
Untargeted LC-MS/MS Metabolomics Study on the MCF-7 Cell Line in the Presence of Valproic Acid

 , , , and
, , , and
Abstract
:1. Introduction
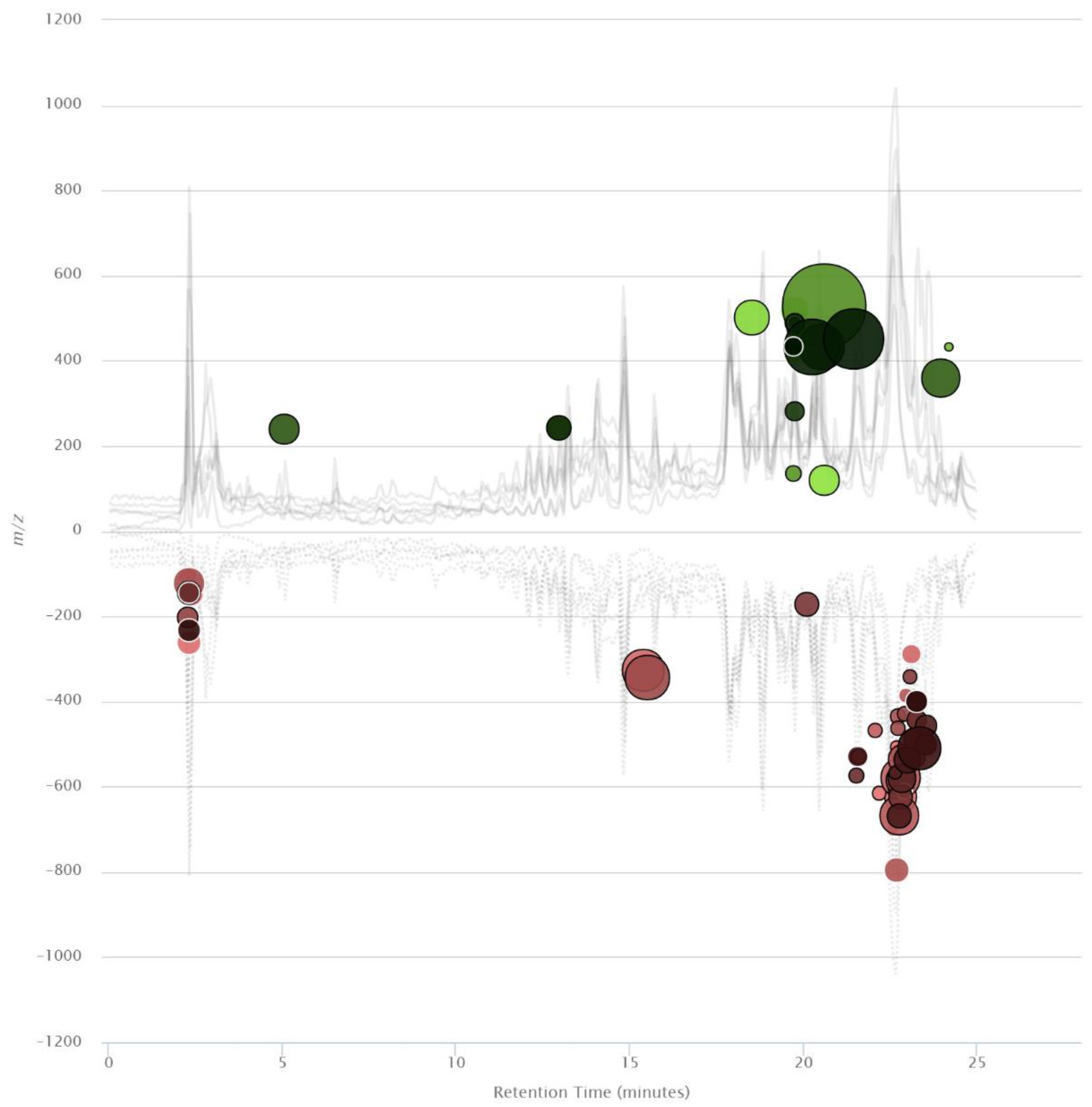
2. Results and Discussion
Metabolomics Analysis of MCF-7 Treated with VPA
3. Material and Methods
3.1. Cell Culture and Treatment
3.2. Steps for Metabolites Extraction
3.2.1. UHPLC-MS/MS Data Acquisition
3.2.2. LC-MS/MS Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BC | breast cancer |
| CHCl3 | Chloroform |
| DPBS | Dulbecco’s phosphate-buffered saline |
| HAT | histone acetyltransferase |
| HDAC | histone deacetylase |
| HDACi | histone deacetylase inhibitor |
| MeOH | methanol |
| NAD | nicotinamide adenine dinucleotide |
| PPP | pentose phosphate pathway |
| ROS | reactive oxygen species |
| RPE | ribulose-5-phosphate epimerase |
| RPI | ribose-5-phosphate isomerase |
| SAHA | suberoylanilide hydroxamic acid |
| TKT | transketolase |
| TALDO1 | transaldolase |
| VPA | valproic acid |
References
- Akram, M.; Iqbal, M.; Daniyal, M.; Khan, A.U. Awareness and current knowledge of breast cancer. Biol. Res. 2017, 50, 33. [Google Scholar] [CrossRef] [Green Version]
- Al-thoubaity, F.K. Molecular classification of breast cancer: A retrospective cohort study. Ann. Med. Surg. 2020, 49, 44–48. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Terry, M.B.; Antoniou, A.C.; Phillips, K.A.; Kast, K.; Mooij, T.M.; Engel, C.; Nogues, C.; Stoppa-Lyonnet, D.; Lasset, C.; et al. Alcohol consumption, cigarette smoking, and risk of breast cancer for BRCA1 and BRCA2 mutation carriers: Results from the BRCA1 and BRCA2 cohort consortium. Cancer Epidemiol. Biomark. Prev. 2020, 29, 368–378. [Google Scholar] [CrossRef]
- Arruebo, M.; Vilaboa, N.; Sáez-Gutierrez, B.; Lambea, J.; Tres, A.; Valladares, M.; González-Fernández, Á. Assessment of the evolution of cancer treatment therapies. Cancers 2011, 3, 3279–3330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.Y.; Ju, D.T.; Chang, C.F.; Muralidhar Reddy, P.; Velmurugan, B.K. A review on the effects of current chemotherapy drugs and natural agents in treating non-small cell lung cancer. Biomedicine 2017, 7, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toh, T.B.; Lim, J.J.; Chow, E.K.H. Epigenetics in cancer stem cells. Mol. Cancer 2017, 16, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Annunziato, A.T.; Hansen, J.C. Role of histone acetylation in the assembly and modulation of chromatin structures. Gene Expr. 2000, 9, 37–61. [Google Scholar] [CrossRef]
- Simone, C.; Peserico, A. Physical and functional HAT/HDAC interplay regulates protein acetylation balance. J. Biomed. Biotechnol. 2010, 2011, 371832. [Google Scholar] [CrossRef] [Green Version]
- Parbin, S.; Kar, S.; Shilpi, A.; Sengupta, D.; Deb, M.; Rath, S.K.; Patra, S.K. Histone Deacetylases: A Saga of Perturbed Acetylation Homeostasis in Cancer. J. Histochem. Cytochem. 2014, 62, 11–33. [Google Scholar] [CrossRef] [Green Version]
