1. Introduction
Cancer is a major global health problem, and the burden of cancer incidence and mortality is rising worldwide [
1]. One part of the problem is the lack of relevant biomarker tools that can, amongst other things, detect cancer earlier and predict survival and response to treatment. Additionally, traditional biomarkers usually involve invasive procedures such as tissue biopsies. A promising non-invasive alternative is liquid biopsies, where biomarkers are measured in the blood and can be assessed easily with minimal discomfort and complications [
2]. Examples include measuring the presence of tumor components such as tumor DNA or tumor cells directly [
3]. This approach, however, is blind to the influence of the tumor microenvironment, which in recent years has proven to be intricately involved in many important aspects of cancer [
4] and components of the tumor microenvironment can be measured in blood [
5]. Therefore, a novel and fruitful approach could be to supplement an assessment of the tumor with an assessment of its environment.
The extracellular matrix (ECM) of the tumor microenvironment, defined as the non-cellular part of tissues, has been recognized as an important part of cancer development [
6]. When assessing the ECM, one cannot overlook the collagens, because they are collectively some of the most abundant proteins in the human body and have also been implicated in a variety of different processes relevant to cancer: tumor stiffness, tumor immunity, and metastasis [
7]. In cancer, the dynamic balance of collagen formation and degradation is knocked askew, and collagen can accumulate to form tumor fibrosis. The collagen superfamily spans 28 different types and most of the collagen cancer research has focused on the abundant and well-characterized collagens, such as type I, III, or IV collagens. As an example, type I collagen is ubiquitous in the tumor microenvironment and is the primary component of tumor fibrosis [
8]. In addition to its structural role, type I collagen can activate signaling that promotes invasion [
9,
10]. Recent reports, however, suggest that collagens, such as type I collagen, can have both pro- and anti-tumor effects [
11,
12,
13].
We have previously shown that these collagens can be used as cancer biomarkers. For example, non-invasive quantification of type III collagen deposition reflects fibrotic activity and is prognostic in several cancers [
14]. Seeing this, we pondered the biomarker potential of the other members of the collagen family. Additionally, in contrast to the abundant collagens, very little is known about the role in cancer of the minor and poorly characterized collagens. Nonetheless, a connection between cancer and the minor collagens can be hypothesized because the expression of many minor collagens is exclusive to tissue development, which shares with cancer the activation of signaling pathways and regulation of ECM components [
15,
16,
17]. This is true for the membrane-bound and multiplexin collagens, which have both structural and signaling functions shared between tissue development and cancer, and their expression is correlated with survival in patients with cancer [
18]. It appears that these specialized collagens are used for specific purposes, namely the development of tissues and cancer—both involving dramatic tissue remodeling [
17]. In support of this hypothesis, we recently discovered elevated levels of a minor collagen, type XIX collagen, in the blood of patients with cancer [
19]. However, many minor collagens remain largely unexplored in the cancer context and this prompted the present study.
Type XX collagen is one such unexplored collagen. Based on its structural features, type XX collagen is part of the fibril-associated collagens with interrupted triple helices (FACIT) family. These collagens are thought to associate with the fibrillar collagens to regulate their organization and interactions [
20]. Structural features of type XX collagen include several fibronectin type III repeats, a von Willebrand factor A domain, a thrombospondin-like domain, as well as collagenous triple-helix domains, interspersed with non-collagenous domains [
20]. Within the FACIT family, type XII and XIV are the closest relatives to type XX collagen [
20]. Little is known about the expression, localization, and function of type XX collagen. It was originally cloned from chick embryos in which expression was primarily in corneal epithelium, but detectable in embryonic skin, lung, sternal cartilage, and tendon [
20]. RNA expression data from the human protein atlas (
https://www.proteinatlas.org/ENSG00000101203-COL20A1/tissue, accessed on 25 February 2022) suggest an enrichment of COL20A1 RNA expression in human brain and minor upticks in testes and spleen tissues [
21]. In the context of cancer, previous studies have mostly found type XX collagen in brain cancers. Based on cDNA microarrays, COL20A1 was elevated in so-called brain-tumor initiating cells versus regular glioma cell lines and normal brain astrocytes [
22]. In a separate study, using biopsy-derived glioma cell models, a downregulation of COL20A1 RNA was observed after treatment with histone deacetylase inhibitors [
23]. Reports on type XX collagen in other cancers are rare, but COL20A1 was included in a 16-gene signature associated with breast cancer recurrence, metastasis, and poor survival in a Chinese population. However, the weight that COL20A1 was assigned in the reported model was minor [
24]. Lastly, COL20A1 gene expression is also upregulated in early stage prostate tumorigenesis [
25]. These studies interrogate type XX collagen at the RNA or DNA levels, so studies at the protein level and its relevance to cancer are lacking.
In this study, we set out to develop a robust ELISA assay targeting the C-terminus of type XX collagen and use it to quantify levels in serum samples from patients with cancer to evaluate its biomarker potential. This study is the first to report on an assay quantifying type XX collagen and the first to show that, not only can it be measured it blood, but it is elevated in sera of patients with cancer and is prognostic for survival.
