
Sequence Requirements for miR-424-5p Regulating and Function in Cancers


Abstract
:1. Introduction
2. Regulation of miR-424-5p Expression in Cancer
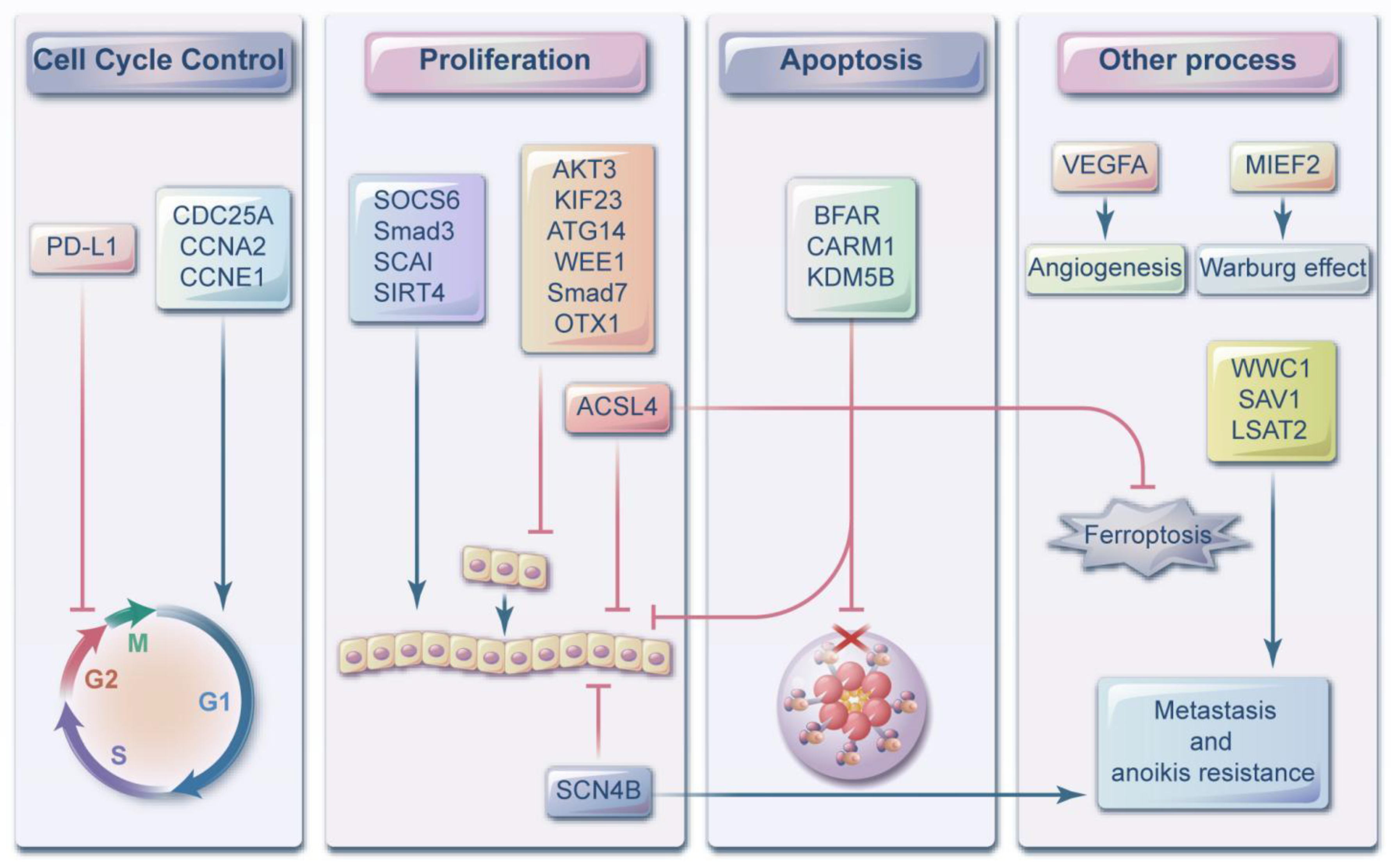
3. MiR-424-5p as a Tumor Suppressor
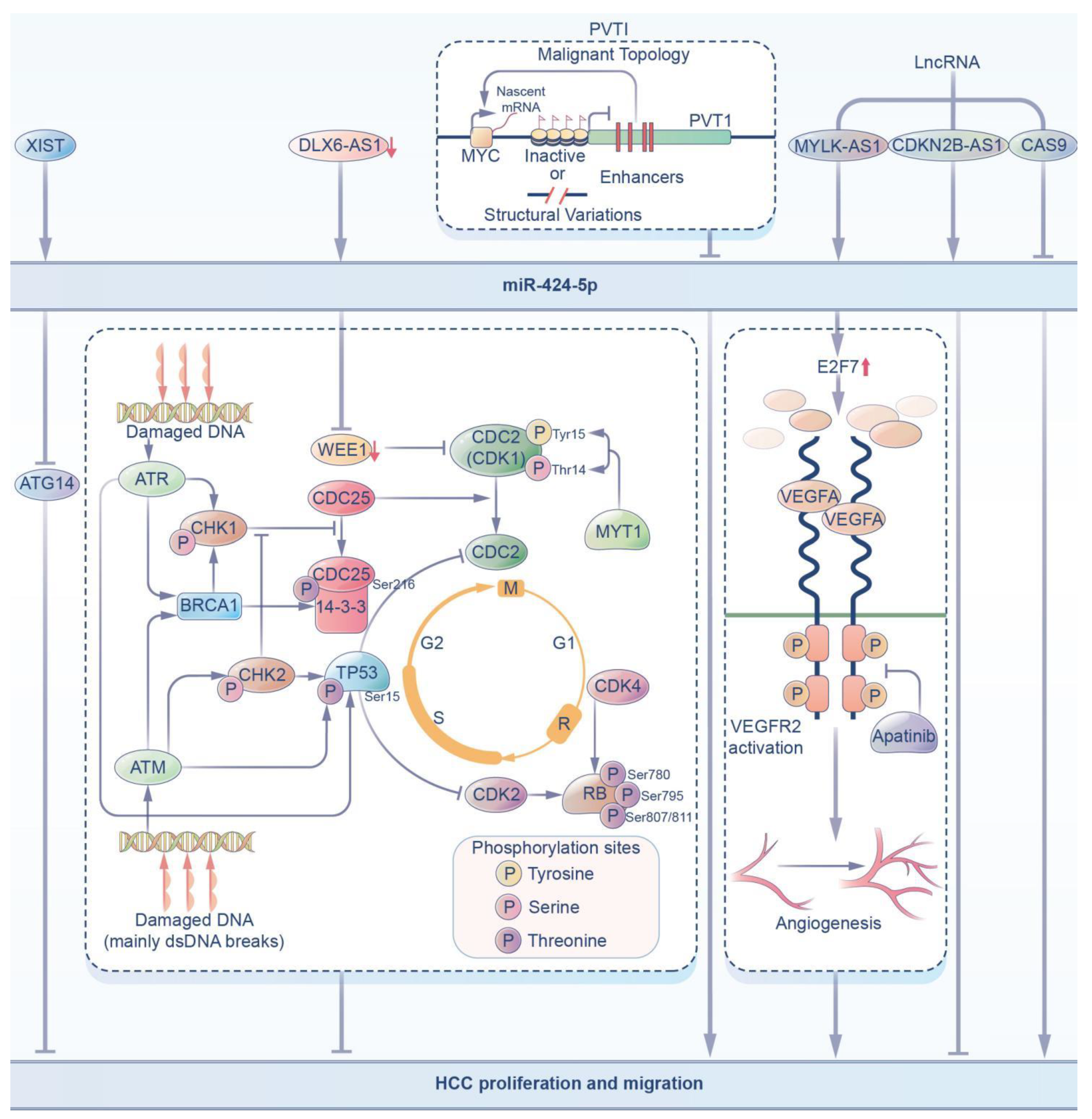
3.1. Hepatocellular Carcinoma
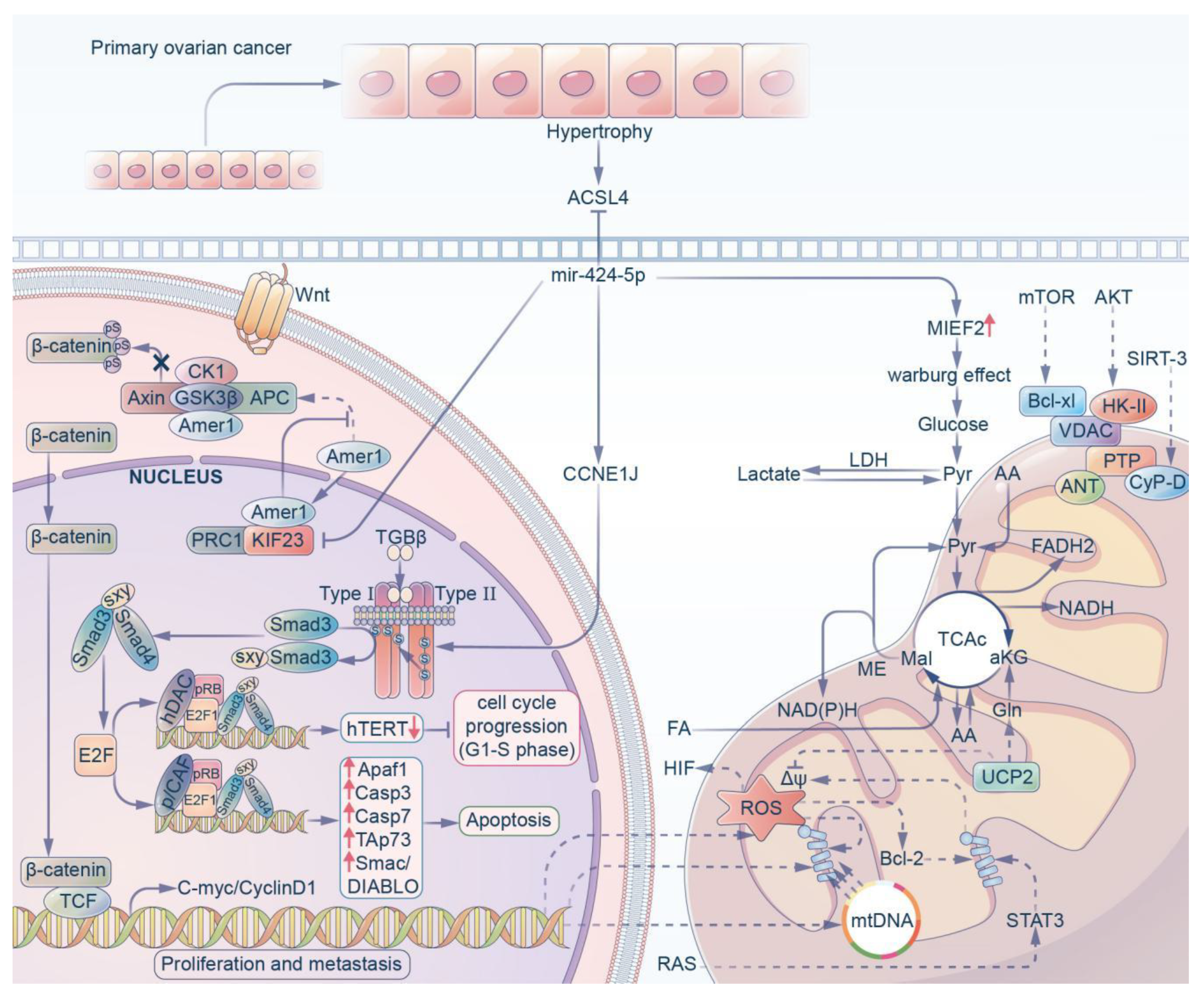
3.2. Ovarian Cancer
3.3. Cervical Cancer
3.4. Neurological Malignancies
3.5. Breast Cancer
3.6. Non-Small-Cell Lung Cancer
3.7. Osteosarcoma
3.8. Nasopharyngeal Carcinoma
4. MiR-424-5p Acts as an Oncogene
4.1. Pancreatic Cancer
4.2. Thyroid Cancer
4.3. Gastric Cancer
5. Controversial or Contradictory Research
5.1. Squamous Cell Carcinoma (SCC)
5.2. Colorectal Cancer
6. MiR-424-5p in the TME
7. MiR-424-5p and Chemotherapy
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lu, T.X.; Rothenberg, M.E. MicroRNA. J. Allergy Clin. Immunol. 2018, 141, 1202–1207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans Heterochronic Gene lin-4 Encodes Small RNAs with Antisense Complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- DeVeale, B.; Swindlehurst-Chan, J.; Blelloch, R. The Roles of microRNAs in Mouse Development. Nat. Rev. Genet. 2021, 22, 307–323. [Google Scholar] [CrossRef] [PubMed]
- Tang, L. Recapitulating miRNA Biogenesis in Cells. Nat. Methods 2022, 19, 35. [Google Scholar] [CrossRef] [PubMed]
- Jet, T.; Gines, G.; Rondelez, Y.; Taly, V. Advances in Multiplexed Techniques for the Detection and Quantification of microRNAs. Chem. Soc. Rev. 2021, 50, 4141–4161. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Target Reconition and Regulatory Functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [Green Version]
- Wojciechowska, A.; Braniewska, A.; Kozar-Kaminska, K. MicroRNA in Cardiovascular Biology and Disease. Adv. Clin. Exp. Med. 2017, 26, 865–874. [Google Scholar] [CrossRef] [Green Version]
- Vasu, S.; Kumano, K.; Darden, C.M.; Rahman, I.; Lawrence, M.C.; Naziruddin, B. MicroRNA Signatures as Future Biomarkers for Diagnosis of Diabetes States. Cells 2019, 8, 1533. [Google Scholar] [CrossRef] [Green Version]
- Kwak, P.B.; Iwasaki, S.; Tomari, Y. The microRNA Pathway and Cancer. Cancer Sci. 2010, 101, 2309–2315. [Google Scholar] [CrossRef]
- Croce, C.M. Causes and Consequences of microRNA Dysregulation in Cancer. Nat. Rev. Genet. 2009, 10, 704–714. [Google Scholar] [CrossRef]
- Liu, B.; Li, J.; Cairns, M.J. Identifying miRNAs, Targets and Functions. Brief. Bioinform. 2014, 15, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Wan, A.; Zhou, Z.; Chen, D.; Liang, H.; Liu, C.; Yan, S.; Niu, Y.; Lin, Z.; Zhan, S.; et al. RNA-Binding Protein RALY Reprogrammes Mitochondrial Metabolism via Mediating miRNA Processing in Colorectal Cancer. Gut 2021, 70, 1698–1712. [Google Scholar] [CrossRef]
- Connolly, M.; Paul, R.; Farre-Garros, R.; Natanek, S.A.; Bloch, S.; Lee, J.; Lorenzo, J.P.; Patel, H.; Cooper, C.; Sayer, A.A.; et al. miR-424-5p Reduces Ribosomal RNA and Protein Synthesis in Muscle Wasting. J. Cachexia Sarcopenia Muscle 2018, 9, 400–416. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Zheng, W.; Shuai, X.; Chang, R.M.; Yu, L.; Fang, F.; Yang, L.Y. MicroRNA-424 Inhibits Akt3/E2F3 Axis and Tumor Growth in Hepatocellular Carcinoma. Oncotarget 2015, 6, 27736–27750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, A.; Papangeli, I.; Park, Y.; Jeong, H.N.; Choi, J.; Kang, H.; Jo, H.N.; Kim, J.; Chun, H.J. A PPARgamma-Dependent miR-424/503-CD40 Axis Regulates Inflammation Mediated Angiogenesis. Sci. Rep. 2017, 7, 2528. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Shi, Z.; Li, M.; Mi, J. Hypoxia-Induced miR-424 Decreases Tumor Sensitivity to Chemotherapy by Inhibiting Apoptosis. Cell Death Dis. 2014, 5, e1301. [Google Scholar] [CrossRef] [Green Version]
