
Functional Redundancy of FLOWERING LOCUS T 3b in Soybean Flowering Time Regulation

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Identification of the FT Homolog GmFT3b in Soybean
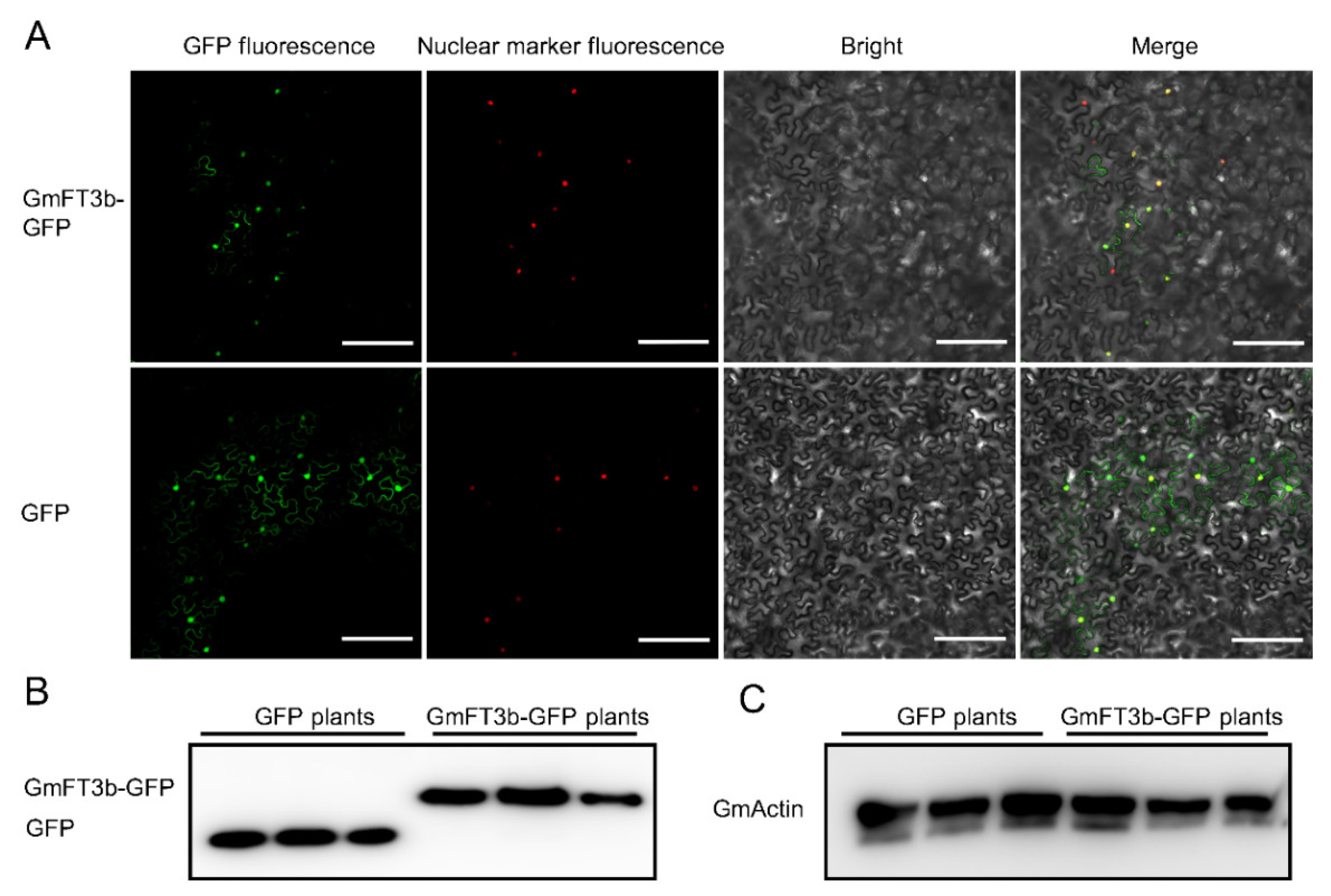
2.2. GmFT3b Is Localized to the Nucleus
2.3. Day Length and Circadian Rhythm Regulate the Expression Pattern of GmFT3b
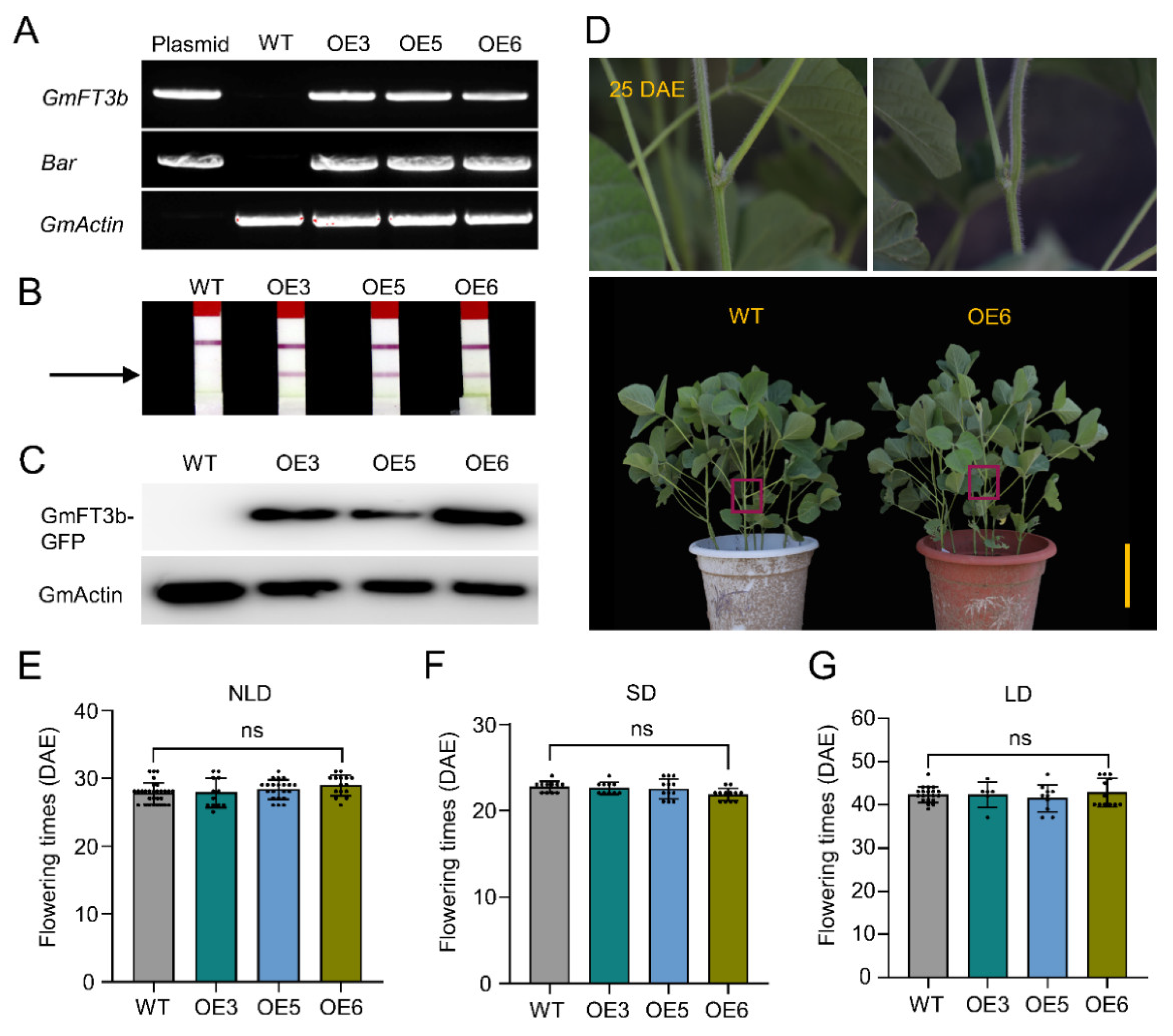
2.4. Evaluation of GmFT3b-Overexpressing Soybean Plants under Different Photoperiods
2.5. ft3b Knockout Did Not Affect Flowering Time
2.6. Expression of Downstream Flowering-Related Genes
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Bioinformatics Analysis
4.3. Generation of GmFT3b-Overexpressing Soybean Plants
4.4. CRISPR-Mediated Mutation of GmFT3b
4.5. Gene Expression Analysis
4.6. Subcellular Localization of GmFT3b
4.7. Western Blot Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Appendix B

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Oligonucleotide Sequence (5′–3′) | Annotation |
|---|---|---|
| KO3b-439F | ATATCCCTTCCCCTCGTCCT | ft3b detection |
| KO3b-439R | GGATCCACCAGGACCTTTGA | ft3b detection |
| FT3b-JC351F | CCTGGATGGCGACAAAAT | OE plants detection |
| FT3b-JC351R | GTAGCGGCTGAAGCACTG | OE plants detection |
| qGmFT3b-202-F | CTATGAAAGCCCACGACCC | GmFT3b qRT-PCR |
| qGmFT3b-202-R | TGTTCTACCACCAGAGCCACT | GmFT3b. qRT-PCR |
| qFT1a-97F | AAGTAGCGTTTCTATGGGGA | GmFT1a qRT-PCR |
| qFT1a-97R | AATTCTTGGTCGATTGAGGA | GmFT1a qRT-PCR |
| qGmFT1b-144-F | TTGAAGTTGGTGGTGATGAC | GmFT1b qRT-PCR |
| qGmFT1b-144-R | CGAAGTTTGCTCCTGTAGTT | GmFT1b qRT-PCR |
| qGmFT3a-269-F | ATAAAGAAGTGGGCAATGGT | GmFT3a qRT-PCR |
| qGmFT3a-269-R | CACAAACACGAAACGATGAA | GmFT3a qRT-PCR |
| qGmFT5b-RT-F | GGGTGTGATTGGGGATGTTC | GmFT5b qRT-PCR |
| qGmFT5b-RT-R | CAGTTCCAAGCCATTGCTAAT | GmFT5b qRT-PCR |
| qGmActin-RT-F | CGGTGGTTCTATCTTGGCATC | GmActin qRT-PCR |
| qGmActin-RT-R | GTCTTTCGCTTCAATAACCCTA | GmActin qRT-PCR |
| qGmAP1a-RT-F | TGAACATGGGTGGCAATTAC | GmAP1a qRT-PCR |
| qGmAP1a-RT-R | TGTCAAATGCCATACCAAAG | GmAP1a qRT-PCR |
| qGmAP1b-RT-F | TGGGAGCAGCCAAACTACAG | GmAP1b qRT-PCR |
| qGmAP1c-RT-F | GAAAGAAAAGGTTGCAGCTTC | GmAP1c qRT-PCR |
| qGmAP1c-RT-R | GCATCCAAGGTGACAGGAAT | GmAP1c qRT-PCR |
| qGmAP1d-F | ATCCGCACAAGGAGGAATGA | GmAP1d qRT-PCR |
| qGmAP1d-R | CCTGTAGTTTGGCTGCTCCC | GmAP1d qRT-PCR |
