
Identification of Zinc Efficiency-Associated Loci (ZEALs) and Candidate Genes for Zn Deficiency Tolerance of Two Recombination Inbred Line Populations in Maize


Abstract
:1. Introduction
2. Results
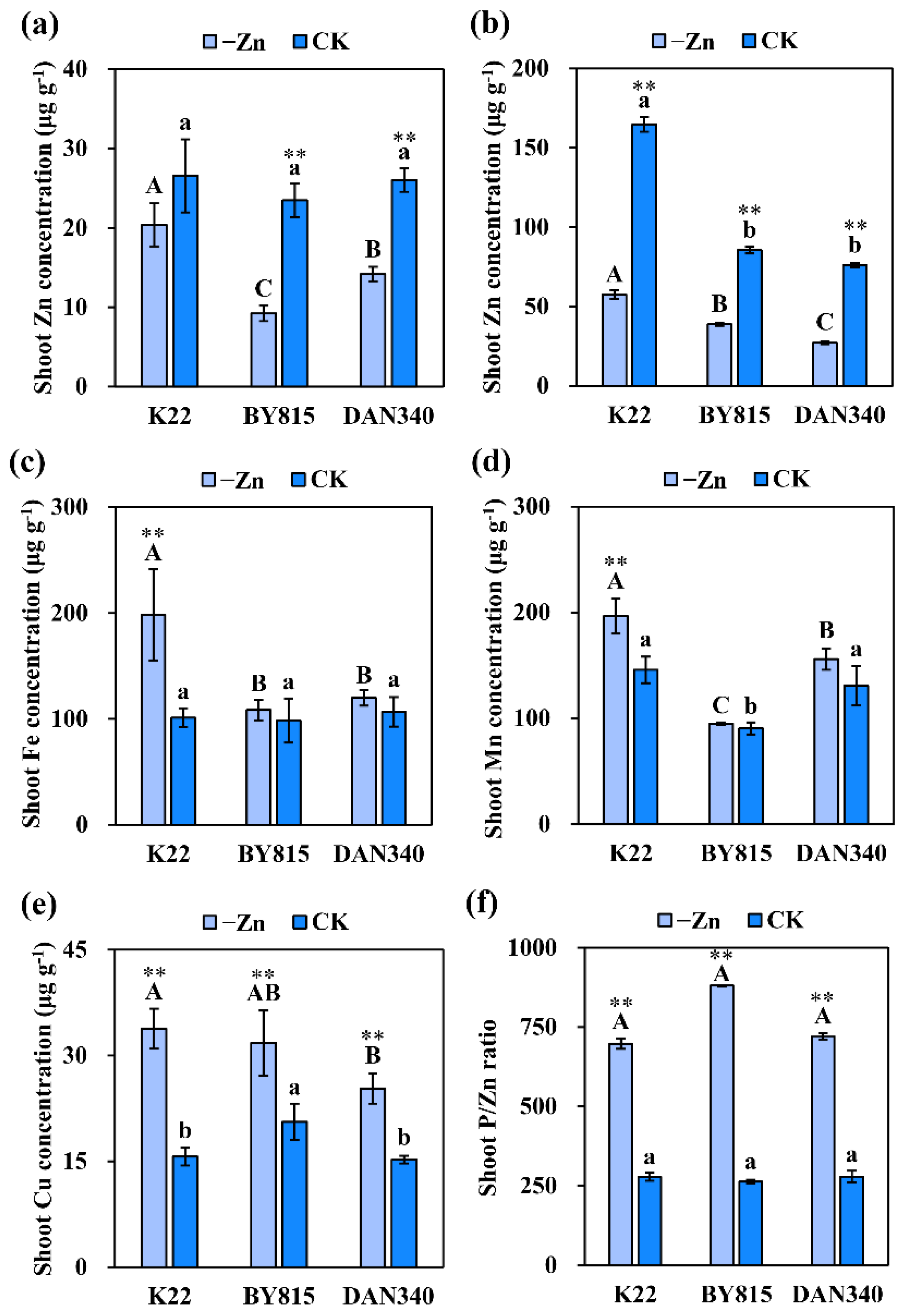
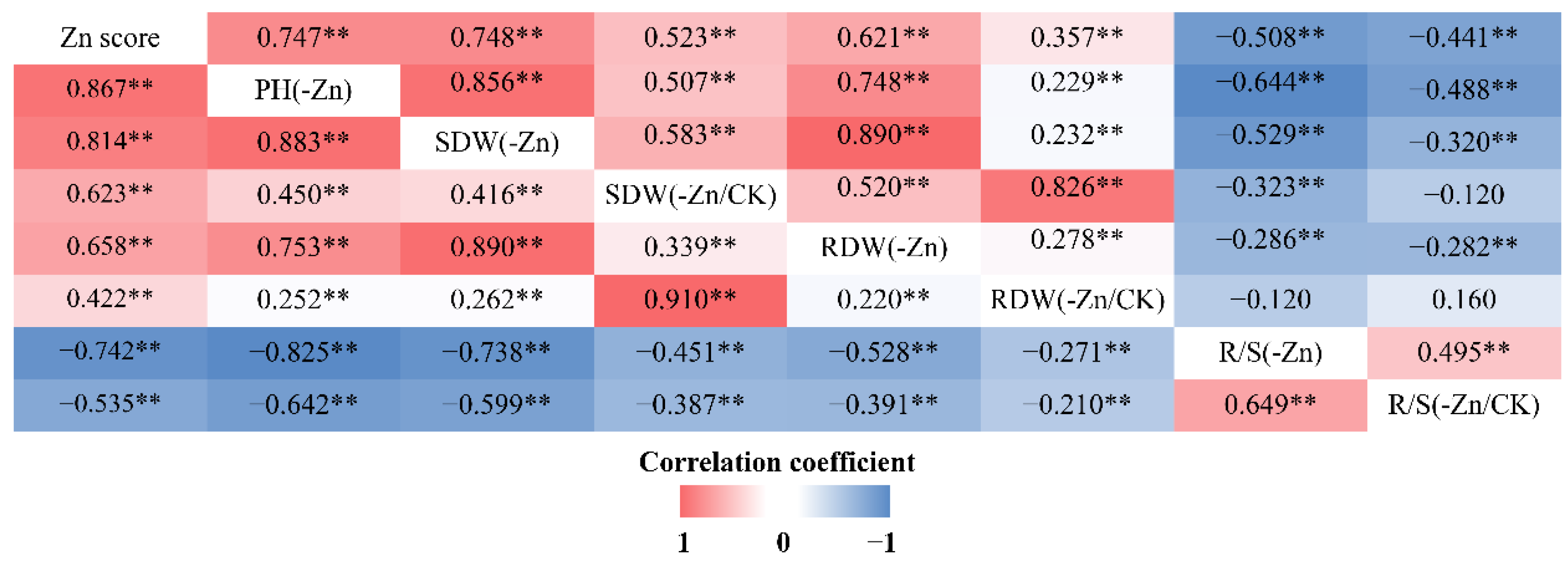
2.1. Phenotypic Variation in Zn Deficiency Tolerance
2.2. QTL Detection
2.3. QTL Colocalization and Candidate Genes Identification
3. Discussion
3.1. Physiological Mechanisms Underlying ZE
3.2. Comparisons of QTLs Identified in this Study with Previous Reports
3.3. Candidate Genes Associated with Zn Uptake and Transport
3.4. Candidate Genes Linked with Hormone Signaling
4. Materials and Methods
4.1. RIL Population
4.2. Plant Culture in Hydroponics
4.3. Phenotyping Methods
4.4. Statistical Analysis
4.5. QTL Mapping
4.6. Annotation of Candidate Genes
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alloway, B.J. Zinc in Soils and Crop Nutrition, 2nd ed.; IZA Publications: Brussels, Belgium; Paris, France, 2008. [Google Scholar]
- Cakmak, I. Enrichment of cereal grains with zinc: Agronomic or genetic biofortification? Plant Soil 2008, 302, 1–17. [Google Scholar] [CrossRef]
- Impa, S.M.; Johnson-Beebout, S.E. Mitigating zinc deficiency and achieving high grain Zn in rice through integration of soil chemistry and plant physiology research. Plant Soil 2012, 361, 3–41. [Google Scholar] [CrossRef]
- Maqbool, M.A.; Beshir, A.R. Zinc biofortification of maize (Zea mays L.): Status and challenges. Plant Breed. 2019, 138, 1–28. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Liang, Z.; He, X.; Wang, X.; Shi, X.; Zou, C.; Chen, X. The effects of controlled release urea on maize productivity and reactive nitrogen losses: A meta-analysis. Environ. Pollut. 2019, 246, 559–565. [Google Scholar] [CrossRef] [PubMed]
- CGIAR. CGIAR Strategy and Results Framework 2016–2030; CGIAR: Montpellier, France, 2016. [Google Scholar]
- Zhao, J.; Yang, X. Spatial patterns of yield-based cropping suitability and its driving factors in the three main maize-growing regions in China. Int. J. Biometeorol. 2019, 63, 1659–1668. [Google Scholar] [CrossRef] [PubMed]
- Grassini, P.; Eskridge, K.M.; Cassman, K.G. Distinguishing between yield advances and yield plateaus in historical crop production trends. Nat. Commun. 2013, 4, 2918. [Google Scholar] [CrossRef] [PubMed]
- FAOSTAT. Food and Agriculture Organization of the United Nations Statistics Division. 2014. Available online: http://faostat3.fao.org/browse/Q/QC/E (accessed on 15 May 2020).
- Sharma, P.N.; Chatterjee, C.; Agarwala, S.C.; Sharma, C.P. Zinc deficiency and pollen fertility in maize (Zea mays). Plant Soil 1990, 124, 221–225. [Google Scholar] [CrossRef]
- Serrago, R.A.; Alzueta, I.; Savin, R.; Slafer, G.A. Understanding grain yield responses to source-sink ratios during grain filling in wheat and barley under contrasting environments. Field Crops Res. 2013, 150, 42–51. [Google Scholar] [CrossRef] [Green Version]
- Alonso-Blanco, C.; Aarts, M.G.M.; Bentsink, L.; Keurentjes, J.J.B.; Reymond, M.; Vreugdenhil, D.; Koornneef, M. What has natural variation taught us about plant development, physiology, and adaptation? Plant Cell 2009, 21, 1877–1896. [Google Scholar] [CrossRef] [Green Version]
- Prasanna, B.M.; Pixley, K.; Warburton, M.L.; Xie, C.X. Molecular marker-assisted breeding options for maize improvement in Asia. Mol. Breed. 2010, 26, 339–356. [Google Scholar] [CrossRef]
- Li, P.; Zhang, Y.; Yin, S.; Zhu, P.; Pan, T.; Xu, Y.; Wang, J.; Hao, D.; Fang, H.; Xu, C. QTL-by-environment interaction in the response of maize root and shoot traits to different water regimes. Front. Plant Sci. 2018, 9, 229. [Google Scholar] [CrossRef] [Green Version]
- Lonergan, P.F.; Pallotta, M.A.; Lorimer, M.; Paull, J.G.; Barker, S.J.; Graham, R.D. Multiple genetic loci for zinc uptake and distribution in barley (Hordeum vulgare). New Phytol. 2009, 184, 168–179. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.; Rengel, Z.; Mohammadi, S.A.; Ebadi-Segherloo, A.; Maqsood, M.A. Mapping QTL associated with remobilization of zinc from vegetative tissues into grains of barley (Hordeum vulgare). Plant Soil 2016, 399, 193–208. [Google Scholar] [CrossRef]