- Drasin, D.J.; Guarnieri, A.L.; Neelakantan, D.; Kim, J.; Cabrera, J.H.; Wang, C.A.; Zaberezhnyy, V.; Gasparini, P.; Cascione, L.; Huebner, K.; et al. TWIST1-Induced miR-424 Reversibly Drives Mesenchymal Programming while Inhibiting Tumor Initiation. Cancer Res. 2015, 75, 1908–1921. [Google Scholar] [CrossRef] [Green Version]
- Linsley, P.S.; Schelter, J.; Burchard, J.; Kibukawa, M.; Martin, M.M.; Bartz, S.R.; Johnson, J.M.; Cummins, J.M.; Raymond, C.K.; Dai, H.; et al. Transcripts Targeted by the microRNA-16 Family Cooperatively Regulate Cell Cycle Progression. Mol. Cell. Biol. 2007, 27, 2240–2252. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Fu, H.; Sun, F.; Zhang, H.; Tie, Y.; Zhu, J.; Xing, R.; Sun, Z.; Zheng, X. miR-16 Family Induces Cell Cycle Arrest by Regulating Multiple Cell Cycle Genes. Nucleic Acids Res. 2008, 36, 5391–5404. [Google Scholar] [CrossRef]
- Shekhar, R.; Priyanka, P.; Kumar, P.; Ghosh, T.; Khan, M.M.; Nagarajan, P.; Saxena, S. The microRNAs miR-449a and miR-424 Suppress Osteosarcoma by Targeting Cyclin A2 Expression. J. Biol. Chem. 2019, 294, 4381–4400. [Google Scholar] [CrossRef] [Green Version]
- Matheson, C.J.; Backos, D.S.; Reigan, P. Targeting WEE1 Kinase in Cancer. Trends Pharmacol. Sci. 2016, 37, 872–881. [Google Scholar] [CrossRef] [PubMed]
- Teng, F.; Zhang, J.X.; Chang, Q.M.; Wu, X.B.; Tang, W.G.; Wang, J.F.; Feng, J.F.; Zhang, Z.P.; Hu, Z.Q. LncRNA MYLK-AS1 Facilitates Tumor Progression and Angiogenesis by Targeting miR-424-5p/E2F7 Axis and Activating VEGFR-2 Signaling Pathway in Hepatocellular Carcinoma. J. Exp. Clin. Cancer Res. 2020, 39, 235. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Yang, B.; Zhang, Y.; Feng, X.; He, B.; Xie, H.; Zhou, L.; Wu, J.; Zheng, S. miR-424-5p Represses the Metastasis and Invasion of Intrahepatic Cholangiocarcinoma by Targeting ARK5. Int. J. Biol. Sci. 2019, 15, 1591–1599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, L.L.; Liang, L.; Zhou, D.; Wang, S.W. Tumor Suppressor miR-424-5p Abrogates Ferroptosis in Ovarian Cancer through Targeting ACSL4. Neoplasma 2021, 68, 165–173. [Google Scholar] [CrossRef]
- Li, T.; Li, Y.; Gan, Y.; Tian, R.; Wu, Q.; Shu, G.; Yin, G. Methylation-Mediated Repression of MiR-424/503 Cluster Promotes Proliferation and Migration of Ovarian Cancer Cells through Targeting the Hub Gene KIF23. Cell Cycle 2019, 18, 1601–1618. [Google Scholar] [CrossRef]
- Zhou, Y.; An, Q.; Guo, R.X.; Qiao, Y.H.; Li, L.X.; Zhang, X.Y.; Zhao, X.L. miR424-5p Functions as an Anti-oncogene in Cervical Cancer Cell Growth by Targeting KDM5B via the Notch Signaling Pathway. Life Sci. 2017, 171, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Shu, H.; Cui, Y.; Zhang, Q.; Zhao, B.; Pan, D.; Chao, Q.; Wang, D. MiR-424-5p Inhibits Proliferation, Invasion and Promotes Apoptosis and Predicts Good Prognosis in Glioma by Directly Targeting BFAR. Pathol. Oncol. Res. 2020, 26, 2327–2335. [Google Scholar] [CrossRef]
- Dastmalchi, N.; Hosseinpourfeizi, M.A.; Khojasteh, S.M.B.; Baradaran, B.; Safaralizadeh, R. Tumor Suppressive Activity of miR-424-5p in Breast Cancer Cells Through Targeting PD-L1 and Modulating PTEN/PI3K/AKT/mTOR Signaling Pathway. Life Sci. 2020, 259, 118239. [Google Scholar] [CrossRef]
