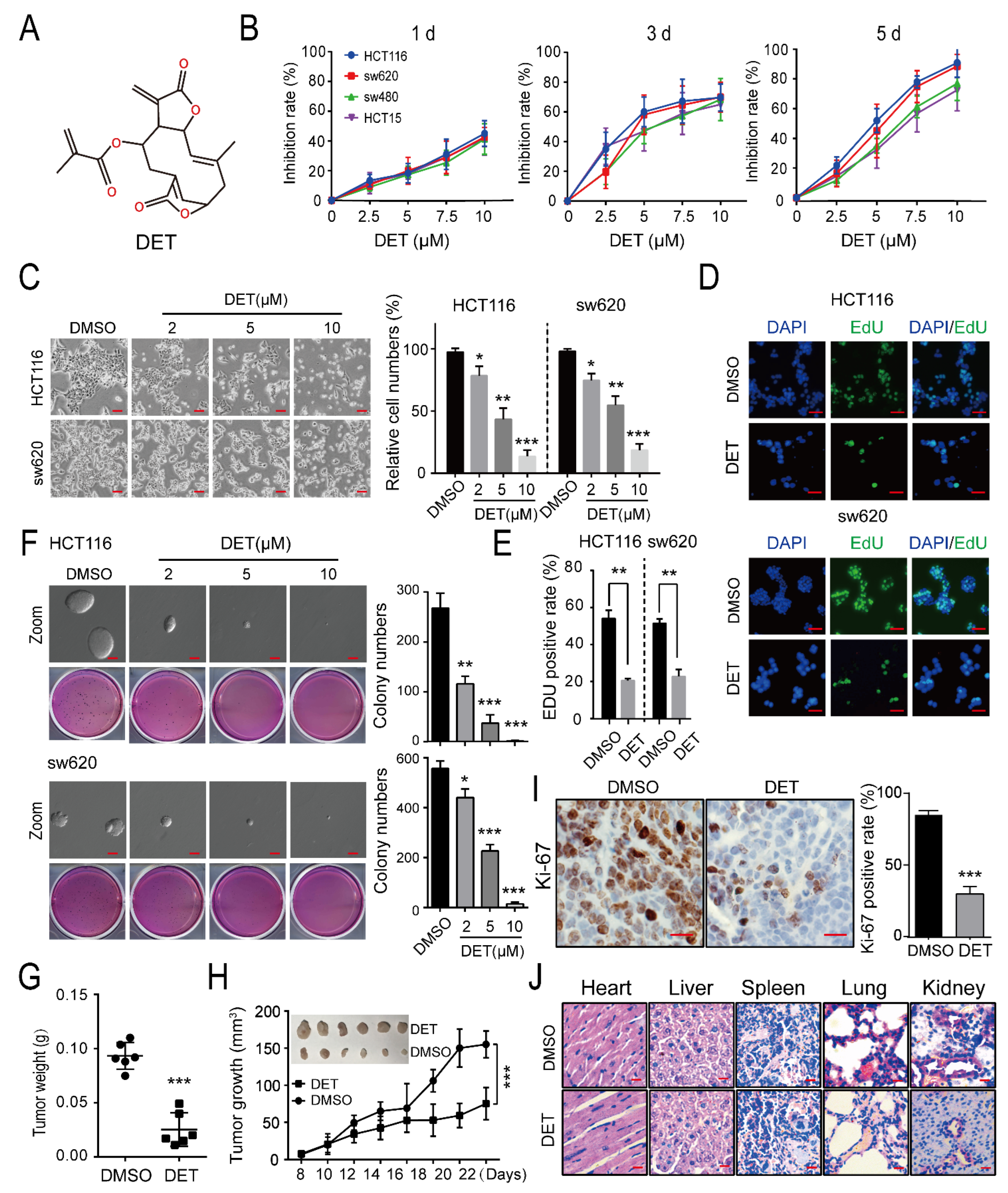
2.1. DET Inhibits Colon Cancer without Obvious Organ Toxicity
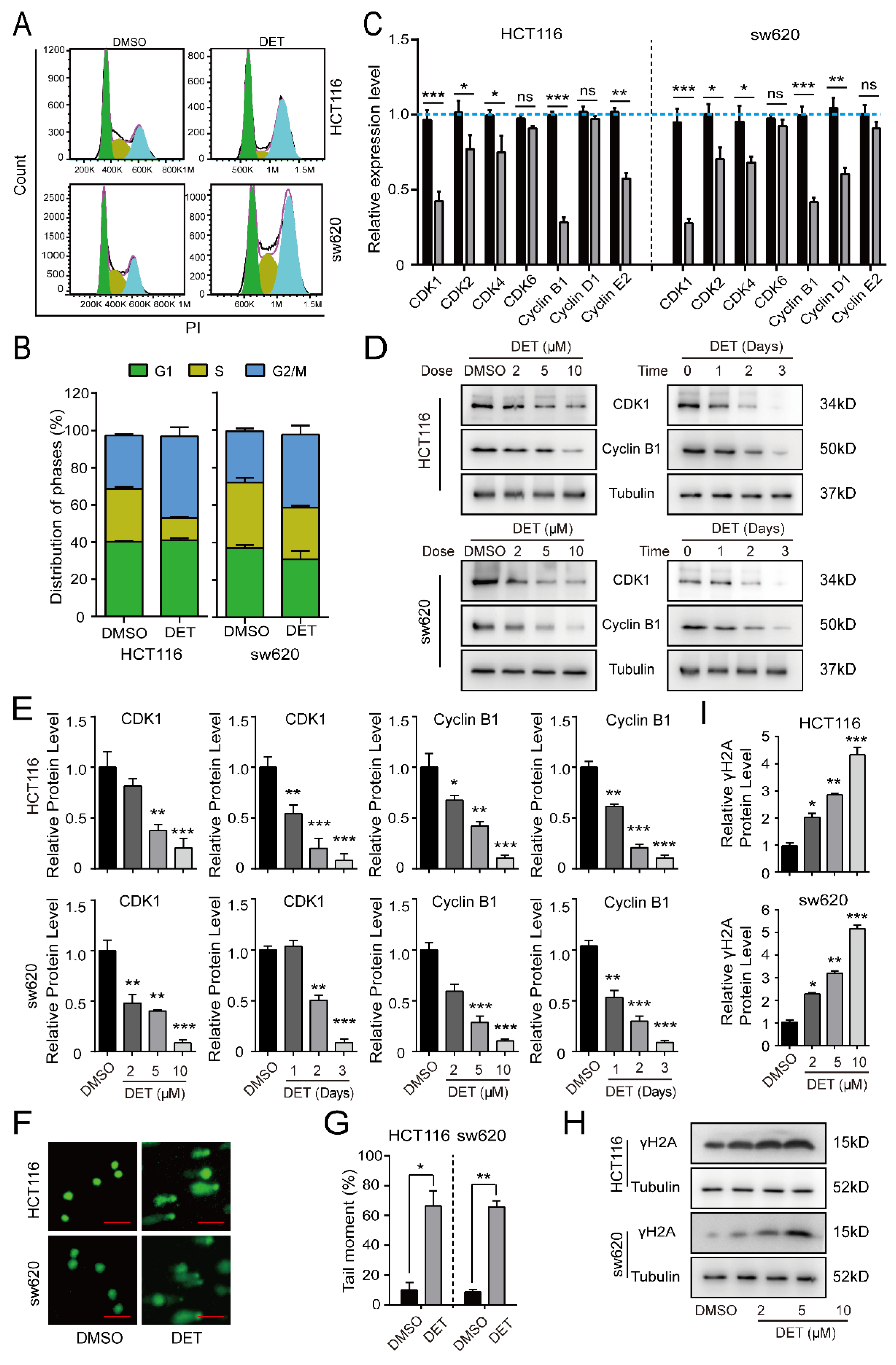
2.2. DET Arrests Colon Cancer Cells at the G2/M Phase by Inducing DNA Damage
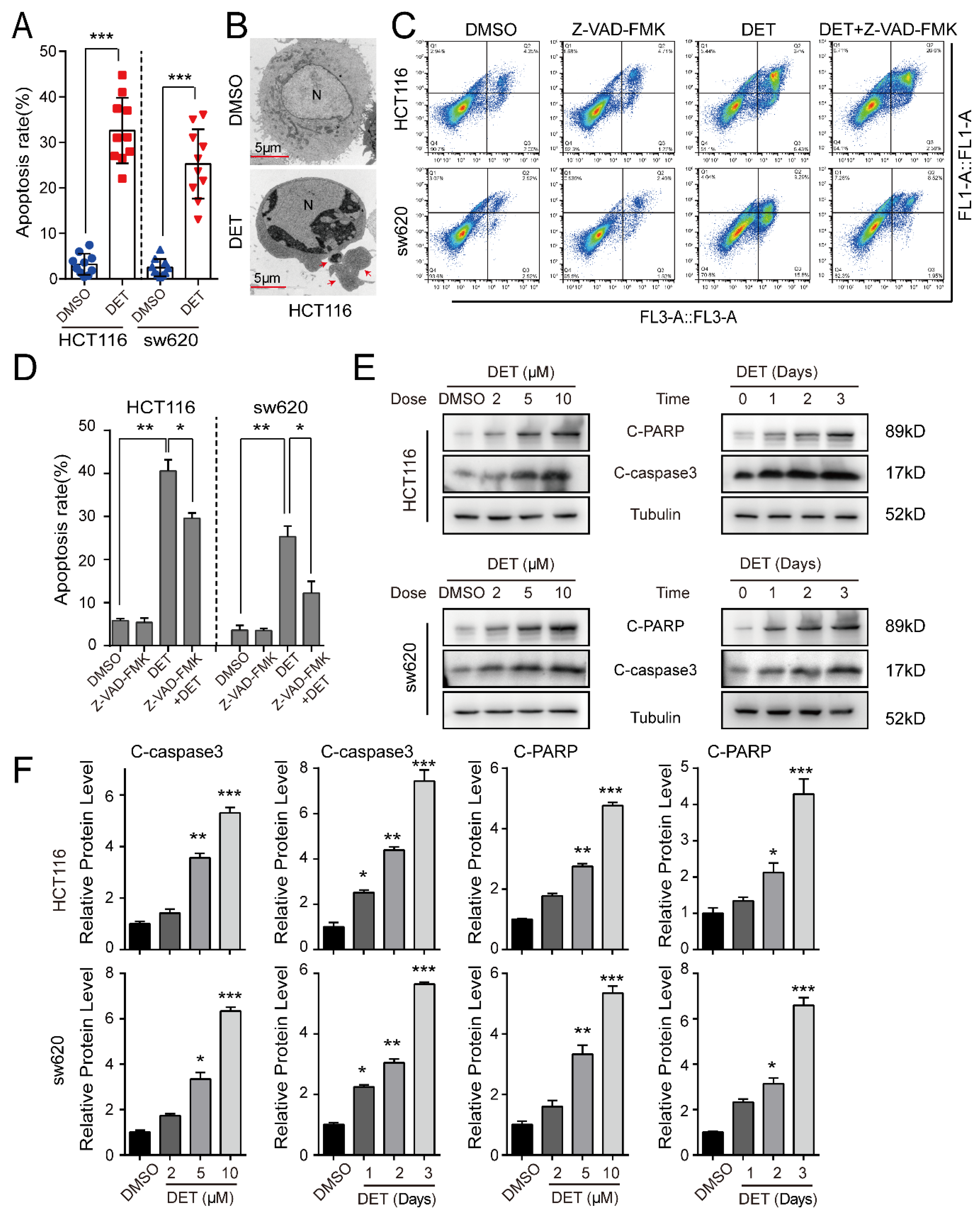
2.3. DET Induces Apoptosis in Colon Cancer Cells
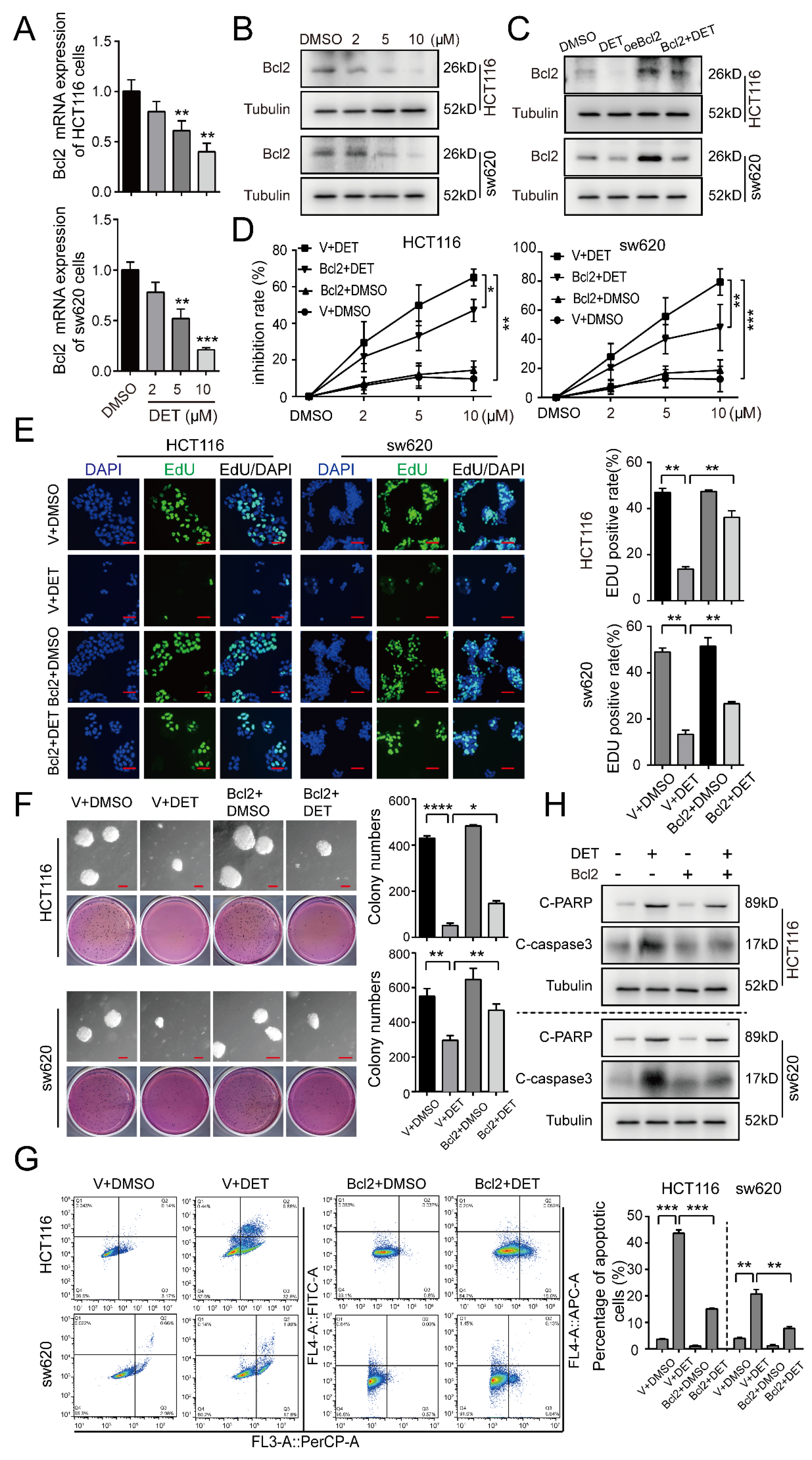
2.4. DET Induces Apoptosis by Inhibiting Bcl2 in Colon Cancer Cells
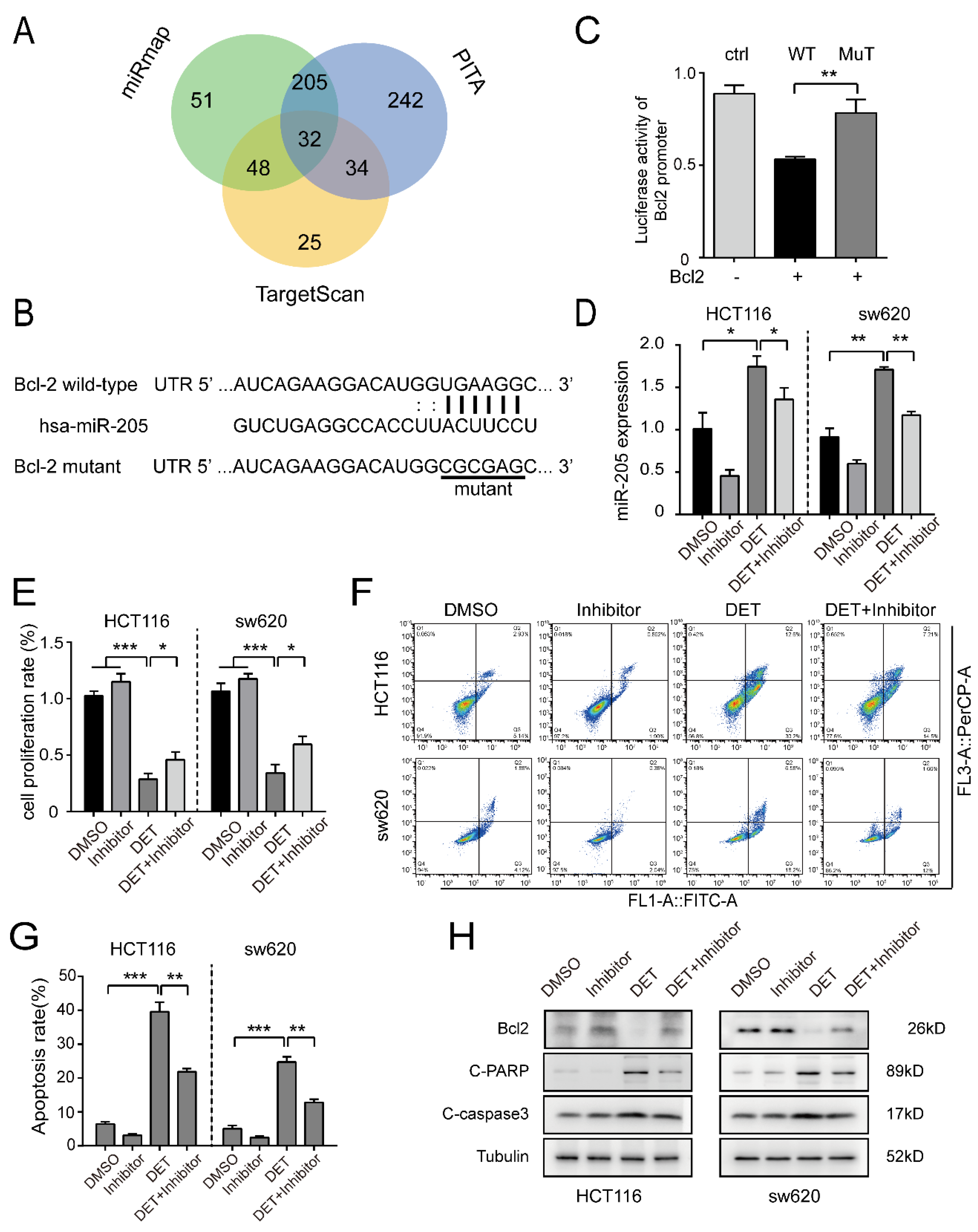
2.5. miR-205 Is Directly Targeted by Bcl2 and Induces the Apoptosis of Colon Cancer Cells
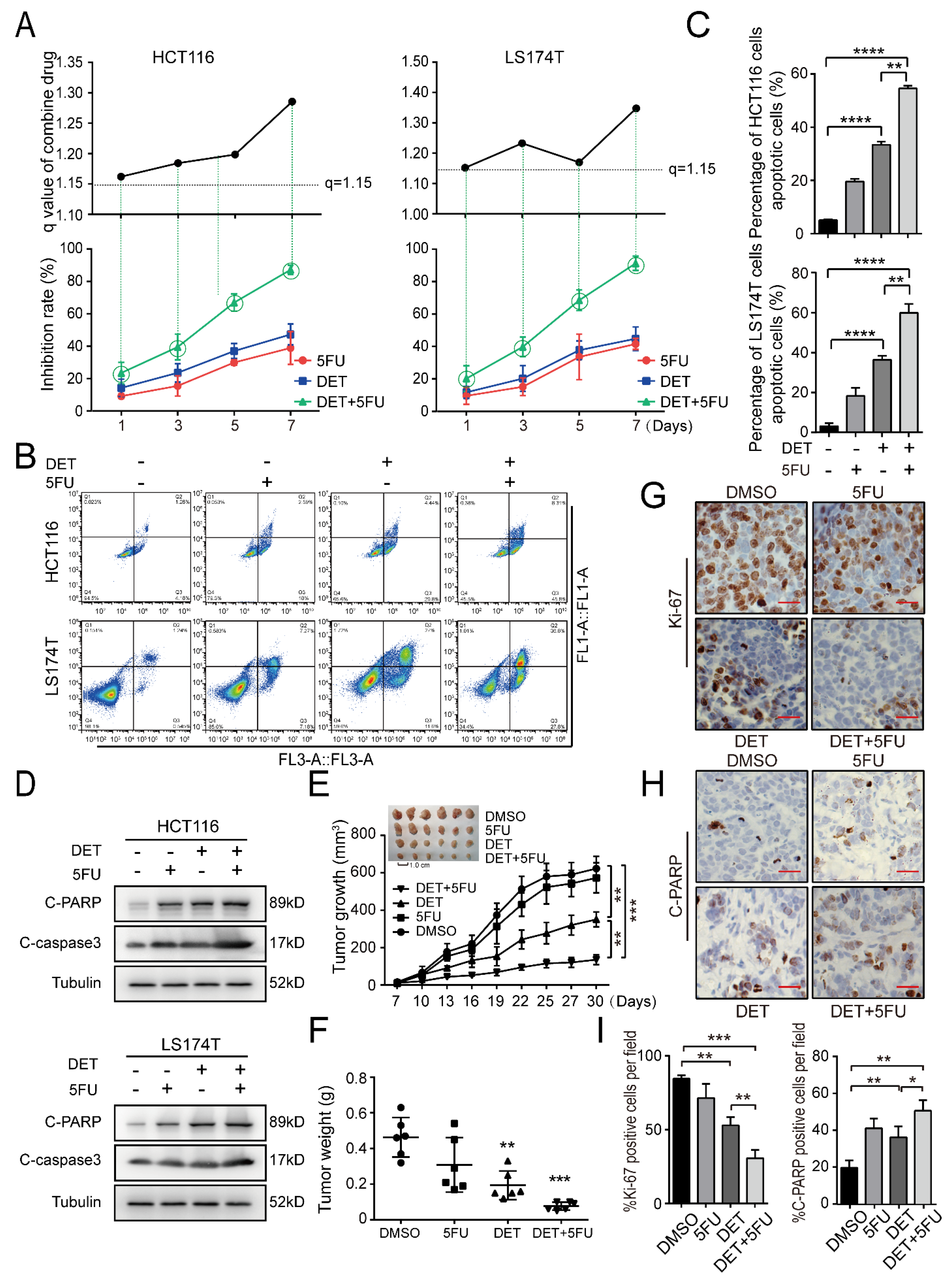
2.6. DET Enhances Chemosensitivity to 5-Fluorouracil (5FU) in Colon Cancer Cells
3. Discussion
4. Materials and Methods
4.1. Preparation of DET
4.2. Lentiviral Constructs and Infection
4.3. RNA Sequencing and qRT-PCR
4.4. Cell Proliferation Analysis
4.5. Detection of Phosphatidylserine Externalization by Annexin V and PI Staining
4.6. Western Blot Assay
4.7. EdU Staining
4.8. Soft Agar
4.9. Luciferase Reporter Assay
4.10. Animal Studies
4.11. Animal Ethics
4.12. Statistics Analysis
5. Conclusions
6. Patent
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| DET | Deoxyelephantopin |
| 5FU | 5-Fluorouracil |
| Bcl2 | B-cell lymphoma-2 |
| CC | Colon Cancer |
| IHC | Immunohistochemistry |
| MTT | 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| DMSO | Dimethyl sulfoxide |
References
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA A Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogino, S.; Galon, J.; Fuchs, C.S.; Dranoff, G. Cancer immunology—Analysis of host and tumor factors for personalized medicine. Nat. Rev. Clin. Oncol. 2011, 8, 711–719. [Google Scholar] [CrossRef] [PubMed]
- O’Kf, S.D. Diet, microorganisms and their metabolites, and colon cancer. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 691–706. [Google Scholar]
- Asmis, T.R.; Saltz, L. Systemic therapy for colon cancer. Gastroenterol. Clin. N. Am. 2008, 37, 287–295. [Google Scholar] [CrossRef]