- Koprinarova, M.; Schnekenburger, M.; Diederich, M. Role of Histone Acetylation in Cell Cycle Regulation. Curr. Top. Med. Chem. 2016, 16, 732–744. [Google Scholar] [CrossRef]
- Narita, T.; Weinert, B.T.; Choudhary, C. Functions and mechanisms of non-histone protein acetylation. Nat. Rev. Mol. Cell Biol. 2019, 20, 156–174. [Google Scholar] [CrossRef] [PubMed]
- Eckschlager, T.; Plch, J.; Stiborova, M.; Hrabeta, J. Histone deacetylase inhibitors as anticancer drugs. Int. J. Mol. Sci. 2017, 18, 1414. [Google Scholar] [CrossRef] [PubMed]
- West, A.C.; Johnstone, R.W. New and emerging HDAC inhibitors for cancer treatment. J. Clin. Investig. 2014, 124, 30–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seto, E.; Yoshida, M. Erasers of histone acetylation: The histone deacetylase enzymes. Cold Spring Harb. Perspect. Biol. 2014, 6, 1–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.Y.; Jun, J.I.A.E.; Jeong, K.J.; Heo, H.J.; Sohn, J.S.; Lee, H.Y.; Park, C.G.; Kang, J. Histone deacetylases 1, 6 and 8 are critical for invasion in breast cancer. Oncol. Rep. 2011, 25, 1677–1681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, B.; Hong, J. An Overview of Naturally Occurring Histone Deacetylase Inhibitors. Curr. Top. Med. Chem. 2015, 14, 2759–2782. [Google Scholar] [CrossRef]
- Mawatari, T.; Ninomiya, I.; Inokuchi, M.; Harada, S.; Hayashi, H.; Oyama, K.; Makino, I.; Nakagawara, H.; Miyashita, T.; Tajima, H.; et al. Valproic acid inhibits proliferation of HER2-expressing breast cancer cells by inducing cell cycle arrest and apoptosis through Hsp70 acetylation. Int. J. Oncol. 2015, 47, 2073–2081. [Google Scholar] [CrossRef] [Green Version]
- Greer, C.B.; Tanaka, Y.; Kim, Y.J.; Xie, P.; Zhang, M.Q.; Park, I.H.; Kim, T.H. Histone Deacetylases Positively Regulate Transcription through the Elongation Machinery. Cell Rep. 2015, 13, 1444–1455. [Google Scholar] [CrossRef] [Green Version]
- Chiang, Y.S.; Huang, Y.F.; Midha, M.K.; Chen, T.H.; Shiau, H.C.; Chiu, K.P. Single cell transcriptome analysis of MCF-7 reveals consistently and inconsistently expressed gene groups each associated with distinct cellular localization and functions. PLoS ONE 2018, 13, e0199471. [Google Scholar] [CrossRef]
- Lo, Y.L.; Ho, C.T.; Tsai, F.L. Inhibit multidrug resistance and induce apoptosis by using glycocholic acid and epirubicin. Eur. J. Pharm. Sci. 2008, 35, 51–67. [Google Scholar] [CrossRef]
- Raju, U.; Katz, J.; Levitz, M. Effect of bile acids and estradiol on thymidine incorporation into DNA in MCF-7 and MCF-10A breast cell lines. Steroids 1997, 62, 643–646. [Google Scholar] [CrossRef]