| qGmAP2-F | TCTTGCTCCACCCTTCTCTA | GmAP2 qRT-PCR |
| qGmAP2-R | CGAGTGGAGGAATGTCATGTT | GmAP2 qRT-PCR |
| qGmAP3-F | GAGGATAGAGAACACCACCAAC | GmAP3 qRT-PCR |
| qGmAP3-R | AAACCTTGGCATCGCATAGA | GmAP3 qRT-PCR |
| GmSOC1a-RT-F | CGAGTTGCTTTTTTTCCCTAG | GmSOC1a qRT-PCR |
| GmSOC1a-RT-R | TGAGTCTTTCCTCTCACCAT | GmSOC1a qRT-PCR |
| GmSOC1b-RT-F | AAGAAGCCCAACTGCAATGT | GmSOC1b qRT-PCR |
| GmSOC1b-RT-R | GGGCTTCAGAAATGAGGAAAGG | GmSOC1b qRT-PCR |
| qGmLFY1-F | TGAACAGCCTTTCCCAGATT | GmLFY1-qRT-PCR |
| qGmLFY1-R | GGAGGTTGTTGCTGTTGTTG | GmLFY1 qRT-PCR |
| GmLFY2-RT-F | TGACGAAGGAAACATTAACACTGG | GmLFY2 qRT-PCR |
| GmLFY2-RT-R | GCCTGAACCTGCATCAAGAA | GmLFY2 qRT-PCR |
| GmFUL1a-RT-F | CTCCCACAACAACACTAGCTC | GmFUL1a qRT-PCR |
| GmFUL1a-RT-R | CCTACAAGACAATTCCAACACGA | GmFUL1a qRT-PCR |
| qGmFUL1b-F | CCCACAACAACACTAGCTCTCA | GmFUL1b qRT-PCR |
| qGmFUL1b-R | AGTAGTAGCACCCTTCAATT | GmFUL1b qRT-PCR |
| qGmFUL2a-F | CTAATGAAGAAACTCCAACCTCA | GmFUL2a qRT-PCR |
| qGmFUL2a-R | GGTATAGTCACCGTCAAATGCCT | GmFUL2a qRT-PCR |
| qGmFUL2b-F | GTAATGAAGAAACTCCAACGTCGA | GmFUL2b qRT-PCR |
| qGmFUL2b-R | GCAGTCAGAAACGTCACACA | GmFUL2b qRT-PCR |
| qGmFUL3a-F | GACTGAAGGTCCACATACTG | GmFUL3a qRT-PCR |
| qGmFUL3a-R | TGTCATAATATCACATGTCAC | GmFUL3a qRT-PCR |
| Cas9-F | CCAGGATTAGAATGATTAGGC | Cas9 detection |
| Cas9-R | GGAAGGAGGAAGACAAGGA | Cas9 detection |
References
- Li, X.M.; Fang, C.; Yang, Y.Q.; Lv, T.X.; Su, T.; Chen, L.Y.; Nan, H.Y.; Li, S.C.; Zhao, X.H.; Lu, S.J.; et al. Overcoming the genetic compensation response of soybean florigens to improve adaptation and yield at low latitudes. Curr. Biol. 2021, 31, 3755–3767. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Cai, Y.P.; Qu, M.G.; Wang, L.W.; Sun, H.B.; Jiang, B.J.; Wu, T.T.; Liu, L.P.; Sun, S.; Wu, C.X.; et al. Soybean adaption to high-latitude regions is associated with natural variations of GmFT2b, an ortholog of FLOWERING LOCUS T. Plant Cell Environ. 2020, 43, 934–944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, S.J.; Zhao, X.H.; Hu, Y.L.; Liu, S.L.; Nan, H.Y.; Li, X.M.; Fang, C.; Cao, D.; Shi, X.Y.; Kong, L.P.; et al. Natural variation at the soybean J locus improves adaptation to the tropics and enhances yield. Nat. Genet. 2017, 49, 773–779. [Google Scholar] [CrossRef] [PubMed]
