
miRNA Expression Profiling in Subcutaneous Adipose Tissue of Monozygotic Twins Discordant for HIV Infection: Validation of Differentially Expressed miRNA and Bioinformatic Analysis

 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Results
2.1. Clinical Data
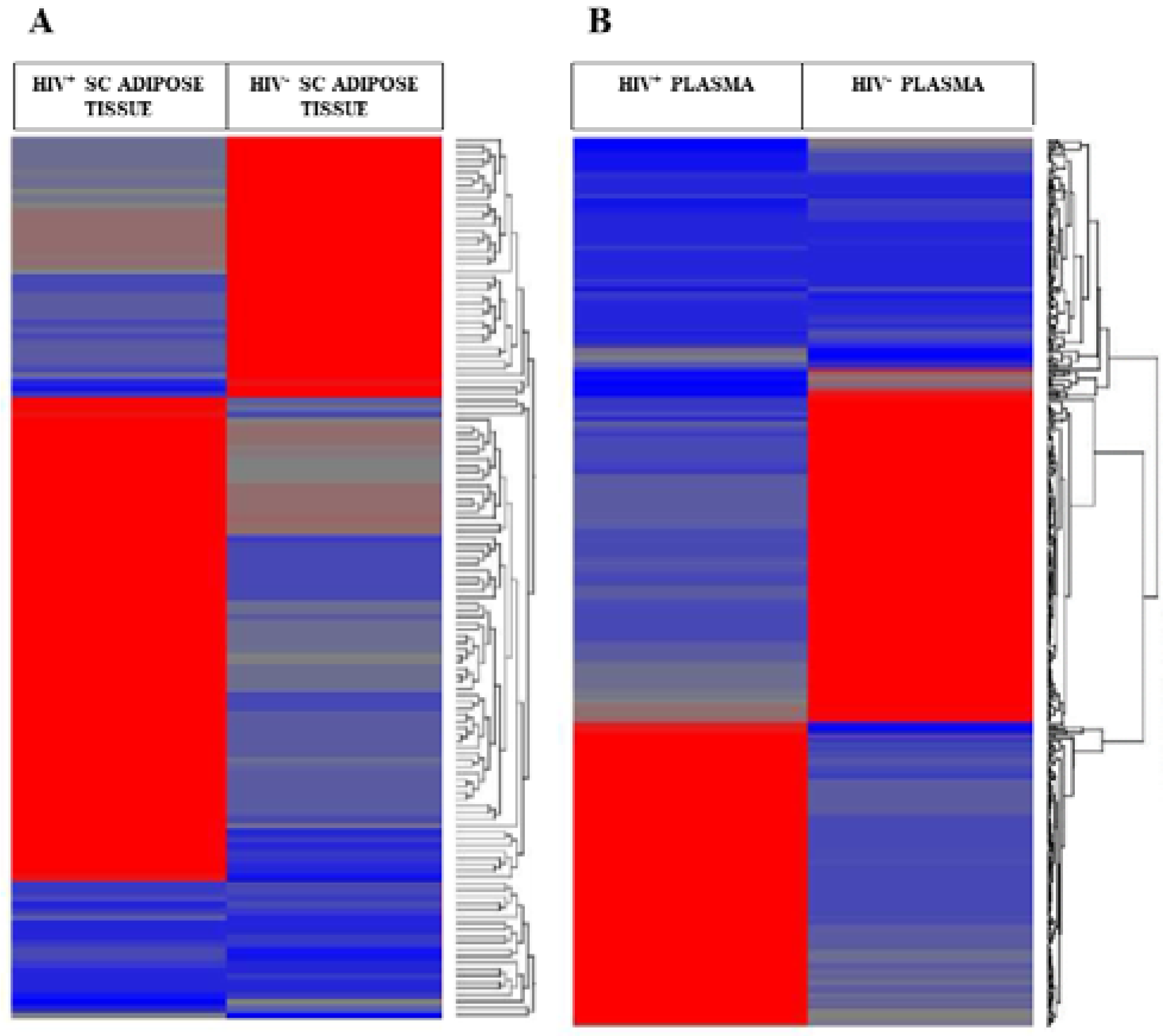
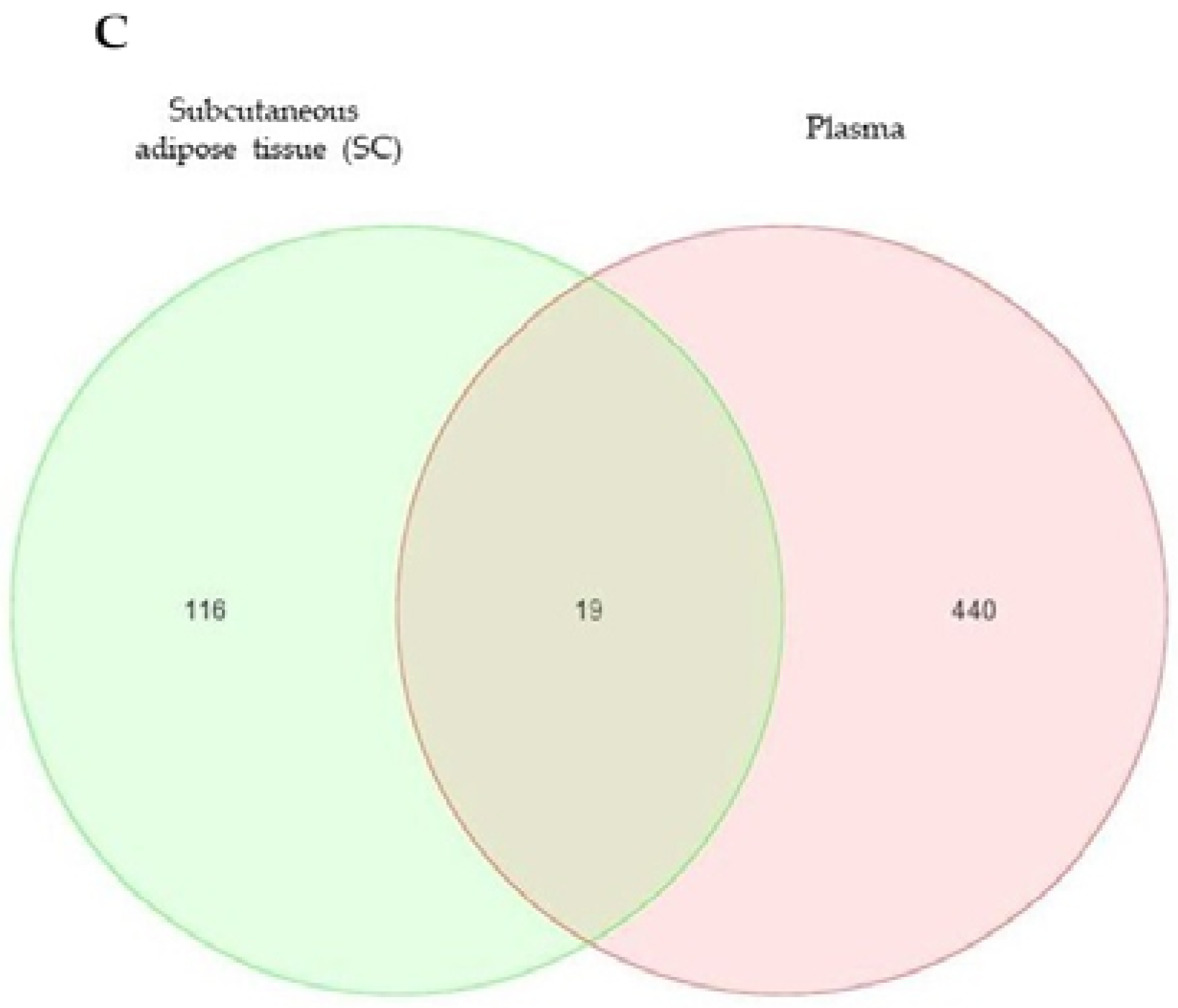
2.2. Differentially Expressed microRNA (DEGs) in the SC Adipose Tissue and the Plasma of the Twins
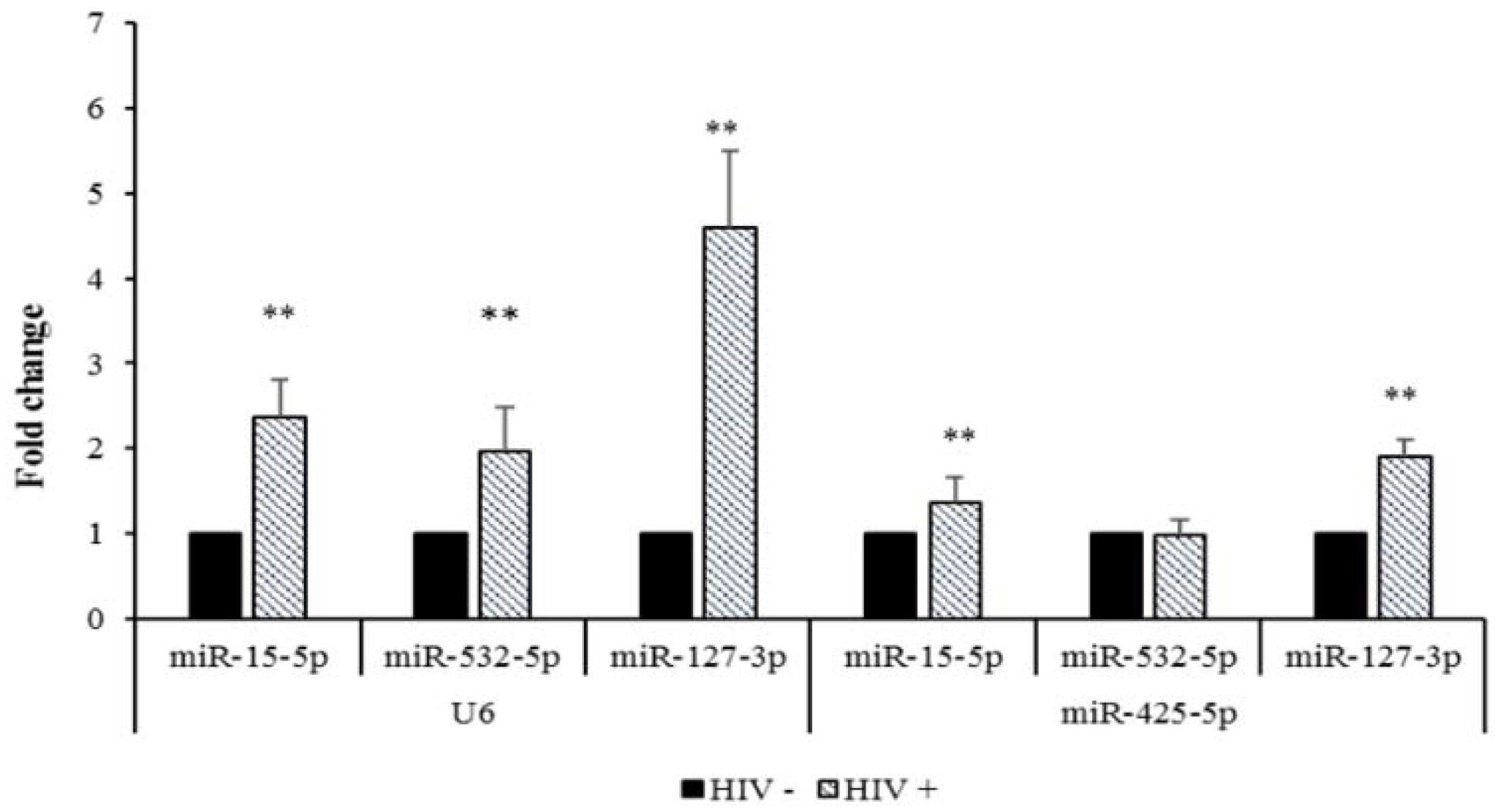
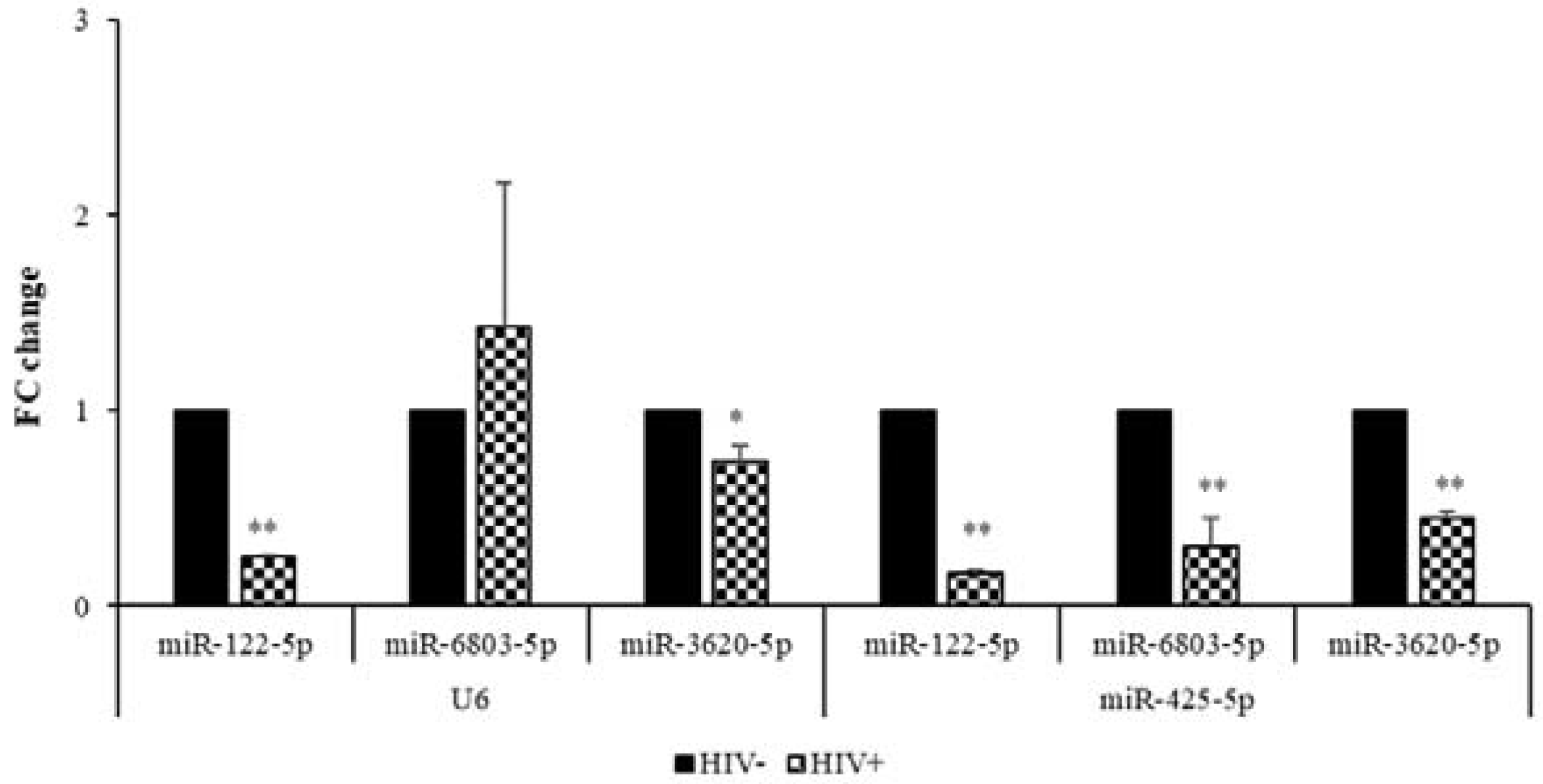
2.3. Validation of microRNA DEGs in SC Adipose Tissue by Real-Time qPCR
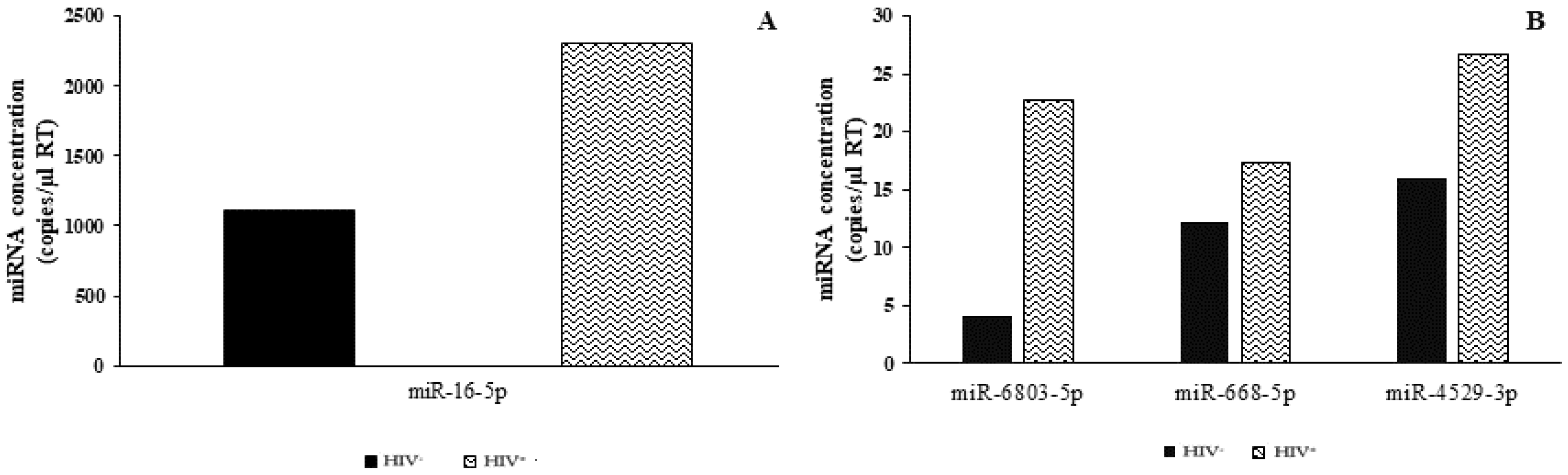
2.4. Analysis of microRNA DEGs in Plasma by Real-Time qPCR
2.5. Analysis of microRNA DEGs in Plasma by ddPCR
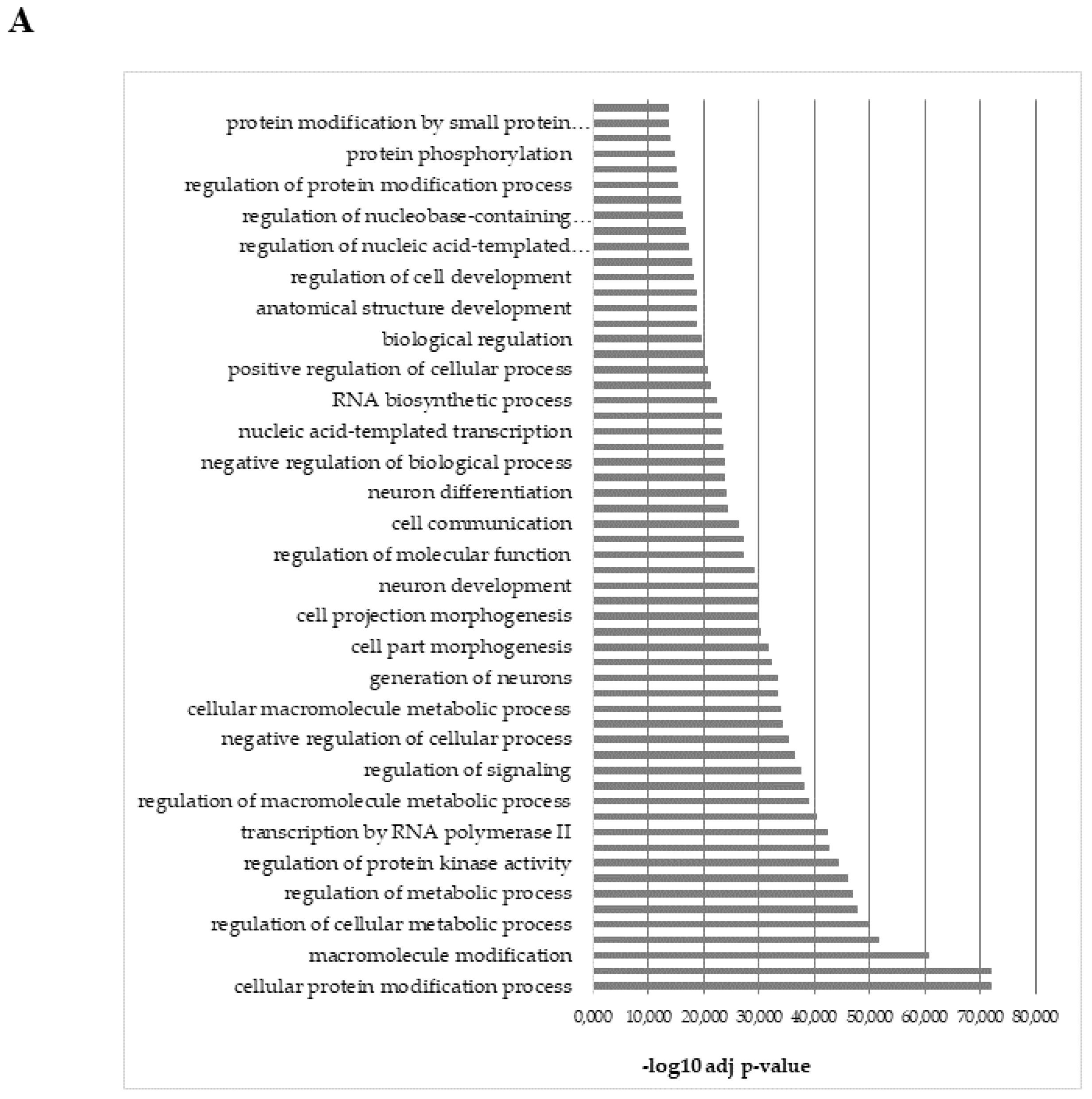
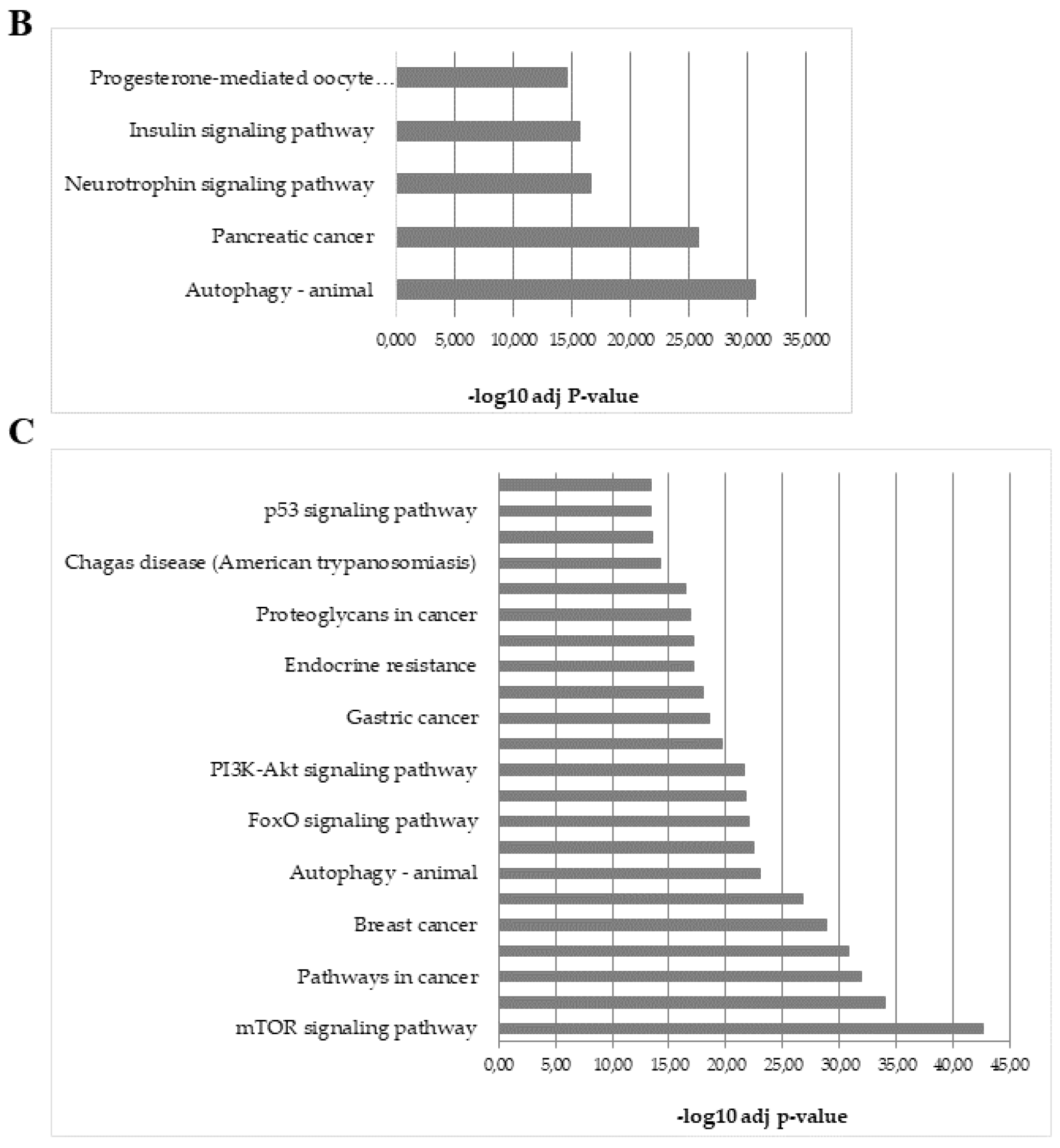
2.6. Analysis of Predicted Target Genes and Functional Annotations of DEGs microRNAs
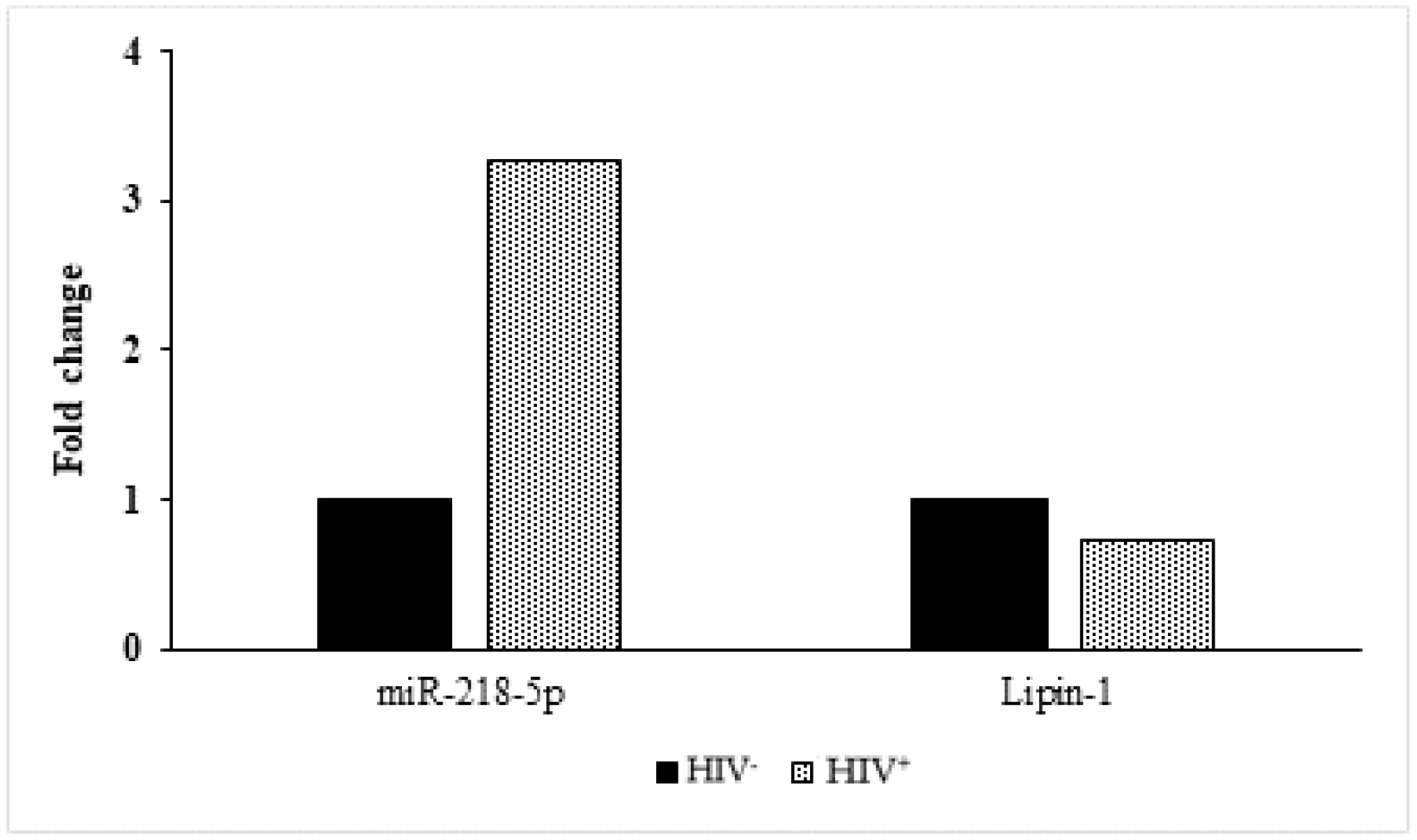
2.7. Analysis of Lipin-1 and miRNA-218-5p Expression in SC Adipose Tissue
3. Discussion
4. Materials and Methods
4.1. Participants
4.2. microRNA Extraction from SC Adipose Tissue and Plasma Samples
4.3. microRNA Microarray
4.4. microRNA Data Analysis
4.5. microRNA Target Prediction and Functional Analysis
4.6. Validation of microRNA Microarray Expression Data by Real-Time qPCR and Droplet Digital PCR (ddPCR)
4.7. Analysis of lipin-1 and miRNA-218-5p Expression in SC Adipose Tissue Samples by Real-Time qPCR
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Koeth, J.R. Adipose Tissue in HIV Infection. Compr. Physiol. 2018, 7, 1339–1357. [Google Scholar]
- Villarroya, F.; Domingo, P.; Giralt, M. Drug-induced lipotoxicity: Lipodystrophy associated with HIV-1 infection and antiretroviral treatment. Biochim. Biophys. Acta 2010, 1801, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Paruthi, J.; Gill, N.; Mantzoros, C.S. Adipokines in the HIV/HAART-associated lipodystrophy syndrome. Metab. Clin. Exp. 2013, 62, 1199–1205. [Google Scholar] [CrossRef]
- Verhaegen, A.A.; Van Gaal, L.F. Drugs That Affect Body Weight, Body Fat Distribution, and Metabolism 2019. In Endotext; Feingold, K.R., Anawalt, B., Boyce, A., Chrousos, G., de Herder, W.W., Dhatariya, K., Dungan, K., Hershman, J.M., Hofland, J., Kalra, S., et al., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2000. [Google Scholar]