- Di Ciaula, A.; Wang, D.Q.H.; Molina, E.M.; Baccetto, R.L.; Calamita, G.; Palmieri, V.O.; Portincasa, P. Bile acids and cancer: Direct and environmental-dependent effects. Ann. Hepatol. 2017, 16, S87–S105. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.; Dayoub, R.; Krautbauer, S.; Liebisch, G.; Wege, A.K.; Melter, M.; Weiss, T.S. Bile acid-induced apoptosis and bile acid synthesis are reduced by over-expression of Augmenter of Liver Regeneration (ALR) in a STAT3-dependent mechanism. Exp. Cell Res. 2019, 374, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Mitro, N.; Godio, C.; De Fabiani, E.; Scotti, E.; Galmozzi, A.; Gilardi, F.; Caruso, D.; Chacon, A.B.V.; Crestani, M. Insights in the regulation of cholesterol 7α-hydroxylase gene reveal a target for modulating bile acid synthesis. Hepatology 2007, 46, 885–897. [Google Scholar] [CrossRef]
- Frederiks, W.M.; Vizan, P.; Bosch, K.S.; Vreeling-Sindelárová, H.; Boren, J.; Cascante, M. Elevated activity of the oxidative and non-oxidative pentose phosphate pathway in (pre)neoplastic lesions in rat liver. Int. J. Exp. Pathol. 2008, 89, 232–240. [Google Scholar] [CrossRef]
- Patra, K.C.; Hay, N. The pentose phosphate pathway and cancer. Trends Biochem. Sci. 2014, 39, 347–354. [Google Scholar] [CrossRef] [Green Version]
- Jin, L.; Zhou, Y. Crucial role of the pentose phosphate pathway in malignant tumors (review). Oncol. Lett. 2019, 17, 4213–4221. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Liu, X.; Cheng, Z.; Zhu, J.; Xu, L.; Wang, F.; Qi, W.; Yan, J.; Liu, N.; Sun, Z.; et al. Quantitative Analysis of Global Proteome and Lysine Acetylome Reveal the Differential Impacts of VPA and SAHA on HL60 Cells. Sci. Rep. 2016, 6, 19926. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Li, Z.; Wang, X.; Jiang, G.; Shan, C.; Liu, S. Metabolomics reveals the effect of valproic acid on MCF-7 and MDA-MB-231 cells. Xenobiotica 2019, 252–260. [Google Scholar] [CrossRef]
- Daher, B.; Vucetic, M.; Pouyssegur, J. Cysteine Depletion, a Key Action to Challenge Cancer Cells to Ferroptotic Cell Death. Front. Oncol. 2020, 10, 723. [Google Scholar] [CrossRef]
- Serpa, J. Cysteine as a Carbon Source, a Hot Spot in Cancer Cells Survival. Front. Oncol. 2020, 10, 947. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Kang, R.; Iii, H.J.Z.; Lotze, M.T. High-Mobility Group Box 1, Oxidative Stress, and Disease. Antioxidants Redox Signal. 2011, 14, 1315–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Contis-Montes de Oca, A.; Rodarte Valle, E.; Rosales Hernández, M.C.; Abarca Rojano, E.; Rojas Hernández, S.; Fragoso Vázquez, M.J.; Mendieta Wejebe, J.E.; Correa Basurto, A.M.; Vázquez Moctezuma, I.; Correa Basurto, J. N-(2’-Hydroxyphenyl)-2-propylpentanamide (OH-VPA), a histone deacetylase inhibitor, induces the release of nuclear HMGB1 and modifies ROS levels in HeLa cells. Oncotarget 2018, 9, 33368–33381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Ferroptosis: An Iron-Dependent Form of Nonapoptotic Cell Death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neagu, M.; Constantin, C.; Popescu, I.D.; Zipeto, D.; Tzanakakis, G.; Nikitovic, D.; Fenga, C.; Stratakis, C.A.; Spandidos, D.A.; Tsatsakis, A.M. Inflammation and metabolism in cancer cell—Mitochondria key player. Front. Oncol. 2019, 9, 348. [Google Scholar] [CrossRef] [Green Version]