- Srikanth, A.; Schmid, M. Regulation of flowering time: All roads lead to Rome. Cell. Mol. Life Sci. 2011, 68, 2013–2037. [Google Scholar] [CrossRef]
- Xia, Z.J.; Zhai, H.; Wu, H.Y.; Xu, K.; Watanabe, S.; Harada, K. The Synchronized efforts to decipher the molecular basis for soybean maturity loci E1, E2, and E3 that regulate flowering and maturity. Front. Plant Sci. 2021, 12, 632754. [Google Scholar] [CrossRef]
- Corbesier, L.; Vincent, C.; Jang, S.H.; Fornara, F.; Fan, Q.Z.; Searle, I.; Giakountis, A.; Farrona, S.; Gissot, L.; Turnbull, C.; et al. FT protein movement contributes to long-distance signaling in floral induction of Arabidopsis. Science 2007, 316, 1030–1033. [Google Scholar] [CrossRef] [Green Version]
- Fornara, F.; de Montaigu, A.; Coupland, G. SnapShot: Control of flowering in Arabidopsis. Cell 2010, 141, 550.e2. [Google Scholar] [CrossRef] [Green Version]
- Kardailsky, I.; Shukla, V.K.; Ahn, J.H.; Dagenais, N.; Christensen, S.K.; Nguyen, J.T.; Chory, J.; Harrison, M.J.; Weigel, D. Activation tagging of the floral inducer FT. Science 1999, 286, 1962–1965. [Google Scholar] [CrossRef] [Green Version]
- Luo, X.; Yin, M.; He, Y. Molecular genetic understanding of photoperiodic regulation of flowering time in Arabidopsis and soybean. Int. J. Mol. Sci. 2021, 23, 466. [Google Scholar] [CrossRef]
- Kim, Y.M.; Park, S.J.; Kim, Y.M.; Nam, H. Subcellular localization of GIGANTEA regulates the timing of leaf senescence and flowering in Arabidopsis. Front. Plant Sci. 2020, 11, 589707. [Google Scholar] [CrossRef]
- Abe, M.; Kobayashi, Y.; Yamamoto, S.; Daimon, Y.; Yamaguchi, A.; Ikeda, Y.; Ichinoki, H.; Notaguchi, M.; Goto, K.; Araki, T. FD, a bZIP protein mediating signals from the floral pathway integrator ft at the shoot apex. Science 2005, 309, 1052–1056. [Google Scholar] [CrossRef]
- Shen, G.; Liu, N.; Zhang, J.; Xu, Y.; Baldwin, I.T.; Wu, J. Cuscuta australis (dodder) parasite eavesdrops on the host plants’ FT signals to flower. Proc. Natl. Acad. Sci. USA 2020, 117, 23125–23130. [Google Scholar] [CrossRef]
- Ferrandiz, C.; Gu, Q.; Martienssen, R.; Yanofsky, M.F. Redundant regulation of meristem identity and plant architecture by FRUITFULL, APETALA1 and CAULIFLOWER. Development 2000, 127, 725–734. [Google Scholar] [CrossRef]