- O’Connell, M.J. Oxaliplatin, fluorouracil, and leucovorin as adjuvant treatment for colon cancer. N. Engl. J. Med. 2004, 350, 2343–2351. [Google Scholar]
- Tournigand, C.; André, T.; Bonnetain, F.; Chibaudel, B.; Lledo, G.; Hickish, T.; Tabernero, J.; Boni, C.; Bachet, J.B.; Teixeira, L.; et al. Adjuvant therapy with fluorouracil and oxaliplatin in stage ii and elderly patients (between ages 70 and 75 years) with colon cancer: Subgroup analyses of the multicenter international study of oxaliplatin, fluorouracil, and leucovorin in the adjuvant trea. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2012, 30, 3353–3360. [Google Scholar] [CrossRef]
- Lee, K.H.; Cowherd, C.M.; Wolo, M.T. Antitumor agents. XV: Deoxyelephantopin, an antitumor principle from Elephantopus carolinianus Willd. J. Pharm. Sci. 1975, 64, 1572–1573. [Google Scholar]
- Farha, A.K.; Dhanya, S.R.; Mangalam, S.N.; Geetha, B.S.; Latha, P.G.; Remani, P. Deoxyelephantopin impairs growth of cervical carcinoma SiHa cells and induces apoptosis by targeting multiple molecular signaling pathways. Cell Biol. Toxicol. 2014, 30, 331–341. [Google Scholar] [CrossRef]
- Mehmood, T.; Maryam, A.; Zhang, H.; Li, Y.; Khan, M.; Ma, T. Deoxyelephantopin induces apoptosis in HepG2 cells via oxidative stress, NF-κB inhibition and mitochondrial dysfunction. Biofactors 2017, 43, 63–72. [Google Scholar] [CrossRef]
- Shiau, J.Y.; Chang, Y.Q.; Nakagawa-Goto, K.; Lee, K.H.; Shyur, L.F. Phytoagent deoxyelephantopin and its derivative inhibit triple negative breast cancer cell activity through ros-mediated exosomal activity and protein functions. Front. Pharmacol. 2017, 8, 398. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.C.; Lo, C.P.; Chiu, C.Y.; Shyur, L.F. Deoxyelephantopin, a novel multifunctional agent, suppresses mammary tumour growth and lung metastasis and doubles survival time in mice. Br. J. Pharmacol. 2010, 159, 856–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuge, W.; Chen, R.; Vladimir, K.; Dong, X.; Zia, K.; Sun, X.; Dai, X.; Bao, M.; Shen, X.; Liang, G. Costunolide specifically binds and inhibits thioredoxin reductase 1 to induce apoptosis in colon cancer. Cancer Lett. 2018, 412, 46–58. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Wang, H.; Niu, J.; Luo, M.; Gou, Y.; Miao, L.; Zou, Z.; Cheng, Y. Induction of ROS overload by alantolactone prompts oxidative DNA damage and apoptosis in colorectal cancer cells. Int. J. Mol. Sci. 2016, 17, 558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salla, M.; Fakhoury, I.; Saliba, N.; Darwiche, N.; Gali-Muhtasib, H. Synergistic anticancer activities of the plant-derived sesquiterpene lactones salograviolide A and iso-seco-tanapartholide. J. Nat. Med. 2013, 67, 468–479. [Google Scholar] [CrossRef]
- Hong, L.; Chen, J.; Wu, F.; Wu, F.; Shen, X.; Zheng, P.; Shao, R.; Lu, K.; Liu, Z.; Chen, D.; et al. Isodeoxyelephantopin Inactivates thioredoxin reductase 1 and activates ROS-Mediated JNK signaling pathway to exacerbate cisplatin effectiveness in human colon cancer cells. Front. Cell Dev. Biol. 2020, 8, 580517. [Google Scholar]
- Zhang, S.; Won, Y.K.; Ong, C.N.; Shen, H.M. Anti-cancer potential of sesquiterpene lactones: Bioactivity and molecular mechanisms. Curr. Med. Chem.-Anti-Cancer Agents 2005, 5, 239–249. [Google Scholar] [CrossRef]
- Liu, X.N.; Li, H.M.; Wang, S.P.; Zhang, J.Z.; Liu, D.L. Sesquiterpene lactones of Aucklandia lappa:Pharmacology, pharmacokinetics, toxicity, and structure-activity relationship. Chin. Herb. Med. 2021, 13, 167–176. [Google Scholar] [CrossRef]
- Gach, K.; Długosz, A.; Janecka, A. The role of oxidative stress in anticancer activity of sesquiterpene lactones. Arch. Für Exp. Pathol. Und Pharmakol. 2015, 388, 477–486. [Google Scholar] [CrossRef]
- Mathema, V.B.; Koh, Y.S.; Thakuri, B.C.; Sillanpää, M. Parthenolide, a sesquiterpene lactone, expresses multiple anti-cancer and anti-inflammatory activities. Inflammation 2012, 35, 560–565. [Google Scholar] [CrossRef]
