
Possible Action of Olaparib for Preventing Invasion of Oral Squamous Cell Carcinoma In Vitro and In Vivo

 ,
,
Abstract
:1. Introduction
2. Results
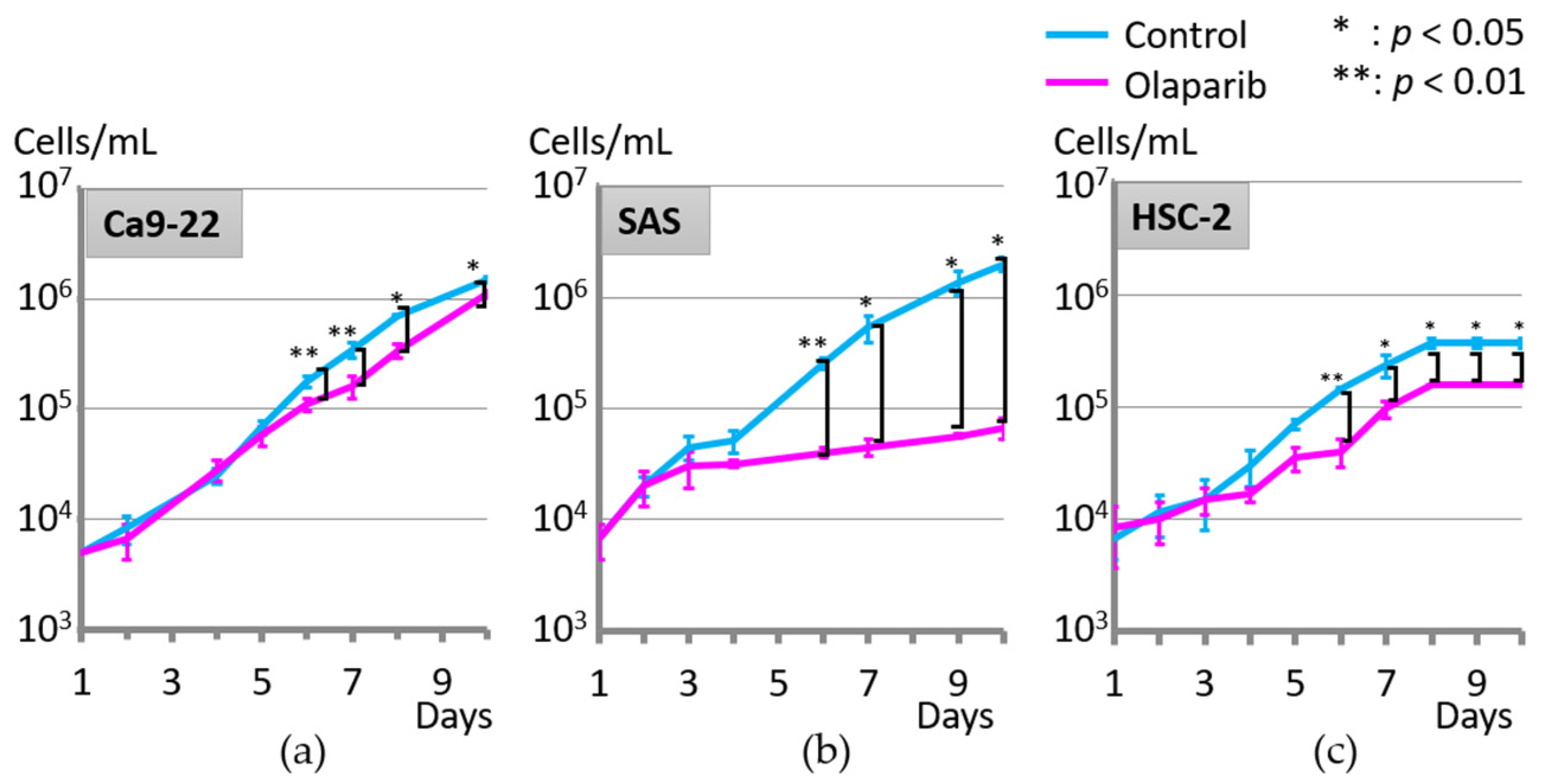
2.1. Effect of Olaparib on Cell Proliferation
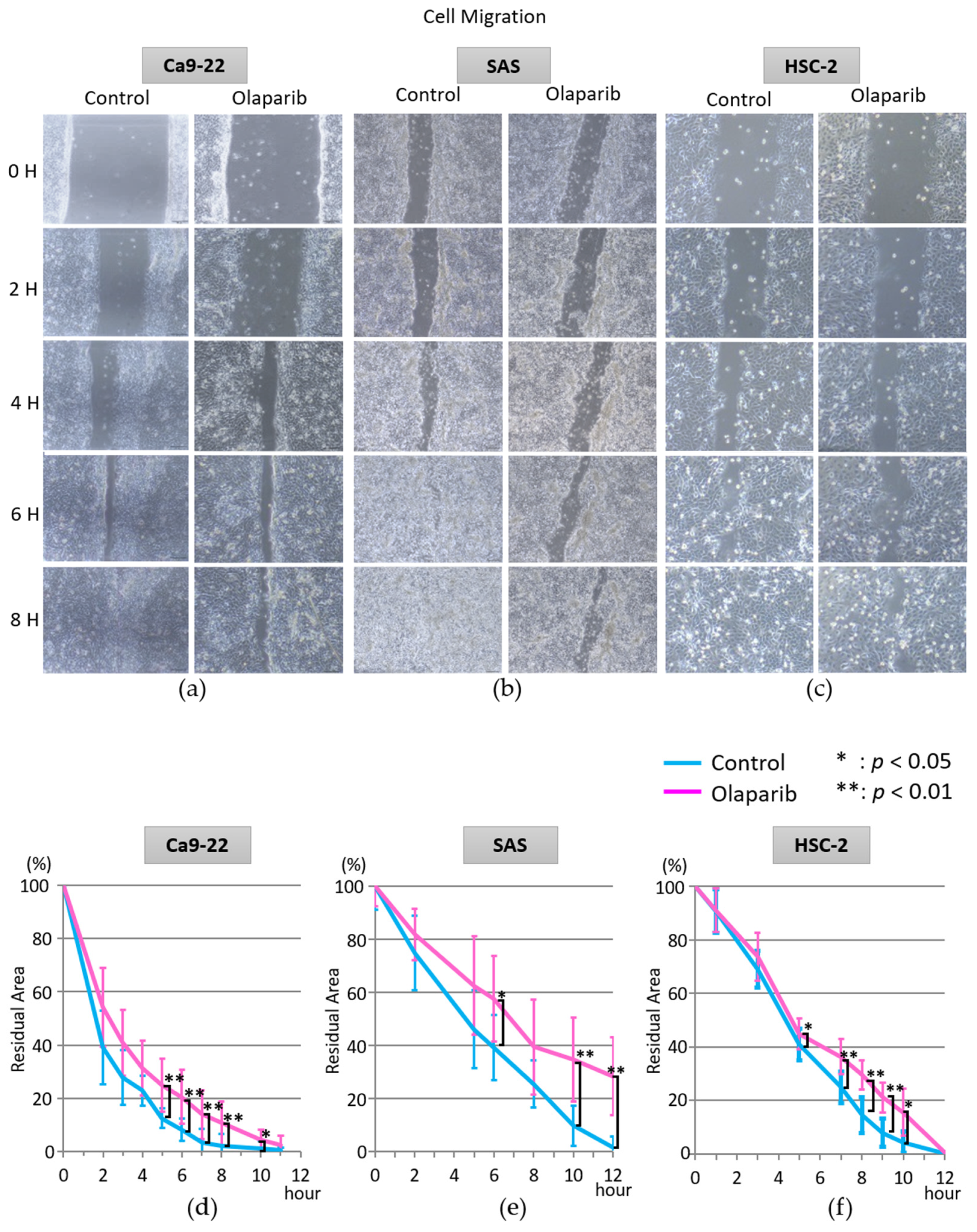
2.2. Effect of Olaparib on Cell Migration
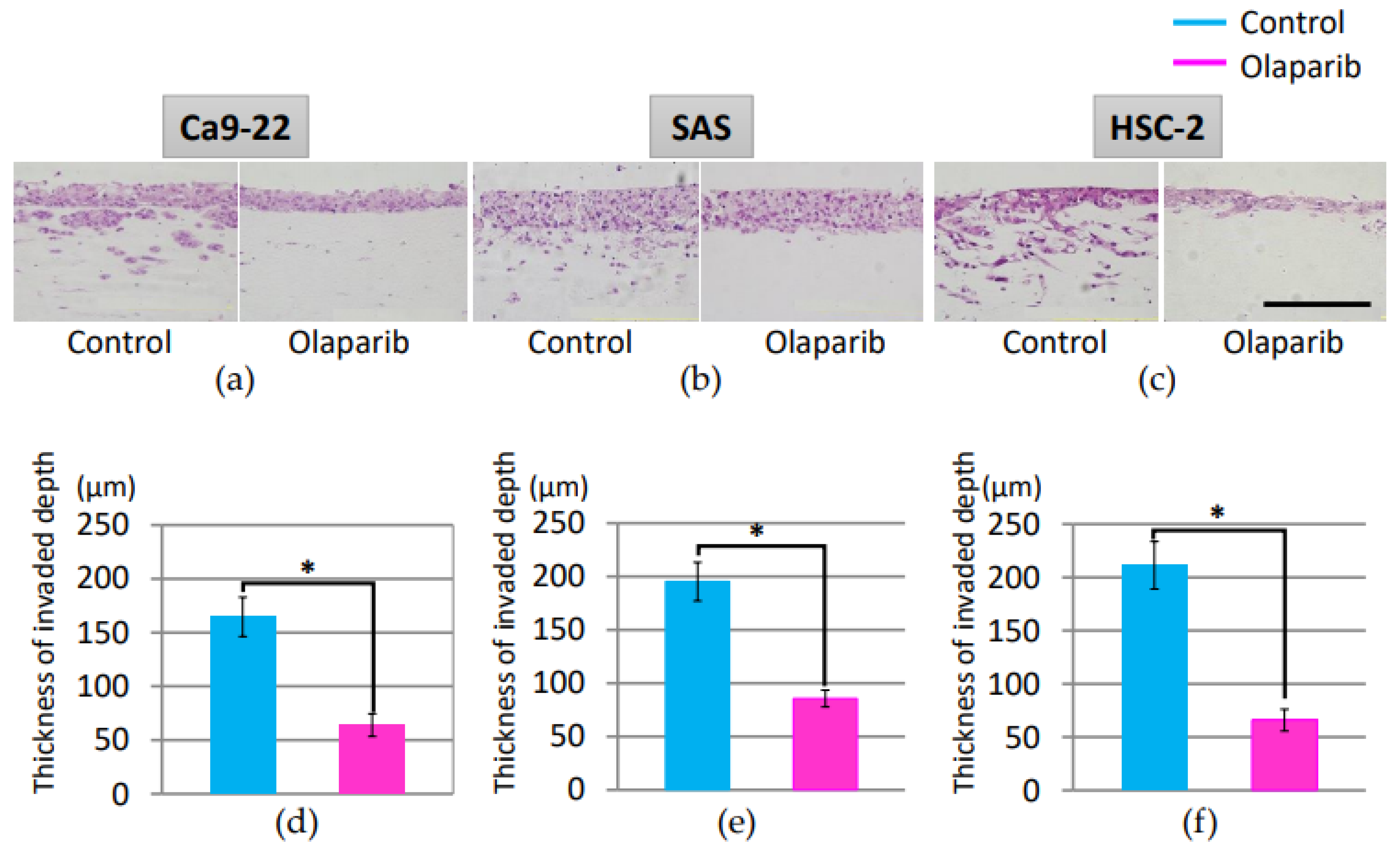
