
Chicoric Acid Prevents Neuroinflammation and Neurodegeneration in a Mouse Parkinson’s Disease Model: Immune Response and Transcriptome Profile of the Spleen and Colon


Abstract
:1. Introduction
2. Results
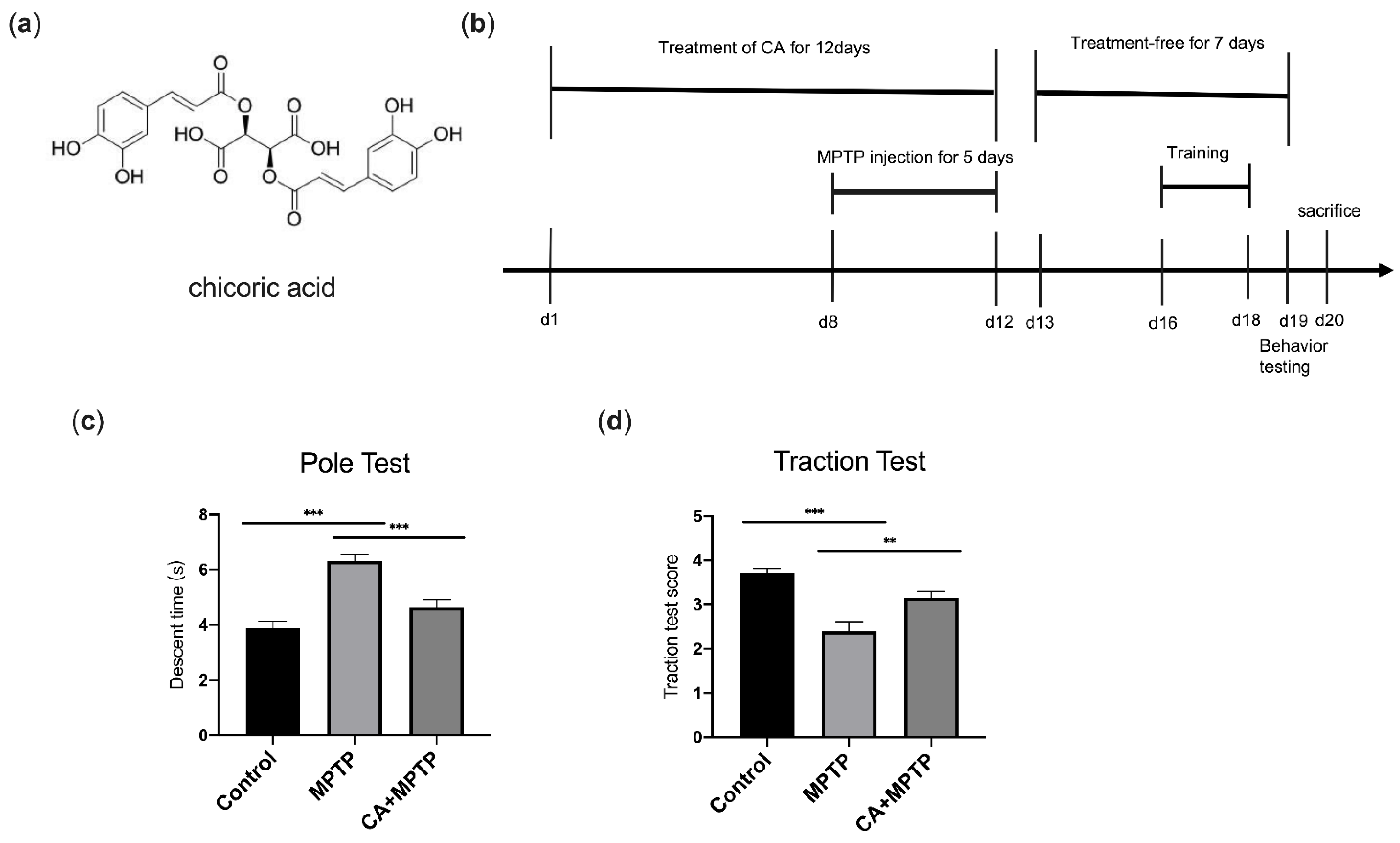
2.1. CA Improves Motor Deficits in PD Mice
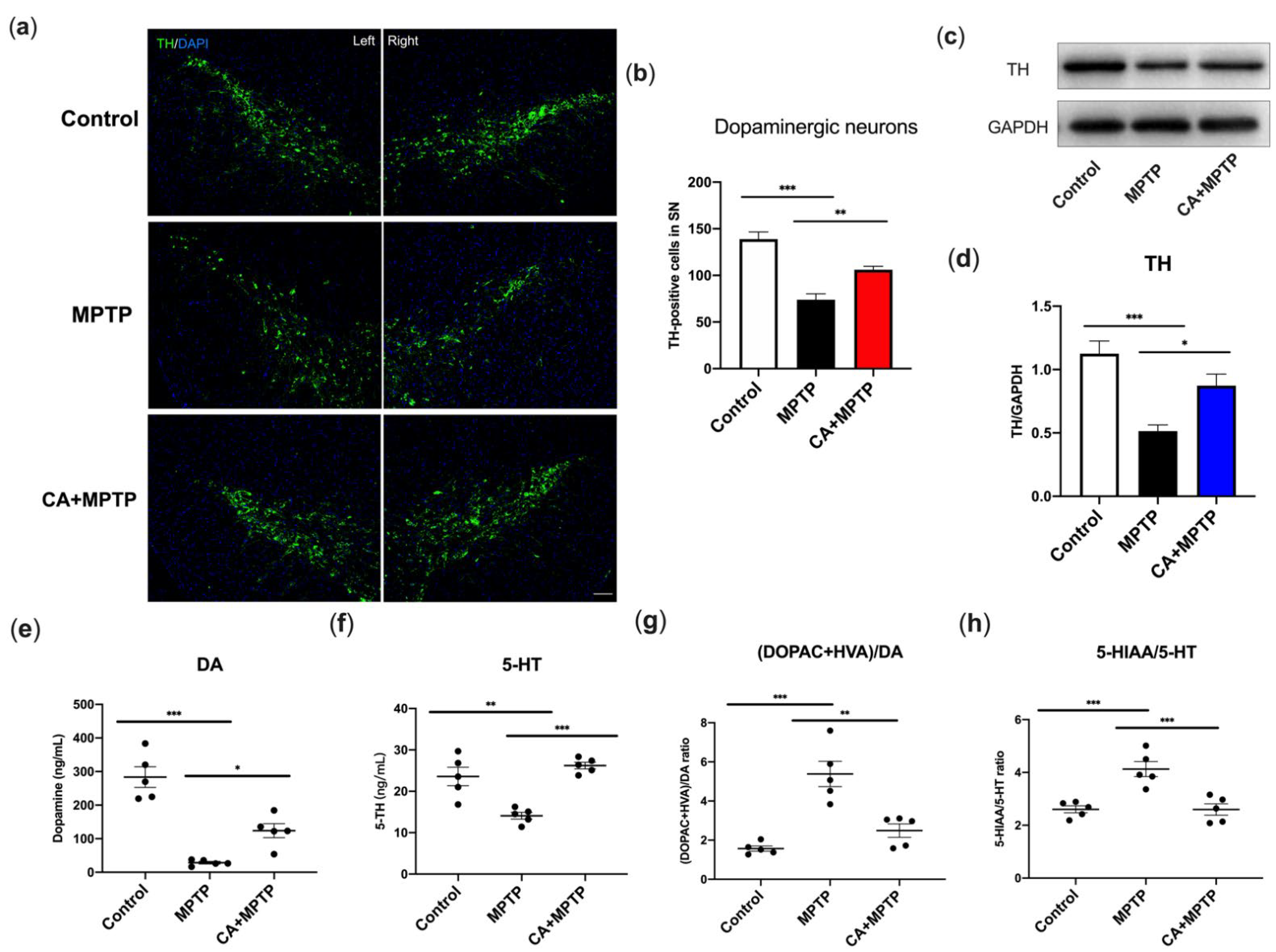
2.2. CA Promoted Dopaminergic Neuron Survival and Striatal TH Levels in PD Mice
2.3. CA Improved the Reduction in Striatal Dopamine and Serotonin of PD Mice
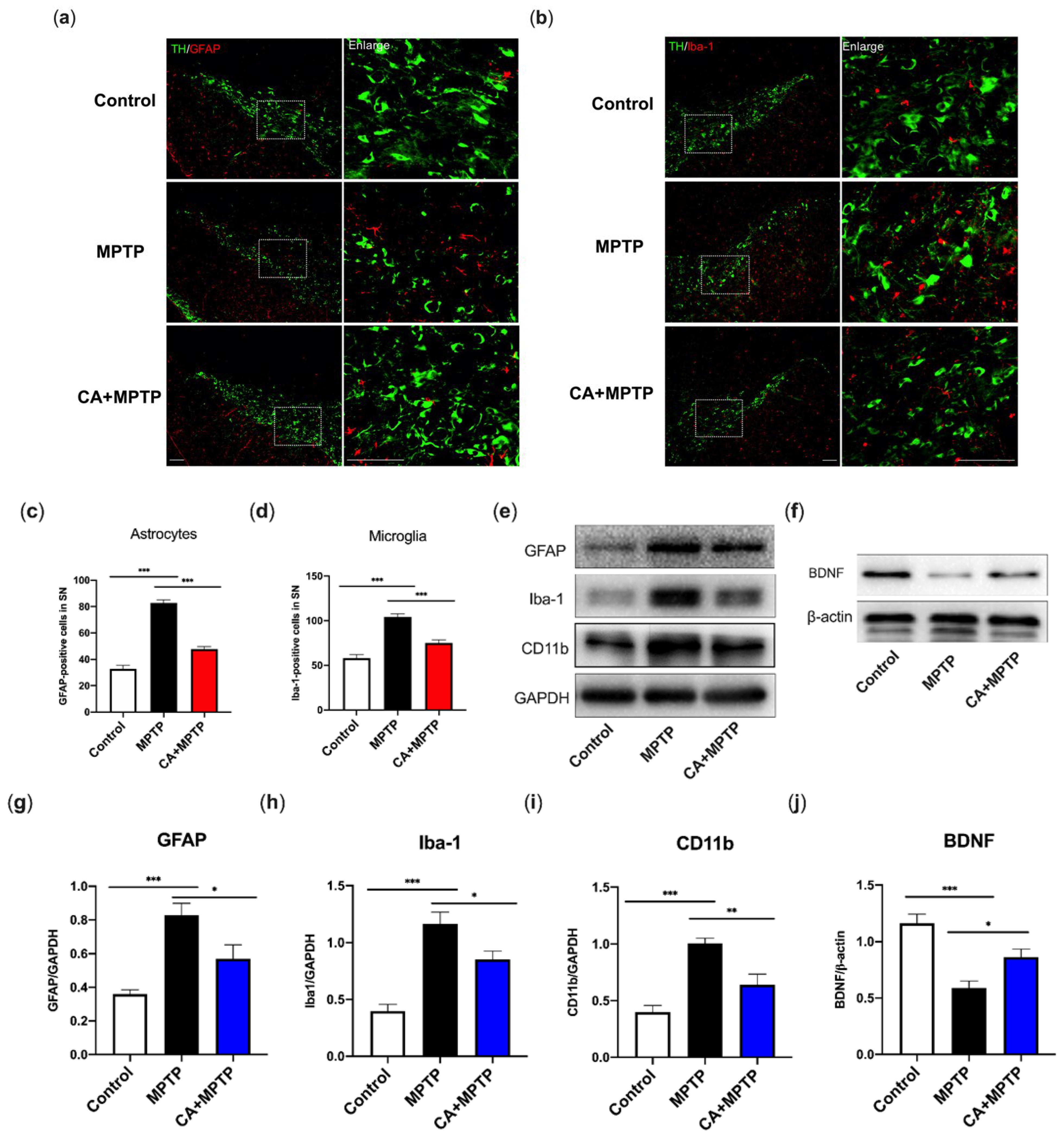
2.4. CA Suppressed Glial-Mediated Neuroinflammation Accompanied by an Increment in Striatal Neurotrophic Factors
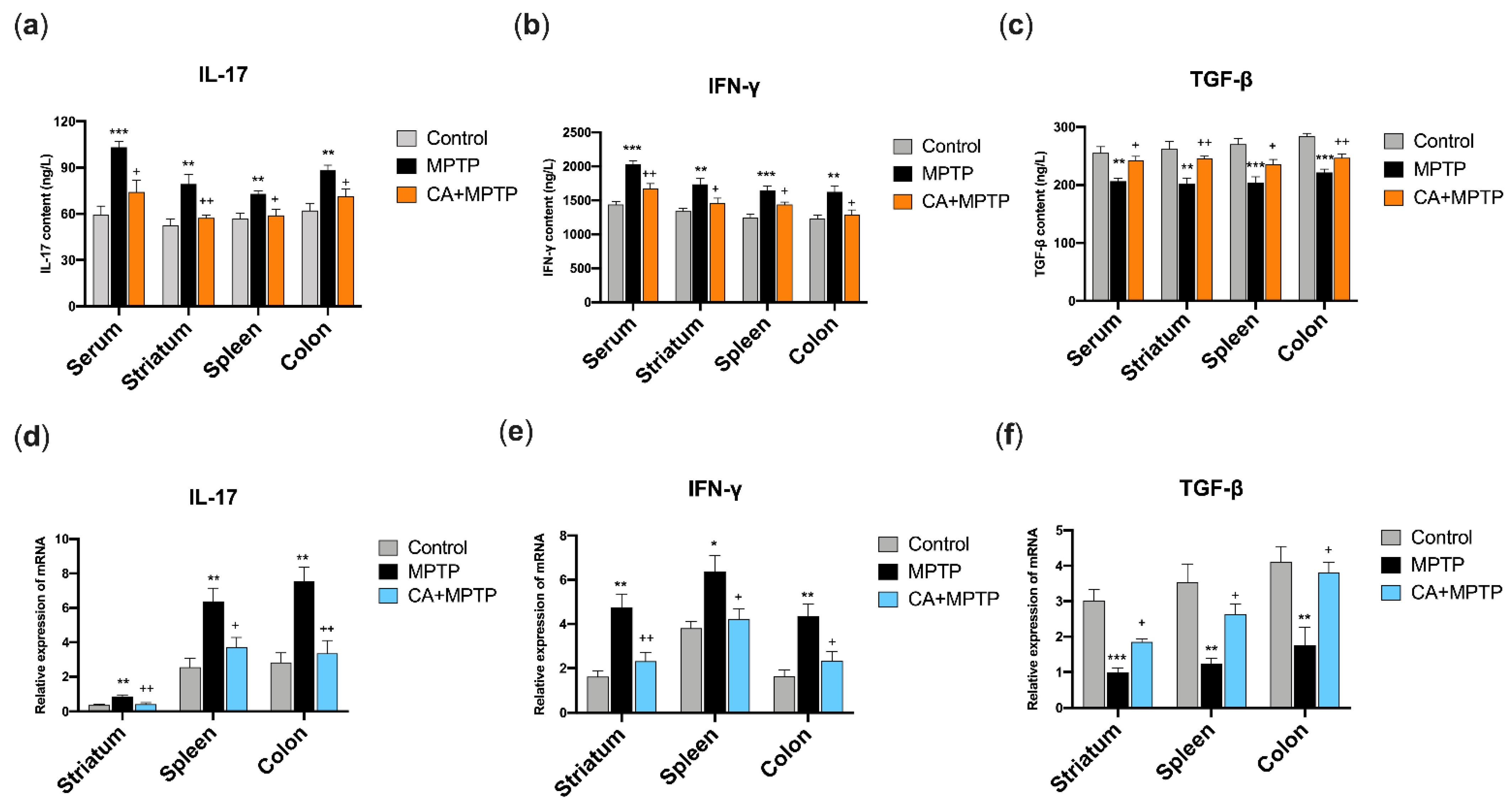
2.5. CA Restored the Protein Levels of IL-17, IFN-γ and TGF-β in the Serum, Striatum, Spleen and Solon of PD Mice
2.6. CA Restored the mRNA Levels of IL-17, IFN-γ and TGF-β in the Striatum, Spleen and Colon of PD Mice
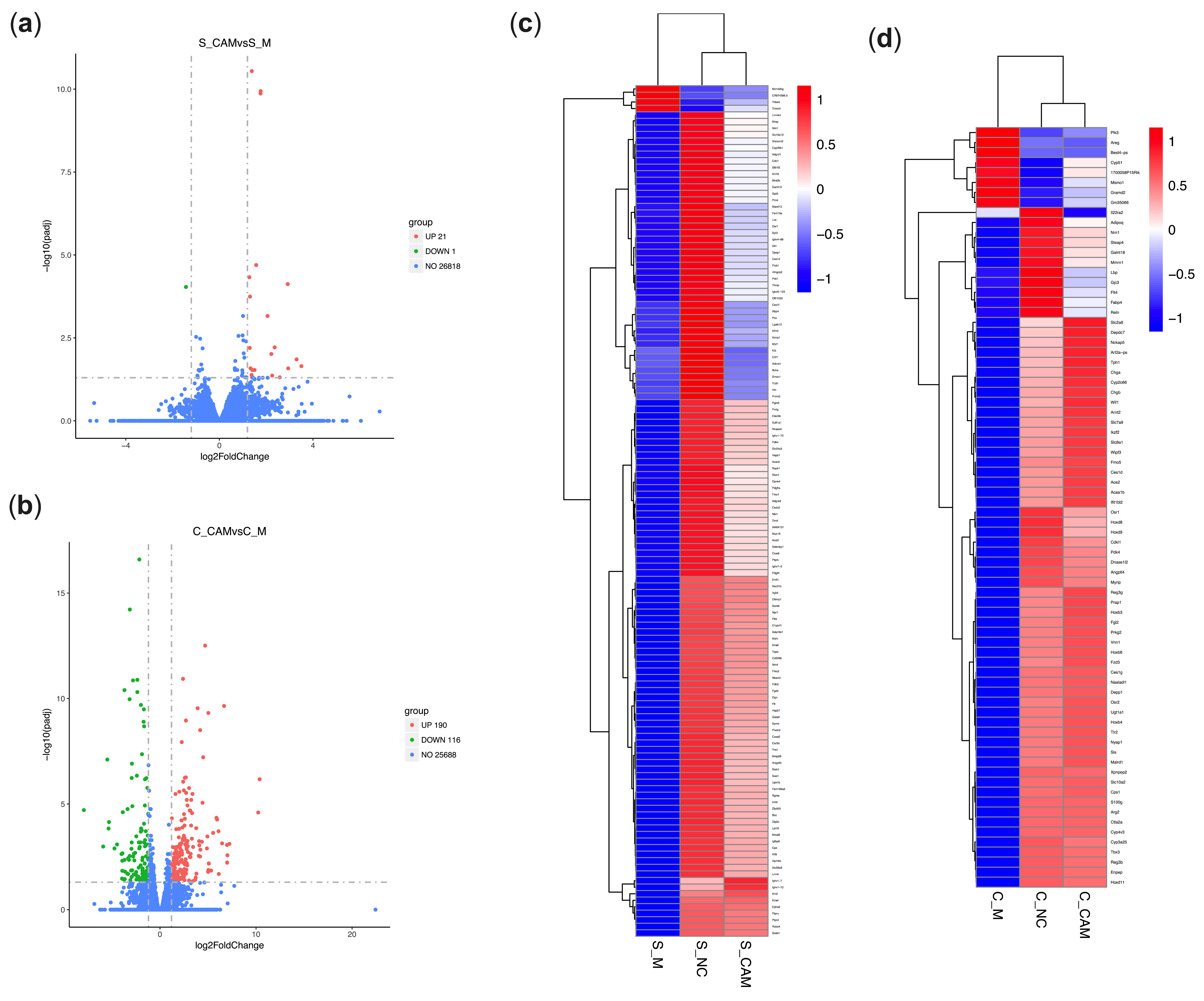
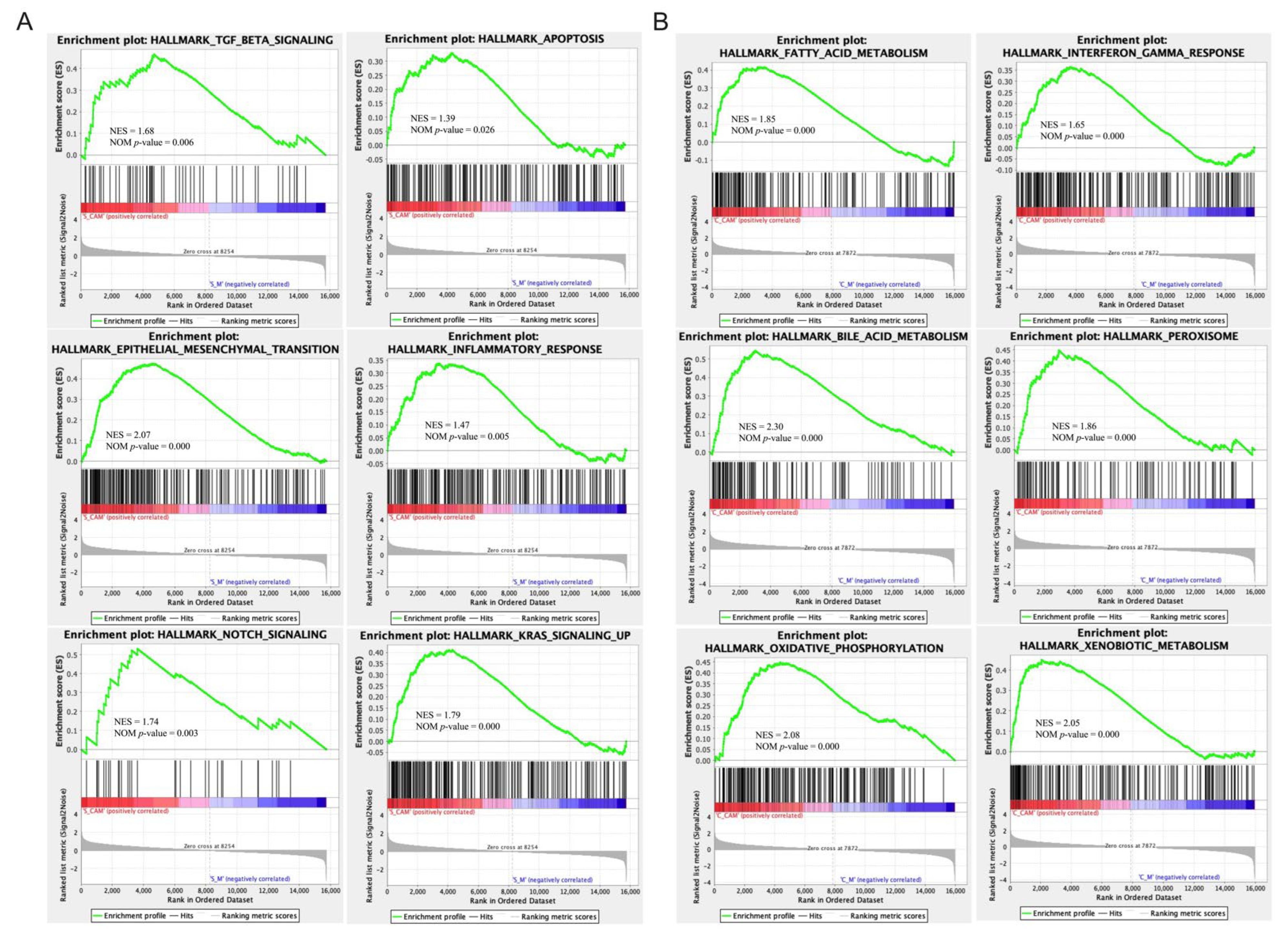
2.7. CA Modulated the Gene Expression of the Spleen in PD Mice Based on Transcriptome Analysis
2.8. CA Modulated the Gene Expression of the Colon in PD Mice Based on Transcriptome Analysis
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Animals and Treatment
4.3. Behavioral Tests
4.4. Measurement of Neurotransmitters
4.5. Immunofluorescence
4.6. Western Blot Analysis
4.7. Enzyme-Linked Immunosorbent Assay (ELISA)
4.8. qPCR for Cytokines IL-17, IFN-γ and TGF-β
4.9. The Isolation of Total RNA
4.10. cDNA Library Construction and Sequencing
4.11. Sequencing Data Analysis
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Olanow, C.W.; Tatton, W.G. Etiology and pathogenesis of Parkinson’s disease. Annu. Rev. Neurosci. 1999, 22, 123–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kish, S.J.; Shannak, K.; Hornykiewicz, O. Uneven pattern of dopamine loss in the striatum of patients with idiopathic Parkinson’s disease. Pathophysiologic and clinical implications. N. Engl. J. Med. 1988, 318, 876–880. [Google Scholar] [CrossRef] [PubMed]
- Forloni, G.; La Vitola, P.; Cerovic, M.; Balducci, C. Inflammation and Parkinson’s disease pathogenesis: Mechanisms and therapeutic insight. Prog. Mol. Biol. Transl. Sci. 2021, 177, 175–202. [Google Scholar] [CrossRef]
- Mok, S.W.; Wong, V.K.; Lo, H.H.; de Seabra Rodrigues Dias, I.R.; Leung, E.L.; Law, B.Y.; Liu, L. Natural products-based polypharmacological modulation of the peripheral immune system for the treatment of neuropsychiatric disorders. Pharmacol. Ther. 2020, 208, 107480. [Google Scholar] [CrossRef] [PubMed]