- Velu, G.; Tutus, Y.; Gomez-Becerra, H.F.; Hao, Y.; Demir, L.; Kara, R.; Crespo-Herrera, L.A.; Orhan, S.; Yazici, A.; Singh, R.P.; et al. QTL mapping for grain zinc and iron concentrations and zinc efficiency in a tetraploid and hexaploid wheat mapping populations. Plant Soil 2017, 411, 81–89. [Google Scholar] [CrossRef]
- Wissuwa, M.; Ismail, A.M.; Yanagihara, S. Effects of zinc deficiency on rice growth and genetic factors contributing to tolerance. Plant Physiol. 2006, 142, 731–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.S.; Sajise, A.G.C.; Gregorio, G.B.; Kretzschmar, T.; Ismail, A.M.; Wissuwa, M. Genetic dissection for zinc deficiency tolerance in rice using bi-parental mapping and association analysis. Theor. Appl. Genet. 2017, 130, 1903–1914. [Google Scholar] [CrossRef]
- Genc, Y.; Verbyla, A.P.; Torun, A.A.; Carmak, I.; Willsmore, K.; Wallwork, H.; McDonald, G.K. Quantitative trait loci analysis of zinc efficiency and grain zinc concentration in wheat using whole genome average interval mapping. Plant Soil 2009, 314, 49–66. [Google Scholar] [CrossRef]
- Gelin, J.R.; Forster, S.; Grafton, K.F.; Mcclean, P.E.; Rojas-Cifuentes, G.A. Analysis of seed zinc and other minerals in a recombinant inbred population of navy bean (Phaseolus vulgaris L.). Crop Sci. 2007, 47, 1361–1366. [Google Scholar] [CrossRef]
- Gu, R.; Chen, F.; Liu, B.; Wang, X.; Liu, J.; Li, P.; Pan, Q.; Pace, J.; Soomro, A.A.; Lübberstedt, T.; et al. Comprehensive phenotypic analysis and quantitative trait locus identification for grain mineral concentration, content, and yield in maize (Zea mays L.). Theor. Appl. Genet. 2015, 128, 1777–1789. [Google Scholar] [CrossRef] [Green Version]
- Jin, T.; Chen, J.; Zhu, L.; Zhao, Y.; Guo, J.; Huang, Y. Comparative mapping combined with homology-based cloning of the rice genome reveals candidate genes for grain zinc and iron concentration in maize. BMC Genet. 2015, 16, 17. [Google Scholar] [CrossRef] [Green Version]
- Hindu, V.; Palacios-Rojas, N.; Babu, R.; Suwarno, W.B.; Rashid, Z.; Usha, R.; Saykhedkar, G.R.; Nair, S.K. Identification and validation of genomic regions influencing kernel zinc and iron in maize. Theor. Appl. Genet. 2018, 131, 1443–1457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Wang, X.; Zhu, H.; Yu, F. Identification and Analysis of Zinc Efficiency-Associated Loci in Maize. Front. Plant Sci. 2021, 12, 739282. [Google Scholar] [CrossRef] [PubMed]
- Eide, D.; Broderius, M.; Fett, J.; Guerinot, M.L. A novel iron-regulated metal transporter from plants identified by functional expression in yeast. Proc. Natl. Acad. Sci. USA 1996, 93, 5624–5628. [Google Scholar] [CrossRef] [Green Version]
- Vert, G.; Grotz, N.; Dédaldéchamp, F.; Gaymard, F.; Guerinot, M.L.; Briat, J.F.; Curie, C. IRT1, an Arabidopsis transporter essential for iron uptake from the soil and for plant growth. Plant Cell 2002, 14, 1223–1233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, C.K.E.; Jarvis, R.S.; Sherson, S.M.; Cobbett, C.S. Functional analysis of the heavy metal binding domains of the Zn/Cd-transporting ATPase, HMA2, in Arabidopsis thaliana. New Phytol. 2009, 181, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Ueno, D.; Yamaji, N.; Kono, I.; Huang, C.F.; Ando, T.; Yano, M.; Ma, J.F. Gene limiting cadmium accumulation in rice. Proc. Natl. Acad. Sci. USA 2010, 107, 16500–16505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, A.; Yamaji, N.; Ma, J.F. Overexpression of OsHMA3 enhances Cd tolerance and expression of Zn transporter genes in rice. J. Exp. Bot. 2014, 65, 6013–6021. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Wang, P.; Wang, P.; Yang, M.; Lian, X.; Tang, Z.; Huang, C.F.; Salt, D.E.; Zhao, F.J. A loss-of-function allele of OsHMA3 associated with high cadmium accumulation in shoots and grain of Japonica rice cultivars. Plant Cell Environ. 2016, 39, 1941–1954. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.L.; Gao, Z.Y.; Shang, L.G.; Yang, C.H.; Ruan, B.P.; Zeng, D.L.; Guo, L.B.; Zhao, F.J.; Huang, C.F.; Qian, Q. Natural variation in the promoter of OsHMA3 contributes to differential grain cadmium accumulation between Indica and Japonica rice. J. Integr. Plant Biol. 2020, 62, 314–329. [Google Scholar] [CrossRef] [Green Version]