- Egaña-Gorroño, L.; Martínez, E.; Pérez, I.; Escribà, T.; Domingo, P.; Gatel, J.M.; Arnedo, M. Contribution of genetic background and antiretroviral therapy to body fat changes in antiretroviral-naive HIV-infected adults. J. Antimicrob. Chemother. 2014, 69, 3076–3084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarr, P.E.; Telenti, A. Genetic screening for metabolic and age-related complications in HIV-infected persons. F1000 Med. Rep. 2010, 2, 83. [Google Scholar] [CrossRef] [Green Version]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [Green Version]
- Thomou, T.; Mori, M.A.; Dreyfuss, J.M.; Konishi, M.; Sakaguchi, M.; Wolfrum, C.; Rao, T.N.; Winnay, J.N.; Garcia-Martin, R.; Grinspoon, S.K.; et al. Adipose-derived circulating miRNAs regulate gene expression in other tissues. Nature 2017, 542, 450–455. [Google Scholar] [CrossRef]
- Wang, J.; Chen, J.; Sen, S. MicroRNA as biomarkers and diagnostics. J. Cell Physiol. 2016, 231, 25–30. [Google Scholar] [CrossRef]
- Jones, A.; Danielson, K.M.; Benton, M.C.; Ziegler, O.; Shah, R.; Stubbs, R.S.; Das, S.; Macartney-Coxson, D. miRNA signatures of insulin resistance in obesity. Obesity 2017, 25, 1734–1744. [Google Scholar] [CrossRef] [Green Version]
- Willeit, P.; Skroblin, P.; Moschen, A.R.; Yin, X.; Kaudewitz, D.; Zampetaki, A.; Barwari, T.; Whitehead, M.; Ramírez, C.M.; Goedeke, L.; et al. Circulating microRNA-122 is associated with the risk of new-onset metabolic syndrome and type 2 diabetes. Diabetes 2017, 66, 347–357. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-Lucena, R.; Camargo, A.; Alcalá-Diaz, J.F.; Romero-Baldonado, C.; Luque, R.M.; van Ommen, B.; Delgado-Lista, J.; Ordovás, J.M.; Pérez-Martínez, P.; Rangel-Zúñiga, O.A.; et al. A plasma circulating miRNAs profile predicts type 2 diabetes mellitus and prediabetes: From the CORDIOPREV study. Exp. Mol. Med. 2018, 50, 168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, M.A.; Thomou, T.; Boucher, J.; Lee, K.Y.; Lallukka, S.; Kim, J.K.; Torriani, M.; Yki-Järvinen, H.; Grinspoon, S.K.; Cypess, A.M.; et al. Altered miRNA processing disrupts brown/white adipocyte determination and associates with lipodystrophy. J. Clin. Investig. 2014, 124, 3339–3351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srinivasa, S.; Garcia-Martin, R.; Torriani, M.; Fitch, K.V.; Carlson, A.R.; Kahn, C.R.; Grinspoon, S.K. Altered pattern of circulating miRNAs in HIV lipodystrophy perturbs key adipose differentiation and inflammation pathways. JCI Insight 2021, 6, e150399. [Google Scholar] [CrossRef] [PubMed]