- Benkler, M.; Agmon-Levin, N.; Hassin-Baer, S.; Cohen, O.S.; Ortega-Hernandez, O.D.; Levy, A.; Moscavitch, S.D.; Szyper-Kravitz, M.; Damianovich, M.; Blank, M.; et al. Immunology, autoimmunity, and autoantibodies in Parkinson’s disease. Clin. Rev. Allergy Immunol. 2012, 42, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Dantzer, R.; O’Connor, J.C.; Freund, G.G.; Johnson, R.W.; Kelley, K.W. From inflammation to sickness and depression: When the immune system subjugates the brain. Nat. Rev. Neurosci. 2008, 9, 46–57. [Google Scholar] [CrossRef] [Green Version]
- Yan, A.; Zhang, Y.; Lin, J.; Song, L.; Wang, X.; Liu, Z. Partial depletion of peripheral M1 macrophages reverses motor deficits in MPTP-treated mouse by suppressing neuroinflammation and dopaminergic neurodegeneration. Front. Aging Neurosci. 2018, 10, 160. [Google Scholar] [CrossRef] [Green Version]
- Olson, K.E.; Namminga, K.L.; Schwab, A.D.; Thurston, M.J.; Lu, Y.; Woods, A.; Lei, L.; Shen, W.; Wang, F.; Joseph, S.B.; et al. Neuroprotective activities of long-acting granulocyte-macrophage colony-stimulating factor (mPDM608) in 1-Methyl-4-Phenyl-1,2,3,6-Tetrahydropyridine-Intoxicated Mice. Neurotherapeutics 2020, 17, 1861–1877. [Google Scholar] [CrossRef]
- Dutta, D.; Kundu, M.; Mondal, S.; Roy, A.; Ruehl, S.; Hall, D.A.; Pahan, K. RANTES-induced invasion of Th17 cells into substantia nigra potentiates dopaminergic cell loss in MPTP mouse model of Parkinson’s disease. Neurobiol. Dis. 2019, 132, 104575. [Google Scholar] [CrossRef]
- Chung, E.S.; Kim, H.; Lee, G.; Park, S.; Kim, H.; Bae, H. Neuro-protective effects of bee venom by suppression of neuroinflammatory responses in a mouse model of Parkinson’s disease: Role of regulatory T cells. Brain Behav. Immun. 2012, 26, 1322–1330. [Google Scholar] [CrossRef]
- Baizabal-Carvallo, J.F.; Alonso-Juarez, M. The link between gut dysbiosis and neuroinflammation in Parkinson’s Disease. Neuroscience 2020, 432, 160–173. [Google Scholar] [CrossRef] [PubMed]
- Sampson, T.R.; Debelius, J.W.; Thron, T.; Janssen, S.; Shastri, G.G.; Ilhan, Z.E.; Challis, C.; Schretter, C.E.; Rocha, S.; Gradinaru, V.; et al. Gut microbiota regulate motor deficits and neuroinflammation in a model of Parkinson’s Disease. Cell 2016, 167, 1469–1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salat-Foix, D.; Tran, K.; Ranawaya, R.; Meddings, J.; Suchowersky, O. Increased intestinal permeability and Parkinson Disease patients: Chicken or Egg? Can. J. Neurol. Sci. 2012, 39, 185–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, R.J.; Walter, G.C.; Wilder, S.L.; Baronowsky, E.A.; Powley, T.L. Alpha-synuclein-immunopositive myenteric neurons and vagal preganglionic terminals: Autonomic pathway implicated in Parkinson’s disease? Neuroscience 2008, 153, 733–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Y.; Sun, Q.C.; Park, Y. The bioactive effects of chicoric acid as a functional food ingredient. J. Med. Food 2019, 22, 645–652. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Chen, Y.; Shen, C.; Xiao, Y.; Wang, Y.; Liu, Z.; Liu, X. Chicoric acid supplementation prevents systemic inflammation-induced memory impairment and amyloidogenesis via inhibition of NF-kappaB. FASEB J. 2017, 31, 1494–1507. [Google Scholar] [CrossRef] [Green Version]
- Lee, N.Y.; Chung, K.S.; Jin, J.S.; Bang, K.S.; Eom, Y.J.; Hong, C.H.; Nugroho, A.; Park, H.J.; An, H.J. Effect of chicoric acid on mast cell-mediated allergic inflammation In Vitro and In Vivo. J. Nat. Prod. 2015, 78, 2956–2962. [Google Scholar] [CrossRef]
- Kour, K.; Bani, S. Augmentation of immune response by chicoric acid through the modulation of CD28/CTLA-4 and Th1 pathway in chronically stressed mice. Neuropharmacology 2011, 60, 852–860. [Google Scholar] [CrossRef]
- Dalile, B.; Van Oudenhove, L.; Vervliet, B.; Verbeke, K. The role of short-chain fatty acids in microbiota-gut-brain communication. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 461–478. [Google Scholar] [CrossRef]
- Devos, D.; Lebouvier, T.; Lardeux, B.; Biraud, M.; Rouaud, T.; Pouclet, H.; Coron, E.; des Varannes, S.B.; Naveilhan, P.; Nguyen, J.M.; et al. Colonic inflammation in Parkinson’s disease. Neurobiol. Dis. 2013, 50, 42–48. [Google Scholar] [CrossRef]
- Yang, H.C.; Won, E.J.; Kim, M.J.; Sung, C.M.; Rhee, J.H.; Nam, K.I. Intralymphatic administration of metagonimus yokogawai-extracted protein attenuates experimental murine allergic rhinitis model. Int. Arch. Allergy Immunol. 2021, 182, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Liu, Z.; Cao, B.B.; Qiu, Y.H.; Peng, Y.P. Treg cells attenuate neuroinflammation and protect neurons in a mouse model of Parkinson’s Disease. J. Neuroimmune Pharm. 2020, 15, 224–237. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Chen, H.Q.; Huang, Y.; Qiu, Y.H.; Peng, Y.P. Transforming growth factor-beta 1 acts via T beta R-I on microglia to protect against MPP+-induced dopaminergic neuronal loss. Brain Behav. Immun. 2016, 51, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Liesz, A.; Suri-Payer, E.; Veltkamp, C.; Doerr, H.; Sommer, C.; Rivest, S.; Giese, T.; Veltkamp, R. Regulatory T cells are key cerebroprotective immunomodulators in acute experimental stroke. Nat. Med. 2009, 15, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, A.D.; Banerjee, R.; Liu, J.N.; Gendelman, H.E.; Mosley, R.L. Neuroprotective activities of CD4+CD25+ regulatory T cells in an animal model of Parkinson’s disease. J. Leukocyte. Biol. 2007, 82, 1083–1094. [Google Scholar] [CrossRef]