- Wang, D.; Hu, Y. Long Non-coding RNA PVT1 Competitively Binds MicroRNA-424-5p to Regulate CARM1 in Radiosensitivity of Non-Small-Cell Lung Cancer. Mol. Ther. Nucleic Acids 2019, 16, 130–140. [Google Scholar] [CrossRef] [Green Version]
- Vimalraj, S.; Saravanan, S.; Raghunandhakumar, S.; Anuradha, D. Melatonin Regulates Tumor Angiogenesis via miR-424-5p/VEGFA Signaling Pathway in Osteosarcoma. Life Sci. 2020, 256, 118011. [Google Scholar] [CrossRef]
- Zhao, C.; Zhao, F.; Chen, H.; Liu, Y.; Su, J. MicroRNA-424-5p Inhibits the Proliferation, Migration, and Invasion of Nasopharyngeal Carcinoma Cells by Decreasing AKT3 Expression. Braz. J. Med. Biol. Res. 2020, 53, e9029. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.T.; Lin, W.Y.; Chang, Y.H.; Lin, P.Y.; Chen, W.C.; Chen, M.F. DNMT1-Dependent Suppression of microRNA424 Regulates Tumor Progression in Human Bladder Cancer. Oncotarget 2015, 6, 24119–24131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Fu, Y.; Zhang, G.; Zhang, D.; Liang, N.; Li, F.; Li, C.; Sui, C.; Jiang, J.; Lu, H.; et al. miR-424-5p Promotes Anoikis Resistance and Lung Metastasis by Inactivating Hippo Signaling in Thyroid Cancer. Mol. Ther. Oncolytics 2019, 15, 248–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, S.; Li, Q.; Li, Z.; Wang, L.; Zhang, L.; Xu, Z. miR-424-5p Promotes Proliferation of Gastric Cancer by Targeting Smad3 through TGF-beta Signaling Pathway. Oncotarget 2016, 7, 75185–75196. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Xie, H.Y.; Ding, S.M.; Xing, C.Y.; Chen, A.; Lai, M.C.; Zhou, L.; Zheng, S.S. CADM1 Regulates the G1/S Transition and Represses Tumorigenicity through the Rb-E2F Pathway in Hepatocellular Carcinoma. Hepatobiliary Pancreat. Dis. Int. 2016, 15, 289–296. [Google Scholar] [CrossRef]
- Peng, H.Y.; Jiang, S.S.; Hsiao, J.R.; Hsiao, M.; Hsu, Y.M.; Wu, G.H.; Chang, W.M.; Chang, J.Y.; Jin, S.L.; Shiah, S.G. IL-8 Induces miR-424-5p Expression and Modulates SOCS2/STAT5 Signaling Pathway in Oral Squamous Cell Carcinoma. Mol. Oncol. 2016, 10, 895–909. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.Y.; Yin, F.; Zhang, Y.M.; Liu, C.L.; Bengio, Y. Drawing and Recognizing Chinese Characters with Recurrent Neural Network. IEEE Trans. Pattern Anal. Mach. Intell. 2018, 40, 849–862. [Google Scholar] [CrossRef] [Green Version]
- Lamouille, S.; Subramanyam, D.; Blelloch, R.; Derynck, R. Regulation of Epithelial-Mesenchymal and Mesenchymal-Epithelial Transitions by microRNAs. Curr. Opin. Cell Biol. 2013, 25, 200–207. [Google Scholar] [CrossRef] [Green Version]
- Hashmi, H.; Maqbool, A.; Ahmed, S.; Ahmed, A.; Sheikh, K.; Ahmed, A. Concurrent Cisplatin-Based Chemoradiation in Squamous Cell Carcinoma of Cervix. J. Coll. Physicians Surg. Pak. 2016, 26, 302–305. [Google Scholar]
- Cheng, C.; Li, H.; Zheng, J.; Xu, J.; Gao, P.; Wang, J. FENDRR Sponges miR-424-5p to Inhibit Cell Proliferation, Migration and Invasion in Colorectal Cancer. Technol. Cancer Res. Treat. 2020, 19, 1533033820980102. [Google Scholar] [CrossRef]
- Dai, W.; Zhou, J.; Wang, H.; Zhang, M.; Yang, X.; Song, W. miR-424-5p Promotes the Proliferation and Metastasis of Colorectal Cancer by Directly Targeting SCN4B. Pathol. Res. Pract. 2020, 216, 152731. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Zhu, G.; Bao, S.; Chen, S. Long Non-Coding RNA LINC00511 Mediates the Effects of ESR1 on Proliferation and Invasion of Ovarian Cancer Through miR-424-5p and miR-370-5p. Cancer Manag. Res. 2019, 11, 10807–10819. [Google Scholar] [CrossRef] [PubMed]
