
Physical Forces in Glioblastoma Migration: A Systematic Review

 , ,
, ,
Abstract
:1. Introduction
2. Results


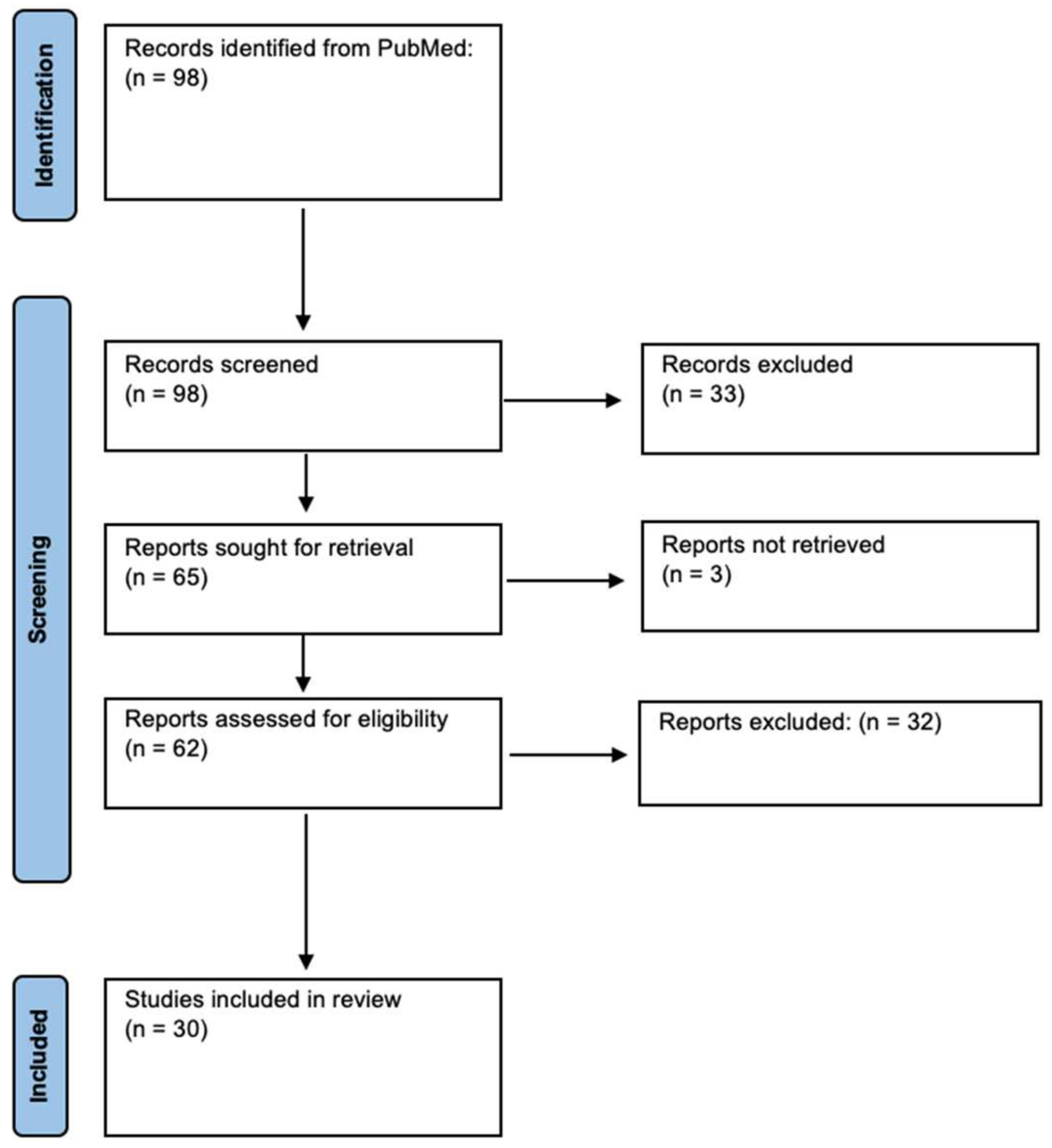{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Author | Stress Marker | Study Design | Effect on GBM | |
|---|---|---|---|---|
| Stiffness | Chen et al. [6] | Piezo/PIEZO1 | Drosophilia glioma model in vivo; mice xenograft experiments; RNA sequencing of two human GBM stem cell lines (G508 and G532) | Regulator of mitosis and tissue stiffness through activation of integrin-FAK signaling; correlated with GBM aggressiveness and decreased survival |
| Miroshnikova et al. [7] | Tenascin C | Patient-derived samples; mouse model | ECM stiffness represses miR-203 expression which activates HIF1α-dependent TNC deposition, which may induce aggressiveness and lead to recurrence | |
| Sen et al. [8] | Talin-1 | U373 MG human glioma cells | Involved in mechanical rigidity sensing; transmits signals from the ECM to the cytoskeleton through interplay of integrins and actin | |
| Khan et al. [9] | N/A | CD 133+ GBM cells | Actively migrating GBMs exhibit higher elastic stiffness at the front end, facilitating traction needed for forward movement through an anchoring effect | |
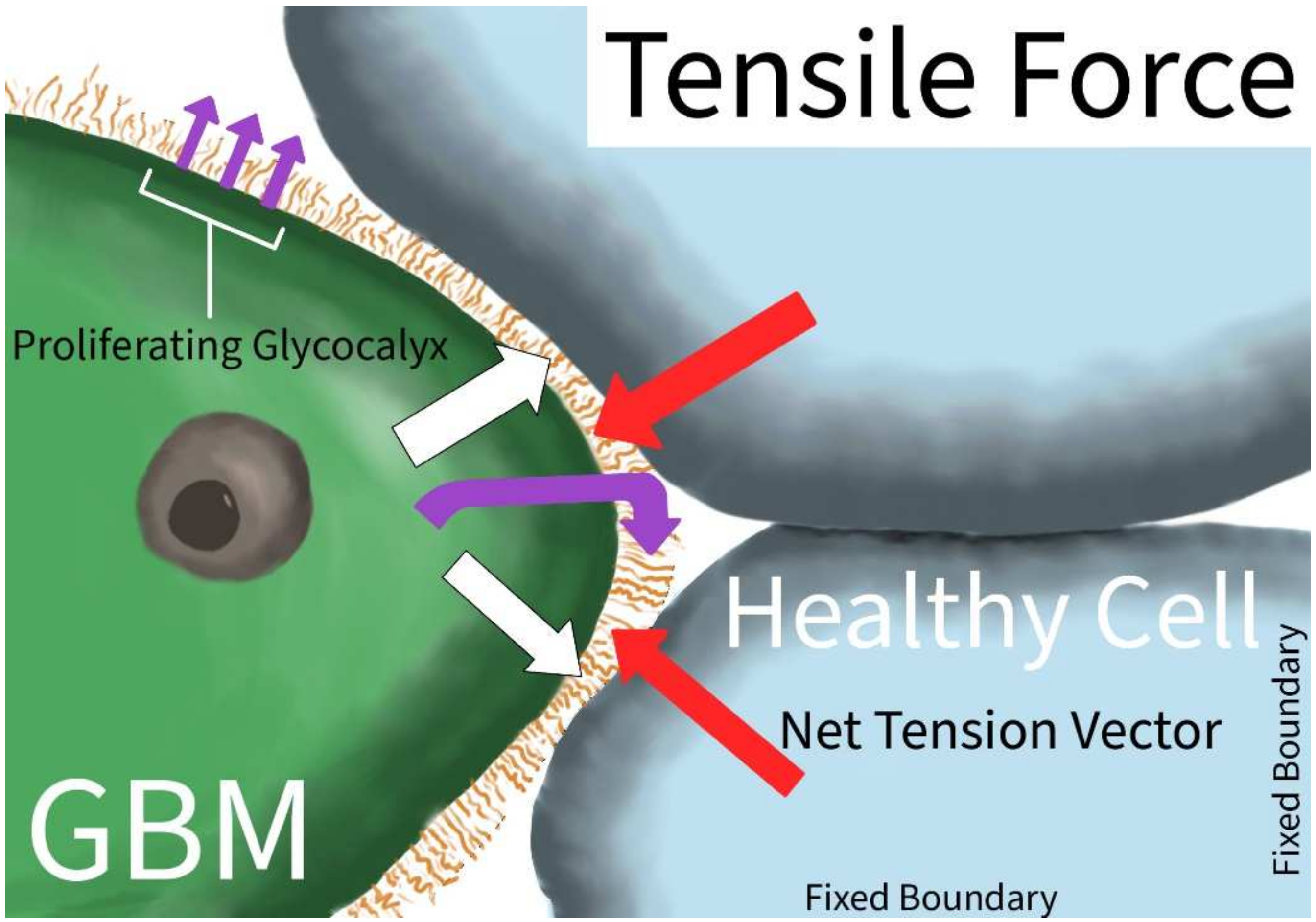
| Tensile Force | Barnes et al. [10] | Tension (tenescin) | Patient-derived samples; mouse model | Tension-mediated glycocalyx–integrin feedback loop which promotes mesenchymal characteritistics |
| Shen et al. [2] | Yes-associated protein (YAP) | G55 GBM cells | Re-localization of YAP to the cell nucleus indicates a higher degree of cytoskeletal tension during migration of GBM cells in a physically confined environment | |
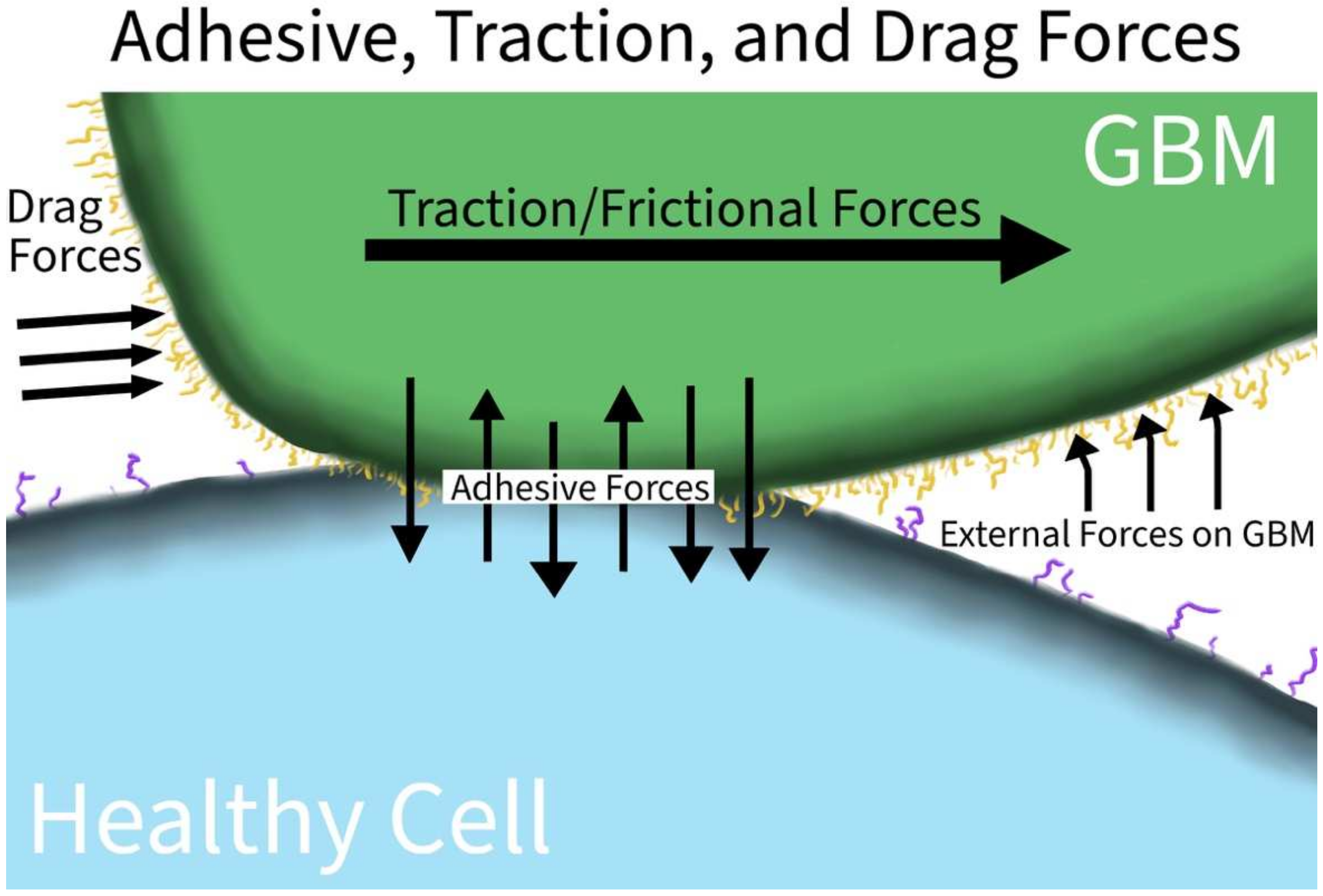
| Traction | de Semir et al. [11] | Pleckstrin homology domain-interacting protein (PHIP) | In vitro and in vivo murine model of U-251 GBM cell lines | Plays a role in activating the actin cytoskeleton, focal adhesion dynamics, migration, and invasion |
| Gordon et al. [12] | Latex beads displacement and cell line volumetric growth | In vitro using human U87MGmEGFR GBM cell line | Demonstrated that tumor cells will grow towards the path of least resistance through traction-mediated forces | |
| Drag Force | Agosti et al. [13] | N/A | U87 GBM cell lines | During proliferation, GBM aggregation is enabled when the adhesive force between cells is of the same magnitude of the drag forces of cells as they expand |
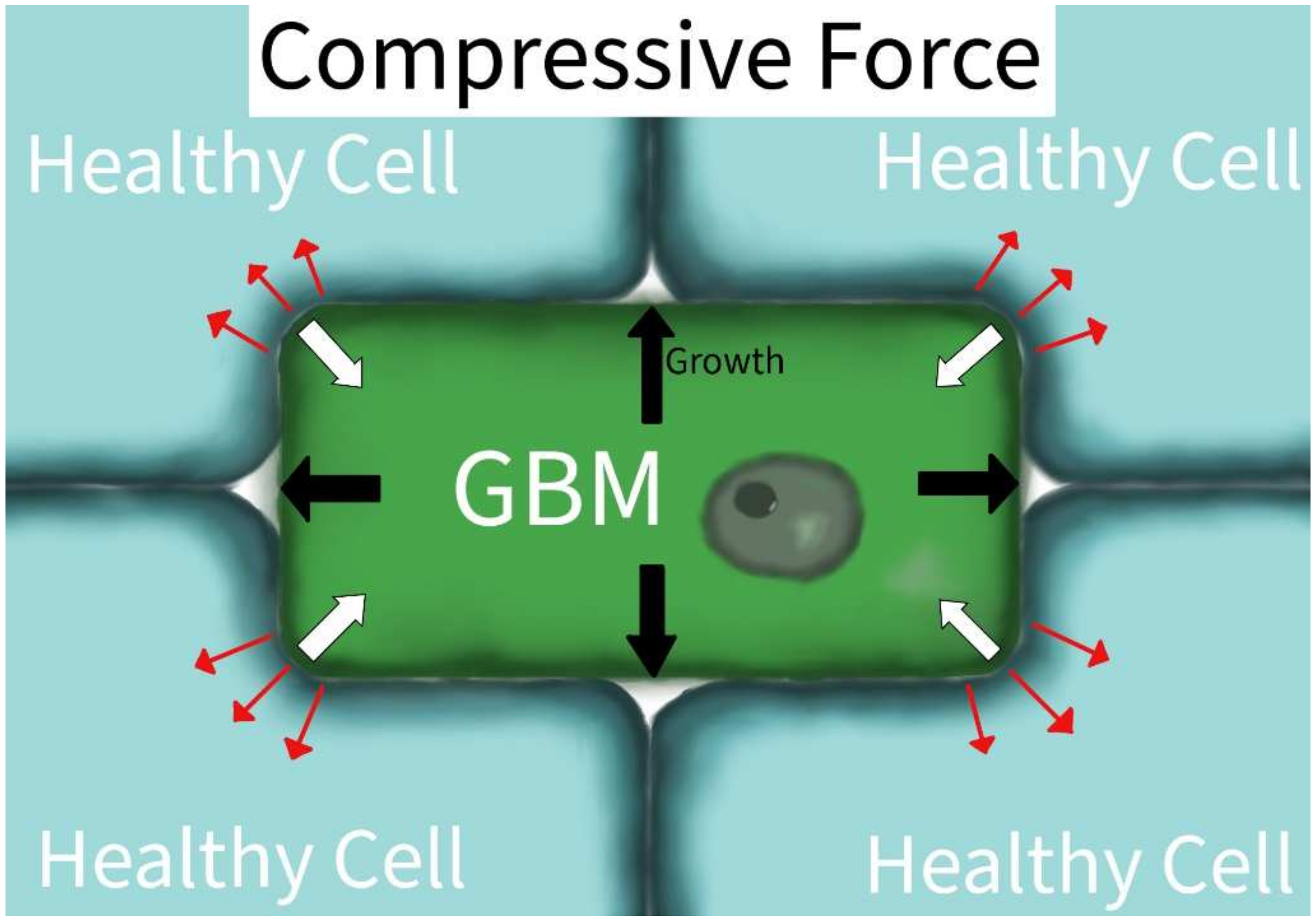
| Compression | Voutouri et al. [14] | Vessel option | Mathematical model | Compression led to hypoxia and resultant angiogenesis |
| Calhoun et al. [15] | miR548 family | LN229 and U251 GBM cell lines; pathway analysis | Increased migration and decreased proliferation, characteristics associated with tumor aggressiveness | |
| Demou et al. [16] | Caveolin-1, integrin-β1, Rac1 | U87 and HGL21 GBM cells | Cell deformation/compression leads to downregulation of E-cadherin (CDH1) and PECAM-1 (CD31) and overexpression of PTEN and Rac1; resultant decrease in cell adhesion and increased migration | |
| Adhesion | Morjen et al. [17] | Kunitz-typeprotease inhibitor (PIVL) | In vitro using U87 cell lines; in vivo mouse model | Disrupted GBM migration, invasion, and adhesion through inhibition of integrin |
| Yao et al. [18] | P311/PTZ17 | In vivo mouse model | Rho GTPase-mediated promotion of migration of epidermal stem cells | |
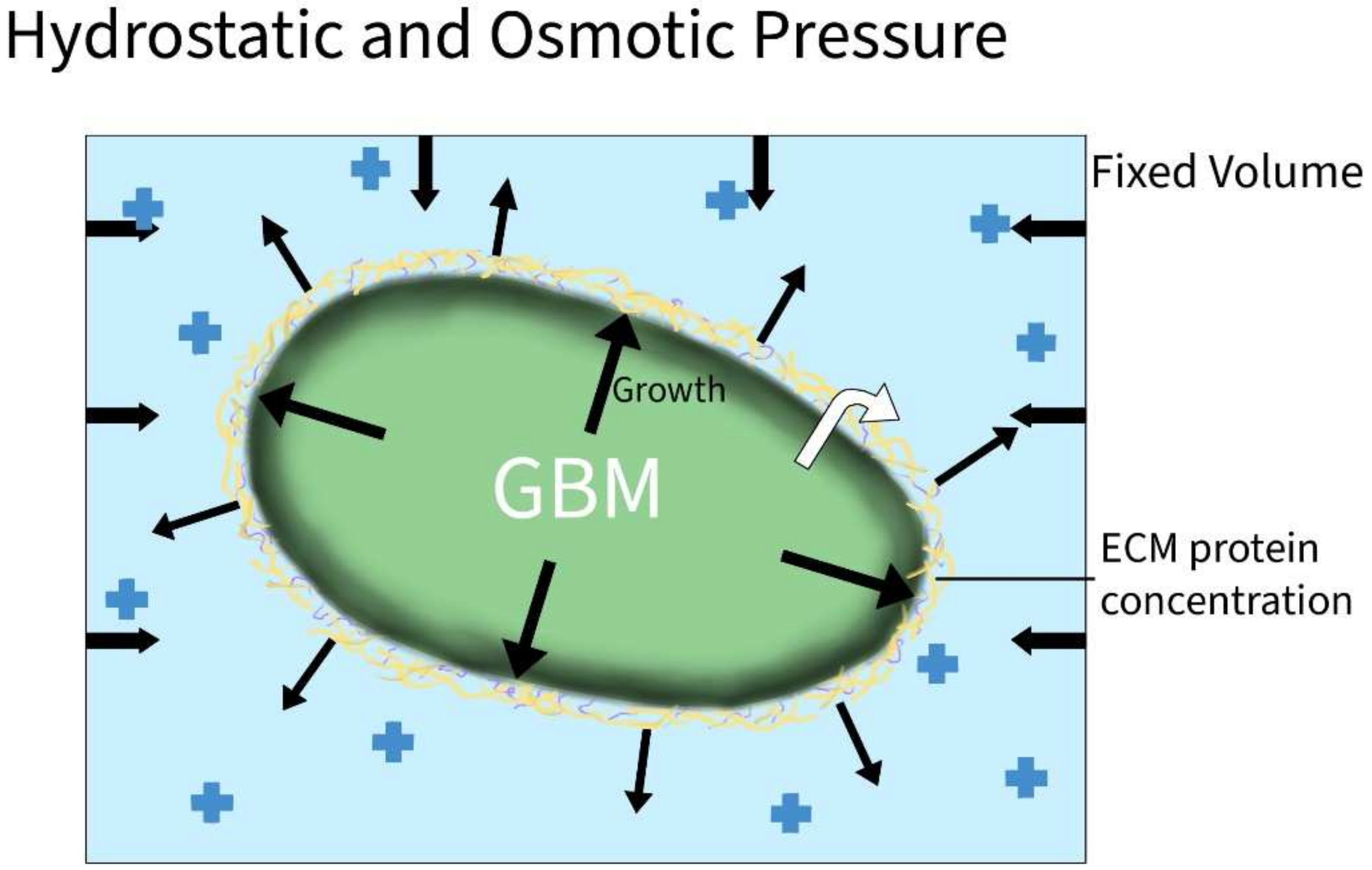
