
ClPIF3-ClHY5 Module Regulates ClPSY1 to Promote Watermelon Fruit Lycopene Accumulation Earlier under Supplementary Red Lighting

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Supplementary Red Lighting Promoted the Lycopene Accumulation Earlier in Watermelon Flesh
2.2. Carotenoid Biosynthesis and Metabolism-Related Genes Expression after Supplementary Red Lighting
2.3. Phylogenetic and Expression Analysis of ClHY5
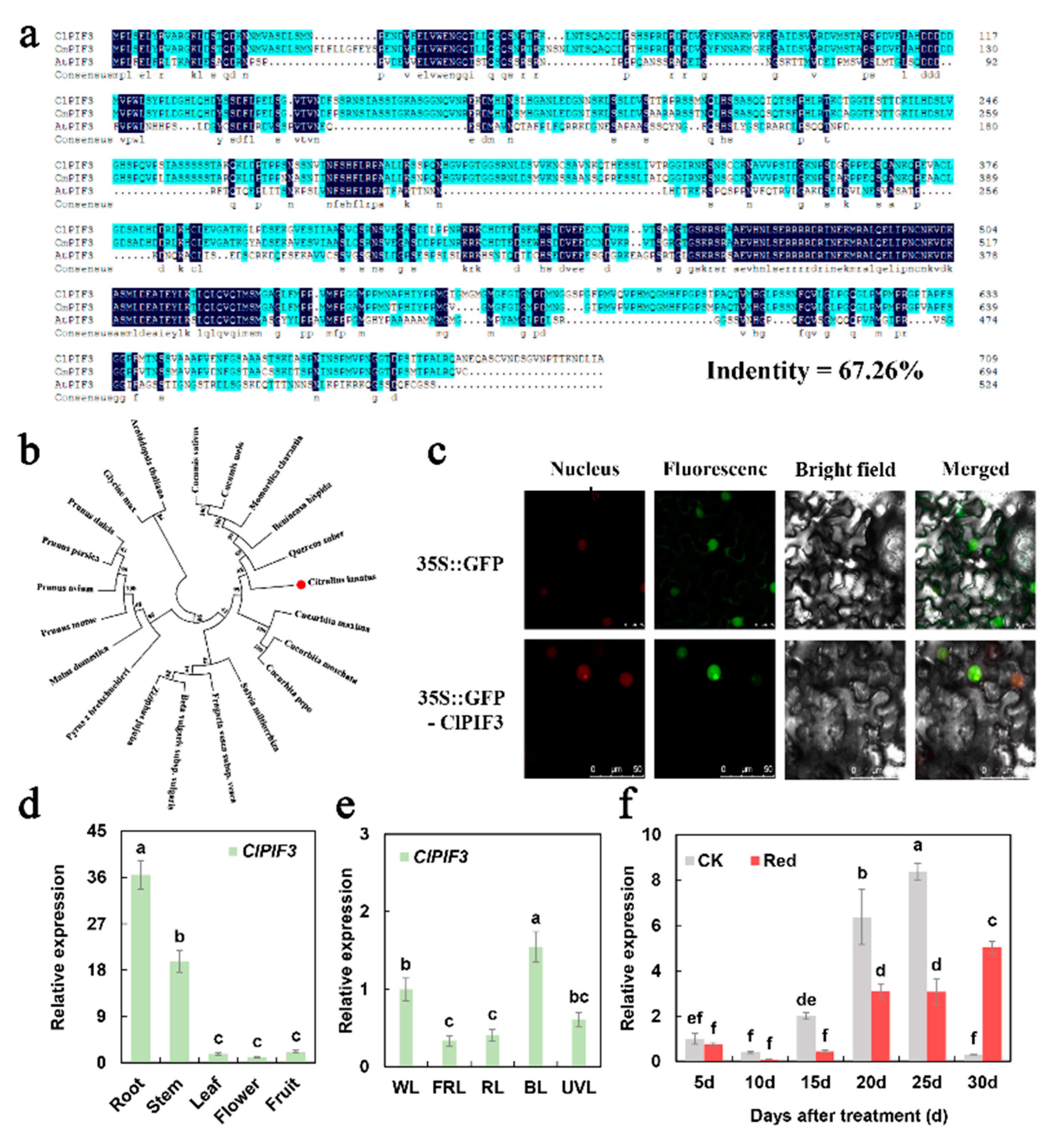
2.4. Phylogenetic and Expression Analysis of ClPIF3
2.5. ClHY5 Positively Regulated While ClPIF3 Negatively Regulated ClPSY1 Expression
3. Discussion
4. Materials and Methods
4.1. Plant Material, Growth Conditions, and Light Treatments
4.2. Flesh Color and Carotenoid Determination
4.3. qRT-PCR Analysis
4.4. Phylogenetic Analysis
4.5. Subcellular Localization Analysis
4.6. Yeast One-Hybrid Assays (Y1H)
4.7. Electrophoretic Mobility Shift Assays (EMSA)
4.8. Luciferase Reporter Assay (LUC)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO Statistics. Available online: https://www.fao.org/home/en/ (accessed on 9 March 2022).