3. Discussion
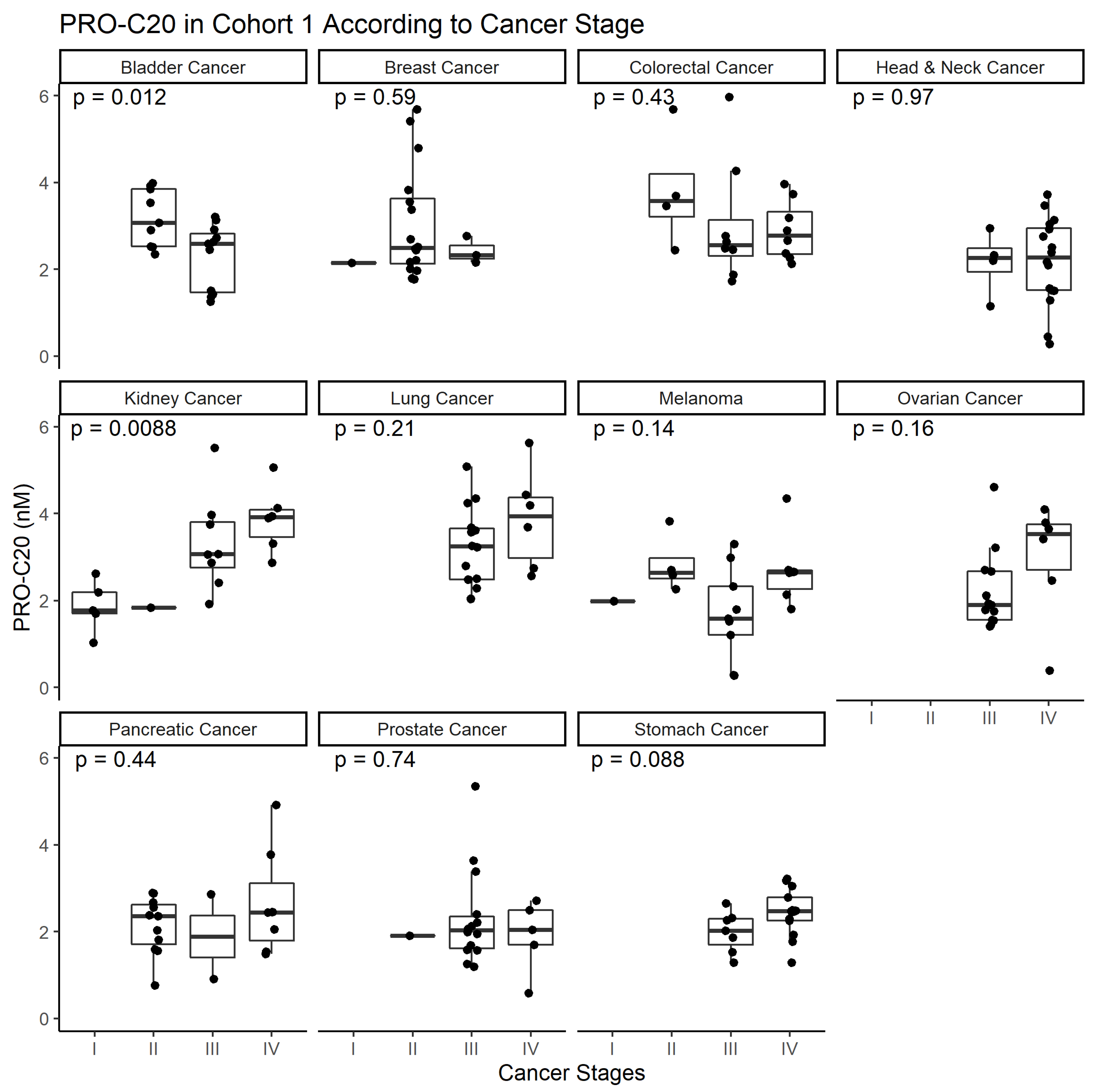
To study type XX collagen and its role in cancer, we developed the PRO-C20 ELISA. The assay proved to be a sensitive assay, capable of detecting nanomolar amounts of type XX collagen. It was also specific, as evidenced by the epitope specificity of the antibody, and it was accurate in the complex sample matrix of serum. Lastly, the analyte and reagent stability of the assay indicate that it is a robust assay. PRO-C20 was elevated in all cancers tested and although a downward trend in bladder and upward trend in kidney cancer was seen as a function of cancer stage, overall PRO-C20 levels did not seem to be associated with cancer stage. Although not the aim of this study, PRO-C20 proved excellent at discriminating between sera from healthy controls and patients with cancer with large AUC values, but was limited in terms of its diagnostic specificity, seeing as a substantial proportion of the healthy individuals would be misclassified using the cutoffs described in the current study. Future studies should pinpoint the discriminatory performance of PRO-C20 and do so with more clinically relevant control subjects, e.g., individuals with comorbidities such as other chronic diseases.
