
Identification and DNA Marker Development for a Wheat-Leymus mollis 2Ns (2D) Disomic Chromosome Substitution

 and
and
Abstract
:1. Introduction
2. Results
2.1. Cytological Characterization of 17DM48
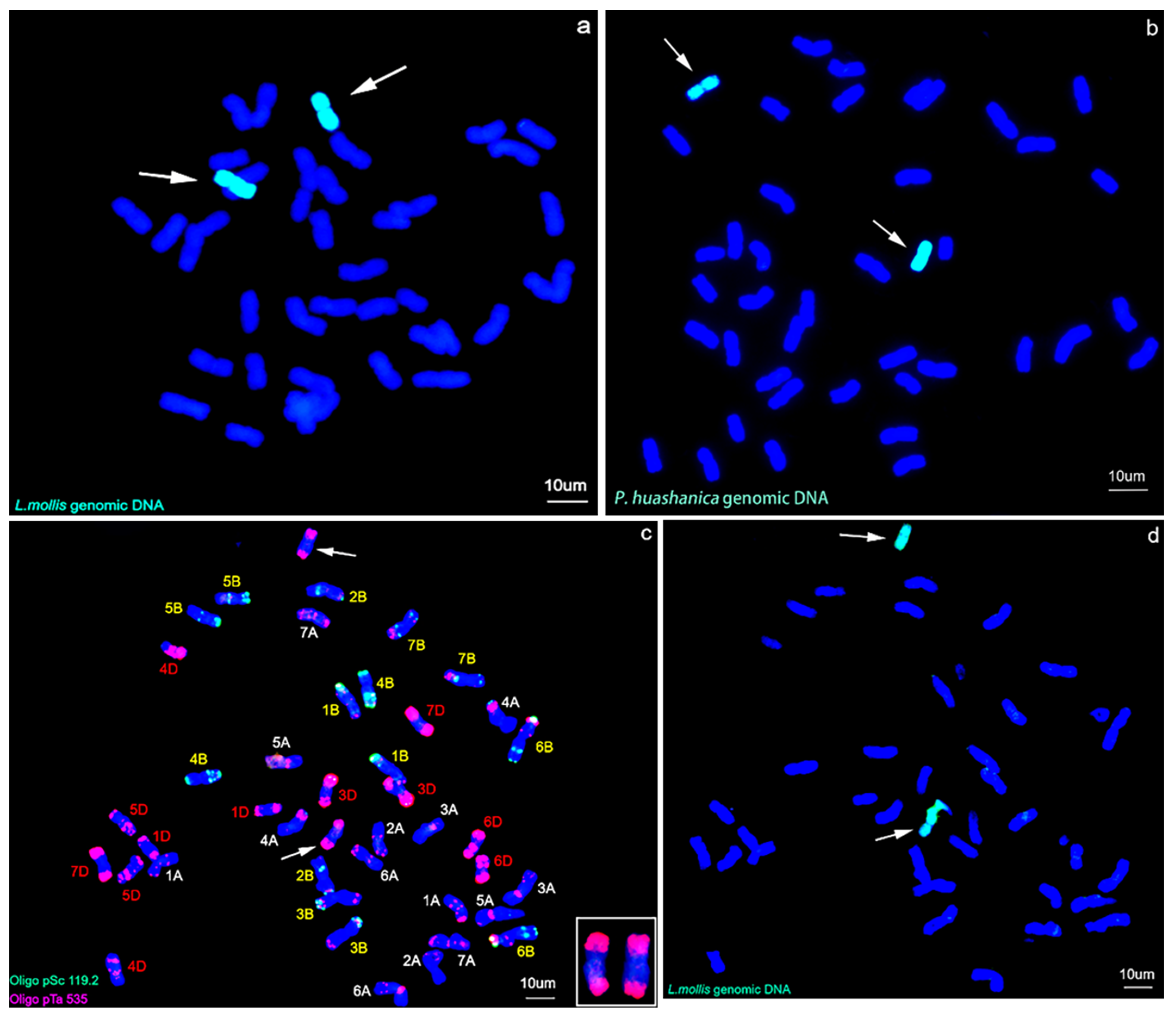
2.2. GISH and Sequential FISH–GISH Analysis
2.3. Wheat 55K SNP Array Analysis
2.4. Molecular Marker Analysis
2.5. Evaluation of Agronomic Traits
2.6. Molecular Marker Development
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Cytological Observation
4.3. GISH, FISH and Sequential FISH–GISH
4.4. Wheat SNP Array Analysis
4.5. Molecular Markers Analysis
4.6. Molecular Marker Development
4.7. Morphological Traits Evaluation
4.8. Disease Reaction Evaluation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wellings, C.R. Global status of stripe rust: A review of historical and current threats. Euphytica 2011, 179, 129–141. [Google Scholar] [CrossRef]
- Friebe, B.; Jiang, J.; Raupp, W.J.; McIntosh, R.A.; Gill, B.S. Characterization of wheat-alien translocations conferring resistance to diseases and pests: Current status. Euphytica 1996, 91, 59–87. [Google Scholar] [CrossRef]
- Merker, A. The Triticeae in cereal breeding. Hereditas 2008, 116, 277–280. [Google Scholar] [CrossRef]
- Fatih, A.M.B. Analysis of the breeding potential of wheat-Agropyron and wheat-Elymus derivatives. Hereditas 2008, 98, 287–295. [Google Scholar] [CrossRef]
- Wang, J.; Chen, X.-H.; Du, W.-L.; Zhao, J.-X.; Wu, J.; Cheng, X.-N.; Pang, Y.-H.; Yang, Q.-H.; Liu, S.-H.; Fu, J. Morphological and molecular cytogenetic characterization of partial octoploid Tritileymus. Genet. Resour. Crop Evol. 2013, 60, 1453–1462. [Google Scholar] [CrossRef]
- Anamthawat-Jónsson, K. Variable genome composition in Triticum × Leymus amphiploids. Theor. Appl. Genet. 1999, 99, 1087–1093. [Google Scholar] [CrossRef]
- Merker, A.; Lantai, K. Hybrids between wheats and perennial Leymus and Thinopyrum species. Acta Agric. Scand. 1997, 47, 48–51. [Google Scholar] [CrossRef]
- Anamthawat-Jónsson, K.; Bodvarsdottir, S.K.; Bragason, B.T.; Gudmundsson, J.; Martin, P.; Koebner, R. Wide hybridization between wheat (Triticum L.) and lymegrass (Leymus Hochst.). Euphytica 1997, 93, 293–300. [Google Scholar] [CrossRef]
- Ceoloni, C.; Kuzmanović, L.; Gennaro, A.; Forte, P.; Giorgi, D.; Grossi, M.R.; Bitti, A. Genomes, Chromosomes and Genes of the Wheatgrass Genus Thinopyrum: The Value of their Transfer into Wheat for Gains in Cytogenomic Knowledge and Sustainable Breeding; Springer Science and Business Media LLC: Dordrecht, The Netherlands, 2014; pp. 333–358. [Google Scholar]
- Melak, S.; Wang, Q.; Tian, Y.; Wei, W.; Zhang, L.; Elbeltagy, A.; Chen, J. Identification and Validation of Marketing Weight-Related SNP Markers Using SLAF Sequencing in Male Yangzhou Geese. Genes 2021, 12, 1203. [Google Scholar] [CrossRef]