- Jalali, S.; Bhartiya, D.; Lalwani, M.K.; Sivasubbu, S.; Scaria, V. Systematic Transcriptome Wide Analysis of lncRNA-miRNA Interactions. PLoS ONE 2013, 8, e53823. [Google Scholar] [CrossRef] [Green Version]
- Hartke, J.; Johnson, M.; Ghabril, M. The Diagnosis and Treatment of Hepatocellular Carcinoma. Semin. Diagn. Pathol. 2017, 34, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Piao, L.; Wang, F.; Wang, Y.; Yang, Z.; Li, Q.; Cui, L.; Yu, Q. miR-424-5p Regulates Hepatoma Cell Proliferation and Apoptosis. Cancer Biother. Radiopharm. 2019, 34, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Xu, Q.; Xiao, S.; Wu, Z.; Gong, J.; Liu, C.; Ren, G.; Wu, H. MicroRNA-424-5p Acts as a Potential Biomarker and Inhibits Proliferation and Invasion in Hepatocellular Carcinoma by Targeting TRIM29. Life Sci. 2019, 224, 1–11. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhu, C.; Chang, Q.; Peng, P.; Yang, J.; Liu, C.; Liu, Y.; Chen, X.; Liu, Y.; Cheng, R.; et al. MiR-424-5p Regulates Cell Cycle and Inhibits Proliferation of Hepatocellular Carcinoma Cells by Targeting E2F7. PLoS ONE 2020, 15, e0242179. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, T.; Guo, P.; Kang, J.; Wei, Q.; Jia, X.; Zhao, W.; Huai, W.; Qiu, Y.; Sun, L.; et al. MiR-424-5p Reversed Epithelial-Mesenchymal Transition of Anchorage-Independent HCC Cells by Directly Targeting ICAT and Suppressed HCC Progression. Sci. Rep. 2014, 4, 6248. [Google Scholar] [CrossRef]
- Zhu, X.J.; Jin, X.; Dai, L.; Liu, X.; Liu, Z. ATG4 Promotes Cell Proliferation, Migration and Invasion in HCC and Predicts a Poor Prognosis. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 12686–12693. [Google Scholar] [CrossRef]
- Li, D.; Tang, X.; Li, M.; Zheng, Y. Long Noncoding RNA DLX6-AS1 Promotes Liver Cancer by Increasing the Expression of WEE1 via Targeting miR-424-5p. J. Cell Biochem. 2019, 120, 12290–12299. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Wen, D.Y.; Zhang, R.; Huang, J.C.; Lin, P.; Ren, F.H.; Wang, X.; He, Y.; Yang, H.; Chen, G.; et al. A Preliminary Investigation of PVT1 on the Effect and Mechanisms of Hepatocellular Carcinoma: Evidence from Clinical Data, a Meta-Analysis of 840 Cases, and In Vivo Validation. Cell Physiol. Biochem. 2018, 47, 2216–2232. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Li, Y.; He, F.; Kong, J. LncRNA CDKN2B-AS1 Promotes Cell Viability, Migration, and Invasion of Hepatocellular Carcinoma via Sponging miR-424-5p. Cancer Manag. Res. 2020, 12, 6807–6819. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Fu, J.; Liu, Y.; Qu, W.; Wang, G.; Yan, Z. LncRNA CASC9 Promotes Proliferation, Migration and Inhibits Apoptosis of Hepatocellular Carcinoma Cells by Down-Regulating miR-424-5p. Ann. Hepatol. 2020, 23, 100297. [Google Scholar] [CrossRef] [PubMed]
- El-Diwany, R.; Pawlik, T.M.; Ejaz, A. Intrahepatic Cholangiocarcinoma. Surg. Oncol. Clin. N. Am. 2019, 28, 587–599. [Google Scholar] [CrossRef]
- Torre, L.A.; Islami, F.; Siegel, R.L.; Ward, E.M.; Jemal, A. Global Cancer in Women: Burden and Trends. Cancer Epidemiol. Biomarkers Prev. 2017, 26, 444–457. [Google Scholar] [CrossRef] [Green Version]