Compared to PRO-C20, PRO-C1 was not effective at discriminating between sera from healthy controls and patients with cancer. Although type I collagen is elevated in several cancers compared to healthy controls [
26], the epitope that the PRO-C1 assay targets is mostly associated with bone metastasis [
27] which the patients included here do not have. Part of the reason to include PRO-C1 in the analysis here was to make a comparison to a well-described and abundant protein. This comparison highlights the merit of investigating the biomarker potential of the otherwise poorly characterized collagens of low abundance. This study also lends merit to the hypothesis that the minor collagens are deregulated in disease and may be more pathology-relevant than abundant proteins. Collagens in general can be predictive of response to treatment [
13] and we have previously demonstrated how non-invasive quantification of fibrillar collagens, such as type III collagen, can be used to assess fibrotic activity from the periphery and is prognostic in several cancers [
14] and predictive of immunotherapy response [
5]. The current study builds upon these discoveries and expands the potential of the approach by demonstrating that the breadth of the collagen family may also have biomarker potential.
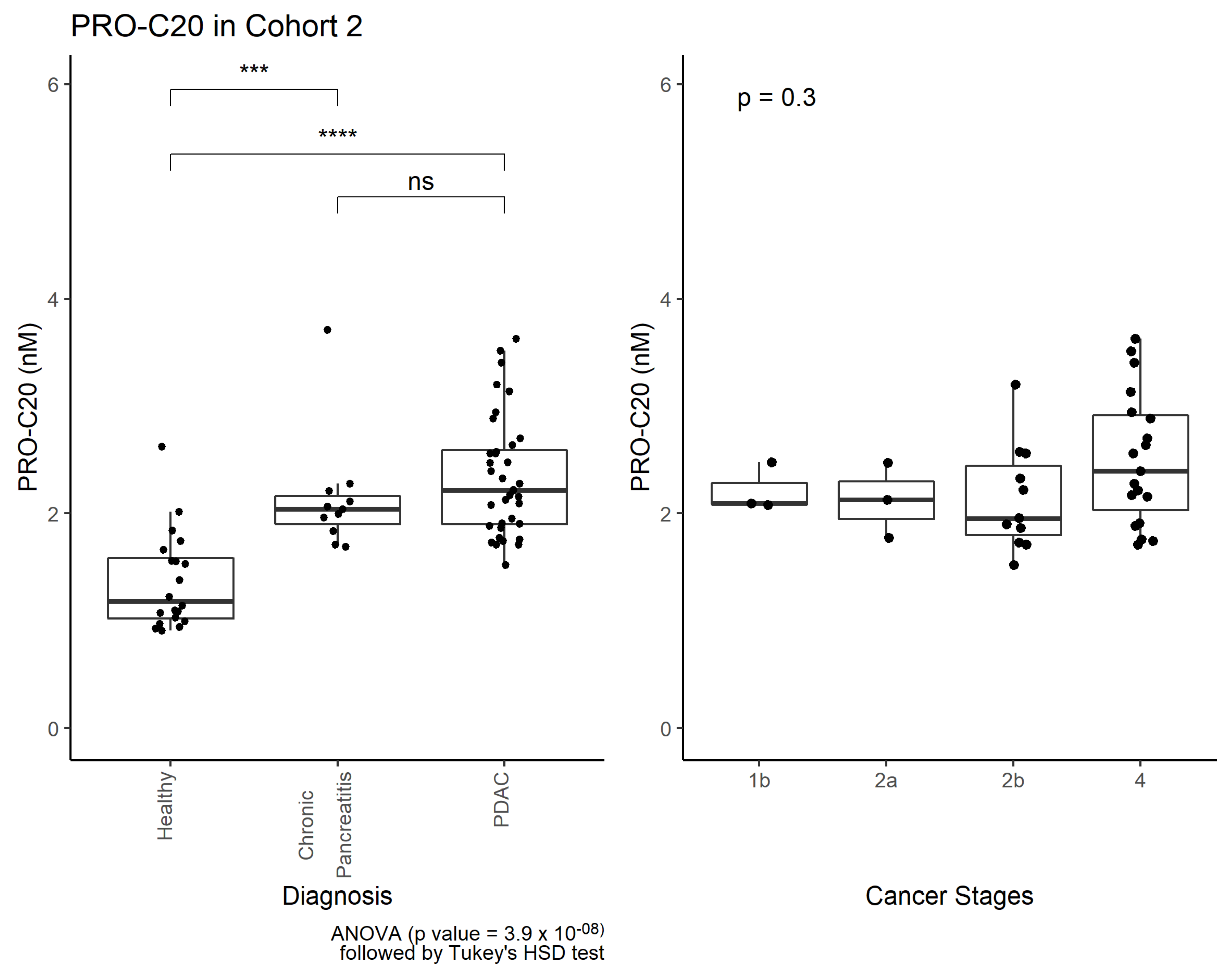
Interestingly, PRO-C20 was also elevated in patients with chronic pancreatitis, indicating PRO-C20 is released to circulation as a function of inflammation and fibrosis. Chronic pancreatitis is a well-known risk factor of cancer. In fact, it can develop as a result of an underlying, undiagnosed, pancreatic cancer [
28], and chronic pancreatitis is characterized by fibrosis and inflammation, similar to cancer, making it difficult to distinguish between the two pathologies [
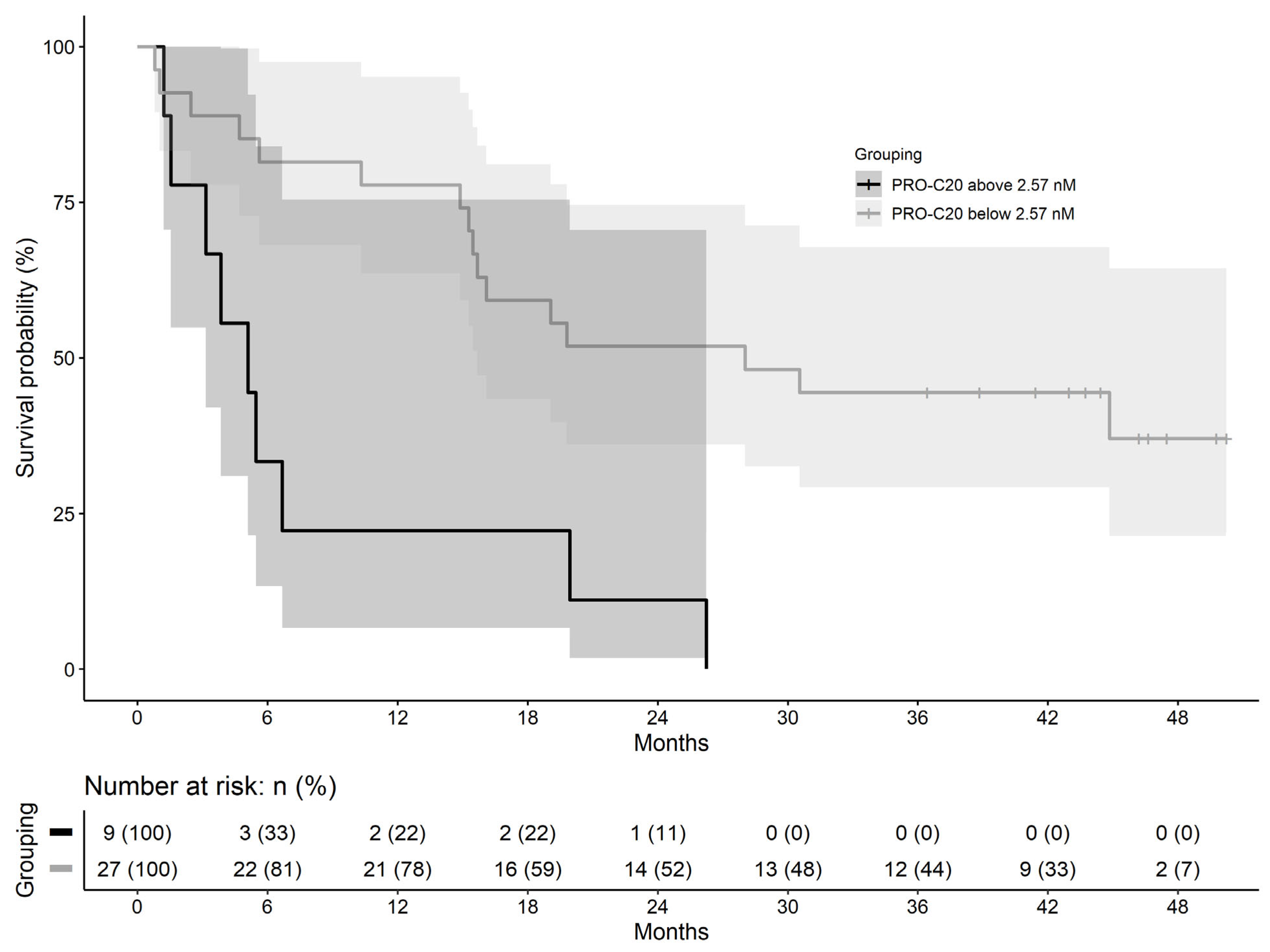