- Wang, S.; Wang, C.; Feng, X.; Zhao, J.; Deng, P.; Wang, Y.; Chen, C.; Wang, B.; Ji, W. Molecular Cytogenetics of Chromosome 2St as Well as Chromosome 3St Derived from Thinopyrum Intermedium and Thinopyrum Ponticum; Research Square Platform LLC: Singapore, 2021. [Google Scholar] [CrossRef]
- Wang, Y.; Cao, Q.; Zhang, J.; Wang, S.; Chen, C.; Wang, C.; Zhang, H.; Wang, Y.; Ji, W. Cytogenetic Analysis and Molecular Marker Development for a New Wheat–Thinopyrum ponticum 1Js (1D) Disomic Substitution Line with Resistance to Stripe Rust and Powdery Mildew. Front. Plant Sci. 2020, 11, 11. [Google Scholar] [CrossRef]
- Chen, S.; Huang, Z.; Dai, Y.; Qin, S.; Gao, Y.; Zhang, L.; Gao, Y.; Chen, J. The Development of 7E Chromosome-Specific Molecular Markers for Thinopyrum elongatum Based on SLAF-seq Technology. PLoS ONE 2013, 8, e65122. [Google Scholar] [CrossRef]
- Tang, Z.; Yang, Z.; Fu, S. Oligonucleotides replacing the roles of repetitive sequences pAs1, pSc119.2, pTa-535, pTa71, CCS1, and pAWRC.1 for FISH analysis. J. Appl. Genet. 2014, 55, 313–318. [Google Scholar] [CrossRef]
- Nguyen, V.; Fleury, D.; Timmins, A.; Laga, H.; Hayden, M.; Mather, D.; Okada, T. Addition of rye chromosome 4R to wheat increases anther length and pollen grain number. Theor. Appl. Genet. 2015, 128, 953–964. [Google Scholar] [CrossRef]
- Silkova, O.G.; Shchapova, A.I.; Shumnyĭ, V.K. Role of rye chromosome 2R from wheat-rye substitution line 2R(2D) (Triticum aestivum L. cv. Saratovskaya 29-Secale cereale L. cv. Onokhoiskaya) in genetic regulation of meiotic restitution in wheat-rye polyhaploids. Genetika 2007, 43, 971–981. [Google Scholar] [CrossRef]
- Silkova, O.G.; Dobrovol’skaia, O.B.; Dubovets, N.I.; Adonina, I.G.; Kravtsova, L.A.; Roder, M.S.; Salina, E.A.; Shchapova, A.I.; Shumnyĭ, V.K. Production of wheat-rye substitution lines and identification of chromosome composition of karyotypes using C-banding, GISH, and SSR markers. Genetika 2006, 42, 793–802. [Google Scholar] [CrossRef]
- Lukaszewski, A.J.; Porter, D.R.; Baker, C.A.; Rybka, K.; Lapinski, B. Attempts to Transfer Russian Wheat Aphid resistance from a Rye Chromosomes in Russian Triticales. Crop Sci. 2001, 41, 1743–1749. [Google Scholar] [CrossRef] [Green Version]
- Kishii, M.; Wang, R.R.-C.; Tsujimoto, H. Characteristics and behaviour of the chromosomes of Leymus mollis and L. racemosus (Triticeae, Poaceae) during mitosis and meiosis. Chromosom. Res. 2003, 11, 741–748. [Google Scholar] [CrossRef]
- Cifuentes, M.; Benavente, E. Wheat-alien metaphase I pairing of individual wheat genomes and D genome chromosomes in interspecific hybrids between Triticum aestivum L. and Aegilops geniculata Roth. Theor. Appl. Genet. 2009, 119, 805–813. [Google Scholar] [CrossRef]