- Symeonides, S.; Gourley, C. Ovarian Cancer Molecular Stratification and Tumor Heterogeneity: A Necessity and a Challenge. Front. Oncol. 2015, 5, 229. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Gu, Z.; Tang, Y.; Hao, J.; Zhang, C.; Yang, X. Tumour-Suppressive microRNA-424-5p directly targets CCNE1 as Potential Prognostic Markers in Epithelial Ovarian Cancer. Cell Cycle 2018, 17, 309–318. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Zhang, X.; Shi, Y.; Cheng, L.; Song, T.; Wu, B.; Li, J.; Yang, H. MIEF2 Over-Expression Promotes Tumor Growth and Metastasis through Reprogramming of Glucose Metabolism in Ovarian Cancer. J. Exp. Clin. Cancer Res. 2020, 39, 286. [Google Scholar] [CrossRef]
- Schwartz, L.; Supuran, C.T.; Alfarouk, K.O. The Warburg Effect and the Hallmarks of Cancer. Anticancer Agents Med. Chem 2017, 17, 164–170. [Google Scholar] [CrossRef]
- Cha, S.Y.; Choi, Y.H.; Hwang, S.; Jeong, J.Y.; An, H.J. Clinical Impact of microRNAs Associated with Cancer Stem Cells as a Prognostic Factor in Ovarian Carcinoma. J. Cancer 2017, 8, 3538–3547. [Google Scholar] [CrossRef] [Green Version]
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global Cancer Statistics. CA Cancer J. Clin. 2011, 61, 69–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Li, Q.; Jin, H.; Zou, H.; Xia, W.; Dai, N.; Dai, X.Y.; Wang, D.; Xu, C.X.; Qing, Y. miR-424 Acts as a Tumor Radiosensitizer by Targeting Aprataxin in Cervical Cancer. Oncotarget 2016, 7, 77508–77515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Li, Y.; Wang, F.; Wang, X.; Cheng, B.; Ye, F.; Xie, X.; Zhou, C.; Lu, W. Suppressed miR-424 Expression via Upregulation of Target Gene Chk1 Contributes to the Progression of Cervical Cancer. Oncogene 2013, 32, 976–987. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Wang, Q.; Li, L.; Xiao-Jin, Z. Upregulation of Long Non-Coding RNA Small Nucleolar RNA Host Gene 12 Contributes to Cell Growth and Invasion in Cervical Cancer by Acting as a Sponge for MiR-424-5p. Cell Physiol. Biochem. 2018, 45, 2086–2094. [Google Scholar] [CrossRef]
- Jansen, E.; Hamisch, C.; Ruess, D.; Heiland, D.H.; Goldbrunner, R.; Ruge, M.I.; Schnell, O.; Grau, S.J. Observation after Surgery for Low Grade Glioma: Long-Term Outcome in the Light of the 2016 WHO Classification. J. Neurooncol. 2019, 145, 501–507. [Google Scholar] [CrossRef]
- De Mariano, M.; Stigliani, S.; Moretti, S.; Parodi, F.; Croce, M.; Bernardi, C.; Pagano, A.; Tonini, G.P.; Ferrini, S.; Longo, L. A Genome-Wide microRNA Profiling Indicates miR-424-5p and miR-503-5p as Regulators of ALK Expression in Neuroblastoma. Oncotarget 2017, 8, 56518–56532. [Google Scholar] [CrossRef] [Green Version]
- Zhou, K.; Li, S.; Du, G.; Fan, Y.; Wu, P.; Sun, H.; Zhang, T. LncRNA XIST Depletion Prevents Cancer Progression in Invasive Pituitary Neuroendocrine Tumor by Inhibiting bFGF via Upregulation of microRNA-424-5p. Onco. Targets Ther. 2019, 12, 7095–7109. [Google Scholar] [CrossRef] [Green Version]
- Xie, D.; Song, H.; Wu, T.; Li, D.; Hua, K.; Xu, H.; Zhao, B.; Wu, C.; Hu, J.; Ji, C.; et al. MicroRNA-424 Serves an Anti-Oncogenic Role by Targeting Cyclin-Dependent Kinase 1 in Breast Cancer Cells. Oncol. Rep. 2018, 40, 3416–3426. [Google Scholar] [CrossRef]
- Yue, X.; Wang, Z. Long Intergenic Non-Coding RNA LINC00922 Aggravates the Malignant Phenotype of Breast Cancer by Regulating the microRNA-424-5p/BDNF Axis. Cancer Manag. Res. 2020, 12, 7539–7552. [Google Scholar] [CrossRef]
- Dastmalchi, N.; Baradaran, B.; Banan Khojasteh, S.M.; Hosseinpourfeizi, M.; Safaralizadeh, R. miR-424: A Novel Potential Therapeutic Target and Prognostic Factor in Malignancies. Cell Biol. Int. 2021, 45, 720–730. [Google Scholar] [CrossRef]