- Keeley, T.S.; Yang, S.; Lau, E. The diverse contributions of fucose linkages in cancer. Cancers 2019, 11, 1241. [Google Scholar] [CrossRef] [Green Version]
- Russo, J.; Fernandez, S.V.; Russo, P.A.; Fernbaugh, R.; Sheriff, F.S.; Lareef, H.M.; Garber, J.; Russo, I.H. 17-Beta-Estradiol induces transformation and tumorigenesis in human breast epithelial cells. FASEB J. 2006, 20, 1622–1634. [Google Scholar] [CrossRef]
- Hardy, S.; El-Assaad, W.; Przybytkowski, E.; Joly, E.; Prentki, M.; Langelier, Y. Saturated fatty acid-induced apoptosis in MDA-MB-231 breast cancer cells. A role for cardiolipin. J. Biol. Chem. 2003, 278, 31861–31870. [Google Scholar] [CrossRef] [Green Version]
- Thomas, W.; Caiazza, F.; Harvey, B.J. Estrogen, phospholipase A and breast cancer. Front. Biosci. 2008, 13, 2604–2613. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Liu, Y.; Geng, C.; Qi, X.; Jiang, J. Estrogen receptor β inhibits estradiol-induced proliferation and migration of MCF-7 cells through regulation of mitofusin 2. Int. J. Oncol. 2013, 42, 1993–2000. [Google Scholar] [CrossRef]
- Wickramasinghe, N.S.M.D.; Jo, H.; McDonald, J.M.K.; Hardy, R.W. Stearate inhibition of breast cancer cell proliferation: A mechanism involving epidermal growth factor receptor and G-proteins. Am. J. Pathol. 1996, 148, 987–995. [Google Scholar] [PubMed]
- Evans, L.M.; Cowey, S.L.; Siegal, G.P.; Hardy, R.W. Stearate Preferentially Induces Apoptosis in Human Breast Cancer Cells. Nutr. Cancer 2009, 61, 746–753. [Google Scholar] [CrossRef] [PubMed]
- Aztopal, N.; Erkisa, M.; Erturk, E.; Ulukaya, E.; Tokullugil, A.H.; Ari, F. Valproic acid, a histone deacetylase inhibitor, induces apoptosis in breast cancer stem cells. Chem. Biol. Interact. 2018, 280, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Yaku, K.; Okabe, K.; Hikosaka, K.; Nakagawa, T. NAD metabolism in cancer therapeutics. Front. Oncol. 2018, 8, 622. [Google Scholar] [CrossRef] [PubMed]
- Navas, L.E.; Carnero, A. NAD+ metabolism, stemness, the immune response, and cancer. Signal Transduct. Target. Ther. 2021, 6, 2. [Google Scholar] [CrossRef]
- Salsaa, M.; Pereira, B.; Liu, J.; Yu, W.; Jadhav, S.; Hüttemann, M.; Greenberg, M.L. Valproate inhibits mitochondrial bioenergetics and increases glycolysis in Saccharomyces cerevisiae. Sci. Reports 2020 101 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Comşa, Ş.; Cîmpean, A.M.; Raica, M. The story of MCF-7 breast cancer cell line: 40 Years of experience in research. Anticancer Res. 2015, 35, 3147–3154. [Google Scholar]
- Fortunati, N.; Bertino, S.; Costantino, L.; Bosco, O.; Vercellinatto, I.; Catalano, M.G.; Boccuzzi, G. Valproic acid is a selective antiproliferative agent in estrogen-sensitive breast cancer cells. Cancer Lett. 2008, 259, 156–164. [Google Scholar] [CrossRef]