28]. Based on our data, PRO-C20 could not separate the two pathologies, suggesting PRO-C20 is not only released to circulation as a result of cancer, but as a result of more general tissue remodeling. Instead, a context of use for PRO-C20 could be as a PDAC prognostic biomarker, where high PRO-C20 levels were associated with poor overall survival. PDAC has a high mortality rate and a prognostic biomarker in this context could, for example, identify the patients where a comprehensive and potentially toxic therapy regimen may not be appropriate. Follow-up studies should be performed to confirm the findings and further delineate the context of use for PRO-C20.
The function of FACITs such as type XX collagen is unclear. Much of the understanding of how FACITs function is derived from biochemical experiments of type IX and XII collagens. These FACITs bind to the collagen fibrils with their C-terminal region, whereas the N-terminal region projects out from the fibril to interact with other components of the ECM [
29,
30]. In this way, FACITs can mediate within and between fibril interactions and be seen as organizers of the collagenous ECM and therefore intricately involved in establishing matrix and tissue structure. Type XX collagen shares many structural features with type XII collagen, including the domain structure of both its C-terminal and N-terminal regions, so it is likely that they share a similar function of integrating collagen fibrils. The connection of type XX collagen to fibrillar collagens is not insignificant: changes in the ECM and collagen composition as a result of cancer has been shown to influence tumor progression, metastasis, and clinical outcome of patients [
7]. Additionally, the minor collagens associated with the fibrillar ones are important for the formation of new fibrils [
31]. Formation of new and remodeling of existing fibrils is relevant to the formation of so-called pre-metastatic niches, wherein the ECM of a metastatic site is remodeled to facilitate the metastasizing cancer cells [
32]. Although we did not see a clear association with PRO-C20 levels and cancer stages, several other members of the FACIT family of collagens seem to be regulated as part of the metastatic cascade: type XII collagen is found upregulated at the invasive front of colon cancer cells [
33] and type XIX collagen is degraded prior to the degradation of the basement membrane and intravasation [
34].