- Squillace, N.; Bresciani, E.; Torsello, A.; Bandera, A.; Sabbatini, F.; Giovannetti, C.; Giunta, G.; Rovati, L.; Del Bene, M.; Locatelli, V.; et al. Changes in subcutaneous adipose tissue microRNA expression in HIV-infected patients. J. Antimicrob. Chemother. 2014, 69, 3067–3075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bresciani, E.; Saletti, C.; Squillace, N.; Rizzi, L.; Molteni, L.; Meanti, R.; Omeljaniuk, R.J.; Biagini, G.; Gori, A.; Locatelli, V.; et al. miRNA-218 Targets Lipin-1 and Glucose Transporter Type 4 Genes in 3T3-L1 Cells Treated with Lopinavir/Ritonavir. Front. Pharmacol. 2019, 10, 461. [Google Scholar] [CrossRef] [Green Version]
- Tarr, P.E.; Telenti, A. Toxicogenetics of antiretroviral therapy: Genetic factors that contribute to metabolic complications. Antivir. Ther. 2007, 12, 999–1013. [Google Scholar]
- Vidal, F.; Domingo, P.; Vilades, C.; Peraire, J.; Arnedo, M.; Alcami, J.; Leal, M.; Villarroya, F.; Gatell, J.M. Pharmacogenetics of the lipodystrophy syndrome associated with HIV infection and combination antiretroviral therapy. Expert Opin. Drug Metab. Toxicol. 2011, 7, 1365–1382. [Google Scholar] [CrossRef]
- Castilhos, J.K.; Sprinz, E.; Lazzaretti, R.K.; Kuhmmer, R.; Mattevi, V.S. Polymorphisms in adiponectin receptor genes are associated with lipodystrophy-related phenotypes in HIV-infected patients receiving antiretroviral therapy. HIV Med. 2015, 16, 494–501. [Google Scholar] [CrossRef] [Green Version]
- Egana-Gorrono, L.; Martinez, E.; Cormand, B.; Escribà, T.; Gatell, J.; Arnedo, M. Impact of genetic factors on dyslipidemia in HIV-infected patients starting antiretroviral therapy. AIDS 2013, 27, 529–538. [Google Scholar] [CrossRef]
- Rotger, M.; Gsponer, T.; Martinez, R.; Taffé, P.; Elzi, L.; Vernazza, P.; Cavassini, M.; Bernasconi, E.; Hirschel, B.; Furrer, H.; et al. Impact of single nucleotide polymorphisms and of clinical risk factors on new-onset diabetes mellitus in HIV-infected individuals. Clin. Infect. Dis. 2010, 51, 1090–1098. [Google Scholar] [CrossRef]
- Arnedo, M.; Taffe, P.; Sahli, R.; Furrer, H.; Hirschel, B.; Elzi, L.; Weber, R.; Vernazza, P.; Bernasconi, E.; Darioli, R.; et al. Contribution of 20 single nucleotide polymorphisms of 13 genes to dyslipidemia associated ith antiretroviral therapy. Pharmacogenet. Genom. 2007, 17, 755–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynn, F.C. Meta-regulation: microRNA regulation of glucose and lipid metabolism. Trends Endocrinol. Metab. 2009, 20, 452–459. [Google Scholar] [CrossRef] [PubMed]