- Pedas, P.; Schjoerring, J.K.; Husted, S. Identification and characterization of zinc-starvation-induced ZIP transporters from barley roots. Plant Physiol. Biochem. 2009, 47, 377–383. [Google Scholar] [CrossRef]
- Tiong, J.; McDonald, G.; Genc, Y.; Shirley, N.; Langridge, P.; Huang, C.Y. Increased expression of six ZIP family genes by zinc (Zn) deficiency is associated with enhanced uptake and root-to-shoot translocation of Zn in barley (Hordeum vulgare). New Phytol. 2015, 207, 1097–1109. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhou, X.; Huang, Y.; Zhu, L.; Zhang, S.; Zhao, Y.; Guo, J.; Chen, J.; Chen, R. Identification and characterization of the zinc-regulated transporters, iron-regulated transporter-like protein (ZIP) gene family in maize. BMC Plant Biol. 2013, 13, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khatun, M.A.; Hossain, M.M.; Bari, M.A.; Abdullahil, K.M.; Parvez, M.S.; Alam, M.F.; Kabir, A.H. Zinc deficiency tolerance in maize is associated with the up-regulation of Zn transporter genes and antioxidant activities. Plant Biol. 2018, 20, 765–770. [Google Scholar] [CrossRef] [PubMed]
- Mager, S.; Schönberger, B.; Ludewig, U. The transcriptome of zinc deficient maize roots and its relationship to DNA methylation loss. BMC Plant Biol. 2018, 18, 372. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Zhao, X.; Liu, Y.; Wang, Y.; Wu, W.; Jiang, Y.; Liao, C.; Xu, X.; Gao, S.; Shen, Y.; et al. Genome-wide identification of ZmHMAs and association of natural variation in ZmHMA2 and ZmHMA3 with leaf cadmium accumulation in maize. PeerJ 2019, 7, e7877. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Zhang, H.; Zhao, P.; Zhang, Z.; Liang, W.; Tian, Z.; Zheng, Y. Mining of candidate maize genes for nitrogen use efficiency by integrating gene expression and QTL data. Plant Mol. Biol. Rep. 2012, 30, 297–308. [Google Scholar] [CrossRef]
- Osman, K.A.; Tang, B.; Wang, Y.; Chen, J.; Yu, F.; Li, L.; Han, X.; Zhang, Z.; Yan, J.; Zheng, Y.; et al. Dynamic QTL analysis and candidate gene mapping for waterlogging tolerance at maize seedling stage. PLoS ONE 2013, 8, e79305. [Google Scholar] [CrossRef] [Green Version]
- Cakmak, I.; Torun, B.; Erenoglu, B.; Ozturk, L.; Marschner, H.; Kalayci, M.; Ekiz, H.; Yilmaz, A. Morphological and physiological differences in the response of cereals to zinc deficiency. Euphytica 1998, 100, 349–357. [Google Scholar] [CrossRef]
- Genc, Y.; McDonald, G.K.; Graham, R.D. Differential expression of zinc efficiency during the growing season of barley. Plant Soil 2004, 263, 273–282. [Google Scholar] [CrossRef]
- Genc, Y.; McDonald, G.K.; Graham, R.D. Contribution of different mechanisms to zinc deficiency in bread wheat during early vegetative stage. Plant Soil 2006, 281, 353–367. [Google Scholar] [CrossRef]
- Khan, H.R.; McDonald, G.K.; Rengel, Z. Chickpea genotypes differ in their sensitivity to Zn deficiency. Plant Soil 1998, 198, 11–18. [Google Scholar] [CrossRef]
- Sadeghzadeh, B.; Rengel, Z.; Li, C. Differential zinc efficiency of barley genotypes grown in soil and chelator-buffered nutrient solution. J. Plant Nutr. 2009, 32, 1744–1767. [Google Scholar] [CrossRef]
- Cakmak, I.; Sari, N.; Marschner, H.; Kalayci, M.; Yilmaz, A.; Eker, S.; Gülüt, K.Y. Dry matter production and distribution of zinc in bread and durum wheat genotypes differing in Zn efficiency. Plant Soil 1996, 180, 173–181. [Google Scholar] [CrossRef]
- Dong, B.; Rengel, Z.; Graham, R.D. Root morphology of wheat genotypes differing in zinc efficiency. J. Plant Nutr. 1995, 18, 2761–2773. [Google Scholar] [CrossRef]
- Rengel, Z.; Graham, R.D. Wheat genotypes differ in zinc efficiency when grown in the chelate-buffered nutrient solution. Plant Soil 1995, 176, 307–316. [Google Scholar] [CrossRef]
- Hacisalihoglu, G.; Hart, J.J.; Wang, Y.; Cakmak, I.; Kochian, L.V. Zinc efficiency is correlated with enhanced expression and activity of zinc-requiring enzymes in wheat. Plant Physiol. 2003, 131, 595–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Impa, S.M.; Morete, M.J.; Ismail, A.M.; Schulin, R.; Johnson-Beebout, S.E. Zn uptake, translocation and grain Zn loading in rice (Oryza sativa L.) genotypes selected for Zn deficiency tolerance and high grain Zn. J. Exp. Bot. 2013, 64, 2739–2751. [Google Scholar] [CrossRef]