| Hydrostatic Pressure | Claus et al. [19] | N/A | Case report | Increased CSF protein concentration caused increased ICP and patient deterioration |
| Takara et al. [20] | N/A | Case report | Increased CSF protein concentration led to hydrostatic pressure build up | |
| Zoi et al. | Polycystin-1 (PC1) | T98G GBM cells subjected to coninuous hydrostatic pressure and/or PC1 blockade | Hydrostatic pressure inhibited proliferation and migration of GBM cells. PC1 had the opposite effect | |
| Magnetic Force | Perez et al. [21] | N/A | (U87) tumor spheroid aggregation methodology based on magnetic cell labeling; spheroid cell invasion w/ Matrigel | Magnetic properties of the spheroids allow for determination of surface tension |
| Chen et al. [22] | Hexagonal superparamagnetic cones | U-343 GBM cell lines | Magnetic field gradient decreased cell growth and migration | |
| Osmotic Pressure | Catacuzzeno et al. [23] | Swelling-activated chloride currents | In vitro using GL-15 GBM cells | Channel activation included shape and volume changes, allowing migration and invasion |
| Pu et al. [24] | Caveolin-1, CAVIN1; uPA and MMPs; AQP1 | U87, U118, and U251 GBM cell lines | Play a role in the response to increased pressure and GBM invasion | |
| Pu et al. [25] | Snail-1, Snail-2, N-cadherin, Twist, and vimentin | GBM cell lines U87 and U251; patient-derived neural oncospheres | EMT and invasion through production of matrix proteases as a response to osmotic/hydrostatic pressure | |
| Shear Stress | Rezk et al. [26] | Nestin and vimentin; actin filaments, vinculin, paxillin, and FAK | Patient-derived samples | Increased migration and proliferation |
| Solid Stress | Ciarletta et al. [27] | N/A | Theoretical calculation of buckling instability from solid stresses | Residual stresses promote buckling instability and promote tumor invasion |
| Stylianopoulos et al. [28] | Collagen, hyaluronan | Mathematical model | Increased perfusion of tumors led to improved oxygenation and drug delivery |
| Author | Stress Marker | Study Design | Effect on GBM | |
|---|---|---|---|---|
| Cellular Volume | Fischer et al. [29] | HAMLET | In vitro using non-transformed human astrocytes CC-2565; in vivo animal models using human GBM xenografted rat models | HAMLET selectively induced GBM apoptosis in rat xenograft models via activation of programmed cell death. HAMLET did not interact with healthy tissue and extended survival by relieving GBM pressure symptoms via volume reduction |
| Sforna et al. [30] | Swelling-activated chloride currents | In vitro using GL-15, U87MG, and U251 cells lines | Acute and cyclic hypoxic conditions (either blood flow interuptions) may enable GBM cells to upregulate I(Cl,swell) conditions, which regulate the cellular volume and prevent cellular death | |
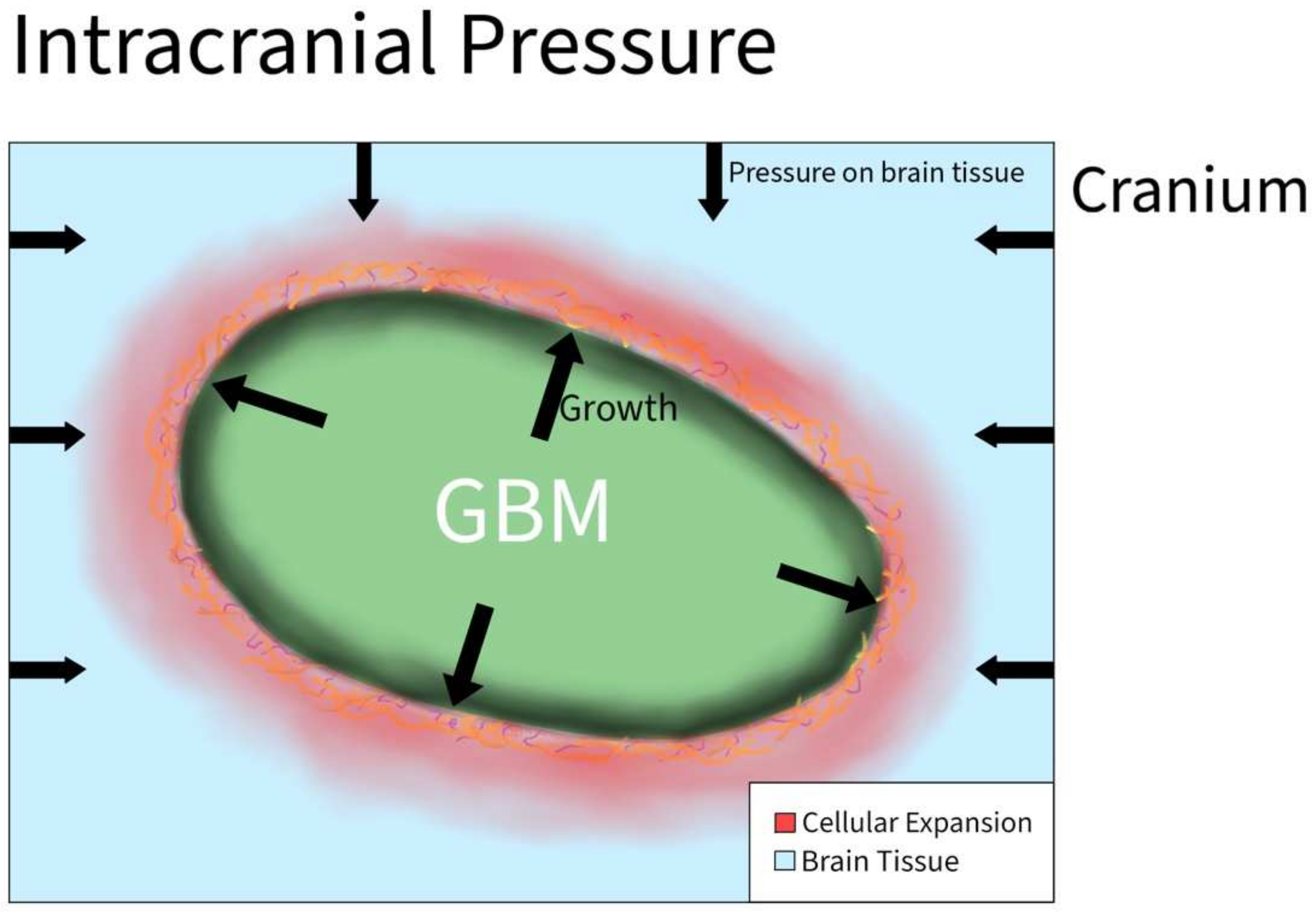
| Intracranial Pressure | Chida et al. [31] | N/A | Case report | Increased high CSF protein and pressure hypothesized to cause aggressive phenogype |
| Rifikinson-Mann et al. [32] | N/A | Case series of hydrocephalus associated with intramedullary spinal GBM | Malignant tumors were associated with tumor extension and ventriculomegaly | |
| Yoo et al. [33] | Hyaluronic acid | U87MG, U373MG, and U251MG glioma cells; transwell assay | In response to radiation, HA production was increased in GBM cells by HA synthase-2 (HAS2), which was transcriptionally upregulated by NF-ĸB. Notably, NF-ĸB was persistently activated by an IL-1α-feedback loop, making HA abundant in tumor microenvironment after radiation |
3. Discussion
3.1. External/Applied/Mechanical Forces