- Torriani, M.; Srinivasa, S.; Fitch, K.V.; Thomou, T.; Wong, K.; Petrow, E.; Kahn, C.R.; Cypess, A.M.; Grinspoon, S.K. Dysfunctional subcutaneous fat with reduced dicer and brown adipose tissue gene expression in HIV-infected patients. J. Clin. Endocrinol. Metab. 2016, 101, 1225–1234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houzet, L.; Yeung, M.L.; de Lame, V.; Desai, D.; Smith, S.M.; Jeang, K.T. MicroRNA profile changes in human immunodeficiency virus type 1 (HIV-1) seropositive individuals. Retrovirology 2008, 5, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koshiol, J.; Wang, E.; Zhao, Y.; Marincola, F.; Landi, M.T. Strengths and limitations of laboratory procedures for microRNA detection. Cancer Epidemiol. Prev. Biomark. 2010, 19, 907–911. [Google Scholar] [CrossRef] [Green Version]
- Faraldi, M.; Gomarasca, M.; Banfi, G.; Lombardi, G. Free Circulating miRNAs Measurement in Clinical Settings: The Still Unsolved Issue of the Normalization. Adv. Clin. Chem. 2018, 87, 113–139. [Google Scholar]
- Schwarzenbach, H.; Machado da Silva, A.; Calin, G.; Pantel, K. Data Normalization Strategies for MicroRNA Quantification. Clin. Chem. 2015, 61, 11. [Google Scholar] [CrossRef]
- Jacobsen, K.S.; Nielsen, K.O.; Winther, T.N.; Glebe, D.; Pociot, F.; Hogh, B. Identification of valid reference genes for microRNA expression studies in a hepatitis B virus replicating liver cell line. BMC Res. Notes 2016, 9, 38. [Google Scholar] [CrossRef] [Green Version]
- Duan, Z.; Ca, G.; Li, J.; Bu, R.; Wang, N.; Yin, P.; Chen, X. U6 can be used as a housekeeping gene for urinary sediment miRNA studies of IgA nephropathy. Sci. Rep. 2018, 8, 10875. [Google Scholar] [CrossRef]
- Lou, G.; Ma, N.; Xu, Y.; Jiang, L.; Yang, J.; Wang, C.; Jiao, Y.; Gao, X. Differential distribution of U6 (RNU6–1) expression in human carcinoma tissues demonstrates the requirement for caution in the internal control gene selection for microRNA quantification. Int. J. Mol. Med. 2015, 36, 1400–1408. [Google Scholar] [CrossRef] [Green Version]
- Hirschberger, S.; Hübner, M.; Strauß, G.; Effinger, D.; Bauer, M.; Weis, S.; Giamarellos-Bourboulis, E.J.; Kreth, S. Identification of suitable controls for miRNA quantification in T-cells and whole blood cells in sepsis. Sci. Rep. 2019, 31, 15735. [Google Scholar] [CrossRef] [PubMed]