- Zielinska, D.; Zielinski, H.; Laparra-Llopis, J.M.; Szawara-Nowak, D.; Honke, J.; Gimenez-Bastida, J.A. Caffeic acid modulates processes associated with intestinal inflammation. Nutrients 2021, 13, 554. [Google Scholar] [CrossRef]
- Park, E.; Kim, N.; Ficarro, S.B.; Zhang, Y.; Lee, B.I.; Cho, A.; Kim, K.; Park, A.K.J.; Park, W.Y.; Murray, B.; et al. Structure and mechanism of activity-based inhibition of the EGF receptor by Mig6. Nat. Struct. Mol. Biol. 2015, 22, 703–711. [Google Scholar] [CrossRef]
- Frosi, Y.; Anastasi, S.; Ballaro, C.; Varsano, G.; Castellani, L.; Maspero, E.; Polo, S.; Alema, S.; Segatto, O. A two-tiered mechanism of EGFR inhibition by RALT/MIG6 via kinase suppression and receptor degradation. J. Cell Biol. 2010, 189, 557–571. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Zeng, F.; Forrester, S.J.; Eguchi, S.; Zhang, M.Z.; Harris, R.C. Expression and function of the epidermal growth factor receptor in physiology and disease. Physiol. Rev. 2016, 96, 1025–1069. [Google Scholar] [CrossRef]
- Wancket, L.M.; Frazier, W.J.; Liu, Y. Mitogen-activated protein kinase phosphatase (MKP)-1 in immunology, physiology, and disease. Life Sci. 2012, 90, 237–248. [Google Scholar] [CrossRef] [Green Version]
- Abraham, S.M.; Clark, A.R. Dual-specificity phosphatase 1: A critical regulator of innate immune responses. Biochem. Soc. Trans. 2006, 34, 1018–1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geng, X.; Shen, J.; Li, F.; Yip, J.; Guan, L.; Rajah, G.; Peng, C.; DeGracia, D.; Ding, Y. Phosphoenolpyruvate carboxykinase (PCK) in the brain gluconeogenic pathway contributes to oxidative and lactic injury after stroke. Mol. Neurobiol. 2021, 58, 2309–2321. [Google Scholar] [CrossRef] [PubMed]
- Ko, C.W.; Counihan, D.; Wu, J.; Hatzoglou, M.; Puchowicz, M.A.; Croniger, C.M. Macrophages with a deletion of the phosphoenolpyruvate carboxykinase 1 (Pck1) gene have a more proinflammatory phenotype. J. Biol. Chem. 2018, 293, 3399–3409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N.; Feng, B.; Hu, B.; Cheng, Y.; Guo, Y.; He, Q. Neuroprotection of chicoric acid in a mouse model of Parkinson’s disease involves gut microbiota and TLR4 signaling pathway. Food Funct. 2022; in press. [Google Scholar] [CrossRef] [PubMed]
- Brunet, A.; Datta, S.R.; Greenberg, M.E. Transcription-dependent and -independent control of neuronal survival by the PI3K-Akt signaling pathway. Curr. Opin. Neurobiol. 2001, 11, 297–305. [Google Scholar] [CrossRef]
- Hakkola, J.; Hukkanen, J.; Turpeinen, M.; Pelkonen, O. Inhibition and induction of CYP enzymes in humans: An update. Arch. Toxicol. 2020, 94, 3671–3722. [Google Scholar] [CrossRef]