3.1.1. Tensile Force and Stiffness
3.1.2. Compressive Force
3.1.3. Adhesive, Traction, and Drag Forces
3.1.4. Hydrostatic and Osmotic Pressure
3.2. Tumor Microenvironment (TME)
3.2.1. Intracranial Pressure
3.2.2. Cellular Volume
3.2.3. Hydraulic Conductivity
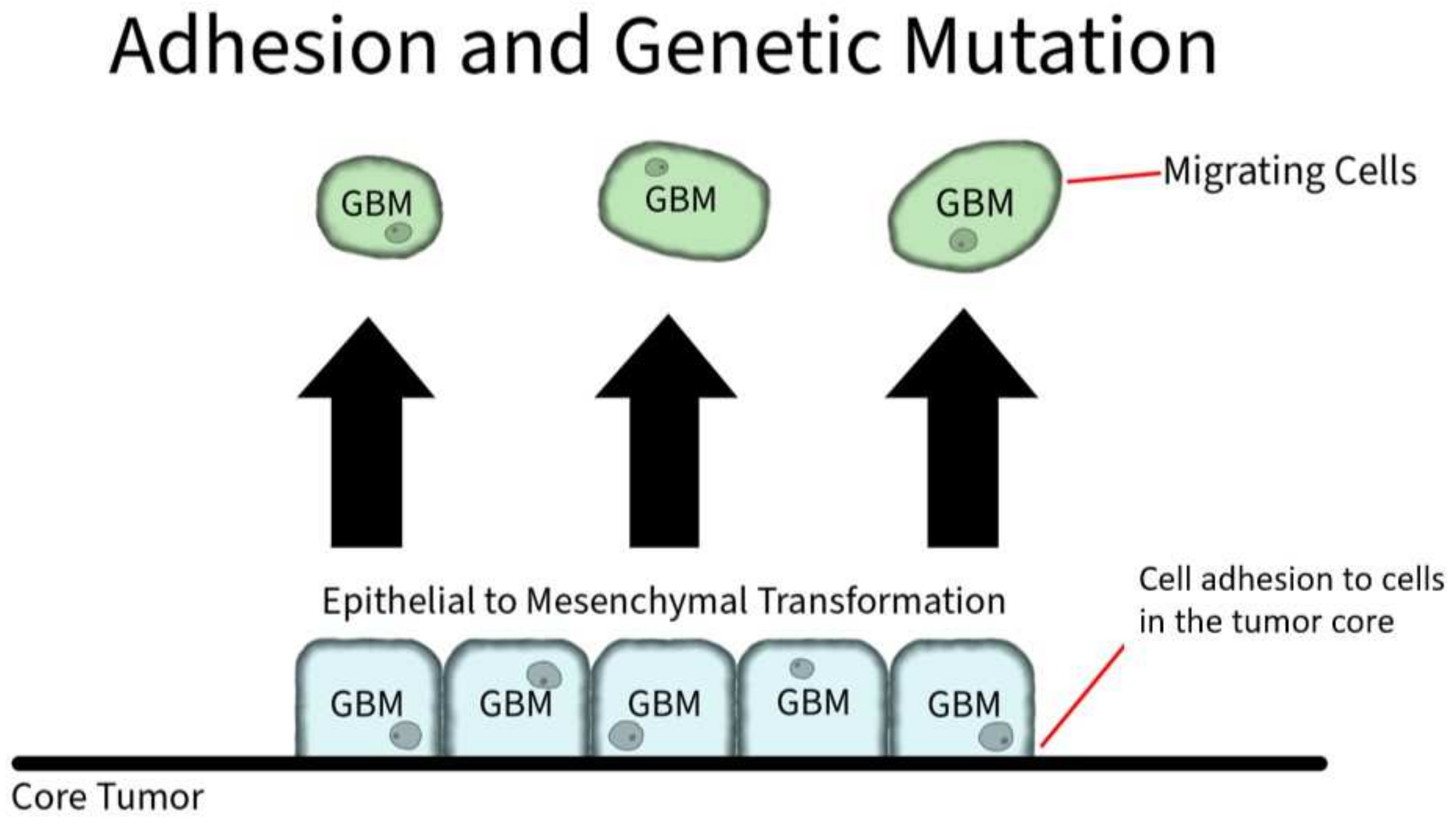
3.2.4. Adhesion Protein Expression
3.3. Major Molecular Mechanisms Associated with Physical Forces in GBM
4. Material and Methods
4.1. Search Strategy
4.2. Eligibility Criteria
4.3. Data Extraction
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Spill, F.; Reynolds, D.S.; Kamm, R.D.; Zaman, M.H. Impact of the physical microenvironment on tumor progression and metastasis. Curr. Opin. Biotechnol. 2016, 40, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Shen, Q.; Hill, T.; Cai, X.; Bui, L.; Barakat, R.; Hills, E.; Almugaiteeb, T.; Babu, A.; Mckernan, P.H.; Zalles, M.; et al. Physical confinement during cancer cell migration triggers therapeutic resistance and cancer stem cell-like behavior. Cancer Lett. 2021, 506, 142–151. [Google Scholar] [CrossRef]
- Mierke, C.T. The matrix environmental and cell mechanical properties regulate cell migration and contribute to the invasive phenotype of cancer cells. Rep. Prog. Phys. 2019, 82, 064602. [Google Scholar] [CrossRef]
- Muldoon, L.L.; Alvarez, J.I.; Begley, D.J.; Boado, R.J.; Del Zoppo, G.J.; Doolittle, N.D.; Engelhardt, B.; Hallenbeck, J.M.; Lonser, R.R.; Ohlfest, J.R.; et al. Immunologic privilege in the central nervous system and the blood-brain barrier. J. Cereb. Blood Flow Metab. 2013, 33, 13–21. [Google Scholar] [CrossRef] [Green Version]
- Van Tellingen, O.; Yetkin-Arik, B.; De Gooijer, M.C.; Wesseling, P.; Wurdinger, T.; De Vries, H.E. Overcoming the blood-brain tumor barrier for effective glioblastoma treatment. Drug Resist. Updates 2015, 19, 1–12. [Google Scholar] [CrossRef]
- Chen, X.; Wanggou, S.; Bodalia, A.; Zhu, M.; Dong, W.; Fan, J.J.; Yin, W.C.; Min, H.; Hu, M.; Draghici, D.; et al. A feedforward mechanism mediated by mechanosensitive ion channel PIEZO1 and tissue mechanics promotes glioma aggression. Neuron 2018, 100, 799–815. [Google Scholar] [CrossRef] [Green Version]
- Miroshnikova, Y.A.; Mouw, J.K.; Barnes, J.M.; Pickup, M.W.; Lakins, J.N.; Kim, Y.; Lobo, K.; Persson, A.I.; Reis, G.F.; McKnight, T.R.; et al. Tissue mechanics promote IDH1-dependent HIF1α-tenascin C feedback to regulate glioblastoma aggression. Nat. Cell Biol. 2016, 18, 1336–1345. [Google Scholar] [CrossRef] [PubMed]
- Sen, S.; Ng, W.P.; Kumar, S. Contributions of talin-1 to glioma cell-matrix tensional homeostasis. J. R. Soc. Interface 2012, 9, 1311–1317. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.; Bui, L.; Bachoo, R.; Kim, Y.T.; Chuong, C.J. Differences in creep response of GBM cells migrating in confinement. Int. Biomech. 2020, 7, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Barnes, J.M.; Kaushik, S.; Bainer, R.O.; Sa, J.K.; Woods, E.C.; Kai, F.; Przybyla, L.; Lee, M.; Lee, H.W.; Tung, J.C.; et al. A tension-mediated glycocalyx-integrin feedback loop promotes mesenchymal-like glioblastoma. Nat. Cell Biol. 2018, 20, 1203–1214. [Google Scholar] [CrossRef] [PubMed]
- De Semir, D.; Bezrookove, V.; Nosrati, M.; Scanlon, K.R.; Singer, E.; Judkins, J.; Rieken, C.; Wu, C.; Shen, J.; Schmudermayer, C.; et al. PHIP drives glioblastoma motility and invasion by regulating the focal adhesion complex. Proc. Natl. Acad. Sci. USA 2020, 117, 9064–9073. [Google Scholar] [CrossRef]