- Zárybnický, T.; Matoušková, P.; Ambrož, M.; Šubrt, Z.; Skálová, L.; Boušová, I. The selection and validation of reference genes for mRNA and microRNA expression studies in human liver slices using RT-qPCR. Genes 2019, 10, 763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalpachidou, T.; Kummer, K.K.; Mitrić, M.; Kress, M. Tissue specific reference genes for microRNA expression analysis in a mouse model of peripheral nerve injury. Front. Mol. Neurosci. 2019, 12, 283. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Liang, H.; Guan, D.; Wang, C.; Hu, X.; Cui, L.; Chen, S.; Zhang, C.; Zhang, J.; Zen, K.; et al. A Combination of Let-7d, Let-7g and Let-7i Serves as a Stable Reference for Normalization of Serum microRNAs. PLoS ONE 2017, 8, e79652. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Zhang, X.; Yuan, J.; Wu, J.; Deng, X.; Peng, J.; Wang, S.; Yang, C.; Ge, J.; Zou, Y. Evaluation of the performance of serum miRNAs as normalizers in microRNA studies focused on cardiovascular disease. J. Thorac. Dis. 2018, 10, 2599–2607. [Google Scholar] [CrossRef]
- Frías, M.; Rivero-Juárez, A.; Téllez, F.; Palacios, R.; Jiménez-Arranz, A.; Pineda, J.A.; Merino, D.; Gómez-Vidal, M.A.; Pérez-Camacho, I.; Camacho, A.; et al. Evaluation of hepatitis C viral RNA persistence in HIV-infected patients with long-term sustained virological response by droplet digital PCR. Sci. Rep. 2019, 9, 12507. [Google Scholar] [CrossRef] [Green Version]
- Alteri, C.; Scutari, R.; Stingone, C.; Maffongelli, G.; Brugneti, M.; Falasca, F.; Martini, S.; Bertoli, A.; Turriziani, O.; Sarmati, L.; et al. Quantification of HIV-DNA and residual viremia in patients starting ART by droplet digital PCR: Their dynamic decay and correlations with immunological parameters and virological success. J. Clin. Virol. 2019, 117, 61–67. [Google Scholar] [CrossRef]
- Beheshti, A.; Stevenson, K.; Vanderburg, C.; Ravi, D.; McDonald, J.T.; Christie, A.L.; Shigemori, K.; Jester, H.; Weinstock, D.M.; Evens, A.M. Identification of circulating serum multi-microRNA signatures in human DLBCL models. Sci. Rep. 2019, 9, 17161. [Google Scholar] [CrossRef]
- D’Antona, P.; Cattoni, M.; Dominioni, L.; Poli, A.; Moretti, F.; Cinquetti, R.; Gini, E.; Daffrè, E.; Noonan, D.M.; Imperatori, A.; et al. Serum miR-223: A validated biomarker for detection of early-stage non–small cell lung cancer. Cancer Epidemiol. Prev. Biomark. 2019, 28, 1926–1933. [Google Scholar] [CrossRef] [Green Version]