- Wagner, N.; Wagner, K.D. The role of PPARs in Disease. Cells 2020, 9, 2367. [Google Scholar] [CrossRef]
- Marie, A.; Darricau, M.; Touyarot, K.; Parr-Brownlie, L.C.; Bosch-Bouju, C. Role and mechanism of vitamin a metabolism in the pathophysiology of Parkinson’s Disease. J. Parkinsons Dis. 2021, 11, 949–970. [Google Scholar] [CrossRef]
- Schiffer, L.; Barnard, L.; Baranowski, E.S.; Gilligan, L.C.; Taylor, A.E.; Arlt, W.; Shackleton, C.H.L.; Storbeck, K.H. Human steroid biosynthesis, metabolism and excretion are differentially reflected by serum and urine steroid metabolomes: A comprehensive review. J. Steroid. Biochem. Mol. Biol. 2019, 194, 105439. [Google Scholar] [CrossRef]
- Han, L.; Shen, W.J.; Bittner, S.; Kraemer, F.B.; Azhar, S. PPARs: Regulators of metabolism and as therapeutic targets in cardiovascular disease. Part I: PPAR-alpha. Future Cardiol. 2017, 13, 259–278. [Google Scholar] [CrossRef]
- Cao, Q.; Qin, L.; Huang, F.; Wang, X.; Yang, L.; Shi, H.; Wu, H.; Zhang, B.; Chen, Z.; Wu, X. Amentoflavone protects dopaminergic neurons in MPTP-induced Parkinson’s disease model mice through PI3K/Akt and ERK signaling pathways. Toxicol Appl. Pharmacol. 2017, 319, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Kuribara, H.; Higuchi, Y.; Tadokoro, S. Effects of central depressants on rota-rod and traction performances in mice. Jpn. J. Pharmacol. 1977, 27, 117–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primers | Reverse Primers |
|---|---|---|
| IL-17 | 5′-TGGACTCTGAGCCGCAATG-3′ | 5′-GGCGGACAATAGAGGAAACG-3′ |
| IFN-γ | 5′-CATAGATGTGGAAGAAAAGAG-3′ | 5′-AGAGTCTGAGGTAGAAAGAGATA-3′ |
| TGF-β | 5′-CGAAGCGGACTACTATGCTA-3′ | 5′-GAATGTCTGACGTATTGAAGAA-3′ |
| β-actin | 5′-CCTCTATGCCAACACAGT-3′ | 5′-AGCCACCAATCCACACAG-3′ |
| Category | GO ID | Description | Gene Name | Count | Up | Down | padj |
|---|---|---|---|---|---|---|---|
| BP | GO:0042036 | negative regulation of cytokine biosynthetic process | Errfi1↑Muc16↑ | 2 | 2 | 0 | 0.037678798 |
| BP | GO:0051384 | response to glucocorticoid | Errfi1↑Dusp1↑Pck1↑ | 3 | 3 | 0 | 0.037678798 |
| BP | GO:0031960 | response to corticosteroid | Errfi1↑Dusp1↑Pck1↑ | 3 | 3 | 0 | 0.037678798 |
| BP | GO:0032869 | cellular response to insulin stimulus | Pdk4↑Errfi1↑Pck1↑ | 3 | 3 | 0 | 0.037678798 |
| BP | GO:1901654 | response to ketone | Errfi1↑Dusp1↑Pck1↑ | 3 | 3 | 0 | 0.037678798 |
| BP | GO:0071549 | cellular response to dexamethasone stimulus | Errfi1↑Pck1↑ | 2 | 2 | 0 | 0.037678798 |
| BP | GO:0071548 | response to dexamethasone | Errfi1↑Pck1↑ | 2 | 2 | 0 | 0.046519989 |