- Huang, X.; Awano, Y.; Maeda, E.; Asada, Y.; Takemoto, H.; Watanabe, T.; Kojima-Yuasa, A.; Kobayashi, Y. Cytotoxic activity of two natural sesquiterpene lactones, isobutyroylplenolin and arnicolide D, on human colon cancer cell line HT-29. Nat. Prod. Res. 2014, 28, 914–916. [Google Scholar] [CrossRef]
- Chan, C.K.; Chan, G.; Awang, K.; Abdul Kadir, H. Deoxyelephantopin from elephantopus scaber inhibits HCT116 human colorectal carcinoma cell growth through apoptosis and cell cycle arrest. Molecules 2016, 21, 385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Jaarsveld, M.T.M.; Deng, D.; Ordoñez-Rueda, D.; Paulsen, M.; Wiemer, E.A.C.; Zi, Z. Cell-type-specific role of CHK2 in mediating DNA damage-induced G2 cell cycle arrest. Oncogenesis 2020, 9, 35. [Google Scholar] [CrossRef] [PubMed]
- Ling, Y.H.; el-Naggar, A.K.; Priebe, W.; Perez-Soler, R. Cell cycle-dependent cytotoxicity, G2/M phase arrest, and disruption of p34cdc2/cyclin B1 activity induced by doxorubicin in synchronized P388 cells. Mol. Pharmacol. 1996, 49, 832–841. [Google Scholar] [PubMed]
- Samuel, T.; Weber, H.O.; Funk, J.O. Linking DNA damage to cell cycle checkpoints. Cell Cycle 2002, 1, 162–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wanzel, M.; Kleine-Kohlbrecher, D.; Herold, S.; Hock, A.; Berns, K.; Park, J.; Hemmings, B.; Eilers, M. Akt and 14-3-3eta regulate Miz1 to control cell-cycle arrest after DNA damage. Nat. Cell Biol. 2005, 7, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Venit, T.; Semesta, K.; Farrukh, S.; Endara-Coll, M.; Havalda, R.; Hozak, P.; Percipalle, P. Nuclear myosin 1 activates p21 gene transcription in response to DNA damage through a chromatin-based mechanism. Commun. Biol. 2020, 3, 115. [Google Scholar] [CrossRef]
- Lawrence, K.S.; Chau, T.; Engebrecht, J. DNA damage response and spindle assembly checkpoint function throughout the cell cycle to ensure genomic integrity. PLoS Genet. 2015, 11, e1005150. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; McLeod, H.L.; Cassidy, J.; Collie-Duguid, E.S. Mechanisms of acquired chemoresistance to 5-fluorouracil and tomudex: Thymidylate synthase dependent and independent networks. Cancer Chemother. Pharmacol. 2007, 59, 839–845. [Google Scholar] [CrossRef]