- Lee, J.; Lee, I. Regulation and function of SOC1, a flowering pathway integrator. J. Exp. Bot. 2010, 61, 2247–2254. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Wang, M.; Zhao, C.; Liu, T.; Liu, Z.; Fan, Y.; Xue, Y.; Li, W.; Zhang, X.; Zhao, L. GmFULc is induced by short days in soybean and may accelerate flowering in transgenic Arabidopsis thaliana. Int. J. Mol. Sci. 2021, 22, 10333. [Google Scholar] [CrossRef]
- Quiroz, S.; Yustis, J.C.; Chavez-Hernandez, E.C.; Martinez, T.; Sanchez, M.D.; Garay-Arroyo, A.; Alvarez-Buylla, E.R.; Garcia-Ponce, B. Beyond the genetic pathways, flowering regulation complexity in Arabidopsis thaliana. Int. J. Mol. Sci. 2021, 22, 5716. [Google Scholar] [CrossRef]
- Taoka, K.; Ohki, I.; Tsuji, H.; Furuita, K.; Hayashi, K.; Yanase, T.; Yamaguchi, M.; Nakashima, C.; Purwestri, Y.A.; Tamaki, S.; et al. 14-3-3 proteins act as intracellular receptors for rice Hd3a florigen. Nature 2011, 476, 332–335. [Google Scholar] [CrossRef]
- Shim, J.S.; Jang, G. Environmental signal-dependent regulation of flowering time in rice. Int. J. Mol. Sci. 2020, 21, 6155. [Google Scholar] [CrossRef]
- Cao, D.; Takeshima, R.; Zhao, C.; Liu, B.; Jun, A.; Kong, F. Molecular mechanisms of flowering under long days and stem growth habit in soybean. J. Exp. Bot. 2017, 68, 1873–1884. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.Y.; Liu, B.H.; Weller, J.L.; Abe, J.; Kong, F.J. Molecular mechanisms for the photoperiodic regulation of flowering in soybean. J. Integr. Plant Biol. 2021, 63, 981–994. [Google Scholar] [CrossRef]
- Carpentleri-Pipolo, V.; de Almeida, L.A.; Kiihl, R.A.D. Inheritance of a long juvenile period under short-day conditions in soybean. Genet. Mol. Biol. 2002, 25, 463–469. [Google Scholar] [CrossRef] [Green Version]
- Bonato, E.R.; Vello, N.A. E-6, a dominant gene conditioning early flowering and maturity in soybeans. Genet. Mol. Biol. 1999, 22, 229–232. [Google Scholar] [CrossRef] [Green Version]
- Cober, E.R.; Molnar, S.J.; Charette, M.; Voldeng, H.D. A new locus for early maturity in soybean. Crop. Sci. 2010, 50, 524–527. [Google Scholar] [CrossRef]
- Cheng, Q.; Dong, L.D.; Su, T.; Li, T.Y.; Gan, Z.R.; Nan, H.Y.; Lu, S.J.; Fang, C.; Kong, L.P.; Li, H.Y.; et al. CRISPR/Cas9-mediated targeted mutagenesis of GmLHY genes alters plant height and internode length in soybean. BMC Plant Biol. 2019, 19, 562. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.; Wang, L.; Chen, L.; Wu, T.; Liu, L.; Sun, S.; Wu, C.; Yao, W.; Jiang, B.; Yuan, S.; et al. Mutagenesis of GmFT2a and GmFT5a mediated by CRISPR/Cas9 contributes for expanding the regional adaptability of soybean. Plant Biotechnol. J. 2020, 18, 298–309. [Google Scholar] [CrossRef] [Green Version]