| BP | GO:0002377 | immunoglobulin production | Igkv9-124↑Igkv14-100↑Igkv4-91↑ | 3 | 3 | 0 | 0.046927892 |
| BP | GO:0032868 | response to insulin | Pdk4↑Errfi1↑Pck1↑ | 3 | 3 | 0 | 0.046927892 |
| ↑ upregulation. | |||||||
| KEGG ID | Description | Gene Name | Count | Up | Down | padj |
|---|---|---|---|---|---|---|
| mmu03320 | PPAR signaling pathway | Pck1↑Plin4↑ | 2 | 2 | 0 | 0.00949221 |
| ↑ upregulation. | ||||||
| Category | GO ID | Description | Gene Name | Count | Up | Down | padj |
|---|---|---|---|---|---|---|---|
| BP | GO:0006805 | xenobiotic metabolic process | Acaa1b↑Cyp2c66↑Ugt1a1↑Lpo↑Cyp2c65↑Cyp2c55↑Nceh1↑Ugt1a7c↑Cyp2d12↓Cyp2d9↓Srd5a2↓Cyp2c68↓Cyp2c69↓Cyp2f2↓ | 14 | 8 | 6 | 1.06 × 10−8 |
| BP | GO:0042737 | drug catabolic process | Cyp2c66↑Cyp2c65↑Akr1c18↑Cyp4b1↑Adh5↑Cyp2c55↑Cubn↑Aldh3b1↑Cyp2d12↓Cyp2d9↓Adh1↓Cyp2c68↓Nt5e↓Cyp2c69↓Cyp2f2↓ | 15 | 8 | 7 | 1.79 × 10−7 |
| BP | GO:0042738 | exogenous drug catabolic process | Cyp2c66↑Cyp2c65↑Cyp4b1↑Cyp2c55↑Cyp2d12↓Cyp2d9↓Cyp2c68↓Cyp2c69↓Cyp2f2↓ | 9 | 4 | 5 | 1.79 × 10−7 |
| BP | GO:0006690 | icosanoid metabolic process | Cyp2c66↑Cyp2c65↑Akr1c18↑Cyp2c55↑Tlr2↑Ggt1↑Cyp2d12↓Cyp2d9↓Pla2g4f↓Cyp2c68↓Pla2g5↓Cyp2c69↓Cyp2f2↓ | 13 | 6 | 7 | 1.79 × 10−7 |
| BP | GO:0006631 | fatty acid metabolic process | Acaa1b↑Ces1f↑Ppara↑Cyp2c66↑Lpin2↑Ces1d↑Pdk4↑Acsf2↑Cyp2c65↑Akr1c18↑Lpl↑Cyp2c55↑Slc27a4↑Ggt1↑Ehhadh↑Abhd3↑Adipoq↑Cyp2d12↓Cyp2d9↓Pla2g4f↓Cyp2c68↓Cyp2c69↓Cyp2f2↓ | 23 | 17 | 6 | 3.37 × 10−7 |
| BP | GO:0016042 | lipid catabolic process | Acaa1b↑Ces1f↑Lpin2↑Ces1d↑Akr1c18↑Lpl↑Ces1g↑Aspg↑Nceh1↑Slc27a4↑Ugt1a7c↑Ehhadh↑Abhd3↑Adipoq↑Pla2g4f↓Srd5a2↓Pla2g2a↓Pla2g5↓Ces3a↓Hexb↓ | 20 | 14 | 6 | 7.85 × 10−7 |
| BP | GO:1901568 | fatty acid derivative metabolic process | Cyp2c66↑Cyp2c65↑Akr1c18↑Cyp2c55↑Tlr2↑Ggt1↑Cyp2d12↓Cyp2d9↓Pla2g4f↓Cyp2c68↓Pla2g5↓Cyp2c69↓Cyp2f2↓ | 13 | 6 | 7 | 2.80 × 10−6 |
| BP | GO:0046394 | carboxylic acid biosynthetic process | Gapdh↑Hkdc1↑Ppara↑Pdk4↑Akr1c18↑Nags↑Lpl↑Malrd1↑Aldh1a1↑Ggt1↑Abhd3↑Ugdh↑Tkfc↑Gpd1↑Rdh16↓Eno3↓Pla2g4f↓Hif1a↓Hk2↓Rdh9↓Pla2g5↓ | 21 | 14 | 7 | 2.80 × 10−6 |
| BP | GO:0016053 | organic acid biosynthetic process | Gapdh↑Hkdc1↑Ppara↑Pdk4↑Akr1c18↑Nags↑Lpl↑Malrd1↑Aldh1a1↑Ggt1↑Abhd3↑Ugdh↑Tkfc↑Gpd1↑Rdh16↓Eno3↓Pla2g4f↓Hif1a↓Hk2↓Rdh9↓Pla2g5↓ | 21 | 14 | 7 | 2.80 × 10−6 |
| BP | GO:0071466 | cellular response to xenobiotic stimulus | Acaa1b↑Cyp2c66↑Ugt1a1↑Lpo↑Cyp2c65↑Cyp2c55↑Nceh1↑Ugt1a7c↑Cyp2d12↓Cyp2d9↓Srd5a2↓Cyp2c68↓Cyp2c69↓Cyp2f2↓ | 14 | 8 | 6 | 2.80 × 10−6 |
| MF | GO:0016712 | oxidoreductase activity, acting on paired donors, with incorporation or reduction of molecular oxygen, reduced flavin or flavoprotein as one donor, and incorporation of one atom of oxygen | Cyp2c66↑Cyp2c65↑Cyp2d26↑Cyp4b1↑Cyp3a44↑Cyp2c55↑Cyp2d12↓Cyp2d9↓Cyp2c68↓Cyp2c69↓Cyp2f2↓ | 11 | 6 | 5 | 2.77 × 10−9 |