- Han, F.; Liu, B.; Fedak, G.; Liu, Z. Genomic constitution and variation in five partial amphiploids of wheat Thinopyrum intermedium as revealed by GISH, multicolor GISH and seed storage protein analysis. Theor. Appl. Genet. 2004, 109, 1070–1076. [Google Scholar] [CrossRef]
- Malysheva, L.; Sjakste, T.; Matzk, F.; Röder, M.; Ganal, M. Molecular cytogenetic analysis of wheat–barley hybrids using genomic in situ hybridization and barley microsatellite markers. Genome 2003, 46, 314–322. [Google Scholar] [CrossRef]
- Nagy, E.D.; Molnár-Láng, M.; Linc, G.; Láng, L. Identification of wheat-barley translocations by sequential GISH and two-colour FISH in combination with the use of genetically mapped barley SSR markers. Genome 2002, 45, 1238–1247. [Google Scholar] [CrossRef]
- Friebe, B.; Kynast, R.G.; Gill, B.S. Gametocidal factor-induced structural rearrangements in rye chromosomes added to common wheat. Chromosom. Res. 2000, 8, 501–511. [Google Scholar] [CrossRef]
- Benavente, E.; Fernández-Calvín, B.; Orellana, J. Relationship between the levels of wheat-rye metaphase I chromosomal pairing and recombination revealed by GISH. Chromosoma 1996, 105, 92–96. [Google Scholar] [CrossRef]
- Rasheed, A.; Hao, Y.; Xia, X.; Khan, A.; Xu, Y.; Varshney, R.; He, Z. Crop Breeding Chips and Genotyping Platforms: Progress, Challenges, and Perspectives. Mol. Plant 2017, 10, 1047–1064. [Google Scholar] [CrossRef] [Green Version]
- Winfield, M.O.; Allen, A.M.; Burridge, A.; Barker, G.L.A.; Benbow, H.R.; Wilkinson, P.A.; Coghill, J.; Waterfall, C.; Davassi, A.; Scopes, G.; et al. High-density SNP genotyping array for hexaploid wheat and its secondary and tertiary gene pool. Plant Biotechnol. J. 2016, 14, 1195–1206. [Google Scholar] [CrossRef]
- Li, J.; Yao, X.; Yang, Z.; Cheng, X.; Yuan, F.; Liu, Y.; Wu, J.; Yang, Q.; Zhao, J.; Chen, X. Molecular cytogenetic characterization of a novel wheat–Psathyrostachys huashanica Keng 5Ns (5D) disomic substitution line with stripe rust resistance. Mol. Breed. 2019, 39, 109. [Google Scholar] [CrossRef]
- Sun, X.; Liu, D.; Zhang, X.; Li, W.; Liu, H.; Hong, W.; Jiang, C.; Guan, N.; Ma, C.; Zeng, H.; et al. SLAF-seq: An Efficient Method of Large-Scale De Novo SNP Discovery and Genotyping Using High-Throughput Sequencing. PLoS ONE 2013, 8, e58700. [Google Scholar] [CrossRef]
- Naranjo, T. The Effect of Chromosome Structure upon Meiotic Homologous and Homoeologous Recombinations in Triticeae. Agronomy 2019, 9, 552. [Google Scholar] [CrossRef] [Green Version]
- Devos, K.M.; Atkinson, M.D.; Chinoy, C.N.; Francis, H.A.; Harcourt, R.L.; Koebner, R.M.D.; Liu, C.J.; Masojć, P.; Xie, D.X.; Gale, M.D. Chromosomal rearrangements in the rye genome relative to that of wheat. Theor. Appl. Genet. 1993, 85, 673–680. [Google Scholar] [CrossRef]