- Sowbiya, M.; Ho, K.C.; Prabhakaran, S.; Abinaya, M.; Gyeong, P.Y.; Ryong, J.B.; Bie, Z. Proteomic Study Related to Vascular Connections in Watermelon Scions Grafted onto Bottle-Gourd Rootstock under Different Light Intensities. PLoS ONE 2015, 10, e0120899. [Google Scholar]
- Tewolde, F.T.; Shiina, K.; Maruo, T.; Takagaki, M.; Kozai, T.; Yamori, W.; Ezura, H. Supplemental LED inter-lighting compensates for a shortage of light for plant growth and yield under the lack of sunshine. PLoS ONE 2018, 13, e0206592. [Google Scholar] [CrossRef] [PubMed]
- Bantis, F.; Smirnakou, S.; Ouzounis, T.; Koukounaras, A.; Radoglou, K. Current status and recent achievements in the field of horticulture with the use of light-emitting diodes (LEDs). Sci. Hortic. 2018, 235, 437–451. [Google Scholar] [CrossRef]
- Joshi, N.C.; Ratner, K.; Eidelman, O.; Bednarczyk, D.; Charuvi, D. Effects of daytime intra-canopy LED illumination on photosynthesis and productivity of bell pepper grown in protected cultivation. Sci. Hortic. 2019, 250, 81–88. [Google Scholar] [CrossRef]
- Wang, J.; Mao, S.; Wu, Q.; Yuan, Y.; Liang, M.; Wang, S.; Huang, K.; Wu, Q. Effects of LED illumination spectra on glucosinolate and sulforaphane accumulation in broccoli seedlings. Food Chem. 2021, 356, 129550. [Google Scholar] [CrossRef]
- Ma, G.; Zhang, L.; Kato, M.; Yamawaki, K.; Kiriiwa, Y.; Yahata, M.; Ikoma, Y.; Matsumoto, H. Effect of the combination of ethylene and red LED light irradiation on carotenoid accumulation and carotenogenic gene expression in the flavedo of citrus fruit. Postharvest Biol. Technol. 2015, 99, 99–104. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.G.; Moon, B.Y.; Kang, N.J. Effects of LED light on the production of strawberry during cultivation in a plastic greenhouse and in a growth chamber. Sci. Hortic. 2015, 189, 22–31. [Google Scholar] [CrossRef]
- Virsile, A.; Brazaityte, A.; Vastakaite-Kairiene, V.; Miliauskiene, J.; Jankauskiene, J.; Novickovas, A.; Lauzike, K.; Samuoliene, G. The distinct impact of multi-color LED light on nitrate, amino acid, soluble sugar and organic acid contents in red and green leaf lettuce cultivated in controlled environment. Food Chem. 2020, 310, 125799. [Google Scholar] [CrossRef]
- Samkumar, A.; Jones, D.; Karppinen, K.; Dare, A.P.; Sipari, N.; Espley, R.V.; Martinussen, I.; Jaakola, L. Red and blue light treatments of ripening bilberry fruits reveal differences in signalling through abscisic acid-regulated anthocyanin biosynthesis. Plant Cell Environ. 2021, 44, 3227–3245. [Google Scholar] [CrossRef]
- Cui, X.H.; Guo, X.O.; Sun, T.Y.; Hong-Yan, Q.I. Effects of LED supplementary lighting on seedling growth and fruit quality of oriental melon. Plant Physiol. J. 2017, 53, 656–667. [Google Scholar]