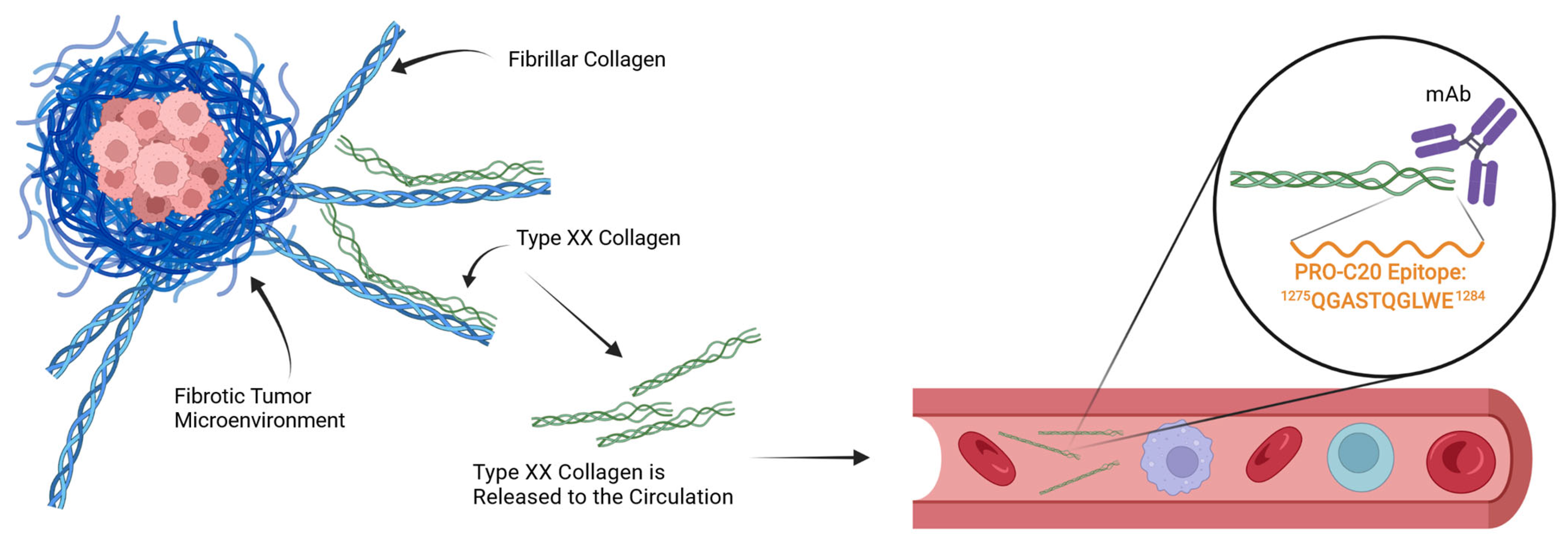
In the case of PRO-C20, which targets the natural C-terminus of the protein, the mechanism of its release to circulation is unclear. Intuitively, a full-length and intact type XX collagen protein would be expected to remain in the tissue to exert its role in integrating collagen fibers. Conversely, a fragmented protein would lose one or more points of interaction with other proteins and is thus less likely to be retained and more likely to be found in circulation (
Figure 6). It is therefore possible that PRO-C20 measures fragments and reflects the degradation of type XX collagen, and may explain why the conclusions drawn from our quantification of type XX collagen differs from some of the gene expression studies outlined in the introduction. This difference could be important, because fragmentation of type XX collagen could be a way to downregulate the integration of fibrillar collagens and possibly expose them to further degradation and remodeling. If true, the fragmentation of type XX collagen and the release of the PRO-C20 epitope could reflect remodeling of the fibrillar collagens. The mechanics of this process are not well described, and the quantification approach presented in this study is therefore limited in this respect. Further studies into the processing and release of type XX collagen are needed.
Research on the role of type XX collagen in cancer is very limited, but looking to its closest collagen relative, type XII collagen, reveals some interesting perspectives for the further study of type XX collagen. Studies of skin and cornea reveal that FACITs can regulate collagen fibril diameter and density [
35,
36]. More and thicker fibrils are associated with an overall stiff matrix, which is a well-known characteristic of cancer [
17]. A stiff matrix can, amongst other things, prevent the host immune cells from attacking the tumor cells [
37]. Further, type XII collagen is secreted by cancer-associated fibroblasts and colon cancer cells at the invasive front of lesions and is associated with myofibroblastic differentiation [
33,
38] and involved in TGFβ signaling [
35,
39]. It is intriguing how our current study supports an important role for another FACIT collagen in the context of cancer, even if the current study provides no clear insight into the mechanisms of this role.