- Gordon, V.D.; Valentine, M.T.; Gardel, M.L.; Andor-Ardó, D.; Dennison, S.; Bogdanov, A.A.; Weitz, D.A.; Deisboeck, T.S. Measuring the mechanical stress induced by an expanding multicellular tumor system: A case study. Exp. Cell Res. 2003, 289, 58–66. [Google Scholar] [CrossRef]
- Agosti, A.; Marchesi, S.; Scita, G.; Ciarletta, P. Modelling cancer cell budding in-vitro as a self-organised, non-equilibrium growth process. J. Theor. Biol. 2020, 492, 110203. [Google Scholar] [CrossRef]
- Voutouri, C.; Kirkpatrick, N.D.; Chung, E.; Mpekris, F.; Baish, J.W.; Munn, L.L.; Fukumura, D.; Stylianopoulos, T.; Jain, R.K. Experimental and computational analyses reveal dynamics of tumor vessel cooption and optimal treatment strategies. Proc. Natl. Acad. Sci. USA 2019, 116, 2662–2671. [Google Scholar] [CrossRef] [Green Version]
- Calhoun, M.A.; Cui, Y.; Elliott, E.E.; Mo, X.; Otero, J.J.; Winter, J.O. MicroRNA-mRNA interactions at low levels of compressive solid stress implicate mir-548 in increased glioblastoma cell motility. Sci. Rep. 2020, 10, 311. [Google Scholar] [CrossRef]
- Demou, Z.N. Gene expression profiles in 3D tumor analogs indicate compressive strain differentially enhances metastatic potential. Ann. Biomed. Eng. 2010, 38, 3509–3520. [Google Scholar] [CrossRef]
- Morjen, M.; Kallech-Ziri, O.; Bazaa, A.; Othman, H.; Mabrouk, K.; Zouari-Kessentini, R.; Sanz, L.; Calvete, J.J.; Srairi-Abid, N.; El Ayeb, M.; et al. PIVL, a new serine protease inhibitor from Macrovipera lebetina transmediterranea venom, impairs motility of human glioblastoma cells. Matrix Biol J. Int. Soc. Matrix Biol. 2013, 32, 52–62. [Google Scholar] [CrossRef]
- Yao, Z.; Li, H.; He, W.; Yang, S.; Zhang, X.; Zhan, R.; Xu, R.; Tan, J.; Zhou, J.; Wu, J.; et al. P311 accelerates skin wound reepithelialization by promoting epidermal stem cell migration through RhoA and Rac1 activation. Stem. Cells Dev. 2017, 26, 451–460. [Google Scholar] [CrossRef]
- Claus, D.; Sieber, E.; Engelhardt, A.; Rechlin, T.; Neubauer, U.; Volk, B. Ascending central nervous spreading of a spinal astrocytoma. J. Neurooncol. 1995, 25, 245–250. [Google Scholar] [CrossRef]
- Takara, E.; Ide, M.; Yamamoto, M.; Imanaga, H.; Jimbo, M.; Imai, M. Case of intracranial and spinal dissemination of primary spinal glioma. No Shinkei Geka. 1985, 13, 301–305. [Google Scholar]
- Perez, J.E.; Nagle, I.; Wilhelm, C. Magnetic molding of tumor spheroids: Emerging model for cancer screening. Biofabrication 2020, 13, 015018. [Google Scholar] [CrossRef]
- Chen, Y.; Hu, Z.; Zhao, D.; Zhou, K.; Huang, Z.; Zhao, W.; Yang, X.; Gao, C.; Cao, Y.; Hsu, Y.; et al. Self-assembled hexagonal superparamagnetic cone structures for fabrication of cell cluster arrays. ACS Appl. Mater. Interfaces 2021, 13, 10667–10673. [Google Scholar] [CrossRef]
- Catacuzzeno, L.; Michelucci, A.; Sforna, L.; Aiello, F.; Sciaccaluga, M.; Fioretti, B.; Castigli, E.; Franciolini, F. Identification of key signaling molecules involved in the activation of the swelling-activated chloride current in human glioblastoma cells. J. Membr. Biol. 2014, 247, 45–55. [Google Scholar] [CrossRef]
- Pu, W.; Qiu, J.; Nassar, Z.D.; Shaw, P.N.; McMahon, K.; Ferguson, C.; Parton, R.G.; Riggins, G.J.; Harris, J.M.; Parat, M. A role for caveola-forming proteins caveolin-1 and CAVIN1 in the pro-invasive response of glioblastoma to osmotic and hydrostatic pressure. J. Cell Mol. Med. 2020, 24, 3724–3738. [Google Scholar] [CrossRef] [PubMed]