- Naranjo, T.; Fernández-Rueda, P. Homoeology of rye chromosome arms to wheat. Theor. Appl. Genet. 1991, 82, 577–586. [Google Scholar] [CrossRef]
- Dvorak, J.; Wang, L.; Zhu, T.; Jorgensen, C.M.; Luo, M.-C.; Deal, K.R.; Gu, Y.Q.; Gill, B.S.; Distelfeld, A.; Devos, K.M.; et al. Reassessment of the evolution of wheat chromosomes 4A, 5A, and 7B. Theor. Appl. Genet. 2018, 131, 2451–2462. [Google Scholar] [CrossRef] [Green Version]
- Devos, K.M.; Gale, M.D. Extended genetic maps of the homoeologous group 3 chromosomes of wheat, rye and barley. Theor. Appl. Genet. 1993, 85, 649–652. [Google Scholar] [CrossRef]
- Naranjo, T.; Fernández-Rueda, P. Pairing and recombination between individual chromosomes of wheat and rye in hybrids carrying the ph1b mutation. Theor. Appl. Genet. 1996, 93, 242–248. [Google Scholar] [CrossRef]
- Larson, S.R.; Kishii, M.; Tsujimoto, H.; Qi, L.; Chen, P.; Lazo, G.R.; Jensen, K.B.; Wang, R.R.-C. Leymus EST linkage maps identify 4NsL–5NsL reciprocal translocation, wheat-Leymus chromosome introgressions, and functionally important gene loci. Theor. Appl. Genet. 2011, 124, 189–206. [Google Scholar] [CrossRef]
- McIntosh, R.; Mu, J.; Han, D.; Kang, Z. Wheat stripe rust resistance gene Yr24/Yr26: A retrospective review. Crop J. 2018, 6, 3–11. [Google Scholar] [CrossRef]
- Yang, X.F.; Wang, C.Y.; Chen, C.H.; Tian, Z.R.; Ji, W.Q. Development and characterization of a wheat–Leymus mollis Lm#7Ns disomic addition line with resistance to stripe rust. Cereal Res. Commun. 2020, 48, 467–476. [Google Scholar] [CrossRef]
- Xin, L.I.; Yang, X.; Bing, M.A.; Chen, Z.; Wang, C.; Chen, C.; Tian, Z.; Wanquan, J.I.; Aamp, N.; University, F. Molecular Cytogenetics Identification of a Wheat-Leymus mollis Derivative with Resistance to Stripe Rust. J. Triticeae Crops. 2016, 8, 556–563. [Google Scholar]
- Yang, X.; Li, X.; Wang, C.; Chen, C.; Tian, Z.; Ji, W. Isolation and molecular cytogenetic characterization of a wheat–Leymus mollis double monosomic addition line and its progenies with resistance to stripe rust. Genome 2017, 60, 1029–1036. [Google Scholar] [CrossRef]
- Zhao, J.; Liu, Y.; Cheng, X.; Pang, Y.; Li, J.; Su, Z.; Wu, J.; Yang, Q.; Bai, G.; Chen, X. Development and identification of a dwarf wheat-Leymus mollis double substitution line with resistance to yellow rust and Fusarium head blight. Crop J. 2019, 7, 516–526. [Google Scholar] [CrossRef]
- Pang, Y.; Chen, X.; Zhao, J.; Du, W.; Cheng, X.; Wu, J.; Li, Y.; Wang, L.; Wang, J.; Yang, Q. Molecular Cytogenetic Characterization of a Wheat–Leymus mollis 3D(3Ns) Substitution Line with Resistance to Leaf Rust. J. Genet. Genom. 2014, 41, 205–214. [Google Scholar] [CrossRef]