2.3. Effect of Olaparib on Cell Invasion
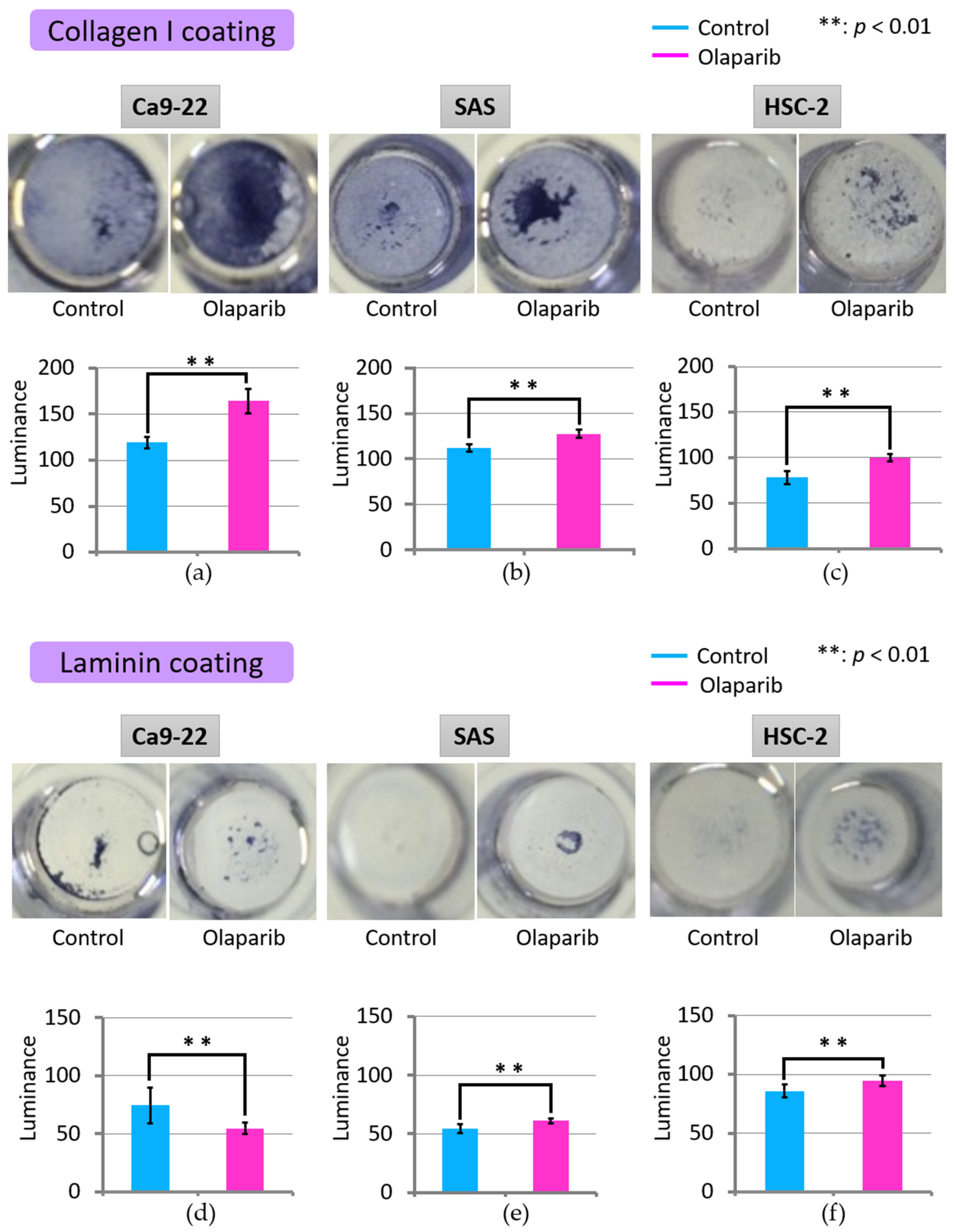
2.4. Effect of Olaparib on Cell Adhesion
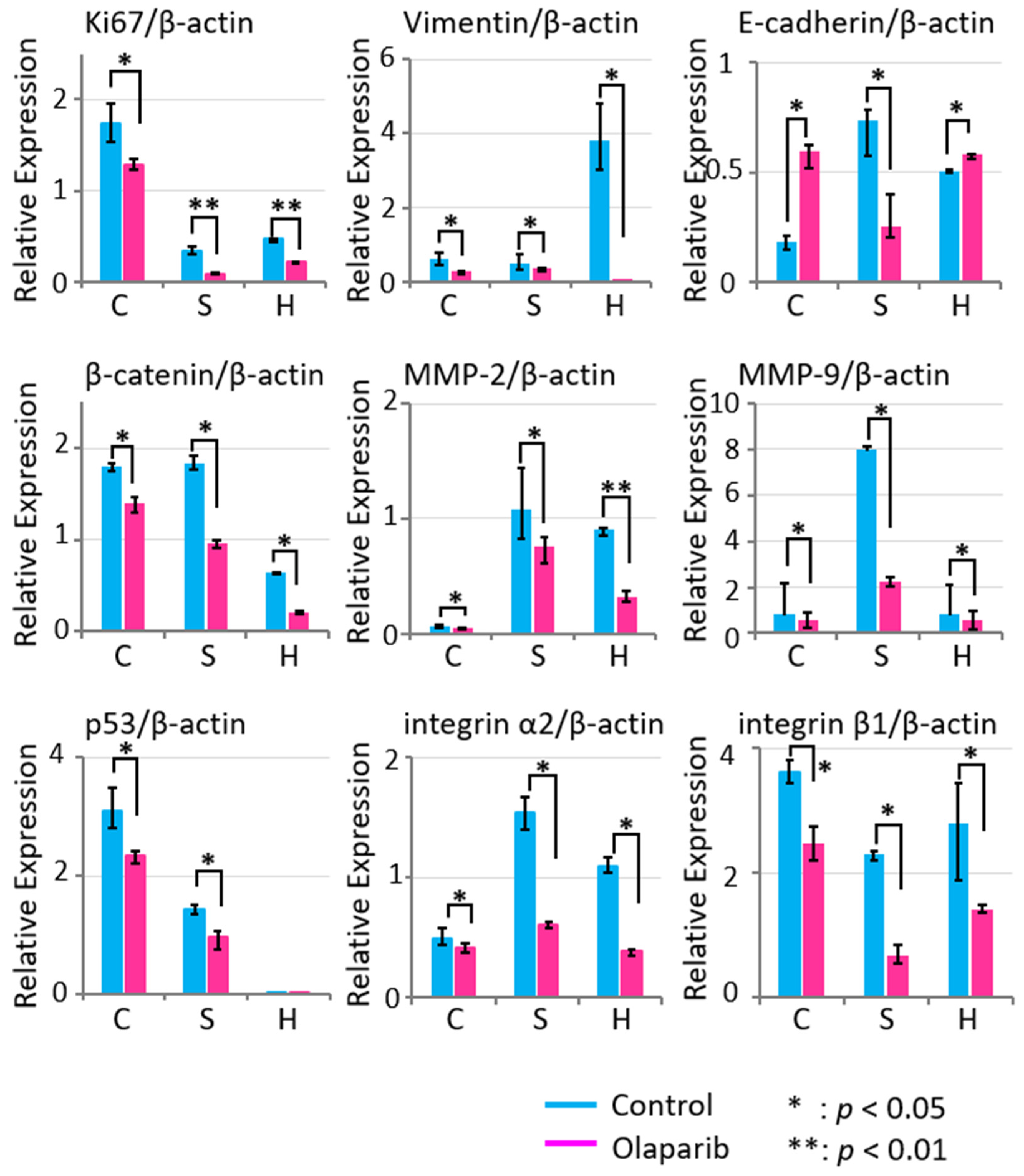
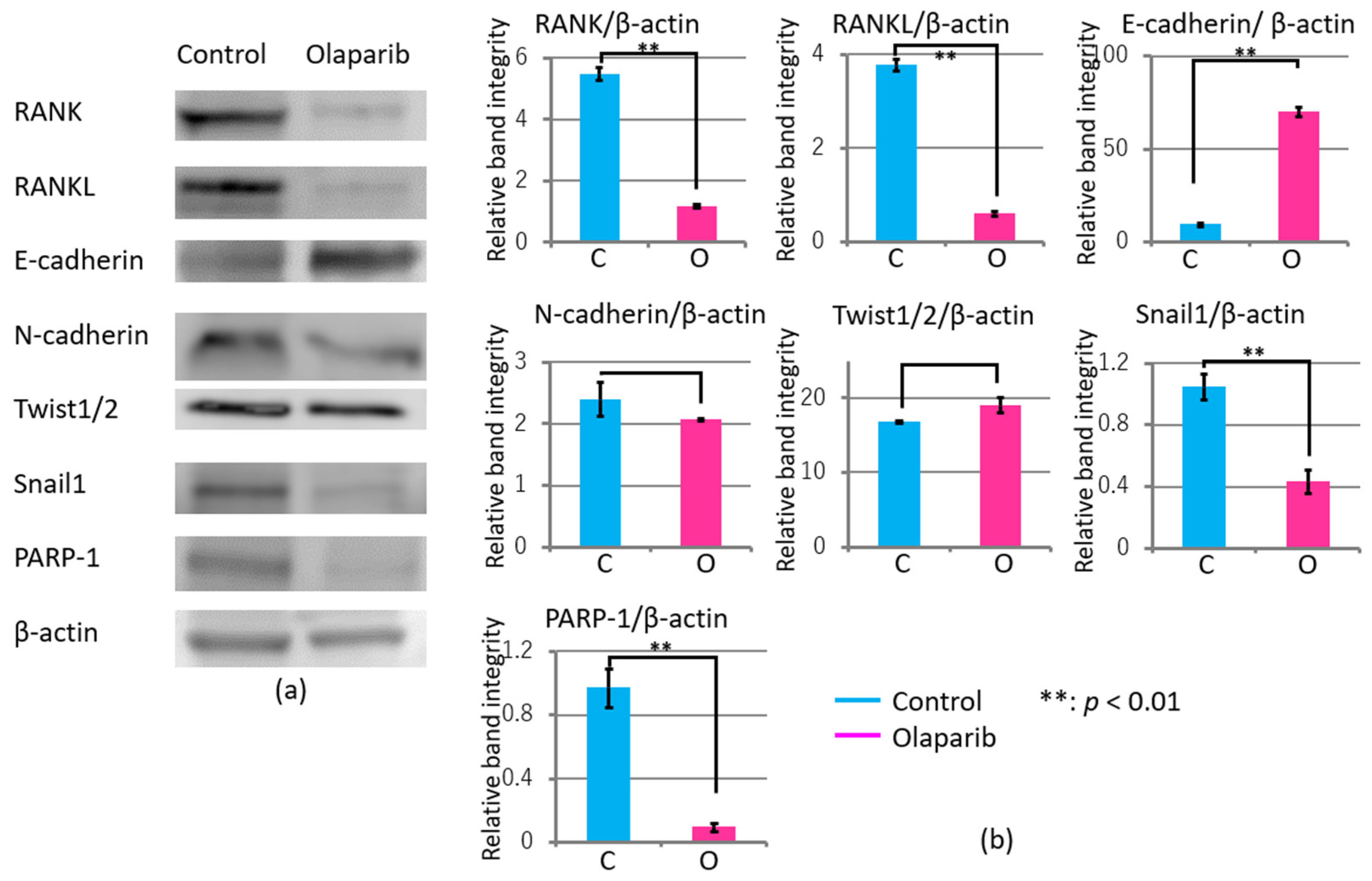
2.5. Effects of Olaparib on Markers Related to Migration, Invasion, and Adhesion
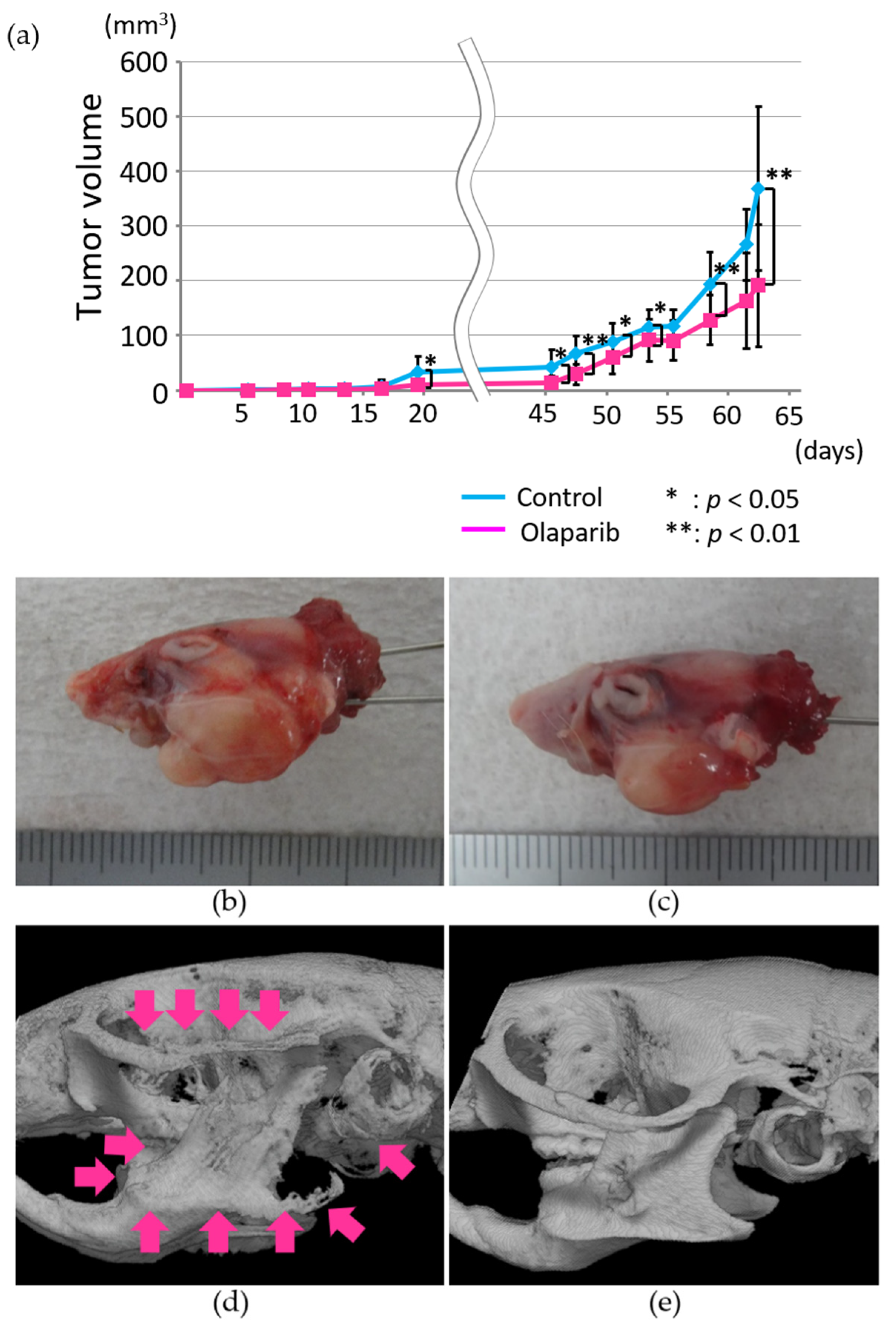
2.6. Radiographic Analysis of the In Vivo Effects of