- Pu, W.; Qiu, J.; Riggins, G.J.; Parat, M.O. Matrix protease production, epithelial-to-mesenchymal transition marker expression and invasion of glioblastoma cells in response to osmotic or hydrostatic pressure. Sci. Rep. 2020, 10, 2634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezk, R.; Jia, B.Z.; Wendler, A.; Dimov, I.; Watts, C.; Markaki, A.E.; Franze, K.; Kabla, A.J.; Notes, A. Spatial heterogeneity of cell-matrix adhesive forces predicts human glioblastoma migration. Neuro-Oncol. Adv. 2020, 2, vdaa081. [Google Scholar] [CrossRef] [PubMed]
- Ciarletta, P. Buckling instability in growing tumor spheroids. Phys. Rev. Lett. 2013, 110, 158102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stylianopoulos, T.; Jain, R.K. Combining two strategies to improve perfusion and drug delivery in solid tumors. Proc. Natl. Acad. Sci. USA 2013, 110, 18632–18637. [Google Scholar] [CrossRef] [Green Version]
- Fischer, W.; Gustafsson, L.; Mossberg, A.K.; Gronli, J.; Mork, S.; Bjerkvig, R.; Svanborg, C. Human alpha-lactalbumin made lethal to tumor cells (HAMLET) kills human glioblastoma cells in brain xenografts by an apoptosis-like mechanism and prolongs survival. Cancer Res. 2004, 64, 2105–2112. [Google Scholar] [CrossRef] [Green Version]
- Sforna, L.; Cenciarini, M.; Belia, S.; Michelucci, A.; Pessia, M.; Franciolini, F.; Catacuzzeno, L. Hypoxia modulates the swelling-activated cl current in human glioblastoma cells: Role in volume regulation and cell survival. J. Cell Physiol. 2017, 232, 91–100. [Google Scholar] [CrossRef]
- Chida, K.; Konno, H.; Sahara, M.; Takase, S. Meningeal seeding of spinal cord glioblastoma multiforme without any signs of myelopathy. Rinsho Shinkeigaku Clin. Neurol. 1995, 35, 1235–1240. [Google Scholar]
- Rifkinson-Mann, S.; Wisoff, J.H.; Epstein, F. The association of hydrocephalus with intramedullary spinal cord tumors: A series of 25 patients. Neurosurgery 1990, 27, 749–754, discussion 754. [Google Scholar] [CrossRef]
- Yoo, K.C.; Suh, Y.; An, Y.; Lee, H.; Jeong, Y.J.; Uddin, N.; Cui, Y.; Roh, T.; Shim, J.; Chang, J.H.; et al. Proinvasive extracellular matrix remodeling in tumor microenvironment in response to radiation. Oncogene 2018, 37, 3317–3328. [Google Scholar] [CrossRef]
- Beckmann, R.; Houben, A.; Tohidnezhad, M.; Kweider, N.; Fragoulis, A.; Wruck, C.J.; Brandenburg, L.O.; Hermanns-Sachweh, B.; Goldring, M.B.; Pufe, T.; et al. Mechanical forces induce changes in VEGF and VEGFR-1/sFlt-1 expression in human chondrocytes. Int. J. Mol. Sci. 2014, 15, 15456–15474. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.Y.; Kim, Y.J.; Lee, S.; Park, J.H. The critical role of ERK in death resistance and invasiveness of hypoxia-selected glioblastoma cells. BMC Cancer 2009, 9, 27. [Google Scholar] [CrossRef] [Green Version]
- Luo, X.; Shen, S.; Yi, S.; Hu, J.; Sun, Y.; Gao, K.; Zhu, L. Screening of differentially expressed miRNAs in tensile strain-treated HepG2 cells by miRNA microarray analysis. Mol. Med. Rep. 2020, 21, 2415–2426. [Google Scholar] [CrossRef] [Green Version]
- Blank, M.A.B.; Antaki, J.F. Breast lesion elastography region of interest selection and quantitative heterogeneity: A systematic review and meta-analysis. Ultrasound Med. Biol. 2017, 43, 387–397. [Google Scholar] [CrossRef] [Green Version]
- Dawwas, M.F.; Taha, H.; Leeds, J.S.; Nayar, M.K.; Oppong, K.W. Diagnostic accuracy of quantitative EUS elastography for discriminating malignant from benign solid pancreatic masses: A prospective, single-center study. Gastrointest. Endosc. 2012, 76, 953–961. [Google Scholar] [CrossRef]
- Das, J.; Agarwal, T.; Chakraborty, S.; Maiti, T.K. Compressive stress-induced autophagy promotes invasion of HeLa cells by facilitating protein turnover in vitro. Exp. Cell Res. 2019, 381, 201–207. [Google Scholar] [CrossRef]