- Takeshima, R.; Nan, H.Y.; Harigai, K.; Dong, L.D.; Zhu, J.H.; Lu, S.J.; Xu, M.L.; Yamagishi, N.; Yoshikawa, N.; Liu, B.H.; et al. Functional divergence between soybean FLOWERING LOCUS T orthologues FT2a and FT5a in post-flowering stem growth. J. Exp. Bot. 2019, 70, 3941–3953. [Google Scholar] [CrossRef]
- Lee, S.H.; Choi, C.W.; Park, K.M.; Jung, W.H.; Chun, H.J.; Baek, D.; Cho, H.M.; Jin, B.J.; Park, M.S.; No, D.H.; et al. Diversification in functions and expressions of soybean FLOWERING LOCUS T genes fine-tunes seasonal flowering. Front. Plant Sci. 2021, 12, 613675. [Google Scholar] [CrossRef]
- Liu, W.; Jiang, B.J.; Ma, L.M.; Zhang, S.W.; Zhai, H.; Xu, X.; Hou, W.S.; Xia, Z.J.; Wu, C.X.; Sun, S.; et al. Functional diversification of Flowering Locus T homologs in soybean: GmFT1a and GmFT2a/5a have opposite roles in controlling flowering and maturation. New Phytol. 2018, 217, 1335–1345. [Google Scholar] [CrossRef] [Green Version]
- Zhai, H.; Lu, S.X.; Liang, S.; Wu, H.Y.; Zhang, X.Z.; Liu, B.H.; Kong, F.J.; Yuan, X.H.; Li, J.; Xia, Z.J. GmFT4, a Homolog of FLOWERING LOCUS T, is positively regulated by E1 and functions as a flowering repressor in soybean. PLoS ONE 2014, 9, e89030. [Google Scholar] [CrossRef] [Green Version]
- Samanfar, B.; Molnar, S.J.; Charette, M.; Schoenrock, A.; Dehne, F.; Golshani, A.; Belzile, F.; Cober, E.R. Mapping and identification of a potential candidate gene for a novel maturity locus, E10, in soybean. Theor. Appl. Genet. 2017, 130, 377–390. [Google Scholar] [CrossRef]
- Pin, P.A.; Benlloch, R.; Bonnet, D.; Wremerth-Weich, E.; Kraft, T.; Gielen, J.J.L.; Nilsson, O. An antagonistic pair of FT homologs mediates the control of flowering time in sugar beet. Science 2010, 330, 1397–1400. [Google Scholar] [CrossRef]
- Kotoda, N.; Hayashi, H.; Suzuki, M.; Igarashi, M.; Hatsuyama, Y.; Kidou, S.; Igasaki, T.; Nishiguchi, M.; Yano, K.; Shimizu, T.; et al. Molecular characterization of FLOWERING LOCUS T-Like genes of apple (Malus × domestica Borkh.). Plant Cell Physiol. 2010, 51, 561–575. [Google Scholar] [CrossRef] [Green Version]