- Chory, J.; Chatterjee, M. From seed germination to flowering, light controls plant development via the pigment phytochrome. Proc. Natl. Acad. Sci. USA 1996, 93, 12066–12071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.I.; Park, J.E.; Zarate, X.; Song, P.S. Phytochrome phosphorylation in plant light signaling. Photochem. Photobiol. Sci. 2005, 4, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Al-Sady, B.; Ni, W.; Kircher, S.; Schäfer, E.; Quail, P.H. Photoactivated phytochrome induces rapid PIF3 phosphorylation prior to proteasome-mediated degradation. Mol. Cell 2006, 23, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Jang, I.C.; Henriques, R.; Seo, H.S.; Nagatani, A.; Chua, N.H. Arabidopsis PHYTOCHROME INTERACTING FACTOR Proteins Promote Phytochrome B Polyubiquitination by COP1 E3 Ligase in the Nucleus. Plant Cell 2010, 22, 2370–2383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leivar, P.; Monte, E. PIFs: Systems Integrators in Plant Development. Plant Cell 2014, 26, 56–78. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Mayba, O.; Pfeiffer, A.; Shi, H.; Tepperman, J.M.; Speed, T.P.; Quail, P.H.; Copenhaver, G.P. A Quartet of PIF bHLH Factors Provides a Transcriptionally Centered Signaling Hub That Regulates Seedling Morphogenesis through Differential Expression-Patterning of Shared Target Genes in Arabidopsis. PLoS Genet. 2013, 9, e1003244. [Google Scholar] [CrossRef] [Green Version]
- Gangappa, S.N.; Botto, J.F. The Multifaceted Roles of HY5 in Plant Growth and Development. Mol. Plant 2016, 9, 1353–1365. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; He, K.; Stolc, V.; Lee, H.; Figueroa, P.; Gao, Y.; Tongprasit, W.; Zhao, H.; Lee, I.; Deng, X.W. Analysis of Transcription Factor HY5 Genomic Binding Sites Revealed Its Hierarchical Role in Light Regulation of Development. Plant Cell Online 2007, 19, 731–749. [Google Scholar] [CrossRef] [Green Version]
- Chattopadhyay, S.; Ang, L.H.; Puente, P.; Deng, X.W.; Wei, N. Arabidopsis bZIP protein HY5 directly interacts with light-responsive promoters in mediating light control of gene expression. Plant Cell 1998, 10, 673–683. [Google Scholar] [CrossRef] [Green Version]
- Nozue, K.; Covington, M.F.; Duek, P.D.; Lorrain, S.; Fankhauser, C.; Harmer, S.L.; Maloof, J.N. Rhythmic growth explained by coincidence between internal and external cues. Nature 2007, 448, 358–361. [Google Scholar] [CrossRef]
- Soy, J.; Leivar, P.; González-Schain, N.; Sentandreu, M.; Prat, S.; Quail, P.H.; Monte, E. Phytochrome-imposed oscillations in PIF3 protein abundance regulate hypocotyl growth under diurnal light/dark conditions in Arabidopsis. Plant J. 2012, 71, 390–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leivar, P.; Quail, P.H. PIFs: Pivotal components in a cellular signaling hub. Trends Plant Sci. 2011, 16, 19–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osterlund, M.T.; Hardtke, C.S.; Wei, N.; Deng, X.W. Targeted destabilization of HY5 during light-regulated development of Arabidopsis. Nature 2000, 405, 462–466. [Google Scholar] [CrossRef] [PubMed]
- Gallé, Á.; Czékus, Z.; Tóth, L.; Galgóczy, L.; Poór, P. Pest and disease management by red light. Plant Cell Environ. 2021, 44, 3197–3210. [Google Scholar] [CrossRef] [PubMed]
- Kami, C.; Lorrain, S.; Hornitschek, P.; Fankhauser, C. Light-regulated plant growth and development. Curr. Top. Dev. Biol. 2010, 91, 29. [Google Scholar]