- Klymenko, Y.; Wates, R.B.; Weiss-Bilka, H.; Lombard, R.; Liu, Y.; Campbell, L.; Kim, O.; Wagner, D.; Ravosa, M.J.; Stack, M.S. Modeling the effect of ascites-induced compression on ovarian cancer multicellular aggregates. Dis. Model. Mech. 2018, 11, dmm034199. [Google Scholar] [CrossRef] [Green Version]
- Boyle, S.T.; Kular, J.; Nobis, M.; Ruszkiewicz, A.; Timpson, P.; Samuel, M.S. Acute compressive stress activates RHO/ROCK-mediated cellular processes. Small GTPases 2020, 11, 354–370. [Google Scholar] [CrossRef] [Green Version]
- Griffon-Etienne, G.; Boucher, Y.; Brekken, C.; Suit, H.D.; Jain, R.K. Taxane-induced apoptosis decompresses blood vessels and lowers interstitial fluid pressure in solid tumors: Clinical implications. Cancer Res. 1999, 59, 3776–3782. [Google Scholar]
- Acheva, A.; Kärki, T.; Schaible, N.; Krishnan, R.; Tojkander, S. Adipokine leptin co-operates with mechanosensitive Ca2+—Channels and triggers actomyosin-mediated motility of breast epithelial cells. Front. Cell Dev. Biol. 2020, 8, 607038. [Google Scholar] [CrossRef]
- Aifuwa, I.; Giri, A.; Longe, N.; Lee, S.H.; An, S.S.; Wirtz, D. Senescent stromal cells induce cancer cell migration via inhibition of RhoA/ROCK/myosin-based cell contractility. Oncotarget 2015, 6, 30516–30531. [Google Scholar] [CrossRef] [Green Version]
- Alcoser, T.A.; Bordeleau, F.; Carey, S.P.; Lampi, M.C.; Kowal, D.R.; Somasegar, S.; Varma, S.; Shin, S.J.; Reinhart-King, C.A. Probing the biophysical properties of primary breast tumor-derived fibroblasts. Cell Mol. Bioeng. 2015, 8, 76–85. [Google Scholar] [CrossRef]
- Chen, S.; Xu, W.; Kim, J.; Nan, H.; Zheng, Y.; Sun, B.; Jiao, Y. Novel inverse finite-element formulation for reconstruction of relative local stiffness in heterogeneous extra-cellular matrix and traction forces on active cells. Phys. Biol. 2019, 16, 036002. [Google Scholar] [CrossRef]
- Cóndor, M.; Mark, C.; Gerum, R.C.; Grummel, N.C.; Bauer, A.; García-Aznar, J.M.; Fabry, B. Breast cancer cells adapt contractile forces to overcome steric hindrance. Biophys. J. 2019, 116, 1305–1312. [Google Scholar] [CrossRef] [Green Version]
- Denoyelle, C.; Albanese, P.; Uzan, G.; Hong, L.; Vannier, J.; Soria, J.; Soria, C. Molecular mechanism of the anti-cancer activity of cerivastatin, an inhibitor of HMG-CoA reductase, on aggressive human breast cancer cells. Cell Signal. 2003, 15, 327–338. [Google Scholar] [CrossRef]
- Desai, S.; Barai, A.; Bukhari, A.B.; De, A.; Sen, S. α-Actinin-4 confers radioresistance coupled invasiveness in breast cancer cells through AKT pathway. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865, 196–208. [Google Scholar] [CrossRef]
- Fokkelman, M.; Balcıoğlu, H.E.; Klip, J.E.; Yan, K.; Verbeek, F.J.; Danen, E.H.J.; Van De Water, B. Cellular adhesome screen identifies critical modulators of focal adhesion dynamics, cellular traction forces and cell migration behaviour. Sci. Rep. 2016, 6, 31707. [Google Scholar] [CrossRef]
- Giri, A.; Bajpai, S.; Trenton, N.; Jayatilaka, H.; Longmore, G.D.; Wirtz, D. The Arp2/3 complex mediates multigeneration dendritic protrusions for efficient 3-dimensional cancer cell migration. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2013, 27, 4089–4099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraning-Rush, C.M.; Califano, J.P.; Reinhart-King, C.A. Cellular traction stresses increase with increasing metastatic potential. PLoS ONE 2012, 7, e32572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Persson, H.; Adolfsson, K.; Abariute, L.; Borgström, M.T.; Hessman, D.; Åström, K.; Oredsson, S.; Prinz, C.N. Cellular traction forces: A useful parameter in cancer research. Nanoscale 2017, 9, 19039–19044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, F.; Zhang, H.; Huang, J.; Xiong, C. Substrate stiffness coupling TGF-β1 modulates migration and traction force of MDA-MB-231 human breast cancer cells in vitro. ACS Biomater. Sci. Eng. 2018, 4, 1337–1345. [Google Scholar] [CrossRef]