- Wei, Y.; Yang, P.; Cao, S.; Zhao, L. The combination of curcumin and 5-fluorouracil in cancer therapy. Arch. Pharmacal Res. 2018, 41, 1–13. [Google Scholar] [CrossRef]
- Chan, C.K.; Supriady, H.; Goh, B.H.; Kadir, H.A. Elephantopus scaber induces apoptosis through ROS-dependent mitochondrial signaling pathway in HCT116 human colorectal carcinoma cells. J. Ethnopharmacol. 2015, 168, 291–304. [Google Scholar] [CrossRef]
- Koe, X.F.; Lim, E.L.; Seah, T.C.; Amanah, A.; Wahab, H.A.; Adenan, M.I.; Sulaiman, S.F.; Tan, M.L. Evaluation of in vitro cytochrome P450 induction and inhibition activity of deoxyelephantopin, a sesquiterpene lactone from Elephantopus scaber L. Food Chem. Toxicol. 2013, 60, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Su, M.; Chung, H.Y.; Li, Y. Deoxyelephantopin from Elephantopus scaber L. induces cell-cycle arrest and apoptosis in the human nasopharyngeal cancer CNE cells. Biochem. Biophys. Res. Commun. 2011, 411, 342–347. [Google Scholar] [CrossRef] [PubMed]
- Kabeer, F.A.; Sreedevi, G.B.; Nair, M.S.; Rajalekshmi, D.S.; Gopalakrishnan, L.P.; Kunjuraman, S.; Prathapan, R. Antineoplastic effects of deoxyelephantopin, a sesquiterpene lactone from Elephantopus scaber, on lung adenocarcinoma (A549) cells. J. Integr. Med. 2013, 11, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Geetha, B.S.; Nair, M.S.; Latha, P.G.; Remani, P. Sesquiterpene lactones isolated from Elephantopus scaber L. inhibits human lymphocyte proliferation and the growth of tumour cell lines and induces apoptosis in vitro. J. Biomed. Biotechnol. 2012, 2012, 721285. [Google Scholar] [CrossRef] [Green Version]
- Zou, J.; Zhang, Y.; Sun, J.; Wang, X.; Tu, H.; Geng, S.; Liu, R.; Chen, Y.; Bi, Z. Deoxyelephantopin Induces Reactive Oxygen Species-Mediated Apoptosis and Autophagy in Human Osteosarcoma Cells. Cell. Physiol. Biochem. 2017, 42, 1812–1821. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Wang, L.; Rodriguez-Aguayo, C.; Yuan, Y.; Debeb, B.G.; Chen, D.; Sun, Y.; You, M.J.; Liu, Y.; Dean, D.C.; et al. miR-205 acts as a tumour radiosensitizer by targeting ZEB1 and Ubc13. Nat. Commun. 2014, 5, 5671. [Google Scholar] [CrossRef] [Green Version]