Olaparib on Xenograft Tumor Growth and Bone Destruction by Tumor Invasion
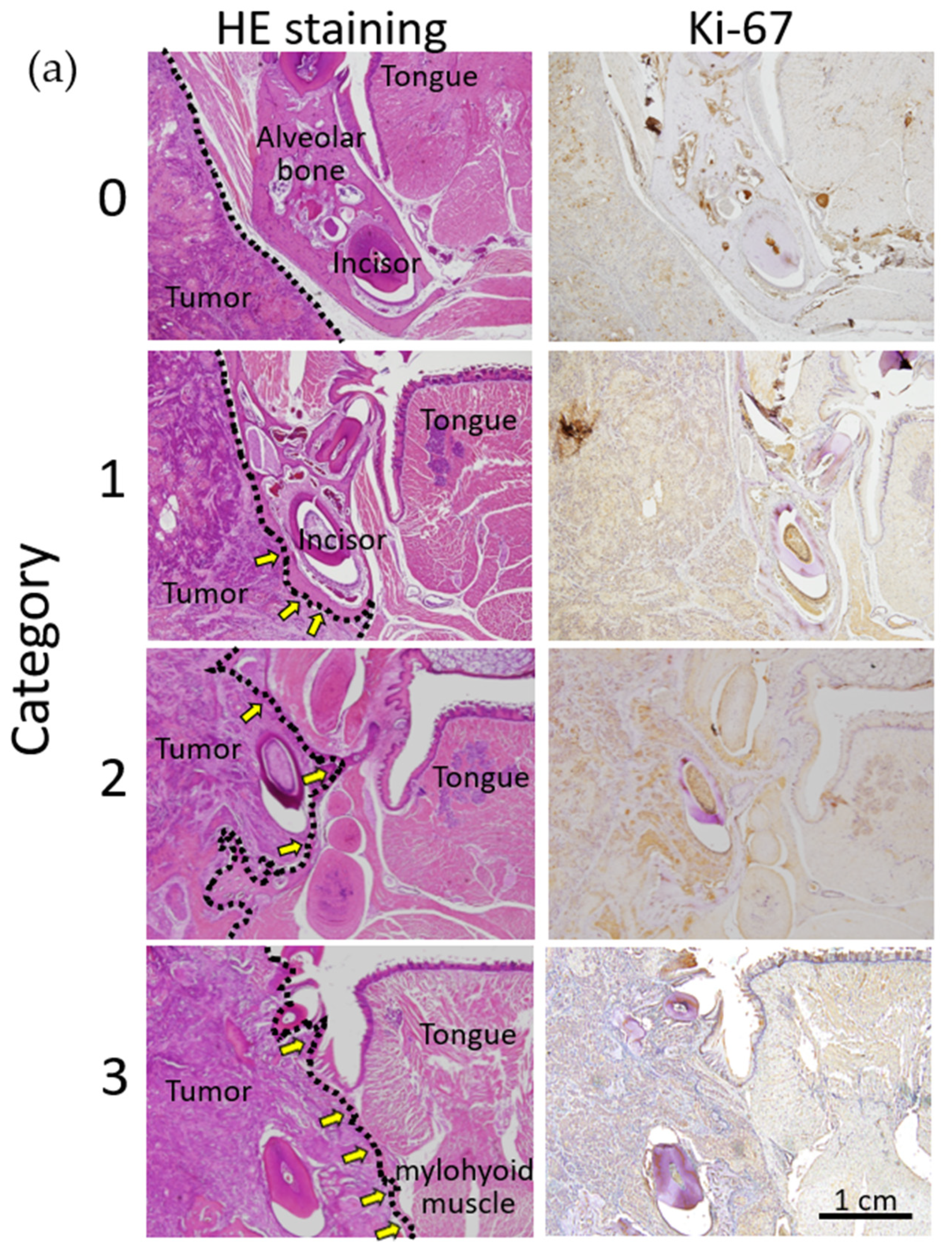
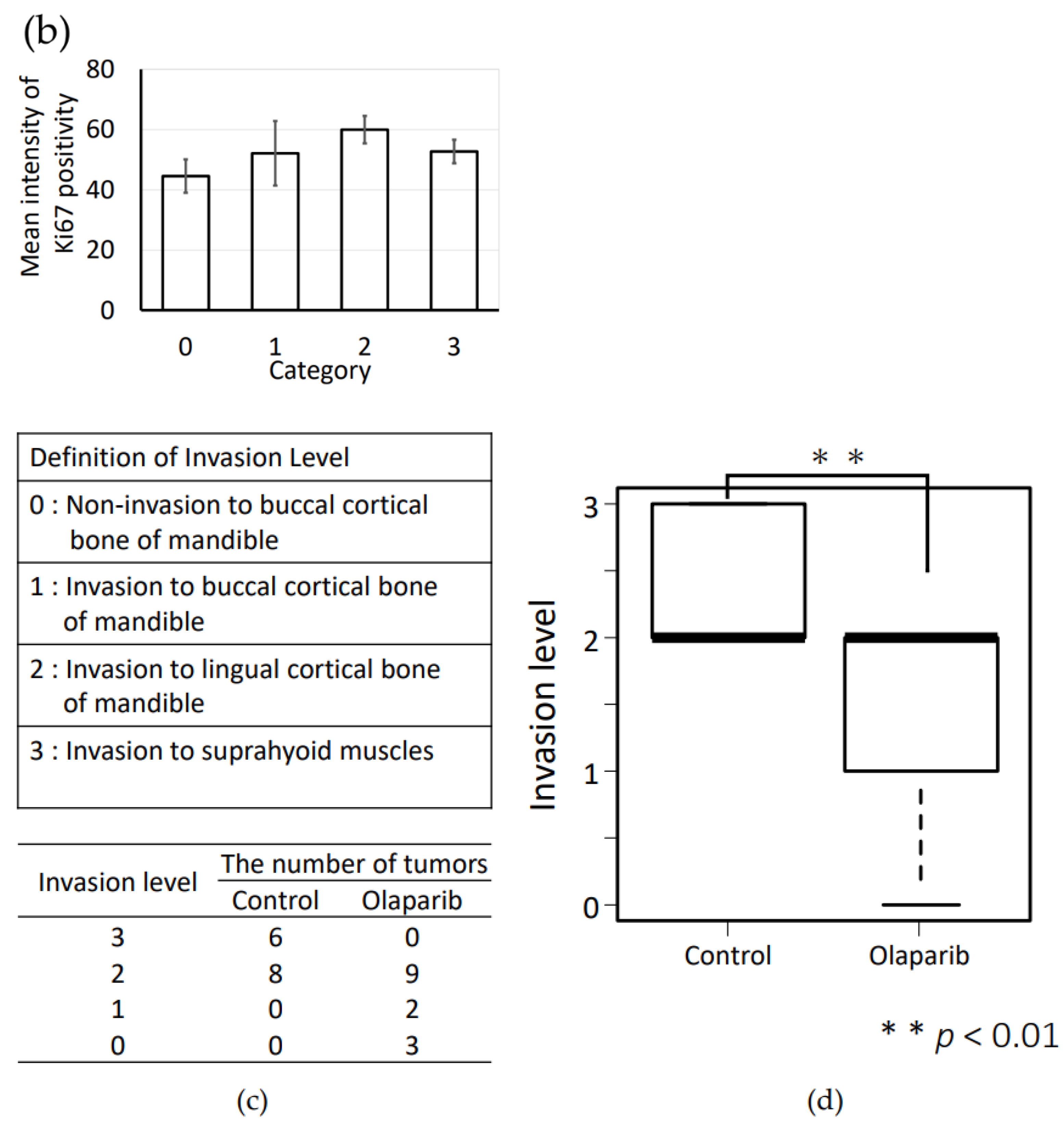
2.7. Histopathological Analysis of the In Vivo Effects of Olaparib on Xenograft Tumor Growth and Bone Destruction by Tumor Invasion
2.8. Olaparib Affected the Expression of Proteins Involved in Invasion and Adhesion in Tumor Tissues
3. Discussion
4. Materials and Methods
4.1. Cell Culture of Cell Lines Derived from Oral Carcinoma
4.2. Proliferation Assay
4.3. Wound Healing Assay
4.4. Invasion Assay
4.5. Adhesion Assay
4.6. Total RNA Extraction and Real-Time PCR
4.7. Animal Models
4.8. Experimental Animal Protocol
4.9. Radiological Analysis by Micro-CT
4.10. Histopathological Analysis
4.11. Immunohistochemistry
4.12. Quantification of the Intensity of Immunohistochemical Positivity
4.13. Western Blot Analysis
4.14. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Oral and Oropharyngeal Cancer: Types of Treatment|Cancer.Net. Available online: https://www.cancer.net/cancer-types/oral-and-oropharyngeal-cancer/types-treatment (accessed on 5 May 2021).