- Massalha, S.; Weihs, D. Metastatic breast cancer cells adhere strongly on varying stiffness substrates, initially without adjusting their morphology. Biomech. Model. Mechanobiol. 2017, 16, 961–970. [Google Scholar] [CrossRef]
- McGrail, D.J.; Mezencev, R.; Kieu, Q.M.N.; McDonald, J.F.; Dawson, M.R. SNAIL-induced epithelial-to-mesenchymal transition produces concerted biophysical changes from altered cytoskeletal gene expression. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2015, 29, 1280–1289. [Google Scholar] [CrossRef] [Green Version]
- Mekhdjian, A.H.; Kai, F.; Rubashkin, M.G.; Prahl, L.S.; Przybyla, L.M.; McGregor, A.L.; Bell, E.S.; Barnes, J.M.; DuFort, C.C.; Ou, G.; et al. Integrin-mediated traction force enhances paxillin molecular associations and adhesion dynamics that increase the invasiveness of tumor cells into a three-dimensional extracellular matrix. Mol. Biol Cell. 2017, 28, 1467–1488. [Google Scholar] [CrossRef]
- Takai, K.; Drain, A.P.; Lawson, D.A.; Littlepage, L.E.; Karpuj, M.; Kessenbrock, K.; Le, A.; Inoue, K.; Weaver, V.M.; Werb, Z. Discoidin domain receptor 1 (DDR1) ablation promotes tissue fibrosis and hypoxia to induce aggressive basal-like breast cancers. Genes Dev. 2018, 32, 244–257. [Google Scholar] [CrossRef]
- Anguiano, M.; Morales, X.; Castilla, C.; Pena, A.R.; Ederra, C.; Martínez, M.; Ariz, M.; Esparza, M.; Amaveda, H.; Mora, M.; et al. The use of mixed collagen-Matrigel matrices of increasing complexity recapitulates the biphasic role of cell adhesion in cancer cell migration: ECM sensing, remodeling and forces at the leading edge of cancer invasion. PLoS ONE 2020, 15, e0220019. [Google Scholar] [CrossRef]
- Biondini, M.; Duclos, G.; Meyer-Schaller, N.; Silberzan, P.; Camonis, J.; Parrini, M.C. RalB regulates contractility-driven cancer dissemination upon TGFβ stimulation via the RhoGEF GEF-H1. Sci. Rep. 2015, 5, 11759. [Google Scholar] [CrossRef] [Green Version]
- Gajula, R.P.; Chettiar, S.T.; Williams, R.D.; Thiyagarajan, S.; Kato, Y.; Aziz, K.; Wang, R.; Gandhi, N.; Wild, A.T.; Vesuna, F.; et al. The twist box domain is required for Twist1-induced prostate cancer metastasis. Mol. Cancer Res. MCR 2013, 11, 1387–1400. [Google Scholar] [CrossRef] [Green Version]
- Galior, K.; Ma, V.P.Y.; Liu, Y.; Su, H.; Baker, N.; Panettieri, R.A., Jr.; Wongtrakool, C.; Salaita, K. Molecular tension probes to investigate the mechanopharmacology of single cells: A step toward personalized mechanomedicine. Adv. Healthc. Mater. 2018, 7, e1800069. [Google Scholar] [CrossRef]
- Cai, H.; Ao, Z.; Wu, Z.; Nunez, A.; Jiang, L.; Carpenter, R.L.; Nephew, K.P.; Guo, F. Profiling cell-matrix adhesion using digitalized acoustic streaming. Anal. Chem. 2020, 92, 2283–2290. [Google Scholar] [CrossRef]
- Zhao, Y.; Lai, H.S.S.; Zhang, G.; Lee, G.B.; Li, W.J. Measurement of single leukemia cell’s density and mass using optically induced electric field in a microfluidics chip. Biomicrofluidics 2015, 9, 022406. [Google Scholar] [CrossRef] [Green Version]