- Wang, J.; Wang, S.; Zhou, J.; Qian, Q. miR-424-5p Regulates Cell Proliferation, Migration and Invasion by Targeting Doublecortin-like Kinase 1 in Basal-like Breast Cancer. Biomed. Pharmacother. 2018, 102, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Paraskevopoulou, M.D.; Hatzigeorgiou, A.G. Analyzing MiRNA-LncRNA Interactions. Methods Mol. Biol. 2016, 1402, 271–286. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhao, L.; Zhao, P.; Liu, Z. Long Non-Coding RNA LINC00641 Suppresses Non-Small-Cell Lung Cancer by Sponging miR-424-5p to Upregulate PLSCR4. Cancer Biomark. 2019, 26, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.J.; Tay, Y. Noncoding RNA:RNA Regulatory Networks in Cancer. Int. J. Mol. Sci. 2018, 19, 1310. [Google Scholar] [CrossRef] [Green Version]
- Kansara, M.; Teng, M.W.; Smyth, M.J.; Thomas, D.M. Translational Biology of Osteosarcoma. Nat. Rev. Cancer 2014, 14, 722–735. [Google Scholar] [CrossRef]
- Chow, J.C.; Ngan, R.K.; Cheung, K.M.; Cho, W.C. Immunotherapeutic Approaches in Nasopharyngeal Carcinoma. Expert Opin. Biol. Ther. 2019, 19, 1165–1172. [Google Scholar] [CrossRef]
- Ansari, D.; Tingstedt, B.; Andersson, B.; Holmquist, F.; Sturesson, C.; Williamsson, C.; Sasor, A.; Borg, D.; Bauden, M.; Andersson, R. Pancreatic Cancer: Yesterday, Today and Tomorrow. Future Oncol. 2016, 12, 1929–1946. [Google Scholar] [CrossRef] [Green Version]
- Nan, B.Y.; Xiong, G.F.; Zhao, Z.R.; Gu, X.; Huang, X.S. Comprehensive Identification of Potential Crucial Genes and miRNA-mRNA Regulatory Networks in Papillary Thyroid Cancer. Biomed. Res. Int. 2021, 2021, 6752141. [Google Scholar] [CrossRef]
- Shah, M.Y.; Ferrajoli, A.; Sood, A.K.; Lopez-Berestein, G.; Calin, G.A. microRNA Therapeutics in Cancer—An Emerging Concept. EBioMedicine 2016, 12, 34–42. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Liu, H.; Hou, L.; Wang, G.; Zhang, R.; Huang, Y.; Chen, X.; Zhu, J. Circular RNA_LARP4 Inhibits Cell Proliferation and Invasion of Gastric Cancer by Sponging miR-424-5p and Regulating LATS1 Expression. Mol. Cancer 2017, 16, 151. [Google Scholar] [CrossRef]
- Sanchez-Danes, A.; Blanpain, C. Deciphering the Cells of Origin of Squamous Cell Carcinomas. Nat. Rev. Cancer 2018, 18, 549–561. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Wistuba, I.I.; Emmert-Buck, M.R.; Erickson, H.S. Squamous Cell Carcinoma—Similarities and Differences among Anatomical Sites. Am. J. Cancer Res. 2011, 1, 275–300. [Google Scholar] [CrossRef]
- Agada, F.O.; Patmore, H.; Alhamarneh, O.; Stafford, N.D.; Greenman, J. Genetic Profile of Head and Neck Squamous Cell Carcinoma: Clinical Implications. J. Laryngol. Otol. 2009, 123, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Gao, W.; Wen, S.; Wu, Y.; Fu, R.; Zhao, D.; Chen, X.; Wang, B. Potential Key Molecular Correlations in Laryngeal Squamous Cell Carcinoma Revealed by Integrated Analysis of mRNA, miRNA and lncRNA Microarray Profiles. Neoplasma 2016, 63, 888–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Liu, J.; Hu, W.; Zhang, Y.; Sang, J.; Li, H.; Ma, T.; Bo, Y.; Bai, T.; Guo, H.; et al. miR-424-5p Promotes Proliferation, Migration and Invasion of Laryngeal Squamous Cell Carcinoma. Onco. Targets Ther. 2019, 12, 10441–10453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farabegoli, F.; Ceccarelli, C.; Santini, D.; Taffurelli, M. Suppressor of Cytokine Signalling 2 (SOCS-2) Expression in Breast Carcinoma. J. Clin. Pathol. 2005, 58, 1046–1050. [Google Scholar] [CrossRef]