- Hashibe, M.; Brennan, P.; Chuang, S.; Boccia, S.; Castellsague, X.; Chen, C.; Curado, M.P.; Maso, L.D.; Daudt, A.W.; Fernandez, L.; et al. Interaction between tobacco and alcohol use and the risk of head and neck cancer: Pooled analysis in the INHANCE consortium. Cancer Epidemiol. Biomark. Prev. 2009, 18, 541–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diana, G.; Corica, C. Human Papilloma Virus vaccine and prevention of head and neck cancer, what is the current evidence? Oral Oncol. 2021, 115, 105168. [Google Scholar] [CrossRef] [PubMed]
- Alsahafi, E.; Begg, K.; Amelio, I.; Raulf, N.; Lucarelli, P.; Sauter, T.; Tavassoli, M. Clinical update on head and neck cancer: Molecular biology and ongoing challenges. Cell Death Dis. 2019, 10, 540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haddad, R.I.; Shin, D.M. Recent advances in head and neck cancer reconstruction. N. Engl. J. Med. 2008, 359, 1143–1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fasano, M.; Della Corte, C.M.; Viscardi, G.; Di Liello, R.; Paragliola, F.; Sparano, F.; Iacovino, M.L.; Castrichino, A.; Doria, F.; Sica, A.; et al. Head and neck cancer: The role of anti-EGFR agents in the era of immunotherapy. Ther. Adv. Med. Oncol. 2021, 13, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Semrau, S.; Gostian, A.-O.; Traxdorf, M.; Eckstein, M.; Rutzner, S.; Von Der Grün, J.; Illmer, T.; Hautmann, M.; Klautke, G.; Laban, S.; et al. Implementation of Double Immune Checkpoint Blockade Increases Response Rate to Induction Chemotherapy in Head and Neck Cancer. Cancers 2021, 13, 1959. [Google Scholar] [CrossRef] [PubMed]
- Oualla, K.; Castelo Branco, L.; Nouiyakh, L.; Amaadour, L.; Benbrahim, Z.; Arifi, S.; Mellas, N. Therapeutic Approaches With Immune Checkpoint Inhibitors in Head and Neck Cancers and the Role of PD-L1 as a Biomarker. Cancer Control 2021, 28, 1–4. [Google Scholar] [CrossRef]
- Yamada, H.; Nakaoka, K.; Horiuchi, T.; Kumagai, K.; Ikawa, T.; Shigeta, Y.; Imamura, E.; Iino, M.; Ogawa, T.; Hamada, Y. Mandibular reconstruction using custom-made titanium mesh tray and particulate cancellous bone and marrow harvested from bilateral posterior ilia. J. Plast. Surg. Hand Surg. 2014, 48, 183–190. [Google Scholar] [CrossRef]
- Chalmers, R.; Schlabe, J.; Yeung, E.; Kerawala, C.; Cascarini, L.; Paleri, V. Robot-Assisted Reconstruction in Head and Neck Surgical Oncology: The Evolving Role of the Reconstructive Microsurgeon. ORL J. Otorhinolaryngol. Relat Spec. 2018, 80, 178–185. [Google Scholar] [CrossRef]
- Christianto, S.; Lau, A.; Li, K.Y.; Yang, W.F.; Su, Y.X. One versus two venous anastomoses in microsurgical head and neck reconstruction: A cumulative meta-analysis. Int. J. Oral Maxillofac. Surg. 2018, 47, 585–594. [Google Scholar] [CrossRef]
- Manjunath, M.; Choudhary, B. Triple-negative breast cancer: A run-through of features, classification and current therapies (Review). Oncol. Lett. 2021, 22, 512. [Google Scholar] [CrossRef] [PubMed]
- Kvízová, J.; Pavlíčková, V.; Kmoníčková, E.; Ruml, T.; Rimpelová, S. Quo Vadis Advanced Prostate Cancer Therapy? Novel Treatment Perspectives and Possible Future Directions. Molecules 2021, 26, 2228. [Google Scholar] [CrossRef] [PubMed]
- Dantzer, F.; De La Rubia, G.; Ménissier-De Murcia, J.; Hostomsky, Z.; De Murcia, G.; Schreiber, V. Base excision repair is impaired in mammalian cells lacking poly(ADP- ribose) polymerase-1. Biochemistry 2000, 39, 7559–7569. [Google Scholar] [CrossRef] [PubMed]
- Cerrato, A.; Morra, F.; Celetti, A. Use of poly ADP-ribose polymerase [PARP] inhibitors in cancer cells bearing DDR defects: The rationale for their inclusion in the clinic. J. Exp. Clin. Cancer Res. 2016, 35, 179. [Google Scholar] [CrossRef] [Green Version]
- McCabe, N.; Turner, N.C.; Lord, C.J.; Kluzek, K.; Białkowska, A.; Swift, S.; Giavara, S.; O’Connor, M.J.; Tutt, A.N.; Zdzienicka, M.Z.; et al. Deficiency in the repair of DNA damage by homologous recombination and sensitivity to poly(ADP-ribose) polymerase inhibition. Cancer Res. 2006, 66, 8109–8115. [Google Scholar] [CrossRef] [Green Version]
- Malanga, M.; Pleschke, J.M.; Kleczkowska, H.E.; Althaus, F.R. Poly(ADP-ribose) binds to specific domains of p53 and alters its DNA binding functions. J. Biol. Chem. 1998, 273, 11839–11843. [Google Scholar] [CrossRef] [Green Version]
- Pleschke, J.M.; Kleczkowska, H.E.; Strohm, M.; Althaus, F.R. Poly(ADP-ribose) binds to specific domains in DNA damage checkpoint proteins. J. Biol. Chem. 2000, 275, 40974–40980. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Luo, W.; Wang, Y. Emerging role of PARP-1 and PARthanatos in ischemic stroke. J. Neurochem. 2021, 160, 74–87. [Google Scholar] [CrossRef]
- Benedetti, F.; Cocchi, F.; Latinovic, O.S.; Curreli, S.; Krishnan, S.; Munawwar, A.; Gallo, R.C.; Zella, D. Role of Mycoplasma Chaperone DnaK in Cellular Transformation. Int. J. Mol. Sci. 2020, 21, 1311. [Google Scholar] [CrossRef] [Green Version]
- Seimiya, H. The telomeric PARP, tankyrases, as targets for cancer therapy. Br. J. Cancer 2006, 94, 341–345. [Google Scholar] [CrossRef]
- Rosado, M.M.; Bennici, E.; Novelli, F.; Pioli, C. Beyond DNA repair, the immunological role of PARP-1 and its siblings. Immunology 2013, 139, 428–437. [Google Scholar] [CrossRef]
- Masutani, M.; Fujimori, H. Poly(ADP-ribosyl)ation in carcinogenesis. Mol. Aspects Med. 2013, 34, 1202–1216. [Google Scholar] [CrossRef] [PubMed]