- Lee, R.; Baldwin, S.; Kenel, F.; McCallum, J.; Macknight, R. FLOWERING LOCUS T genes control onion bulb formation and flowering. Nat. Commun. 2013, 4, 2884. [Google Scholar] [CrossRef] [Green Version]
- Harig, L.; Beinecke, F.A.; Oltmanns, J.; Muth, J.; Muller, O.; Ruping, B.; Twyman, R.M.; Fischer, R.; Prufer, D.; Noll, G.A. Proteins from the FLOWERING LOCUS T-like subclade of the PEBP family act antagonistically to regulate floral initiation in tobacco. Plant J. 2012, 72, 908–921. [Google Scholar] [CrossRef]
- Wickland, D.P.; Hanzawa, Y. The FLOWERING LOCUS T/TERMINAL FLOWER 1 gene family: Functional evolution and molecular mechanisms. Mol. Plant 2015, 8, 983–997. [Google Scholar] [CrossRef] [Green Version]
- Hymowitz, T. On the domestication of soybean. Econ. Bot. 1970, 24, 408–421. [Google Scholar] [CrossRef]
- Wu, F.Q.; Sedivy, E.J.; Price, W.B.; Haider, W.; Hanzawa, Y. Evolutionary trajectories of duplicated FT homologues and their roles in soybean domestication. Plant J. 2017, 90, 941–953. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.H.; Singh, M.B.; Bhalla, P.L. Molecular characterization of a soybean FT homologue, GmFT7. Sci. Rep. 2021, 11, 3651. [Google Scholar] [CrossRef]
- Cai, Y.; Chen, L.; Liu, X.; Guo, C.; Sun, S.; Wu, C.; Jiang, B.; Han, T.; Hou, W. CRISPR/Cas9-mediated targeted mutagenesis of GmFT2a delays flowering time in soya bean. Plant Biotechnol. J. 2018, 16, 176–185. [Google Scholar] [CrossRef] [Green Version]
- Kaessmann, H. Origins, evolution, and phenotypic impact of new genes. Genome Res. 2010, 20, 1313–1326. [Google Scholar] [CrossRef] [Green Version]
- Conant, G.C.; Wolfe, K.H. Turning a hobby into a job: How duplicated genes find new functions. Nat. Rev. Genet. 2008, 9, 938–950. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.P.; Zhu, P.P.; Shi, H.; Guo, L.W.; Zhang, Q.H.; Chen, Y.N.; Chen, S.M.; Zhang, Z.; Peng, J.R.; Chen, J. PTC-bearing mRNA elicits a genetic compensation response via Upf3a and COMPASS components. Nature 2019, 568, 259–263. [Google Scholar] [CrossRef]
- Blackman, B.K.; Strasburg, J.L.; Raduski, A.R.; Michaels, S.D.; Rieseberg, L.H. The role of recently derived FT paralogs in sunflower domestication. Curr. Biol. 2010, 20, 629–635. [Google Scholar] [CrossRef] [Green Version]