- Falke, K.C.; Sušić, Z.; Wilde, P.; Wortmann, H.; Möhring, J.; Piepho, H.-P.; Geiger, H.H.; Miedaner, T. Testcross performance of rye introgression lines developed by marker-assisted backcrossing using an Iranian accession as donor. Theor. Appl. Genet. 2009, 118, 1225–1238. [Google Scholar] [CrossRef]
- Endo, T.R. The gametocidal chromosome as a tool for chromosome manipulation in wheat. Chromosom. Res. 2007, 15, 67–75. [Google Scholar] [CrossRef]
- Masoudi-Nejad, A.; Nasuda, S.; McIntosh, R.A.; Endo, T.R. Transfer of rye chromosome segments to wheat by a gametocidal system. Chromosom. Res. 2002, 10, 349–357. [Google Scholar] [CrossRef]
- Friebe, B.; Hatchett, J.H.; Gill, B.S.; Mukai, Y.; Sebesta, E.E. Transfer of Hessian fly resistance from rye to wheat via radiation-induced terminal and intercalary chromosomal translocations. Theor. Appl. Genet. 1991, 83, 33–40. [Google Scholar] [CrossRef]
- Han, F.; Lamb, J.C.; Birchler, J.A. High frequency of centromere inactivation resulting in stable dicentric chromosomes of maize. Proc. Natl. Acad. Sci. USA 2006, 103, 3238–3243. [Google Scholar] [CrossRef] [Green Version]
- Doyle, J. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Liu, L.; Luo, Q.; Teng, W.; Li, B.; Li, H.; Li, Y.; Li, Z.; Zheng, Q. Development of Thinopyrum ponticum-specific molecular markers and FISH probes based on SLAF-seq technology. Planta 2018, 247, 1099–1108. [Google Scholar] [CrossRef]
- Tiwari, V.K.; Wang, S.; Sehgal, S.; Vrána, J.; Friebe, B.; Kubaláková, M.; Chhuneja, P.; Doležel, J.; Akhunov, E.; Kalia, B.; et al. SNP Discovery for mapping alien introgressions in wheat. BMC Genom. 2014, 15, 273. [Google Scholar] [CrossRef] [Green Version]
- Georgieva, M.; Sepsi, A.; Tyankova, N.; Molnár-Láng, M. Molecular cytogenetic characterization of two high protein wheat-Thinopyrum intermedium partial amphiploids. J. Appl. Genet. 2011, 52, 269–277. [Google Scholar] [CrossRef]
- Sanguinetti, C.J.; Dias-Neto, E.; Simpson, A.J. Rapid silver staining and recovery of PCR products separated on polyacrylamide gels. Biotechniques 1994, 17, 914–921. [Google Scholar]
- Bao, Y.; Wang, J.; He, F.; Ma, H.; Wang, H. Molecular cytogenetic identification of a wheat (Triticum aestivum)-American dune grass (Leymus mollis) translocation line resistant to stripe rust. Genet. Mol. Res. 2012, 11, 3198–3206. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate long-read alignment with Burrows–Wheeler transform. Bioinformatics 2010, 26, 589–595. [Google Scholar] [CrossRef] [Green Version]
- Saari, E.E.; Prescott, J.M. A scale for appraising the foliar intensity of wheat disease. Plant Dis. Rep. 1975, 59, 377–380. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Material | Plant Height (cm) | Tiller Number | Spike Length (cm) | Florets Per Spike | Spikelets Per Spike | Kernels Per Spike | Thousand Kernel Weight (g) | Kernel Length (mm) | Kernel Width (mm) | Awn Type | Stripe Rust Reaction (IT) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| M842 | 86.10 ± 3.32 Aa | 9 ± 5 Bb | 11.75 ± 1.21 Bb | 89 ± 6 BCc | 21 ± 1 Cc | 43 ± 17 Bb | 34.81 ± 1.27 Cc | 8.53 ± 0.08 Aa | 3.34 ± 0.03 Dd | short awn | 0 |