- Yasukawa, M.; Fujihara, H.; Fujimori, H.; Kawaguchi, K.; Yamada, H.; Nakayama, R.; Yamamoto, N.; Kishi, Y.; Hamada, Y.; Masutani, M. Synergetic Effects of PARP Inhibitor AZD2281 and Cisplatin in Oral Squamous Cell Carcinoma in Vitro and in Vivo. Int. J. Mol. Sci. 2016, 17, 272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruggeri, E.M.; Carlini, P.; Pollera, C.F.; De Marco, S.; Ruscito, P.; Pinnarò, P.; Nardi, M.; Giannarelli, D.; Cognetti, F. Long-term survival in locally advanced oral cavity cancer: An analysis of patients treated with neoadjuvant cisplatin-based chemotherapy followed by surgery. Head Neck 2005, 27, 452–458. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Li, R. Contemporary Treatment of Locally Advanced Oral Cancer. Curr. Treat. Options Oncol. 2019, 20, 32. [Google Scholar] [CrossRef]
- Omura, K. Current status of oral cancer treatment strategies: Surgical treatments for oral squamous cell carcinoma. Int. J. Clin. Oncol. 2014, 19, 423–430. [Google Scholar] [CrossRef]
- Lönn, P.; van der Heide, L.P.; Dahl, M.; Hellman, U.; Heldin, C.H.; Moustakas, A. PARP-1 attenuates Smad-mediated transcription. Mol. Cell 2010, 40, 521–532. [Google Scholar] [CrossRef]
- Rebecca, V.W.; Somasundaram, R.; Herlyn, M. Pre-clinical modeling of cutaneous melanoma. Nat. Commun. 2020, 11, 2858. [Google Scholar] [CrossRef]
- Kim, K.M.; Im, A.-R.; Kim, S.H.; Hyun, J.W.; Chae, S. Timosaponin AIII inhibits melanoma cell migration by suppressing COX-2 and in vivo tumor metastasis. Cancer Sci. 2016, 107, 181–188. [Google Scholar] [CrossRef] [Green Version]
- Calabrò, M.L.; Lazzari, N.; Rigotto, G.; Tonello, M.; Sommariva, A. Role of Epithelial-Mesenchymal Plasticity in Pseudomyxoma Peritonei: Implications for Locoregional Treatments. Int. J. Mol. Sci. 2020, 21, 9120. [Google Scholar] [CrossRef]
- Kumar, M.; Jaiswal, R.K.; Yadava, P.K.; Singh, R.P. An assessment of poly (ADP-ribose) polymerase-1 role in normal and cancer cells. BioFactors 2020, 46, 894–905. [Google Scholar] [CrossRef] [PubMed]
- Kolijn, K.; Verhoef, E.I.; van Leenders, G.J.L.H. Morphological and immunohistochemical identification of epithelial-to-mesenchymal transition in clinical prostate cancer. Oncotarget 2015, 6, 24488–24498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyuno, D.; Yamaguchi, H.; Ito, T.; Kono, T.; Kimura, Y.; Imamura, M.; Konno, T.; Hirata, K.; Sawada, N.; Kojima, T. Targeting tight junctions during epithelial to mesenchymal transition in human pancreatic cancer. World J. Gastroenterol. 2014, 20, 10813–10824. [Google Scholar] [CrossRef] [PubMed]
- Lamouille, S.; Xu, J.; Derynck, R. Molecular mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell Biol 2014, 15, 178–196. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, M.I.; González-Flores, A.; Dantzer, F.; Collard, J.; de Herreros, A.G.; Oliver, F.J. Poly(ADP-ribose)-dependent regulation of Snail1 protein stability. Oncogene 2011, 30, 4365–4372. [Google Scholar] [CrossRef] [Green Version]
- Berkholtz, C.B.; Lai, B.E.; WoodruV, T.K.; Shea, L.D. Distribution of extracellular matrix proteins type I collagen, type IV collagen, fibronectin, and laminin in mouse folliculogenesis. Histochem. Cell Biol. 2006, 126, 583–592. [Google Scholar] [CrossRef] [Green Version]
- Mittag, F.; Falkenberg, E.-M.; Janczyk, A.; Götze, M.; Felka, T.; Aicher, W.K.; Kluba, T. Laminin-5 and type I collagen promote adhesion and osteogenic differentiation of animal serum-free expanded human mesenchymal stromal cells. Orthop. Rev. 2012, 4, e36. [Google Scholar] [CrossRef]
- Pearson, G.W. Control of invasion by epithelial-to-mesenchymal transition programs during metastasis. J. Clin. Med. 2019, 8, 646. [Google Scholar] [CrossRef] [Green Version]
- Li, C.H.; Hsu, T.I.; Chang, Y.C.; Chan, M.H.; Lu, P.J.; Hsiao, M. Stationed or relocating: The seesawing emt/met determinants from embryonic development to cancer metastasis. Biomedicines 2021, 9, 1265. [Google Scholar] [CrossRef]
- Zheng, K.; He, Z.; Kitazato, K.; Wang, Y. Selective Autophagy Regulates Cell Cycle in Cancer Therapy. Theranostics 2019, 9, 104–125. [Google Scholar] [CrossRef]
- Chen, T.; Wang, W.; Li, J.R.; Xu, H.Z.; Peng, Y.C.; Fan, L.F.; Yan, F.; Gu, C.; Wang, L.; Chen, G. PARP inhibition attenuates early brain injury through NF-κB/MMP-9 pathway in a rat model of subarachnoid hemorrhage. Brain Res. 2016, 1644, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, P.; Dey, P.; De, D.; Ghosh, U. Gamma ray-induced in vitro cell migration via EGFR/ERK/Akt/p38 activation is prevented by olaparib pretreatment. Int. J. Radiat. Biol. 2020, 96, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, P.; Dey, P.; Ghosh, S.; Sarma, A.; Ghosh, U. Reduction of metastatic potential by inhibiting EGFR/Akt/p38/ERK signaling pathway and epithelial-mesenchymal transition after carbon ion exposure is potentiated by PARP-1 inhibition in non-small-cell lung cancer. BMC Cancer 2019, 19, 829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheli, Y.; Williams, S.A.; Ballotti, R.; Nugent, D.J.; Kunicki, T.J. Enhanced binding of poly(ADP-ribose)polymerase-1 and Ku80/70 to the ITGA2 promoter via an extended cytosine-adenosine repeat. PLoS ONE 2010, 5, e8743. [Google Scholar] [CrossRef]
- Tao, X.; Chen, X.; Hao, S.; Hou, Z.; Lu, T.; Sun, M.; Liu, B. Protective actions of PJ34, a poly(ADP-ribose)polymerase inhibitor, on the blood-brain barrier after traumatic brain injury in mice. Neuroscience 2015, 291, 26–36. [Google Scholar] [CrossRef]
- Rom, S.; Zuluaga-Ramirez, V.; Reichenbach, N.L.; Dykstra, H.; Gajghate, S.; Pacher, P.; Persidsky, Y. PARP inhibition in leukocytes diminishes inflammation via effects on integrins/ cytoskeleton and protects the blood-brain barrier. J. Neuroinflammation 2016, 13, 254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; López-Ozuna, V.M.; Baloch, T.; Bithras, J.; Amin, O.; Kessous, R.; Kogan, L.; Laskov, I.; Yasmeen, A. Biguanides in combination with olaparib limits tumorigenesis of drug-resistant ovarian cancer cells through inhibition of Snail. Cancer Med. 2020, 9, 1307–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pu, H.; Horbinski, C.; Hensley, P.J.; Matuszak, E.A.; Atkinson, T.; Kyprianou, N. PARP-1 regulates epithelial-mesenchymal transition (EMT) in prostate tumorigenesis. Carcinogenesis 2014, 35, 2592–2601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okuyama, K.; Suzuki, K.; Yanamoto, S.; Naruse, T.; Tsuchihashi, H.; Yamashita, S.; Umeda, M. Anaplastic transition within the cancer microenvironment in early-stage oral tongue squamous cell carcinoma is associated with local recurrence. Int. J. Oncol. 2018, 53, 1713–1720. [Google Scholar] [CrossRef] [Green Version]