- Jiang, B.J.; Zhang, S.W.; Song, W.W.; Abdul, M.; Khan, A.; Sun, S.; Zhang, C.S.; Wu, T.T.; Wu, C.X.; Han, T.F. Natural variations of FT family genes in soybean varieties covering a wide range of maturity groups. BMC Genom. 2019, 20, 230. [Google Scholar] [CrossRef]
- Kazan, K.; Lyons, R. The link between flowering time and stress tolerance. J. Exp. Bot. 2015, 67, 47–60. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Cai, Y.P.; Liu, X.J.; Yao, W.W.; Guo, C.; Sun, S.; Wu, C.X.; Jiang, B.J.; Han, T.F.; Hou, W.S. Improvement of soybean Agrobacterium-mediated transformation efficiency by adding glutamine and asparagine into the culture media. Int. J. Mol. Sci. 2018, 19, 3039. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.; Chen, L.; Sun, S.; Wu, C.; Yao, W.; Jiang, B.; Han, T.; Hou, W. CRISPR/Cas9-mediated deletion of large genomic fragments in soybean. Int. J. Mol. Sci. 2018, 19, 3835. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Lyu, J.; Cai, Z.D.; Li, Y.H.; Suo, H.C.; Yi, R.; Zhang, S.; Nian, H. The floral repressor GmFLC-like is involved in regulating flowering time mediated by low temperature in soybean. Int. J. Mol. Sci. 2020, 21, 1322. [Google Scholar] [CrossRef] [Green Version]
- Su, Q.; Wang, K.; Zhang, Z. Ecotopic expression of the antimicrobial peptide DmAMP1W improves resistance of transgenic wheat to two diseases: Sharp eyespot and common root rot. Int. J. Mol. Sci. 2020, 21, 647. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, Q.; Chen, L.; Cai, Y.; Chen, Y.; Yuan, S.; Li, M.; Zhang, J.; Sun, S.; Han, T.; Hou, W. Functional Redundancy of FLOWERING LOCUS T 3b in Soybean Flowering Time Regulation. Int. J. Mol. Sci. 2022, 23, 2497. https://doi.org/10.3390/ijms23052497
Su Q, Chen L, Cai Y, Chen Y, Yuan S, Li M, Zhang J, Sun S, Han T, Hou W. Functional Redundancy of FLOWERING LOCUS T 3b in Soybean Flowering Time Regulation. International Journal of Molecular Sciences. 2022; 23(5):2497. https://doi.org/10.3390/ijms23052497
Chicago/Turabian StyleSu, Qiang, Li Chen, Yupeng Cai, Yingying Chen, Shan Yuan, Min Li, Jialing Zhang, Shi Sun, Tianfu Han, and Wensheng Hou. 2022. "Functional Redundancy of FLOWERING LOCUS T 3b in Soybean Flowering Time Regulation" International Journal of Molecular Sciences 23, no. 5: 2497. https://doi.org/10.3390/ijms23052497