- Tautenhahn, R.; Patti, G.J.; Rinehart, D.; Siuzdak, G. XCMS Online: A web-based platform to process untargeted metabolomic data. Anal. Chem. 2013, 84, 5035–5039. [Google Scholar] [CrossRef] [Green Version]
- Guijas, C.; Montenegro-Burke, J.R.; Domingo-Almenara, X.; Palermo, A.; Warth, B.; Hermann, G.; Koellensperger, G.; Huan, T.; Uritboonthai, W.; Aisporna, A.E.; et al. METLIN: A Technology Platform for Identifying Knowns and Unknowns. Anal. Chem. 2018, 90, 3156–3164. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Metabolite | Dysregulation | Fold Change | p-Value | m/z | RT (min) |
|---|---|---|---|---|---|
| Glycocholate | Up | 2.7 | 2.3 × 10−4 | 488.3007 | 19.75 |
| 7α, 12α-dihydroxycholest-4-en-3-one | Down | 3.2 | 5.5 × 10−4 | 400.3111 | 23.27 |
| Calcitriol | Down | 3.2 | 5.5 × 10−4 | 400.3111 | 23.27 |
| L-cysteine | Down | 3.0 | 2.6 × 10−3 | 145.0173 | 2.32 |
| β-L-fucose 1-phosphate | Down | 3.5 | 8.7 × 10−3 | 262.0682 | 2.32 |
| D-glyceraldehyde 3-phosphate | Down | 2.4 | 1.3 × 10−2 | 188.0317 | 2.32 |
| 2-deoxy-α-D-ribose 1-phosphate | Down | 3.7 | 1.7 × 10−2 | 232.0577 | 2.32 |
| 17 β-estradiol | Down | 2.5 | 2.0 × 10−2 | 273.1849 | 21.81 |
| Nicotinamide | Down | 3.1 | 2.7 × 10−2 | 123.0555 | 3.36 |
| Stearate | Up | 1.7 | 3.8 × 10−2 | 267.2683 | 23.91 |
| D-sedoheptulose 7-phosphate | Down | 2.6 | 4.1 × 10−2 | 157.0177 | 2.32 |
| 17-hydro(p)docosahexaenoate | Down | 2.7 | 4.4 × 10−2 | 378.2652 | 18.81 |
| Pathway | Pathway Number of Metabolites Found/Total Number of Metabolites | Putative Metabolites Identity | p Value |
|---|---|---|---|
| 2′-deoxy-α-D-ribose 1-phosphate degradation | 2/2 | 2-deoxy-α-D-ribose 1-phosphate | 4.4 × 10−3 |
| D-glyceraldehyde 3-phosphate | |||
| Pentose phosphate pathway (nonoxidative branch) | 2/4 | D-sedoheptulose 7-phosphate | 8 × 10−3 |
| D-glyceraldehyde 3-phosphate | |||
| Bile acid biosynthesis, neutral pathway | 2/8 | 7α, 12α-dihydroxycholest-4-en-3-one | 2 × 10−2 |
| Glycocholate |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Estrada-Pérez, A.R.; Rosales-Hernández, M.C.; García-Vázquez, J.B.; Bakalara, N.; Fromager, B.; Correa-Basurto, J. Untargeted LC-MS/MS Metabolomics Study on the MCF-7 Cell Line in the Presence of Valproic Acid. Int. J. Mol. Sci. 2022, 23, 2645. https://doi.org/10.3390/ijms23052645
Estrada-Pérez AR, Rosales-Hernández MC, García-Vázquez JB, Bakalara N, Fromager B, Correa-Basurto J. Untargeted LC-MS/MS Metabolomics Study on the MCF-7 Cell Line in the Presence of Valproic Acid. International Journal of Molecular Sciences. 2022; 23(5):2645. https://doi.org/10.3390/ijms23052645
Chicago/Turabian StyleEstrada-Pérez, Alan Rubén, Martha Cecilia Rosales-Hernández, Juan Benjamín García-Vázquez, Norbert Bakalara, Benedicte Fromager, and José Correa-Basurto. 2022. "Untargeted LC-MS/MS Metabolomics Study on the MCF-7 Cell Line in the Presence of Valproic Acid" International Journal of Molecular Sciences 23, no. 5: 2645. https://doi.org/10.3390/ijms23052645