Recently, collagen binding proteins have been used in conjunction with immune checkpoint inhibitors in the form of fusion constructs to target the therapy towards the tumor and limit adverse effects. For example, the collagen-binding domain of von Willebrand Factor fused to checkpoint inhibitor antibodies can concentrate therapy effects towards the fibrotic stroma of the tumor microenvironment [
40]. Minor collagens, which bind fibrillar collagens may be useful in a similar way. The interaction partners of type XX collagen are not currently known, but if they are similar to type XII collagen, it is possible that they interact with major fibrillar collagens such as type I collagen and may, therefore, be useful as a homing device to deliver therapies to fibrotic tumors.
The limitations of this study are substantial. Sample sizes for both cohorts are small, increasing the likelihood of bias and false positives. Thankfully, effect sizes seemed large, with large fold-increases in PRO-C20 levels in samples from cancer patients compared to healthy controls. Prognostic evaluation of PRO-C20 was also limited by the small sample size and the dichotomization of biomarker levels, which can introduce bias [
41]. Clinical information was especially lacking for the first cohort, limiting the analysis to a basic comparison between indications with little consideration for their clinical outcome. This underlines the need to validate the current findings in a secondary and well-characterized cohort. The goal of this study was to demonstrate the biological relevance and biomarker potential of type XX collagen and a robust assay to quantify it in complex samples. This study was merely one of our first steps to explore the interesting biology of FACIT collagens. The next step for the further validation of PRO-C20 involves confirming the findings presented here in another cohort of sufficient size to determine the cutoffs needed in both a diagnostic and prognostic settings. Special attention should be paid to the comparison to controls, which could preferably be individuals with other chronic diseases than cancer to evaluate, e.g., Chronic pancreatitis to PDAC—which is a comparison more applicable to the clinic. Further, how PRO-C20 levels are affected by treatment or other clinically relevant factors should be evaluated. It seems clear, however, that PRO-C20 is a biomarker with the potential to make an impact for patients in the future, and further investigation into PRO-C20 and the biology of the minor collagens is warranted.
4. Materials and Methods
4.1. Generation of Monoclonal Antibodies Targeting the PRO-C20 Epitope
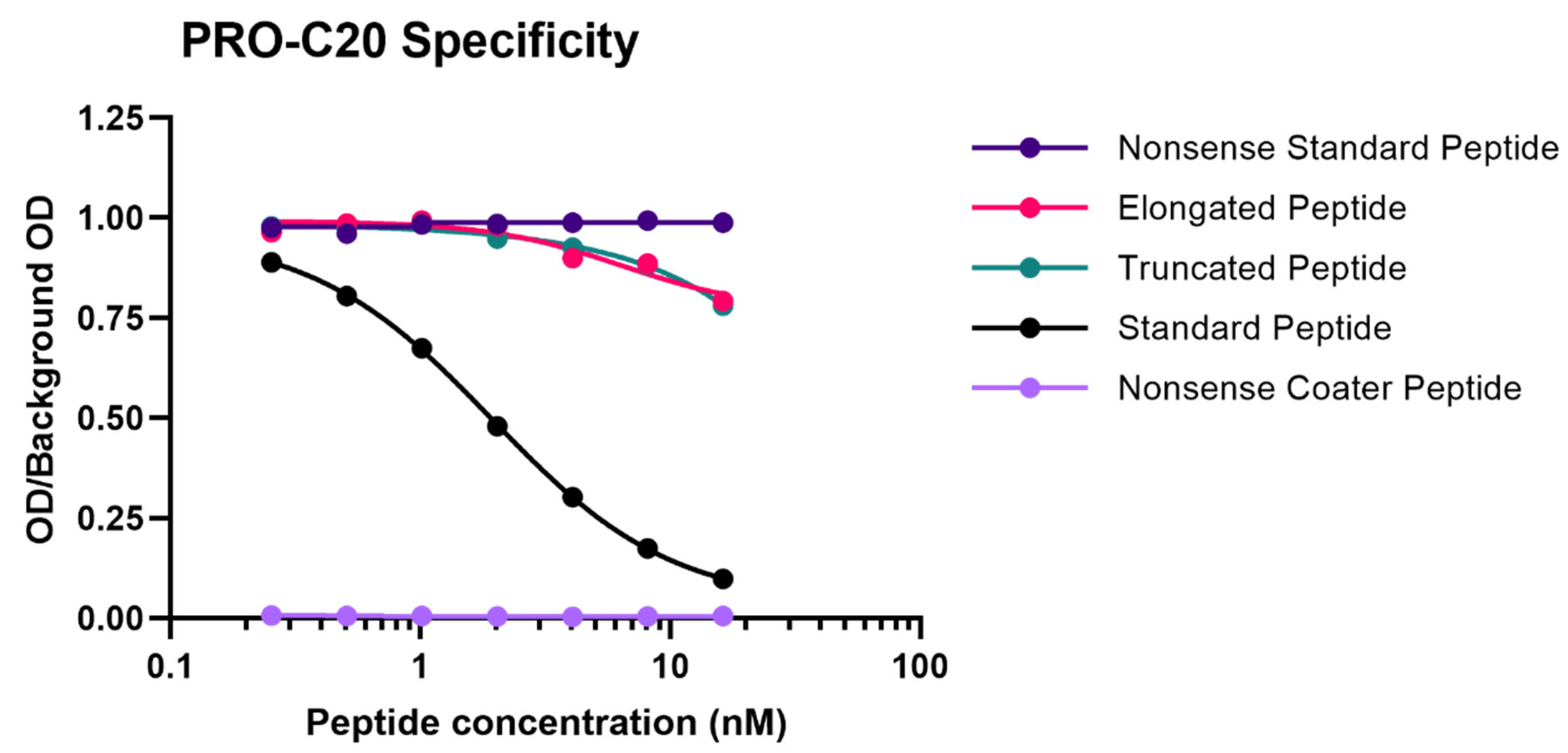