- Rengel, Z. Physiological mechanisms underlying differential nutrient efficiency of crop genotypes. In Mineral Nutrition of Crops—Fundamental Mechanisms and Implications; Rengel, Z., Ed.; Food Products Press: New York, NY, USA, 1999. [Google Scholar]
- Hacisalihoglu, G.; Kochian, L. V How do some plants tolerate low levels of soil zinc? Mechanisms of zinc efficiency in crop plants. New Phytol. 2003, 159, 341–350. [Google Scholar] [CrossRef]
- White, P.J.; Broadley, M.R. Physiological limits to zinc biofortification of edible crops. Front. Plant Sci. 2011, 2, 80. [Google Scholar] [CrossRef] [Green Version]
- Erenoglu, B.; Nikolic, M.; Römheld, V.; Cakmak, I. Uptake and transport of foliar applied zinc (65Zn) in bread and durum wheat cultivars differing in zinc efficiency. Plant Soil 2002, 241, 251–257. [Google Scholar] [CrossRef]
- Rengel, Z.; Graham, R.D. Uptake of zinc from chelate-buffered nutrient solutions by wheat genotypes differing in zinc efficiency. J. Exp. Bot. 1996, 47, 217–226. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, D.; Li, C.; Cui, Z.; Chen, X.; Russell, Y.; Zou, C. Zinc accumulation and remobilization in winter wheat as affected by phosphorus application. Field Crops Res. 2015, 184, 155–161. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, D.; Liu, Y.; Cui, Z.; Chen, X.; Zou, C. Zinc uptake and accumulation in winter wheat relative to changes in root morphology and mycorrhizal colonization following varying phosphorus application on calcareous soil. Field Crops Res. 2016, 197, 74–82. [Google Scholar] [CrossRef]
- Rose, T.J.; Impa, S.M.; Rose, M.T.; Pariasca-Tanaka, J.; Mori, A.; Heuer, S.; Johnson-Beebout, S.E.; Wissuwa, M. Enhancing phosphorus and zinc acquisition efficiency in rice: A critical review of root traits and their potential utility in rice breeding. Ann. Bot. 2013, 112, 331–345. [Google Scholar] [CrossRef] [Green Version]
- Shahzad, Z.; Rouached, H.; Rakha, A. Combating mineral malnutrition through iron and zinc biofortification of cereals. Comprehen. Rev. Food Sci. Food Saf. 2014, 13, 329–346. [Google Scholar] [CrossRef]
- Huang, C.; Barker, S.J.; Langridge, P.; Smith, F.W.; Graham, R.D. Zinc deficiency up-regulates expression of high-affinity phosphate transporter genes in both phosphate-sufficient and -deficient barley roots. Plant Physiol. 2000, 124, 415–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, G.A.; Bouraine, S.; Wege, S.; Li, Y.; de Carbonnel, M.; Berthomieu, P.; Rouached, H.; Poirier, Y. Coordination between zinc and phosphate homeostasis involves the transcription factor PHR1, the phosphate exporter PHO1, and its homologue PHO1; H3 in Arabidopsis. J. Exp. Bot. 2014, 65, 871–884. [Google Scholar] [CrossRef] [Green Version]
- Qin, H.; Cai, Y.; Liu, Z.; Wang, G.; Wang, J.; Guo, Y.; Wang, H. Identification of QTL for zinc and iron concentration in maize kernel and cob. Euphytica 2012, 187, 345–358. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, J.; Jin, T.; Huang, Y.; Chen, J.; Zhu, L.; Zhao, Y.; Guo, J. Identification of quantitative trait locus and prediction of candidate genes for grain mineral concentration in maize across multiple environments. Euphytica 2017, 213, 90. [Google Scholar] [CrossRef]
- Ju, C.; Zhang, W.; Liu, Y.; Gao, Y.; Wang, X.; Yan, J.; Yang, X.; Li, J. Genetic analysis of seedling root traits reveals the association of root trait with other agronomic traits in maize. BMC Plant Biol. 2018, 18, 171. [Google Scholar] [CrossRef]
- Ma, L.; Qing, C.; Zhang, M.; Zou, C.; Pan, G.; Shen, Y. GWAS with a PCA uncovers candidate genes for accumulations of microelements in maize seedlings. Physiol. Plantarum 2021, 172, 2170–2180. [Google Scholar] [CrossRef]