- Zhang, Y.; Tang, K.; Chen, L.; Du, M.; Qu, Z. Exosomal CircGDI2 Suppresses Oral Squamous Cell Carcinoma Progression Through the Regulation of MiR-424-5p/SCAI Axis. Cancer Manag. Res. 2020, 12, 7501–7514. [Google Scholar] [CrossRef]
- Enzinger, P.C.; Mayer, R.J. Esophageal Cancer. N. Engl. J. Med. 2003, 349, 2241–2252. [Google Scholar] [CrossRef] [Green Version]
- Harada, K.; Baba, Y.; Ishimoto, T.; Shigaki, H.; Kosumi, K.; Yoshida, N.; Watanabe, M.; Baba, H. The Role of microRNA in Esophageal Squamous Cell Carcinoma. J. Gastroenterol. 2016, 51, 520–530. [Google Scholar] [CrossRef]
- Dai, F.; Mei, L.; Meng, S.; Ma, Z.; Guo, W.; Zhou, J.; Zhang, J. The Global Expression Profiling in Esophageal Squamous Cell Carcinoma. Genomics 2017, 109, 241–250. [Google Scholar] [CrossRef]
- Cui, Y.; Yang, J.; Bai, Y.; Zhang, Y.; Yao, Y.; Zheng, T.; Liu, C.; Wu, F. miR-424-5p Regulates Cell Proliferation and Migration of Esophageal Squamous Cell Carcinoma by Target-Ing SIRT4. J. Cancer 2020, 11, 6337–6347. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wang, J.; Yang, X.; Chen, D.; Wang, L. MiR-424-5p Participates in Esophageal Squamous Cell Carcinoma Invasion and Metastasis via SMAD7 Pathway Mediated EMT. Diagn. Pathol. 2016, 11, 88. [Google Scholar] [CrossRef] [Green Version]
- Rothenberg, S.M.; Ellisen, L.W. The Molecular Pathogenesis of Head and Neck Squamous Cell Carcinoma. J. Clin. Invest. 2012, 122, 1951–1957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahami-Fard, M.H.; Kheirandish, S.; Sheikhha, M.H. Expression Levels of miR-143-3p and -424-5p in Colorectal Cancer and Their Clinical Significance. Cancer Biomark. 2019, 24, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Kandhavelu, J.; Subramanian, K.; Khan, A.; Omar, A.; Ruff, P.; Penny, C. Computational Analysis of miRNA and their Gene Targets Significantly Involved in Colorectal Cancer Progression. Microrna 2019, 8, 68–75. [Google Scholar] [CrossRef]
- Yan, W.; Jiang, X.; Wang, G.; Li, W.; Zhai, C.; Chen, S.; Shang, F.; Zhao, Z.; Yu, W. Cyto-biological Effects of microRNA-424-5p on Human Colorectal Cancer Cells. Oncol. Lett. 2020, 20, 120. [Google Scholar] [CrossRef]
- Zou, W. Immune Regulation in the Tumor Microenvironment and Its Relevance in Cancer Therapy. Cell. Mol. Immunol. 2022, 19, 1–2. [Google Scholar] [CrossRef]
- Yi, M.; Niu, M.; Xu, L.; Luo, S.; Wu, K. Regulation of PD-L1 Expression in the Tumor Microenvironment. J. Hematol. Oncol. 2021, 14, 10. [Google Scholar] [CrossRef]
- Genova, C.; Dellepiane, C.; Carrega, P.; Sommariva, S.; Ferlazzo, G.; Pronzato, P.; Gangemi, R.; Filaci, G.; Coco, S.; Croce, M. Therapeutic Implications of Tumor Microenvironment in Lung Cancer: Focus on Immune Checkpoint Blockade. Front. Immunol. 2021, 12, 799455. [Google Scholar] [CrossRef]
- Zhou, Y.; Yamamoto, Y.; Takeshita, F.; Yamamoto, T.; Xiao, Z.; Ochiya, T. Delivery of miR-424-5p via Extracellular Vesicles Promotes the Apoptosis of MDA-MB-231 TNBC Cells in the Tumor Microenvironment. Int. J. Mol. Sci. 2021, 22, 844. [Google Scholar] [CrossRef]
- Christopher, A.F.; Kaur, R.P.; Kaur, G.; Kaur, A.; Gupta, V.; Bansal, P. MicroRNA Therapeutics: Discovering Novel Targets and Developing Specific Therapy. Perspect. Clin. Res. 2016, 7, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Wu, M.; Lu, Y.; Zhao, Z.; Liu, T.; Fu, W.; Li, W. MicroRNA-424 Regulates Cisplatin Resistance of Gastric Cancer by Targeting SMURF1 Based on GEO database and Primary Validation in Human Gastric Cancer Tissues. OncoTargets Ther. 2019, 12, 7623–7636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dastmalchi, N.; Safaralizadeh, R.; Hosseinpourfeizi, M.A.; Baradaran, B.; Khojasteh, S.M.B. MicroRNA-424-5p Enhances Chemosensitivity of Breast Cancer Cells to Taxol and Regulates Cell Cycle, Apoptosis, and Proliferation. Mol. Biol. Rep. 2021, 48, 1345–1357. [Google Scholar] [CrossRef]