| MF | GO:0004497 | monooxygenase activity | Tph1↑Fmo5↑Cyp2c66↑Cyp2c65↑Cyp2d26↑Akr1c18↑Cyp4b1↑Cyp3a44↑Cyp2c55↑Akr1c19↑Cyp2d12↓Cyp2d9↓Cyp2c68↓Cyp2c69↓Cyp2f2↓ | 15 | 10 | 5 | 2.02 × 10−8 |
| MF | GO:0008395 | steroid hydroxylase activity | Cyp2c66↑Cyp2c65↑Cyp3a44↑Cyp2c55↑Cyp2d12↓Cyp2d9↓Cyp2c68↓Cyp2c69↓Cyp2f2↓ | 9 | 4 | 5 | 8.37 × 10−7 |
| MF | GO:0052689 | carboxylic ester hydrolase activity | Ces1f↑Ces1d↑Ces2b↑Car1↑Lpl↑Ces1g↑Aspg↑Nceh1↑Abhd3↑Bche↑Pla2g4f↓Pla2g2a↓Pla2g5↓Ces3a↓ | 14 | 10 | 4 | 8.37 × 10−7 |
| MF | GO:0016705 | oxidoreductase activity, acting on paired donors, with incorporation or reduction of molecular oxygen | Tph1↑Fmo5↑Cyp2c66↑Cyp2c65↑Cyp2d26↑Akr1c18↑Cyp4b1↑Cyp3a44↑Cyp2c55↑Akr1c19↑Cyp2d12↓Cyp2d9↓Cyp2c68↓Cyp2c69↓Cyp2f2↓ | 15 | 10 | 5 | 2.51 × 10−6 |
| MF | GO:0008392 | arachidonic acid epoxygenase activity | Cyp2c66↑Cyp2c65↑Cyp2c55↑Cyp2c68↓Cyp2c69↓Cyp2f2↓ | 6 | 3 | 3 | 5.79 × 10−6 |
| MF | GO:0008391 | arachidonic acid monooxygenase activity | Cyp2c66↑Cyp2c65↑Cyp2c55↑Cyp2c68↓Cyp2c69↓Cyp2f2↓ | 6 | 3 | 3 | 1.12 × 10−5 |
| MF | GO:0046906 | tetrapyrrole binding | Cyp2c66↑Lpo↑Cyp2c65↑Cyp2d26↑Cyp4b1↑Cyp3a44↑Cyp2c55↑Cubn↑Cyp2d12↓Cyp2d9↓Cyp2c68↓Cyp2c69↓Cyp2f2↓ | 13 | 8 | 5 | 1.56 × 10−5 |
| MF | GO:0005506 | iron ion binding | Tph1↑Cyp2c66↑Cyp2c65↑Cyp2d26↑Cyp4b1↑Cyp3a44↑Cyp2c55↑Cyp2d12↓Cyp2d9↓Cyp2c68↓Nt5e↓Cyp2c69↓Cyp2f2↓ | 13 | 7 | 6 | 4.05 × 10−5 |
| MF | GO:0020037 | heme binding | Cyp2c66↑Lpo↑Cyp2c65↑Cyp2d26↑Cyp4b1↑Cyp3a44↑Cyp2c55↑Cyp2d12↓Cyp2d9↓Cyp2c68↓Cyp2c69↓Cyp2f2↓ | 12 | 7 | 5 | 4.27 × 10−5 |
| ↑upregulation | |||||||
| ↓downregulation | |||||||
| KEGG ID | Description | Gene Name | Count | Up | Down | padj |
|---|---|---|---|---|---|---|
| mmu00830 | Retinol metabolism | Ugt2b5↑Cyp2c66↑Ugt1a1↑Cyp2c65↑Cyp3a44↑Aldh1a1↑Ugt2b36↑Adh5↑Cyp2c55↑Gm15368↑Ugt1a7c↑Rdh16↓Adh1↓Cyp2c68↓Rdh9↓ | 15 | 11 | 4 | 1.76 × 10−10 |
| mmu00140 | Steroid hormone biosynthesis | Ugt2b5↑Cyp2c66↑Ugt1a1↑Cyp2c65↑Cyp2d26↑Akr1c18↑Cyp3a44↑Ugt2b36↑Cyp2c55↑Gm15368↑Ugt1a7c↑Cyp2d12↓Cyp2d9↓Srd5a2↓Cyp2c68↓ | 15 | 11 | 4 | 1.99 × 10−10 |
| mmu05204 | Chemical carcinogenesis | Ugt2b5↑Cyp2c66↑Ugt1a1↑Cyp2c65↑Cyp3a44↑Ugt2b36↑Adh5↑Cyp2c55↑Gm15368↑Ugt1a7c↑Aldh3b1↑Adh1↓Cyp2c68↓ | 13 | 11 | 2 | 4.60 × 10−7 |
| mmu00591 | Linoleic acid metabolism | Cyp2c66↑Cyp2c65↑Cyp3a44↑Cyp2c55↑Pla2g4f↓Cyp2c68↓Pla2g2a↓Pla2g5↓ | 8 | 4 | 4 | 1.34 × 10−5 |
| mmu04976 | Bile secretion | Ugt2b5↑Ugt1a1↑Slc51a↑Slc10a2↑Sct↑Ugt2b36↑Gm15368↑Nceh1↑Ugt1a7c↑Adcy9↑ | 10 | 10 | 0 | 0.000109402 |
| mmu00053 | Ascorbate and aldarate metabolism | Ugt2b5↑Ugt1a1↑Ugt2b36↑Gm15368↑Ugt1a7c↑Ugdh↑ | 6 | 6 | 0 | 0.000126403 |