- Azevedo, G.C.; Cheavegatti-Gianotto, A.; Negri, B.F.; Hufnagel, B.; Silva, L.D.C.E.; Magalhaes, J.V.; Garcia, A.A.; Lana, U.G.; de Sousa, S.M.; Guimaraes, C.T. Multiple interval QTL mapping and searching for PSTOL1 homologs associated with root morphology, biomass accumulation and phosphorus content in maize seedlings under low-P. BMC Plant Biol. 2015, 15, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Zhuang, Z.; Cai, H.; Cheng, S.; Soomro, A.A.; Liu, Z.; Gu, R.; Mi, G.; Yuan, L.; Chen, F. Use of genotype-environment interactions to elucidate the pattern of maize root plasticity to nitrogen deficiency. J. Integr. Plant Biol. 2016, 58, 242–253. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Zhao, Y.; Zhang, R.; Xing, J.; Duan, M.; Li, J.; Wang, N.; Wang, W.; Zhang, S.; Chen, Z.; et al. Mapping of a major QTL for salt tolerance of mature field-grown maize plants based on SNP markers. BMC Plant Biol. 2017, 17, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Li, Z.; Fang, H.; Zhang, M.; Duan, L. Analysis of the genetic basis of plant height-related traits in response to ethylene by QTL mapping in maize (Zea mays L.). PLoS ONE 2018, 13, e0193072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burton, A.L.; Johnson, J.; Foerster, J.; Hanlon, M.T.; Kaeppler, S.M.; Lynch, J.P.; Brown, K.M. QTL mapping and phenotypic variation of root anatomical traits in maize (Zea mays L.). Theor. Appl. Genet. 2015, 128, 93–106. [Google Scholar] [CrossRef]
- Li, S.; Zhou, X.; Li, H.; Liu, Y.; Zhu, L.; Guo, J.; Liu, X.; Fan, Y.; Chen, J.; Chen, R. Overexpression of ZmIRT1 and ZmZIP3 enhances iron and zinc accumulation in transgenic Arabidopsis. PLoS ONE 2015, 10, e0136647. [Google Scholar] [CrossRef]
- Rogers, E.E.; Eide, D.J.; Guerinot, M.L. Altered selectivity in an Arabidopsis metal transporter. Proc. Natl. Acad. Sci. USA 2000, 97, 12356–12360. [Google Scholar] [CrossRef] [Green Version]
- Pedas, P.; Ytting, C.K.; Fuglsang, A.T.; Jahn, T.P.; Schjoerring, J.K.; Husted, S. Manganese efficiency in barley: Identification and characterization of the metal ion transporter HvIRT1. Plant Physiol. 2008, 148, 455–466. [Google Scholar] [CrossRef] [Green Version]
- Korshunova, Y.O.; Eide, D.; Clark, W.G.; Guerinot, M.L.; Pakrasi, H.B. The IRT1 protein from Arabidopsis thaliana is a metal transporter with a broad substrate range. Plant Mol. Biol. 1999, 40, 37–44. [Google Scholar] [CrossRef]
- Lee, S.; An, G. Over-expression of OsIRT1 leads to increased iron and zinc accumulations in rice. Plant Cell Environ. 2009, 32, 408–416. [Google Scholar] [CrossRef] [PubMed]
- Morel, M.; Crouzet, J.; Gravot, A.; Auroy, P.; Leonhardt, N.; Vavasseur, A.; Richaud, P. AtHMA3, a P1B-ATPase allowing Cd/Zn/Co/Pb vacuolar storage in Arabidopsis. Plant Physiol. 2009, 149, 894–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, H.M.; Huang, S.; Che, J.; Yamaji, N.; Ma, J.F. The tonoplast-localized transporter OsHMA3 plays an important role in maintaining Zn homeostasis in rice. J. Exp. Bot. 2019, 70, 2717–2725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.F.; Ueno, D.; Zhao, F.J.; McGrath, S.P. Subcellular localisation of Cd and Zn in the leaves of a Cd-hyperaccumulating ecotype of Thlaspi caerulescens. Planta 2005, 220, 731–736. [Google Scholar] [CrossRef]
- Nevo, Y.; Nelson, N. The NRAMP family of metal-ion transporters. Biochim. Biophys. Acta-Mol. Cell Res. 2006, 1763, 609–620. [Google Scholar] [CrossRef] [Green Version]
- Bozzi, A.T.; Bane, L.B.; Weihofen, W.A.; McCabe, A.L.; Singharoy, A.; Chipot, C.J.; Schulten, K.; Gaudet, R. Conserved methionine dictates substrate preference in Nramp-family divalent metal transporters. Proc. Natl. Acad. Sci. USA 2016, 113, 10310–10315. [Google Scholar] [CrossRef] [Green Version]
- Weber, M.; Harada, E.; Vess, C.; Roepenack-Lahaye, E.V.; Clemens, S. Comparative microarray analysis of Arabidopsis thaliana and Arabidopsis halleri roots identifies nicotianamine synthase, a ZIP transporter and other genes as potential metal hyperaccumulation factors. Plant J. 2004, 37, 269–281. [Google Scholar] [CrossRef]
- Lanquar, V.; Lelièvre, F.; Barbier-Brygoo, H.; Thomine, S. Regulation and function of AtNRAMP4 metal transporter protein. Soil Sci. Plant Nutr. 2004, 50, 1141–1150. [Google Scholar] [CrossRef]
- Pottier, M.; Oomen, R.; Picco, C.; Giraudat, J.; Scholz-Starke, J.; Richaud, P.; Carpaneto, A.; Thomine, S. Identification of mutations allowing Natural Resistance Associated Macrophage Proteins (NRAMP) to discriminate against cadmium. Plant J. 2015, 83, 625–637. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Xu, Y.H.; Yi, H.Y.; Gong, J.M. Vacuolar membrane transporters OsVIT1 and OsVIT2 modulate iron translocation between flag leaves and seeds in rice. Plant J. 2012, 72, 400–410. [Google Scholar] [CrossRef]