- Saucier, D.; Wajnberg, G.; Roy, J.; Beauregard, A.P.; Chacko, S.; Crapoulet, N.; Fournier, S.; Ghosh, A.; Lewis, S.M.; Marrero, A.; et al. Identification of a circulating miRNA signature in extracellular vesicles collected from amyotrophic lateral sclerosis patients. Brain Res. 2019, 1708, 100–108. [Google Scholar] [CrossRef]
- Maciejak, A.; Kostarska-Srokosz, E.; Gierlak, W.; Dluzniewski, M.; Kuch, M.; Marchel, M.; Opolski, G.; Kiliszek, M.; Matlak, K.; Dobrzycki, S.; et al. Circulating miR-30a-5p as a prognostic biomarker of left ventricular dysfunction after acute myocardial infarction. Sci. Rep. 2018, 8, 9883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolmeson, C.; Esguerra, J.L.; Salehi, A.; Speidel, D.; Eliasson, L.; Cilio, C.M. Differences in islet-enriched miRNAs in healthy and glucose intolerant human subjects. Biochem. Biophys. Res. Commun. 2011, 404, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Latorre, J.; Moreno-Navarrete, J.M.; Mercader, J.M.; Sabater, M.; Rovira, Ò.; Gironès, J.; Ricart, W.; Fernández-Real, J.M.; Ortega, F.J. Decreased lipid metabolism but increased FA biosynthesis are coupled with changes in liver microRNAs in obese subjects with NAFLD. Int. J. Obes. 2017, 41, 620–630. [Google Scholar] [CrossRef] [PubMed]
- Giralt, M.; Domingo, P.; Villarroya, F. Adipose tissue biology and HIV-infection. Best Pract. Res. Clin. Endocrinol. Metab. 2011, 25, 487–499. [Google Scholar] [CrossRef]
- Xi, Y.; Shen, W.; Ma, L.; Zhao, M.; Zheng, J.; Bu, S.; Hino, S.; Nakao, M. HMGA2 promotes adipogenesis by activating C/EBPb-mediated expression of PPARγ. Biochem. Biophys. Res. Commun. 2016, 472, 617–623. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Rui, B.; Tang, L.; Hu, C. Lipin family proteins—Key regulators in lipid metabolism. Ann. Nutr. Metab. 2015, 66, 10–18. [Google Scholar] [CrossRef]
- Biswas, S.; Haleyurgirisetty, M.; Lee, S.; Hewlett, I.; Devadas, K. Development and validation of plasma miRNA biomarker signature panel for the detection of early HIV-1 infection. EBioMedicine 2019, 43, 307–316. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Ding, Y.; Ma, Q.; Zhao, L.; Guo, X.; Shao, Y.; Niu, C.; He, Y.; Zhang, F.; Zheng, D.; et al. Identification of novel circulating miRNAs biomarkers for the diagnosis of esophageal squamous cell carcinoma and squamous dysplasia. Cancer Epidemiol. Prev. Biomark. 2019, 28, 1212–1220. [Google Scholar] [CrossRef] [Green Version]