A 10 amino-acid peptide 1275QGASTQGLWE1284 corresponding to the C-terminus of type XX collagen (UniprotKB: Q9P218) was purchased from Genscript (Piscataway, NJ, USA) and used for immunization. This sequence was incorporated into an immunogenic peptide (KLH-CGG-QGASTQGLWE) by covalently cross-linking the target peptide to Keyhole Limpet Hemocyanin (KLH) carrier protein using sulfosuccinimidyl 4-(N-maleimidomethyl) cyclohexane-1-carboxylate (SMCC, Thermo Scientific, Waltham, MA, USA, cat. no. 22322). Glycine and cysteine residues were added at the N-terminal end of the peptide to ensure correct linking of the carrier protein. Monoclonal antibodies were generated by subcutaneous immunization of six-week-old Balb/C mice with 200 µL emulsified antigen containing 100 µg immunogenic peptide mixed with Sigma Adjuvant System (Sigma cat. no. S6322). Consecutive immunizations were performed at 2-week intervals until stable sera titer levels were reached. The mouse with the highest titer was rested for four weeks and was then boosted with 100 µg immunogenic peptide in 100 µL 0.9% NaCl solution intravenously. Hybridoma cells were produced by fusing spleen cells with SP2/0 myeloma cells. The resultant hybridoma cells were then cultured in 96-well microtiter plates and limited dilution was used to secure monoclonal growth. The best antibody clone for the epitope of interest was selected based on a preliminary competitive ELISA for the reactivity towards the selection peptide (the target peptide, QGASTQGLWE), and not an elongated peptide (QGASTQGLWES), a truncated peptide (QGASTQGLW), or a non-sense KLH-conjugated peptide (IRQCPDRTYG-GGC-KLH). The monoclonal antibodies were purified using protein-G-columns according to the manufacturer’s instructions (GE Healthcare Life Sciences, Little Chalfont, UK, cat. no. 17-0404-01). The purified antibodies were labeled with horseradish peroxidase (HRP) using a peroxidase labeling kit (Roche Diagnostics GmbH, Mannheim, Germany, cat. no. 11829696001).
4.2. PRO-C20 ELISA Protocol
Several optimizations were made to the ELISA, including the choice of assay buffer, incubation time and temperature, as well as concentrations of antibody and peptides. The final PRO-C20 protocol was performed as follows: a 96-well streptavidin-coated ELISA plate was coated with 100 µL/well of 1.25 ng/mL biotinylated QGASTQGLWE peptide dissolved in assay buffer (25 mM tris-buffered saline (TBS), 1% bovine serum albumin (BSA) (w/v), 0.1% Tween-20 (w/v), 2 g/L NaCl, pH 8.0) and incubated for 30 min at 20 °C with shaking at 300 RPM. After washing five times with washing buffer (25 mM Tris, 50 mM NaCl, pH 7.2), 20 µL/well of sample was added in duplicates followed by 100 µL/well of 50 ng/mL HRP-labelled monoclonal antibody in assay buffer and incubated for 1 h at 20 °C with shaking at 300 RPM. After a second washing cycle, 100 µL/well of 3, 3′, 5, 5′-tetramethylbenzidine (TMB) was added and incubated for 15 min in darkness at 20 °C with shaking at 300 RPM. The reaction was stopped by adding 100 µL/well of 1% H2SO4 (v/v). Absorbance was measured at 450 nm with 650 nm as reference. To generate a standard curve, 20 µL/well of 50 ng/mL RHLEGRGEPGAVGQMGSPGQQGASTQGLWE peptide, serially diluted twofold, was added to appropriate wells and a four-parametric logistic regression model was used to fit a curve. Each plate included five quality control samples comprising one human serum, one horse serum, one human plasma, and two standard peptide in assay buffer samples to monitor intra- and inter-assay variation.