- Rellan-Alvarez, R.; Abadia, J.; Alvarez-Fernandez, A. Formation of metal-nicotianamine complexes as affected by pH, ligand exchange with citrate and metal exchange. A study by electrospray ionization time-of-flight mass spectrometry. Rapid Commun. Mass. Spectrom. 2008, 22, 1553–1562. [Google Scholar] [CrossRef] [PubMed]
- Shojima, S.; Nishizawa, N.K.; Fushiya, S.; Nozoe, S.; Irifune, T.; Mori, S. Biosynthesis of phytosiderophores: In Vitro biosynthesis of 2’-deoxymugineic acid from l-methionine and nicotianamine. Plant Physiol. 1990, 93, 1497–1503. [Google Scholar] [CrossRef] [Green Version]
- Higuchi, K.; Suzuki, K.; Nakanishi, H.; Yamaguchi, H.; Nishizawa, N.K.; Mori, S. Cloning of nicotianamine synthase genes, novel genes involved in the biosynthesis of phytosiderophores. Plant Physiol. 1999, 119, 471–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, M.; Yamaguchi, H.; Nakanishi, H.; Shioiri, T.; Nishizawa, N.K.; Mori, S. Cloning two genes for nicotianamine aminotransferase, a critical enzyme in iron acquisition (strategy II) in graminaceous plants. Plant Physiol. 1999, 121, 947–956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bashir, K.; Inoue, H.; Nagasaka, S.; Takahashi, M.; Nakanishi, H.; Mori, S.; Nishizawa, N.K. Cloning and characterization of deoxymugineic acid synthase genes from graminaceous plants. J. Biol. Chem. 2006, 281, 32395–32402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nozoye, T.; Nagasaka, S.; Kobayashi, T.; Takahashi, M.; Sato, Y.; Uozumi, N.; Nakanishi, H.; Nishizawa, N.K. Phytosiderophore efflux transporters are crucial for iron acquisition in graminaceous plants. J. Biol. Chem. 2011, 286, 5446–5454. [Google Scholar] [CrossRef] [Green Version]
- Nozoye, T.; Nagasaka, S.; Kobayashi, T.; Sato, Y.; Uozumi, N.; Nakanishi, H.; Nishizawa, N.K. The phytosiderophore efflux transporter TOM2 is involved in metal transport in rice. J. Biol. Chem. 2015, 290, 27688–27699. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Kaur, G.; Kumar, A.; Meena, V.; Kaur, J.; Pandey, A.K. Overlapping transcriptional expression response of wheat zinc-induced facilitator-like transporters emphasize important role during Fe and Zn stress. BMC Mol. Biol. 2019, 20, 22. [Google Scholar] [CrossRef] [Green Version]
- Murakami, T.; Ise, K.; Hayakawa, M.; Kamei, S.; Takagi, S.I. Stabilities of metal-complexes of mugineic acids and their specific affinities for iron (III). Chem. Lett. 1989, 12, 2137–2140. [Google Scholar] [CrossRef]
- Xuan, Y.; Scheuermann, E.B.; Meda, A.R.; Hayen, H.; von Wireìn, N.; Weber, G. Separation and identification of phytosiderophores and their metal complexes in plants by zwitterionic hydrophilic interaction liquid chromatography coupled to electrospray ionization mass spectrometry. J. Chromatogr. A 2006, 1136, 73–81. [Google Scholar] [CrossRef]
- Nishiyama, R.; Kato, M.; Nagata, S.; Yanagisawa, S.; Yoneyama, T. Identification of Zn-nicotianamine and Fe-20-deoxymugineic acid in the phloem sap from rice plants (Oryza sativa L.). Plant Cell Physiol. 2012, 53, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Tsukamoto, T.; Inoue, H.; Watanabe, S.; Matsuhashi, S.; Takahashi, M.; Nakanishi, H.; Mori, S.; Nishizawa, N.K. Deoxymugineic acid increases Zn translocation in Zn-deficient rice plants. Plant Mol. Biol. 2008, 66, 609–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnold, T.; Kirk, G.J.; Wissuwa, M.; Frei, M.; Zhao, F.J.; Mason, T.F.; Weiss, D.J. Evidence for the mechanisms of zinc uptake by rice using isotope fractionation. Plant Cell Environ. 2010, 33, 370–381. [Google Scholar] [CrossRef] [PubMed]
- Bashir, K.; Ishimaru, Y.; Nishizawa, N.K. Molecular mechanisms of zinc uptake and translocation in rice. Plant Soil 2012, 361, 189–201. [Google Scholar] [CrossRef]
- Yancey, P.H. Organic osmolytes as compatible, metabolic and counteracting cytoprotectants in high osmolarity and other stresses. J. Exp. Biol. 2005, 208, 2819–2830. [Google Scholar] [CrossRef] [Green Version]
- Mittler, R. Abiotic stress, the field environment and stress combination. Trends Plant Sci. 2006, 11, 15–19. [Google Scholar] [CrossRef]