- Dunaeva, M.; Blom, J.; Thurlings, R.; Pruijn, G.J.M. Circulating serum miR-223–3p and miR-16–5p as possible biomarkers of early rheumatoid arthritis. Clin. Exp. Immunol. 2018, 193, 376–385. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Song, Y.; Zhang, C.; Zhi, X.; Fu, H.; Ma, Y.; Chen, Y.; Pan, F.; Wang, K.; Ni, J.; et al. Circulating miR-16–5p and miR-19b-3p as two novel potential biomarkers to indicate progression of gastric cancer. Theranostics 2015, 5, 733–745. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Liu, J.; Chen, J.; Wang, H.; Yang, L.; Chen, F.; Fan, S.; Wang, J.; Shao, B.; Yin, D.; et al. A serum microRNA signature predicts trastuzumab benefit in HER2-positive metastatic breast cancer patients. Nat. Commun. 2018, 9, 1614. [Google Scholar] [CrossRef] [PubMed]
- Vickers, K.; Palmisano, B.; Shoucri, B.; Shamburek, R.D.; Remaley, A.T. MicroRNAs are transported in plasma and delivered to recipient cells by high-density lipoproteins. Nat. Cell Biol. 2011, 13, 423–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Xu, Z.; Ou, D.; Liu, J.; Zhang, J. The miR-15a/16 gene cluster in human cancer: A systematic review. J. Cell Physiol. 2019, 234, 5496–5506. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.J.; Pu, X.M.; Zhao, Z.F.; Zhao, Y.N.; Kang, X.J.; Wu, W.D.; Zou, Y.M.; Wu, C.Y.; Qu, Y.Y.; Zhang, D.Z.; et al. The expression profiles of microRNAs in Kaposi’s sarcoma. Tumor Biol. 2015, 36, 437–446. [Google Scholar] [CrossRef]
- Pla, A.; Zhong, X.; Rayner, S. miRAW: A deep learning-based approach to predict microRNA targets by analyzing whole microRNA transcripts. PLoS Comput. Biol. 2018, 14, e1006185. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | HIV+ | HIV− |
|---|---|---|
| BMI | 21.8 | 27.3 |
| Waist circumference (cm) | 80 | 93 |
| SBP (mmHg) | 120 | 120 |
| DBP (mmHg) | 60 | 80 |
| Glucose (mg/dL) | 92 | 94 |
| ALT (U/L) | 32 | 15 |
| Total cholesterol (mg/dL) | 179 | 172 |
| HDL-Cholesterol (mg/dL) | 41 | 46 |
| Triglycerides (mg/dL) | 78 | 124 |
| Real-Time qPCR FC Means ± SEM | |||||
|---|---|---|---|---|---|
| miRNAs Expressed in SC ADIPOSE TISSUE | Microarray Expression Values (HIV + vs. HIV −) | Microarray FC (>+2 or <−2) | Reference Gene U6 | Reference Gene miR-425-5p | |
| Upregulated | miR-15b-5p | 215/89 | 2.4 | 2.36 ± 0.44 | 1.36 ± 0.29 |
| miR-532-3p | 118/77 | 2.1 | 1.96 ± 0.53 | 0.98 ± 0.17 | |
| miR-127-3p | 76/32.8 | 2.3 | 4.59 ± 0.92 | 1.90 ± 0.20 | |
| Downregulated | miR-122-5p | 637.8/2150 | −3.4 | 0.25 ± 0.01 | 0.16 ± 0.02 |
| miR-6803-5p | 275.8/694.4 | −2.5 | 1.42 ± 0.74 | 0.30 ± 0.15 | |
| miR-3620-5p | 121.3/263.9 | −2.2 | 0.74 ± 0.08 | 0.45 ± 0.04 | |
| Real-Time qPCR FC Means | |||||