- El Bezawy, R.; Tinelli, S.; Tortoreto, M.; Doldi, V.; Zuco, V.; Folini, M.; Stucchi, C.; Rancati, T.; Valdagni, R.; Gandellini, P.; et al. miR-205 enhances radiation sensitivity of prostate cancer cells by impairing DNA damage repair through PKCε and ZEB1 inhibition. J. Exp. Clin. Cancer Res. 2019, 38, 51. [Google Scholar] [CrossRef]
- Alla, V.; Kowtharapu, B.S.; Engelmann, D.; Emmrich, S.; Schmitz, U.; Steder, M.; Pützer, B.M. E2F1 confers anticancer drug resistance by targeting ABC transporter family members and Bcl-2 via the p73/DNp73-miR-205 circuitry. Cell Cycle 2012, 11, 3067–3078. [Google Scholar] [CrossRef] [Green Version]
- Juin, P.; Geneste, O.; Gautier, F.; Depil, S.; Campone, M. Decoding and unlocking the BCL-2 dependency of cancer cells. Nat. Rev. Cancer 2013, 13, 455–465. [Google Scholar] [CrossRef]
- Certo, M.; Del Gaizo Moore, V.; Nishino, M.; Wei, G.; Korsmeyer, S.; Armstrong, S.A.; Letai, A. Mitochondria primed by death signals determine cellular addiction to antiapoptotic BCL-2 family members. Cancer Cell 2006, 9, 351–365. [Google Scholar] [CrossRef] [Green Version]
- Beroukhim, R.; Mermel, C.H.; Porter, D.; Wei, G.; Raychaudhuri, S.; Donovan, J.; Barretina, J.; Boehm, J.S.; Dobson, J.; Urashima, M.; et al. The landscape of somatic copy-number alteration across human cancers. Nature 2010, 463, 899–905. [Google Scholar] [CrossRef] [PubMed]
- Iorio, M.V.; Croce, C.M. MicroRNA dysregulation in cancer: Diagnostics, monitoring and therapeutics. A comprehensive review. EMBO Mol. Med. 2012, 4, 143–159. [Google Scholar] [CrossRef] [PubMed]
- Rupaimoole, R.; Slack, F.J. MicroRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef] [PubMed]