- Lv, P.; Na, L.; Liu, H.; Gu, H.; Zhao, W.E. Changes in carotenoid profiles and in the expression pattern of the genes in carotenoid metabolisms during fruit development and ripening in four watermelon cultivars. Food Chem. 2015, 174, 52–59. [Google Scholar] [CrossRef]
- Auldridge, M.E.; Mccarty, D.R.; Klee, H.J. Plant carotenoid cleavage oxygenases and their apocarotenoid products. Curr. Opin. Plant Biol. 2006, 9, 315–321. [Google Scholar] [CrossRef]
- Walter, M.H.; Strack, D. Carotenoids and their cleavage products: Biosynthesis and functions. Nat. Prod. Rep. 2011, 28, 663–692. [Google Scholar] [CrossRef]
- Perkins-Veazie, P.; Collins, J.K.; Davis, A.R.; Roberts, W. Carotenoid content of 50 watermelon cultivars. J. Agric. Food Chem. 2006, 54, 2593. [Google Scholar] [CrossRef]
- Tadmor, Y.; King, S.; Levi, A.; Davis, A.; Meir, A.; Wasserman, B.; Hirschberg, J.; Lewinsohn, E. Comparative fruit colouration in watermelon and tomato. Food Res. Int. 2005, 38, 837–841. [Google Scholar] [CrossRef]
- Gady, A.L.; Vriezen, W.H.; Van de Wal, M.H.; Huang, P.; Bovy, A.G.; Visser, R.G.; Bachem, C.W. Induced point mutations in the phytoene synthase 1 gene cause differences in carotenoid content during tomato fruit ripening. Mol. Breed. 2012, 29, 801–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, A.J.; Vinyard, B.T.; Wiley, E.R.; Brown, E.D.; Collins, J.K.; Perkins-Veazie, P.; Baker, R.A.; Clevidence, B.A. Consumption of Watermelon Juice Increases Plasma Concentrations of Lycopene and β-Carotene in Humans. J. Nutr. 2003, 133, 1043–1050. [Google Scholar] [CrossRef] [PubMed]
- Molecular Bioinformatics Center. Available online: http://cello.life.nctu.edu.tw/ (accessed on 11 March 2022).
- Alba, R. Fruit-localized phytochromes regulate lycopene accumulation independently of ethylene production in tomato. Plant Physiol. 2000, 123, 363–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.H.; Zabaras, D.; Bennett, L.E.; Aguas, P.; Woonton, B.W. Effects of UV-C, red light and sun light on the carotenoid content and physical qualities of tomatoes during post-harvest storage. Food Chem. 2009, 115, 495–500. [Google Scholar] [CrossRef]
- Nisar, N.; Li, L.; Lu, S.; Khin, N.C.; Pogson, B. Carotenoid Metabolism in Plants. Mol. Plant 2015, 8, 68–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Sun, H.; Guo, S.; Ren, Y.; Li, M.; Wang, J.; Zhang, H.; Gong, G.; Xu, Y. Decreased Protein Abundance of Lycopene β-Cyclase Contributes to Red Flesh in Domesticated Watermelon1. Plant Physiol. 2020, 183, 1171–1183. [Google Scholar] [CrossRef] [PubMed]
- Koornneef, M.; Rolff, E.; Spruit, C.J.P. Genetic control of light-inhibited hypocotyl elongation in Arabidopsis thaliana (L.) Heynh. Pflanzenphysiologie 1980, 100, 147–160. [Google Scholar] [CrossRef]