- Marschner, H. Mineral Nutrition of Higher Plants, 2nd ed.; Academic Press: London, UK, 1995. [Google Scholar]
- Cakmak, I. Possible roles of zinc in protecting plant cells from damage by reactive oxygen species. New Phytol. 2000, 146, 185–205. [Google Scholar] [CrossRef]
- Marschner, H. Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: London, UK, 2012. [Google Scholar]
- Alloway, B.J. Zinc in Soils and Crop Nutrition. International Zinc Association Communications; IZA Publications: Brussels, Belgium, 2004. [Google Scholar]
- Zeng, H.; Wu, H.; Yan, F.; Yi, K.; Zhu, Y. Molecular regulation of zinc deficiency responses in plants. J. Plant Physiol. 2021, 261, 153419. [Google Scholar] [CrossRef]
- Guilfoyle, T.J.; Hagen, G. Auxin response factors. Curr. Opin. Plant Biol. 2007, 10, 453–460. [Google Scholar] [CrossRef]
- Kazan, K. Auxin and the integration of environmental signals into plant root development. Annu. Bot. 2013, 112, 1655–1665. [Google Scholar] [CrossRef] [Green Version]
- Guilfoyle, T.J. The PB1 domain in auxin response factor and Aux/IAA proteins: A versatile protein interaction module in the auxin response. Plant Cell 2015, 27, 33–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jian, Y.; Wu, Y.; Hu, N.; Li, H.; Jiao, J. Interactions of bacterial-feeding nematodes and indole-3-acetic acid (IAA)-producing bacteria promotes growth of Arabidopsis thaliana by regulating soil auxin status. Appl. Soil Ecol. 2020, 147, 103447. [Google Scholar] [CrossRef]
- Jain, M.; Kaur, N.; Garg, R.; Thakur, J.K.; Tyagi, A.K.; Khurana, J.P. Structure and expression analysis of early auxin-responsive Aux/IAA gene family in rice (Oryza sativa). Funct. Integr. Genom. 2006, 6, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.L. The gene OsIAA9 encoding auxin/indole-3-acetic acid proteins is a negative regulator of auxin-regulated root growth in rice. Biol. Plant. 2019, 63, 210–218. [Google Scholar] [CrossRef]
- Iqbal, N.; Trivellini, A.; Masood, A.; Ferrante, A.; Khan, N.A. Current understanding on ethylene signaling in plants: The influence of nutrient availability. Plant Physiol. Biochem. 2013, 73, 128–138. [Google Scholar] [CrossRef]
- Peng, J.; Li, Z.; Wen, X.; Li, W.; Shi, H.; Yang, L.; Zhu, H.; Guo, H. Salt-induced stabilization of EIN3/EIL1 confers salinity tolerance by deterring ROS accumulation in Arabidopsis. PLoS Genet. 2014, 10, e1004664. [Google Scholar] [CrossRef] [Green Version]
- Tao, J.J.; Chen, H.W.; Ma, B.; Zhang, W.K.; Chen, S.Y.; Zhang, J.S. The role of ethylene in plants under salinity stress. Front. Plant Sci. 2015, 6, 1059. [Google Scholar] [CrossRef]
- Muchate, N.S.; Nikalje, G.C.; Rajurkar, N.S.; Suprasanna, P.; Nikam, T.D. Plant salt stress: Adaptive responses, tolerance mechanism and bioengineering for salt tolerance. Bot. Rev. 2016, 82, 371–406. [Google Scholar] [CrossRef]
- Khan, M.I.R.; Jahan, B.; Alajmi, M.F.; Rehman, M.T.; Khan, N.A. Exogenously-sourced ethylene modulates defense mechanisms and promotes tolerance to zinc stress in mustard (Brassica juncea L.). Plants 2019, 8, 540. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Tong, H.; Yang, X.; Xu, S.; Pan, Q.; Qiao, F.; Raihan, M.S.; Luo, Y.; Liu, H.; Zhang, X.; et al. Genome-wide dissection of the maize ear genetic architecture using multiple populations. New Phytol. 2016, 210, 1095–1106. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Z.B. Precision mapping of quantitative trait loci. Genetics 1994, 136, 999–1009. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait a | Treatment a | Parents | RIL Population | ||||
|---|---|---|---|---|---|---|---|
| K22♀ | BY815♂ | b | Mean | CV (%) | H2 (%)c | ||
| ZnSc | -Zn | 1 | 3 | ** | 2.5 | 55.4 | 83.1 |
| PH (cm) | -Zn | 29.5 | 67.2 | ** | 41.8 | 26.1 | 72.4 |
| SDW (g) | -Zn | 0.29 | 1.88 | ** | 0.79 | 46.6 | 81.2 |
| SDW | -Zn/CK | 0.23 | 1.16 | ** | 0.53 | 50.2 | NA |
| RDW (g) | -Zn | 0.12 | 0.64 | ** | 0.28 | 41.0 | 74.8 |
| RDW | -Zn/CK | 0.29 | 1.11 | ** | 0.64 | 47.2 | NA |
| R/S | -Zn | 0.34 | 0.37 | ** | 0.34 | 19.6 | 64.7 |
| R/S | -Zn/CK | 1.31 | 1.12 | * | 1.26 | 21.9 | NA |
| Trait | Treatment | Parents | RIL Population | ||||
|---|---|---|---|---|---|---|---|