- Shen, R.; Cai, X.; Shen, D.; Zhang, R.; Zhang, W.; Zhang, Y.; Li, Y.; Wang, A.; Zeng, Y.; Zhu, J.; et al. Long noncoding RNA LINC00518 contributes to proliferation and metastasis in lung adenocarcinoma via the miR-335-3p/CTHRC1 Axis. Cell Death Discov. 2022, 8, 98. [Google Scholar] [CrossRef]
- Cvetanova, B.; Li, M.Y.; Yang, C.C.; Hsiao, P.W.; Yang, Y.C.; Feng, J.H.; Shen, Y.C.; Nakagawa-Goto, K.; Lee, K.H.; Shyur, L.F. Sesquiterpene lactone deoxyelephantopin isolated from elephantopus scaber and its derivative DETD-35 suppress BRAFV600E mutant melanoma lung metastasis in mice. Int. J. Mol. Sci. 2021, 22, 3226. [Google Scholar] [CrossRef]
- Feng, J.H.; Nakagawa-Goto, K.; Lee, K.H.; Shyur, L.F. A novel plant sesquiterpene lactone derivative, DETD-35, suppresses BRAFV600E mutant melanoma growth and overcomes acquired vemurafenib resistance in mice. Mol. Cancer Ther. 2016, 15, 1163–1176. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.C.; Lin, K.J.; Cheng, Y.W.; Hsu, C.A.; Yang, S.S.; Shyur, L.F. Hepatoprotective effect and mechanistic insights of deoxyelephantopin, a phyto-sesquiterpene lactone, against fulminant hepatitis. J. Nutr. Biochem. 2013, 24, 516–530. [Google Scholar] [CrossRef]
- Peters, G.J.; Backus, H.H.J.; Freemantle, S.; Van Triest, B.; Codacci-Pisanelli, G.; Van der Wilt, C.L.; Smid, K.; Lunec, J.; Calvert, A.H.; Marsh, S.; et al. Induction of thymidylate synthase as a 5-fluorouracil resistance mechanism. Biochim. Biophys. Acta 2002, 1587, 194–205. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; He, J.; Li, J.; Peng, X.; Wang, X.; Dong, Z.; Zhao, E.; Liu, Y.; Wu, Z.; Cui, H. Demethylzeylasteral inhibits cell proliferation and induces apoptosis through suppressing MCL1 in melanoma cells. Cell Death Dis. 2017, 8, e3133. [Google Scholar] [CrossRef] [Green Version]
- Yang, R.; Wu, Y.; Wang, M.; Sun, Z.; Zou, J.; Zhang, Y.; Cui, H. HDAC9 promotes glioblastoma growth via TAZ-mediated EGFR pathway activation. Oncotarget 2015, 6, 7644. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Fu, G.; Pan, G.; Li, C.; Shen, L.; Hu, R.; Zhu, S.; Chen, Y.; Cui, H. Demethylzeylasteral inhibits glioma growth by regulating the miR-30e-5p/MYBL2 axis. Cell Death Dis. 2018, 9, 1035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Q.; Hou, J.; Feng, L.; Lv, A.; Ke, X.; Liang, H.; Wang, F.; Zhang, K.; Chen, K.; Cui, H. PHF19 promotes the proliferation, migration, and chemosensitivity of glioblastoma to doxorubicin through modulation of the SIAH1/β-catenin axis. Cell Death Dis. 2018, 9, 1049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; He, J.; Li, Y.; Xu, M.; Peng, X.; Mao, J.; Xu, B.; Cui, H. PHF14 promotes cell proliferation and migration through the AKT and ERK1/2 pathways in gastric cancer cells. BioMed Res. Int. 2020, 2020, 6507510. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Lv, A.; Deng, Q.; Zhang, G.; Hu, X.; Cui, H. TROP2 promotes the proliferation and metastasis of glioblastoma cells by activating the JAK2/STAT3 signaling pathway. Oncol. Rep. 2019, 41, 753–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; He, J.; Li, Y.; Lv, S.; Cui, H. NUSAP1 potentiates chemoresistance in glioblastoma through its SAP domain to stabilize ATR. Signal Transduct. Target. Ther. 2020, 5, 44. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, H.; Zhang, K.; Pan, G.; Li, C.; Li, C.; Hu, X.; Yang, L.; Cui, H. Deoxyelephantopin Induces Apoptosis and Enhances Chemosensitivity of Colon Cancer via miR-205/Bcl2 Axis. Int. J. Mol. Sci. 2022, 23, 5051. https://doi.org/10.3390/ijms23095051
Ji H, Zhang K, Pan G, Li C, Li C, Hu X, Yang L, Cui H. Deoxyelephantopin Induces Apoptosis and Enhances Chemosensitivity of Colon Cancer via miR-205/Bcl2 Axis. International Journal of Molecular Sciences. 2022; 23(9):5051. https://doi.org/10.3390/ijms23095051
Chicago/Turabian StyleJi, Haoyan, Kui Zhang, Guangzhao Pan, Changhong Li, Chongyang Li, Xin Hu, Liqun Yang, and Hongjuan Cui. 2022. "Deoxyelephantopin Induces Apoptosis and Enhances Chemosensitivity of Colon Cancer via miR-205/Bcl2 Axis" International Journal of Molecular Sciences 23, no. 9: 5051. https://doi.org/10.3390/ijms23095051