- Zuo, H.; Yang, D.; Yang, Q.; Tang, H.; Fu, Y.-X.; Wan, Y. Differential regulation of breast cancer bone metastasis by PARP1 and PARP2. Nat. Commun. 2020, 11, 1578. [Google Scholar] [CrossRef] [Green Version]
- Beranger, G.E.; Momier, D.; Rochet, N.; Carle, G.F.; Scimeca, J.-C. Poly(adp-ribose) polymerase-1 regulates Tracp gene promoter activity during RANKL-induced osteoclastogenesis. J. Bone Miner. Res. 2008, 23, 564–571. [Google Scholar] [CrossRef] [PubMed]
- Kishi, Y.; Fujihara, H.; Kawaguchi, K.; Yamada, H.; Nakayama, R.; Yamamoto, N.; Fujihara, Y.; Hamada, Y.; Satomura, K.; Masutani, M. PARP Inhibitor PJ34 Suppresses Osteogenic Differentiation in Mouse Mesenchymal Stem Cells by Modulating BMP-2 Signaling Pathway. Int. J. Mol. Sci. 2015, 16, 24820–24838. [Google Scholar] [CrossRef] [PubMed]
- Almasoud, N.; Binhamdan, S.; Younis, G.; Alaskar, H.; Alotaibi, A.; Manikandan, M.; Alfayez, M.; Kassem, M.; AlMuraikhi, N. Tankyrase inhibitor XAV-939 enhances osteoblastogenesis and mineralization of human skeletal (mesenchymal) stem cells. Sci. Rep. 2020, 10, 16746. [Google Scholar] [CrossRef] [PubMed]
- Weston, V.J.; Oldreive, C.E.; Skowronska, A.; Oscier, D.G.; Pratt, G.; Dyer, M.J.S.; Smith, G.; Powell, J.E.; Rudzki, Z.; Kearns, P.; et al. The PARP inhibitor olaparib induces significant killing of ATM-deficientlymphoid tumor cells in vitro and in vivo. Blood 2010, 116, 4578–4587. [Google Scholar] [CrossRef] [Green Version]
- Chen, I.H.; Chang, F.R.; Wu, Y.C.; Kung, P.H.; Wu, C.C. 3,4-Methylenedioxy-β-nitrostyrene inhibits adhesion and migration of human triple-negative breast cancer cells by suppressing β1 integrin function and surface protein disulfide isomerase. Biochimie 2015, 110, 81–92. [Google Scholar] [CrossRef]
- He, H.; Zheng, L.; Sun, Y.P.; Zhang, G.W.; Yue, Z.G. Steroidal saponins from Paris polyphylla suppress adhesion, migration and invasion of human lung cancer A549 cells via down-regulating MMP-2 and MMP-9. Asian Pac. J. Cancer Prev. 2014, 15, 10911–10916. [Google Scholar] [CrossRef]
- Taguchi, S.; Kawachi, Y.; Ishitsuka, Y.; Fujisawa, Y.; Furuta, J.; Nakamura, Y.; Xu, X.; Ikebe, D.; Kato, M.; Otsuka, F. Overexpression of the transcription factor yin-yang-1 suppresses differentiation of HaCaT cells in three-dimensional cell culture. J. Investig. Dermatol. 2011, 131, 37–45. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, N.; Kawaguchi, K.; Fujihara, H.; Hasebe, M.; Kishi, Y.; Yasukawa, M.; Kumagai, K.; Hamada, Y. Detection accuracy for epithelial dysplasia using an objective autofluorescence visualization method based on the luminance ratio. Int. J. Oral Sci. 2017, 9, e2. [Google Scholar] [CrossRef] [Green Version]
- Kirihara, Y.; Takechi, M.; Kurosaki, K.; Kobayashi, Y.; Kurosawa, T. Anesthetic effects of a mixture of medetomidine, midazolam and butorphanol in two strains of mice. Exp. Anim. 2013, 62, 173–180. [Google Scholar] [CrossRef] [Green Version]
- Kawai, S.; Takagi, Y.; Kaneko, S.; Kurosawa, T. Effect of three types of mixed anesthetic agents alternate to ketamine in mice. Exp. Anim. 2011, 60, 481–487. [Google Scholar] [CrossRef] [Green Version]
- Kelavkar, U.P.; Hutzley, J.; McHugh, K.; Allen, K.G.D.; Parwani, A. Prostate tumor growth can be modulated by dietarily targeting the 15-lipoxygenase-1 and cyclooxygenase-2 enzymes. Neoplasia 2009, 11, 692–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Length | Forward Sequence (5’ > 3’) | Reverse Sequence (5’ > 3’) |
|---|---|---|---|
| Ki67 | 129 | ATCGTCCCAGGTGGAAGAGTT | ATAGTAACCAGGCGTCTCGTGG |
| vimentin | 72 | AGCCGAAAACACCCTGCAAT | CGTTCAAGGTCAAGACGTGC |
| E-cadherin | 196 | GGTGCTCTTCCAGGAACCTC | GGAAACTCTCTCGGTCCAGC |
| β-catenin | 116 | GAGTGCTGAAGGTGCTATCTGTCTG | GTTCTGAACAAGACGTTGACTTGGA |
| MMP2 | 266 | GTGTTCTTTGCAGGGAATGAAT | ACGACGGCATCCAGGTTATC |
| MMP9 | 80 | CGCCCATTTCGACGATGAC | CGCCATCTGCGTTTCCAA |
| p53 | 200 | CTACAAGCAGTCACAGCAC | AGTCAGAGCCAACCTCAG |
| Integrin β1 | 185 | CAAGCAGGGCCAAATTGTGG | CCTTTGCTACGGTTGGTTACATT |
| Integrin α2 | 224 | CTGGAGTGGCTTTCCTGAG | ACTG ATTCCCACATTGCTG |
| β-actin | 262 | GGCTGTATTCCCCTCCATCG | GTTGGCCTTAGGGTTCAGGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakamura, N.; Fujihara, H.; Kawaguchi, K.; Yamada, H.; Nakayama, R.; Yasukawa, M.; Kishi, Y.; Hamada, Y.; Masutani, M. Possible Action of Olaparib for Preventing Invasion of Oral Squamous Cell Carcinoma In Vitro and In Vivo. Int. J. Mol. Sci. 2022, 23, 2527. https://doi.org/10.3390/ijms23052527
Nakamura N, Fujihara H, Kawaguchi K, Yamada H, Nakayama R, Yasukawa M, Kishi Y, Hamada Y, Masutani M. Possible Action of Olaparib for Preventing Invasion of Oral Squamous Cell Carcinoma In Vitro and In Vivo. International Journal of Molecular Sciences. 2022; 23(5):2527. https://doi.org/10.3390/ijms23052527
Chicago/Turabian StyleNakamura, Nanami, Hisako Fujihara, Koji Kawaguchi, Hiroyuki Yamada, Ryoko Nakayama, Masaaki Yasukawa, Yuta Kishi, Yoshiki Hamada, and Mitsuko Masutani. 2022. "Possible Action of Olaparib for Preventing Invasion of Oral Squamous Cell Carcinoma In Vitro and In Vivo" International Journal of Molecular Sciences 23, no. 5: 2527. https://doi.org/10.3390/ijms23052527