4.3. Technical Validation of the PRO-C20 ELISA
Antibody specificity was evaluated by the inhibition of signal by two different versions of the standard peptide: a 10 amino acid version (QGASTQGLWE) and 30 amino acid version (RHLEGRGEPGAVGQMGSPGQQGASTQGLWE), both tested in twofold dilution series. In addition, an elongated (QGASTQGLWES) and truncated peptide (QGASTQGLW) of the PRO-C20 epitope was tested, as well as a non-sense standard peptide (SHAHQRTGGN) and a non-sense coater peptide (Biotin-SHAHQRTGGN) with sequences corresponding to the C-terminus of type XIX collagen. Linearity of dilution was evaluated by twofold dilutions of human serum samples and then calculating the percentage recovery of the measured concentration relative to the predicted concentration. Accuracy was evaluated by either spiking a known quantity of the standard peptide into human serum samples or by spiking one human serum sample into another human serum sample at different ratios (100:0, 75:25, 50:50, 25:75, or 0:100), followed in both cases by calculating the percentage recovery of the spiked sample relative to the non-spiked sample. The influence of commonly interfering substances including hemoglobin, lipids, and biotin were evaluated by spiking human serum samples with a known quantity of the interfering substances (hemoglobin low = 2.5 mg/mL, high = 5 mg/mL; lipids low = 1.5 mg/mL, high = 5 mg/mL; and biotin low = 5 ng/mL, high = 100 ng/mL) and calculating the percentage recovery relative to the non-spiked sample. Assay variation was tested by running ten independent runs of the assay using ten quality control samples in double determinations. Five quality control samples were one human serum, one horse serum, one human plasma, and five samples of standard peptide in assay buffer of varying concentrations. Intra-assay variation was calculated as the mean coefficient of variance (CV%) between double determinations during each run of the assay. Inter-assay variation was calculated as the mean CV% between all determinations across all ten runs. Lower- and upper-limit of the measurement range were determined as the concentrations that denote the limits of the linear range of the assay, defined as the intersection between the line drawn through the standard points that comprise the linear range of the assay and the line drawn horizontally at either the maximum or minimum OD. Lower limit of detection was calculated as the mean interpolated concentration of 21 blank samples only containing assay buffer plus three standard deviations. Upper limit of detection was calculated as the mean interpolated concentration of standard peptide corresponding to the highest concentration of the standard curve minus three standard deviations. Analyte stability was evaluated for three human serum samples incubated at either 4 or 20 °C for 2, 4, 24, or 48 h and calculating the percentage recovery of the incubated samples relative to the corresponding control sample kept at −20 °C. Freeze–thaw stability was evaluated by repeatedly freezing and thawing human serum samples for up to 4 cycles and calculating the percentage recovery of the cycled samples relative to the corresponding control samples that underwent a single freeze–thaw cycle.
4.4. PRO-C1 ELISA Protocol
The PRO-C1 ELISA (Nordic Bioscience, Herlev, Denmark, cat. no. 2800) measures the N-terminal pro-peptide of type I collagen, sometimes called PINP in the literature. PRO-C1 levels reflect type I collagen formation, as the release of the pro-peptide is a critical step in the maturation process of type I collagen. A detailed description and protocol of the ELISA has already been published [
42].
4.5. Patient Samples
The first cohort included serum samples from 219 patients with cancer and 33 healthy controls. It included 10 groups, each with 20 patients with bladder, breast, colorectal, head and neck, kidney, lung, pancreatic, prostate, and stomach cancer or melanoma. In addition, 19 patients with ovarian cancer and 33 age-matched healthy controls were included. Serum samples from cancer patients and healthy controls were obtained from Proteogenex (Los Angeles, CA, USA) and BioIVT (Westbury, NY, USA), respectively. Samples were stored at −80 °C prior to analysis. A summary of the cohort characteristics can be found in
Table 2. According to the vendors, sample collection was approved by an Institutional Review Board or Independent Ethical Committee and patients gave their informed consent: Russian Oncological Research Centre n.a. Blokhin RAMS (PG-ONC 2003/1) and Western Institutional Review Board, Inc. (WIRB
®Protocol #20161665). All investigations were carried out according to the Helsinki Declaration.
The second cohort included serum samples from 36 patients with PDAC, 11 patients with chronic pancreatitis, and 20 healthy controls. Serum samples from healthy controls were obtained from Valley Biomedical (Winchester, VA, USA). Samples were stored at −80 °C prior to analysis. All patients with PDAC and chronic pancreatitis were included in the Danish BIOPAC study (
ClinicalTrials.gov ID: NCT03311776) and were collected prospectively prior to treatment. A detailed description of the cohort has already been published elsewhere [
43]. A summary of the cohort characteristics can be found in
Table 4.
4.6. Statistics
Comparisons of biomarker levels across groups were investigated using ordinary one-way ANOVA. In cohort 1, ANOVA was followed by pair-wise comparisons to the control group using the Dunnett test. In cohort 2, ANOVA was followed by Tukey’s Honestly Significant Difference Test. Differences in PRO-C20 levels across cancer stages were evaluated by ordinary one-way ANOVA. Diagnostic accuracy was tested by ROC curve analysis, including the area under the curve as well as sensitivity, specificity, positive predictive value, and negative predictive value at the PRO-C20 cutoff where the Youden-index was maximized. In the survival analysis, a cut-off value was defined using the maximally selected rank statistic and used to stratify patients into PRO-C20 high and low groups. Survival of these two groups was estimated using Kaplan–Meier curves. Cox proportional hazards regression was used to model survival and estimate the hazard ratios. A
p-value below 0.05 was considered statistically significant. Asterisks indicate the following significance levels: *
p < 0.05; **
p < 0.01; ***
p < 0.001; and ****
p < 0.0001. Statistical analysis and graphs were compiled in GraphPad Prism (version 9.1.0 for Windows, GraphPad Software, San Diego, CA, USA,
www.graphpad.com, accessed on 17 March 2022) and R version 4.0.4 (R Core Team (2021), R Foundation for Statistical Computing, Vienna, Austria,
https://www.R-project.org, accessed on 17 March 2022).


 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}