| D4286 | 72.88 ± 4.58 Bc | 10 ± 2 Bb | 9.69 ± 0.72 Cc | 71 ± 6 Cd | 24 ± 1 Bb | 47 ± 13 Bb | 57.01 ± 1.58 Aa | 8.08 ± 0.05 Cc | 3.56 ± 0.02 Bb | long awn | 1 |
| 7182 | 83.15 ± 5.75 ABab | 16 ± 4 Aa | 11.08 ± 1.04 Bb | 109 ± 7 Bb | 24 ± 1 Bb | 65 ± 11 Aa | 30.53 ± 1.34 Dd | 6.45 ± 0.06 Dd | 3.44 ± 0.01 Cc | long awn | 3 |
| 17DM48 | 78.55 ± 7.07 Bb | 9 ± 3 Bb | 19.05 ± 1.34 Aa | 203 ± 34 Aa | 28 ± 2 Aa | 8 ± 4 Cc | 54.95 ± 1.08 Bb | 8.31 ± 0.01 Bb | 3.69 ± 0.01 Aa | short awn | 1 |
| Huixianhong | 4 |
| Marker | Tm (°C) | Primer (5′-3′) |
|---|---|---|
| LM19474 | 52 | F: TCGTCTGGGTTTGCTTAT |
| R: CACCGATTTCCAAGTTTC | ||
| LM19428 | 56 | F: CGTCATCCTCCACCACCT |
| R: ACGCAATCTGCTCAACCC | ||
| LM13006 | 56 | F: TGCGGTTGCGTCTATTGG |
| R: TGCTGGTGCATCATCATCG | ||
| LM17228 | 56 | F: GCTCCTTTCTCGCTTGCT |
| R: TGGACCGCTACGTTTGAC | ||
| LM25058 | 54 | F: AGGAAGGGTCGGAAACTC |
| R: AACACCACGGAATGAAGC | ||
| LM152390 | 52 | F: TTTCTAGCCGCTAAAGGT |
| R: TTTCCAAGCCTACTCCTG | ||
| LM65677 | 54 | F: CAGAGCATAACCCAGGAG |
| R: CCATAGGAACAAGCCAGA | ||
| LM224473 | 54 | F: GGACGGTGAGCAAGAAGG |
| R: CGTAATGCCCACGAAACA | ||
| LM33865 | 52 | F: GCTAGTAAATCGGAGGAC |
| R: TAGCCATAACACCAATCC | ||
| LM7529 | 52 | F: AGGTTTCCAAATAAGGGAT |
| R: CGGACCGTGAATACTCTG | ||
| LM12508 | 54 | F: TCACGGCATACAACAAGG |
| R: TATCCACCGACCACTCAA | ||
| LM23891 | 56 | F: TGGGCAACCGATGCTCTA |
| R: ACTGGCACGAATCCGTCT | ||
| LM51499 | 56 | F: CAGCAGTGGCTTCTGTTCC |
| R: TGTATGTGCGGGAGTGGA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, X.; Du, X.; Wang, S.; Deng, P.; Wang, Y.; Shang, L.; Tian, Z.; Wang, C.; Chen, C.; Zhao, J.; et al. Identification and DNA Marker Development for a Wheat-Leymus mollis 2Ns (2D) Disomic Chromosome Substitution. Int. J. Mol. Sci. 2022, 23, 2676. https://doi.org/10.3390/ijms23052676
Feng X, Du X, Wang S, Deng P, Wang Y, Shang L, Tian Z, Wang C, Chen C, Zhao J, et al. Identification and DNA Marker Development for a Wheat-Leymus mollis 2Ns (2D) Disomic Chromosome Substitution. International Journal of Molecular Sciences. 2022; 23(5):2676. https://doi.org/10.3390/ijms23052676
Chicago/Turabian StyleFeng, Xianbo, Xin Du, Siwen Wang, Pingchuan Deng, Yongfu Wang, Lihui Shang, Zengrong Tian, Changyou Wang, Chunhuan Chen, Jixin Zhao, and et al. 2022. "Identification and DNA Marker Development for a Wheat-Leymus mollis 2Ns (2D) Disomic Chromosome Substitution" International Journal of Molecular Sciences 23, no. 5: 2676. https://doi.org/10.3390/ijms23052676