- Toledo-Ortiz, G.; Johansson, H.; Lee, K.P.; Bou-Torrent, J.; Stewart, K.; Steel, G.; Rodríguez-Concepción, M.; Halliday, K.J.; Qu, L.J. The HY5-PIF Regulatory Module Coordinates Light and Temperature Control of Photosynthetic Gene Transcription. PLoS Genet. 2014, 10, e1004416. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Yao, Q.; Gao, X.; Jiang, C.; Fu, X. Shoot-to-Root Mobile Transcription Factor HY5 Coordinates Plant Carbon and Nitrogen Acquisition. Curr. Biol. 2016, 26, 640–646. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Chen, P.; Jiang, L.; Chen, F.; Tang, H. Cloning and Expression Analysis of Light Responsive bZIP Transcription Factor FaHY5 from Strawberry. Genom. Appl. Biol. 2015, 34, 2695–2701. [Google Scholar]
- Jiang, B.; Shi, Y.; Zhang, X.; Xin, X.; Yang, S. PIF3 is a negative regulator of the CBF pathway and freezing tolerance in Arabidopsis. Proc. Natl. Acad. Sci. USA 2017, 114, E6695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Shang, F.; Huai, J.; Xu, G.; Tang, W.; Jing, Y.; Lin, R. A PIF1/PIF3-HY5-BBX23 transcription factor cascade affects photomorphogenesis. Plant Physiol. 2017, 174, 2487–2500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Job, N.; Datta, S. PIF3/HY5 module regulates BBX11 to suppress protochlorophyllide levels in dark and promote photomorphogenesis in light. New Phytol. 2021, 230, 190–204. [Google Scholar] [CrossRef] [PubMed]
- Pokhilko, A.; Ramos, J.A.; Holtan, H.; Maszle, D.R.; Khanna, R.; Millar, A.J. Ubiquitin ligase switch in plant photomorphogenesis: A hypothesis. J. Theor. Biol. 2011, 270, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Chory, J.; Fankhauser, C. Light signal transduction in higher plants. Annu. Rev. Genet. 2004, 38, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, J.; Terzaghi, W.; Deng, X.W.; Chen, H. Multiple photomorphogenic repressors work in concert to regulate Arabidopsis seedling development. Plant Signal. Behav. 2015, 10, e1011934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakuraba, Y.; Kanno, S.; Mabuchi, A.; Monda, K.; Iba, K.; Yanagisawa, S. A phytochrome-B-mediated regulatory mechanism of phosphorus acquisition. Nat. Plants 2018, 4, 1089–1101. [Google Scholar] [CrossRef]
- Liu, S.; Gao, P.; Wang, X.; Davis, A.R.; Baloch, A.M.; Luan, F. Mapping of quantitative trait loci for lycopene content and fruit traits in Citrullus lanatus. Euphytica 2015, 202, 411–426. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, T.; Zhao, L.; Zhang, S.; Guan, J.; Liu, W.; Qi, H. ClPIF3-ClHY5 Module Regulates ClPSY1 to Promote Watermelon Fruit Lycopene Accumulation Earlier under Supplementary Red Lighting. Int. J. Mol. Sci. 2022, 23, 4145. https://doi.org/10.3390/ijms23084145
Lv T, Zhao L, Zhang S, Guan J, Liu W, Qi H. ClPIF3-ClHY5 Module Regulates ClPSY1 to Promote Watermelon Fruit Lycopene Accumulation Earlier under Supplementary Red Lighting. International Journal of Molecular Sciences. 2022; 23(8):4145. https://doi.org/10.3390/ijms23084145
Chicago/Turabian StyleLv, Tinghui, Lili Zhao, Shuting Zhang, Jingyue Guan, Wei Liu, and Hongyan Qi. 2022. "ClPIF3-ClHY5 Module Regulates ClPSY1 to Promote Watermelon Fruit Lycopene Accumulation Earlier under Supplementary Red Lighting" International Journal of Molecular Sciences 23, no. 8: 4145. https://doi.org/10.3390/ijms23084145