- Kathagen-Buhmann, A.; Schulte, A.; Weller, J.; Holz, M.; Herold-Mende, C.; Glass, R.; Lamszus, K. Glycolysis and the pentose phosphate pathway are differentially associated with the dichotomous regulation of glioblastoma cell migration versus proliferation. Neuro-Oncology 2016, 18, 1219–1229. [Google Scholar] [CrossRef] [Green Version]
- Poignard, C.; Silve, A.; Campion, F.; Mir, L.M.; Saut, O.; Schwartz, L. Ion fluxes, transmembrane potential, and osmotic stabilization: A new dynamic electrophysiological model for eukaryotic cells. Eur. Biophys. J. EBJ 2011, 40, 235–246. [Google Scholar] [CrossRef] [Green Version]
- McEvoy, E.; Han, Y.L.; Guo, M.; Shenoy, V.B. Gap junctions amplify spatial variations in cell volume in proliferating tumor spheroids. Nat. Commun. 2020, 11, 6148. [Google Scholar] [CrossRef]
- Heimer, S.; Knoll, G.; Steixner, C.; Calance, D.N.; Trinh, D.T.; Ehrenschwender, M. Hypertonicity-imposed BCL-XL addiction primes colorectal cancer cells for death. Cancer Lett. 2018, 435, 23–31. [Google Scholar] [CrossRef]
- Ueki, H. Study on changes in extravascular lung water during early postoperative periods in thoracic esophageal cancer—With special emphasis on their relation to postoperative renal function. Zasshi J. Nihon Kyobu Geka Gakkai. 1990, 38, 442–456. [Google Scholar]
- Heimer, S.; Knoll, G.; Neubert, P.; Hammer, K.P.; Wagner, S.; Bauer, R.J.; Jantsch, J.; Ehrenschwender, M. Hypertonicity counteracts MCL-1 and renders BCL-XL a synthetic lethal target in head and neck cancer. FEBS J. 2021, 288, 1822–1838. [Google Scholar] [CrossRef]
- Song, Y.; Li, C.; Fu, Y.; Xie, Q.; Guo, J.; Li, G.; Wu, H. Inward tension of Talin and integrin-related osmotic pressure are involved synergetically in the invasion and metastasis of non-small cell lung cancer. J. Cancer 2020, 11, 5032–5041. [Google Scholar] [CrossRef]
- Haslene-Hox, H.; Oveland, E.; Berg, K.C.; Kolmannskog, O.; Woie, K.; Salvesen, H.B.; Tenstad, O.; Wiig, H. A new method for isolation of interstitial fluid from human solid tumors applied to proteomic analysis of ovarian carcinoma tissue. PLoS ONE 2011, 6, e19217. [Google Scholar] [CrossRef] [Green Version]
- Havard, M.; Dautry, F.; Tchénio, T. A dormant state modulated by osmotic pressure controls clonogenicity of prostate cancer cells. J. Biol. Chem. 2011, 286, 44177–44186. [Google Scholar] [CrossRef] [Green Version]
- Semenova, N.; Tuchin, V.V. 3D models of the dynamics of cancer cells under external pressure. Chaos Woodbury N. 2021, 31, 083122. [Google Scholar] [CrossRef]
- Knepper, P.A.; McLone, D.G. Glycosaminoglycans and outflow pathways of the eye and brain. Pediatr. Neurosurg. 1985, 12, 240–251. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grossen, A.; Smith, K.; Coulibaly, N.; Arbuckle, B.; Evans, A.; Wilhelm, S.; Jones, K.; Dunn, I.; Towner, R.; Wu, D.; et al. Physical Forces in Glioblastoma Migration: A Systematic Review. Int. J. Mol. Sci. 2022, 23, 4055. https://doi.org/10.3390/ijms23074055
Grossen A, Smith K, Coulibaly N, Arbuckle B, Evans A, Wilhelm S, Jones K, Dunn I, Towner R, Wu D, et al. Physical Forces in Glioblastoma Migration: A Systematic Review. International Journal of Molecular Sciences. 2022; 23(7):4055. https://doi.org/10.3390/ijms23074055
Chicago/Turabian StyleGrossen, Audrey, Kyle Smith, Nangorgo Coulibaly, Benjamin Arbuckle, Alexander Evans, Stefan Wilhelm, Kenneth Jones, Ian Dunn, Rheal Towner, Dee Wu, and et al. 2022. "Physical Forces in Glioblastoma Migration: A Systematic Review" International Journal of Molecular Sciences 23, no. 7: 4055. https://doi.org/10.3390/ijms23074055