| DAN340♀ | K22♂ | a | Mean | CV (%) | H2 (%) | ||
| ZnSc | -Zn | 3 | 1 | ** | 2.4 | 53.1 | 86.8 |
| PH (cm) | -Zn | 59.4 | 31 | ** | 37.5 | 30.2 | 81.5 |
| SDW (g) | -Zn | 1.04 | 0.36 | ** | 0.61 | 50.1 | 84.1 |
| SDW | -Zn/CK | 0.8 | 0.26 | ** | 0.54 | 53.8 | NA |
| RDW (g) | -Zn | 0.27 | 0.13 | ** | 0.18 | 37.1 | 77.4 |
| RDW | -Zn/CK | 0.86 | 0.31 | * | 0.56 | 48.7 | NA |
| R/S | -Zn | 0.36 | 0.33 | ** | 0.34 | 29.0 | 72.2 |
| R/S | -Zn/CK | 1.16 | 1.34 | ** | 1.17 | 23.9 | NA |
| Chr | Interval (Mb) a | QTL | Trait b | Physical Interval (Mb) a | LOD | R2 (%) | RIL Population c |
|---|---|---|---|---|---|---|---|
| 1 | 261.4–265.2 | qKB-PH1-1 | PH | 260.2–265.2 | 5.7 | 10.3 | KB |
| qDK-PH1-1 | PH | 261.4–266.6 | 9.5 | 14.2 | DK | ||
| 1 | 270.4–270.8 | qKB-RDW1-1 | RDW | 270.4–275.1 | 3 | 5.9 | KB |
| qDK-RDW1-1 | RDW | 267.1–270.8 | 6.4 | 11.4 | DK | ||
| 2 | 7.9–9.4 | qKB-ZnSc2-1 | ZnSc | 7.9–9.4 | 4.5 | 7.8 | KB |
| qZEAL-ZnSc2-1 | ZnSc | 4.9–14.4 | |||||
| 2 | 36.2–38.4 | qDK-ZnSc2-1 | ZnSc | 36.2–38.4 | 10 | 16.4 | DK |
| qZEAL-ZnSc2-2 | ZnSc | 33.5–111.8 | 3.9 | 10.1 | YW | ||
| 2 | 108.6–134.3 | qDK-SDW2-1 | SDW | 74.6–153.0 | 12.4 | 23.2 | DK |
| qZEAL-SDW2-1 | SDW | 111.8–176.3 | 6.1 | 20.7 | YW | ||
| qZEAL-SDW2-2 | SDW | 111.8–176.3 | 6.3 | 17.3 | YW |
| QTL | Chr | Gene ID | Gene Position (bp) a | Description |
|---|---|---|---|---|
| qDK-R/S1-3 | 1 | Zm00001eb010040 | 31742085–31745237 | ZmDMAS1—Deoxymugineic acid synthase 1 |
| qDK-R/S1-3 | 1 | Zm00001eb011850 | 38561776–38563168 | ZmEBF1—EIN3-binding F box protein 1 |
| qKB-PH1-1, qDK-PH1-1 | 1 | Zm00001eb051790 | 262493944–262496834 | ZmNRAMP—Metal transporter Nramp 6 |
| 1 | Zm00001eb052440 | 264886590–264888963 | ZmIRT1—Iron-regulated transporter 1 | |
| qDK-PH1-1 | 1 | Zm00001eb052890 | 266321457–266323150 | ZmNAS3—Nicotianamine synthase 3 |
| qKB-RDW1-1 | 1 | Zm00001eb054060 | 270891072–270897163 | ZmEIN2—Ethylene-insensitive protein 2 |
| qKB-RDW1-1 | 1 | Zm00001eb054170 | 271172369–271177408 | ZmETR—Signal transduction histidine kinase hybrid-type ethylene sensor |
| qDK-ZnSc2-2 | 2 | Zm00001eb085030 | 67296998–67301921 | ZmAFB—Auxin signaling F-box |
| qDK-ZnSc2-2, qDK-PH2-1, qDK-SDW2-1, qDK-R/S2-1 | 2 | Zm00001eb093430 | 152901305–152906124 | ZmTOM3—Transporter of mugineic acid 3 |
| qDK-ZnSc2-3, qDK-PH2-2, qDK-R/S2-2 | 2 | Zm00001eb095010 | 162889727–162893463 | ZmHMA4—Cadmium/Zinc-transporting ATPase HMA4 |
| 2 | Zm00001eb095020 | 162910876–162914856 | ZmHMA3—Cadmium/Zinc-transporting ATPase HMA3 | |
| qDK-ZnSc2-3, qDK-PH2-2, qDK-R/S2-2 | 2 | Zm00001eb096080 | 170130733–170150672 | ZmCTR3—Constitutive triple response 3 |
| qDK-PH5-1, qDK-RDW5-1 | 5 | Zm00001eb248740 | 193961138–193962096 | ZmVIT—Vacuolar iron transporter (VIT) family protein |
| qKB-R/S5-1, qZEAL-RDW5-1 | 5 | Zm00001eb258220 | 222819625–222821061 | ZmIAA17—auxin-responsive Aux/IAA family member |
| qKB-R/S7-2 | 7 | Zm00001eb331080 | 182932191–182934361 | ZmEIN3—Ethylene insensitive 3 family protein |
| qKB-PH9-1, qKB-ZnSc9-2, qKB-SDW9-1 | 9 | Zm00001eb389830 | 114577311–114585730 | ZmHMA9—Heavy metal translocating P-type ATPase transports |
| qDK-PH10-1 | 10 | Zm00001eb421340 | 113391837–113396456 | ZmAFB—Auxin signaling F-box |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, J.; Qin, X.; Ni, Z.; Chen, F.; Fu, X.; Yu, F. Identification of Zinc Efficiency-Associated Loci (ZEALs) and Candidate Genes for Zn Deficiency Tolerance of Two Recombination Inbred Line Populations in Maize. Int. J. Mol. Sci. 2022, 23, 4852. https://doi.org/10.3390/ijms23094852
Xu J, Qin X, Ni Z, Chen F, Fu X, Yu F. Identification of Zinc Efficiency-Associated Loci (ZEALs) and Candidate Genes for Zn Deficiency Tolerance of Two Recombination Inbred Line Populations in Maize. International Journal of Molecular Sciences. 2022; 23(9):4852. https://doi.org/10.3390/ijms23094852
Chicago/Turabian StyleXu, Jianqin, Xiaoxin Qin, Zhongfu Ni, Fanjun Chen, Xiuyi Fu, and Futong Yu. 2022. "Identification of Zinc Efficiency-Associated Loci (ZEALs) and Candidate Genes for Zn Deficiency Tolerance of Two Recombination Inbred Line Populations in Maize" International Journal of Molecular Sciences 23, no. 9: 4852. https://doi.org/10.3390/ijms23094852