|---|---|---|---|---|---|
| miRNAs Expressed in PLASMA | Microarray Expression Values (HIV + vs. HIV −) | Microarray FC | Reference Gene Let-7i | Reference Gene miR-425-5p | |
| Upregulated | miR-4529-3p | 950.5/399.7 | 2.4 | Indetermined | Indetermined |
| miR-16-5p | 857.2/118 | 7.3 | 0.90 | 0.92 | |
| miR- 6803-5p | 333.8/52 | 6.4 | 3.91 | 4.12 | |
| miR-4707-5p | 262.8/71 | 3.1 | Indetermined | Indetermined | |
| miR-668-5p | 336.8/156.4 | 2.2 | Indetermined | Indetermined | |
| Downregulated | miR-877-5p | 82/244.5 | −3.0 | 2.32 | 1.89 |
| miR-642b-3p | 1.8/6.0 | −3.4 | Indetermined | Indetermined | |
| SC ADIPOSE TISSUE and PLASMA PRIMER SET | |
|---|---|
| Candidate miRNAs | Exiqon ID |
| has-miR-15b-5p | 002042432 |
| has-miR-532-3p | 00204003 |
| has-miR-127-3p | 00204048 |
| has-miR-122-5p | 00205664 |
| has-miR-6803-5p | 02111873 |
| has-miR-3620-5p | 02102506 |
| has-miR-16-5p | 00205702 |
| has-miR-4529-3p | 02118987 |
| has-miR-877-5p | 00205626 |
| has-miR-642b-3p | 02103759 |
| has-miR-4707-5p | 02116591 |
| has-miR-668-5p | 02108164 |
| has-miR-425-5p | 00204337 |
| has-Let7i-5p | 204394 |
| snU6 | 203907 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bresciani, E.; Squillace, N.; Orsini, V.; Piolini, R.; Rizzi, L.; Molteni, L.; Meanti, R.; Soria, A.; Lapadula, G.; Bandera, A.; et al. miRNA Expression Profiling in Subcutaneous Adipose Tissue of Monozygotic Twins Discordant for HIV Infection: Validation of Differentially Expressed miRNA and Bioinformatic Analysis. Int. J. Mol. Sci. 2022, 23, 3486. https://doi.org/10.3390/ijms23073486
Bresciani E, Squillace N, Orsini V, Piolini R, Rizzi L, Molteni L, Meanti R, Soria A, Lapadula G, Bandera A, et al. miRNA Expression Profiling in Subcutaneous Adipose Tissue of Monozygotic Twins Discordant for HIV Infection: Validation of Differentially Expressed miRNA and Bioinformatic Analysis. International Journal of Molecular Sciences. 2022; 23(7):3486. https://doi.org/10.3390/ijms23073486
Chicago/Turabian StyleBresciani, Elena, Nicola Squillace, Valentina Orsini, Roberta Piolini, Laura Rizzi, Laura Molteni, Ramona Meanti, Alessandro Soria, Giuseppe Lapadula, Alessandra Bandera, and et al. 2022. "miRNA Expression Profiling in Subcutaneous Adipose Tissue of Monozygotic Twins Discordant for HIV Infection: Validation of Differentially Expressed miRNA and Bioinformatic Analysis" International Journal of Molecular Sciences 23, no. 7: 3486. https://doi.org/10.3390/ijms23073486