- Li, H.; An, X.; Li, Q.; Yu, H.; Li, Z. Construction and Analysis of Competing Endogenous RNA Network of MCF-7 Breast Cancer Cells Based on the Inhibitory Effect of 6-Thioguanine on Cell Proliferation. Oncol. Lett. 2021, 21, 104. [Google Scholar] [CrossRef] [PubMed]
- Nakhjavani, M.; Hardingham, J.E.; Palethorpe, H.M.; Tomita, Y.; Smith, E.; Price, T.J.; Townsend, A.R. Ginsenoside Rg3: Potential Molecular Targets and Therapeutic Indication in Metastatic Breast Cancer. Medicines 2019, 6, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.; Ji, H.W.; Kim, H.W.; Yun, S.H.; Park, J.E.; Kim, S.J. Ginsenoside Rg3 Prevents Oncogenic Long Noncoding RNA ATXN8OS from Inhibiting Tumor-Suppressive microRNA-424-5p in Breast Cancer Cells. Biomolecules 2021, 11, 118. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Type of Cancer | Effect of mir-424-5p | Mechanisms | Reference |
|---|---|---|---|
| Hepatocellular carcinoma (HCC) | Tumor suppressor | Inhibit ATG14, WEE1, activate E2F7 | [21,22] |
| Intrahepatic cholangiocarcinoma (ICC) | Tumor suppressor | Regulate ARK5, inhibit mTOR phosphorylation | [23] |
| Ovarian cancer | Tumor suppressor | Inhibit ACSL4, KIF23, activate CCNE1J, MIEF2 | [24,25] |
| Cervical cancer | Tumor suppressor | Activate KDM5B-Notch | [26] |
| Malignant tumors of the nervous system | Tumor suppressor | Target BFAR and ALK receptor directly, increase βFGF | [27] |
| Breast cancer | Tumor suppressor | Target PD-L1, regulate PTEN/PI3K/AKT/mTOR | [28] |
| Non-small cell carcinoma (NSCLC) | Tumor suppressor | Inhibit CARM1 | [29] |
| Osteosarcoma | Tumor suppressor | Target VEGFA, inhibit angiogenesis | [30] |
| Nasopharyngeal carcinoma | Tumor suppressor | Inhibit AKT3 | [31] |
| Pancreatic cancer | Oncogene | Inhibit SOC6 to regulate ERK1/2 | [32] |
| Thyroid cancer | Oncogene | Target WWC1, SAV1, LSAT2 to inhibit Hippo | [33] |
| Gastric (stomach cancer) | Oncogene | Target Smad3 to regulate TGF-β | [34] |
| Laryngeal squamous-cell Carcinoma | Tumor suppressor | Inhibit CADM1 | [35] |
| Oral squamous-cell Carcinoma | Tumor suppressor | Activate IL-8 to inhibit SOCS2; activate SCAI | [36] |
| Esophageal squamous-cell Carcinoma | Evidence for both | Oncogene: target SIRT4;Tumor suppressor: inhibit Smad7 | [37,38] |
| Cervical squamous-cell carcinoma | oncogene | Target CCND1, CCNE1, WEE1 to regulate cell cycle | [39] |
| Colorectal cancer | Evidence for both | Oncogene: target SCN4B;Tumor suppressor: inhibit OTX1 | [40,41] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xuan, J.; Liu, Y.; Zeng, X.; Wang, H. Sequence Requirements for miR-424-5p Regulating and Function in Cancers. Int. J. Mol. Sci. 2022, 23, 4037. https://doi.org/10.3390/ijms23074037
Xuan J, Liu Y, Zeng X, Wang H. Sequence Requirements for miR-424-5p Regulating and Function in Cancers. International Journal of Molecular Sciences. 2022; 23(7):4037. https://doi.org/10.3390/ijms23074037
Chicago/Turabian StyleXuan, Jiangying, Yingxia Liu, Xiaoping Zeng, and Hongmei Wang. 2022. "Sequence Requirements for miR-424-5p Regulating and Function in Cancers" International Journal of Molecular Sciences 23, no. 7: 4037. https://doi.org/10.3390/ijms23074037