| mmu00982 | Drug metabolism - cytochrome P450 | Ugt2b5↑Fmo5↑Ugt1a1↑Ugt2b36↑Adh5↑Gm15368↑Ugt1a7c↑Aldh3b1↑Adh1↓ | 9 | 8 | 1 | 0.000135848 |
| mmu00980 | Metabolism of xenobiotics by cytochrome P450 | Ugt2b5↑Ugt1a1↑Ugt2b36↑Adh5↑Gm15368↑Ugt1a7c↑Aldh3b1↑Adh1↓Cyp2f2↓ | 9 | 7 | 2 | 0.000182884 |
| mmu04726 | Serotonergic synapse | Tph1↑Cyp2c66↑Cyp2c65↑Cyp2d26↑Slc18a1↑Cyp2c55↑Cyp2d12↓Cyp2d9↓Pla2g4f↓Cyp2c68↓Htr4↓ | 11 | 6 | 5 | 0.000408708 |
| mmu00590 | Arachidonic acid metabolism | Cyp2c66↑Cyp2c65↑Cyp2c55↑Ggt1↑Pla2g4f↓Cyp2c68↓Pla2g2a↓Pla2g5↓ | 8 | 4 | 4 | 0.001096684 |
| mmu00040 | Pentose and glucuronate interconversions | Ugt2b5↑Ugt1a1↑Ugt2b36↑Gm15368↑Ugt1a7c↑Ugdh↑ | 6 | 6 | 0 | 0.001096684 |
| mmu00983 | Drug metabolism - other enzymes | Ces1f↑Ugt2b5↑Ugt1a1↑Ces1d↑Ces2b↑Ugt2b36↑Gm15368↑Ugt1a7c↑Gm45727↑ | 9 | 9 | 0 | 0.0021122 |
| mmu03320 | PPAR signaling pathway | Acaa1b↑Ppara↑Lpl↑Angptl4↑Hmgcs2↑Slc27a4↑Ehhadh↑Adipoq↑ | 8 | 8 | 0 | 0.002234301 |
| mmu00860 | Porphyrin and chlorophyll metabolism | Ugt2b5↑Ugt1a1↑Ugt2b36↑Gm15368↑Ugt1a7c↑ | 5 | 5 | 0 | 0.008876674 |
| mmu00010 | Glycolysis / Gluconeogenesis | Hkdc1↑Adh5↑Pgm2↑Aldh3b1↑Eno3↓Adh1↓Hk2↓ | 7 | 4 | 3 | 0.008876674 |
| mmu00592 | alpha-Linolenic acid metabolism | Acaa1b↑Pla2g4f↓Pla2g2a↓Pla2g5↓ | 4 | 1 | 3 | 0.01005902 |
| mmu00500 | Starch and sucrose metabolism | Hkdc1↑Sis↑Pgm2↑Hk2↓ | 4 | 3 | 1 | 0.030913453 |
| mmu00520 | Amino sugar and nucleotide sugar metabolism | Hkdc1↑Pgm2↑Ugdh↑Hk2↓Hexb↓ | 5 | 3 | 2 | 0.036511025 |
| mmu00052 | Galactose metabolism | Hkdc1↑Sis↑Pgm2↑Hk2↓ | 4 | 3 | 1 | 0.041823524 |
| ↑upregulation | ||||||
| ↓downregulation | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, N.; Li, R.; Feng, B.; Cheng, Y.; Guo, Y.; Qian, H. Chicoric Acid Prevents Neuroinflammation and Neurodegeneration in a Mouse Parkinson’s Disease Model: Immune Response and Transcriptome Profile of the Spleen and Colon. Int. J. Mol. Sci. 2022, 23, 2031. https://doi.org/10.3390/ijms23042031
Wang N, Li R, Feng B, Cheng Y, Guo Y, Qian H. Chicoric Acid Prevents Neuroinflammation and Neurodegeneration in a Mouse Parkinson’s Disease Model: Immune Response and Transcriptome Profile of the Spleen and Colon. International Journal of Molecular Sciences. 2022; 23(4):2031. https://doi.org/10.3390/ijms23042031
Chicago/Turabian StyleWang, Ning, Rui Li, Bainian Feng, Yuliang Cheng, Yahui Guo, and He Qian. 2022. "Chicoric Acid Prevents Neuroinflammation and Neurodegeneration in a Mouse Parkinson’s Disease Model: Immune Response and Transcriptome Profile of the Spleen and Colon" International Journal of Molecular Sciences 23, no. 4: 2031. https://doi.org/10.3390/ijms23042031