

A Fucosylated Lactose-Presenting Tetravalent Glycocluster Acting as a Mutual Ligand of Pseudomonas aeruginosa Lectins A (PA-IL) and B (PA-IIL)—Synthesis and Interaction Studies

 , ,
, ,
Abstract
:1. Introduction
2. Results
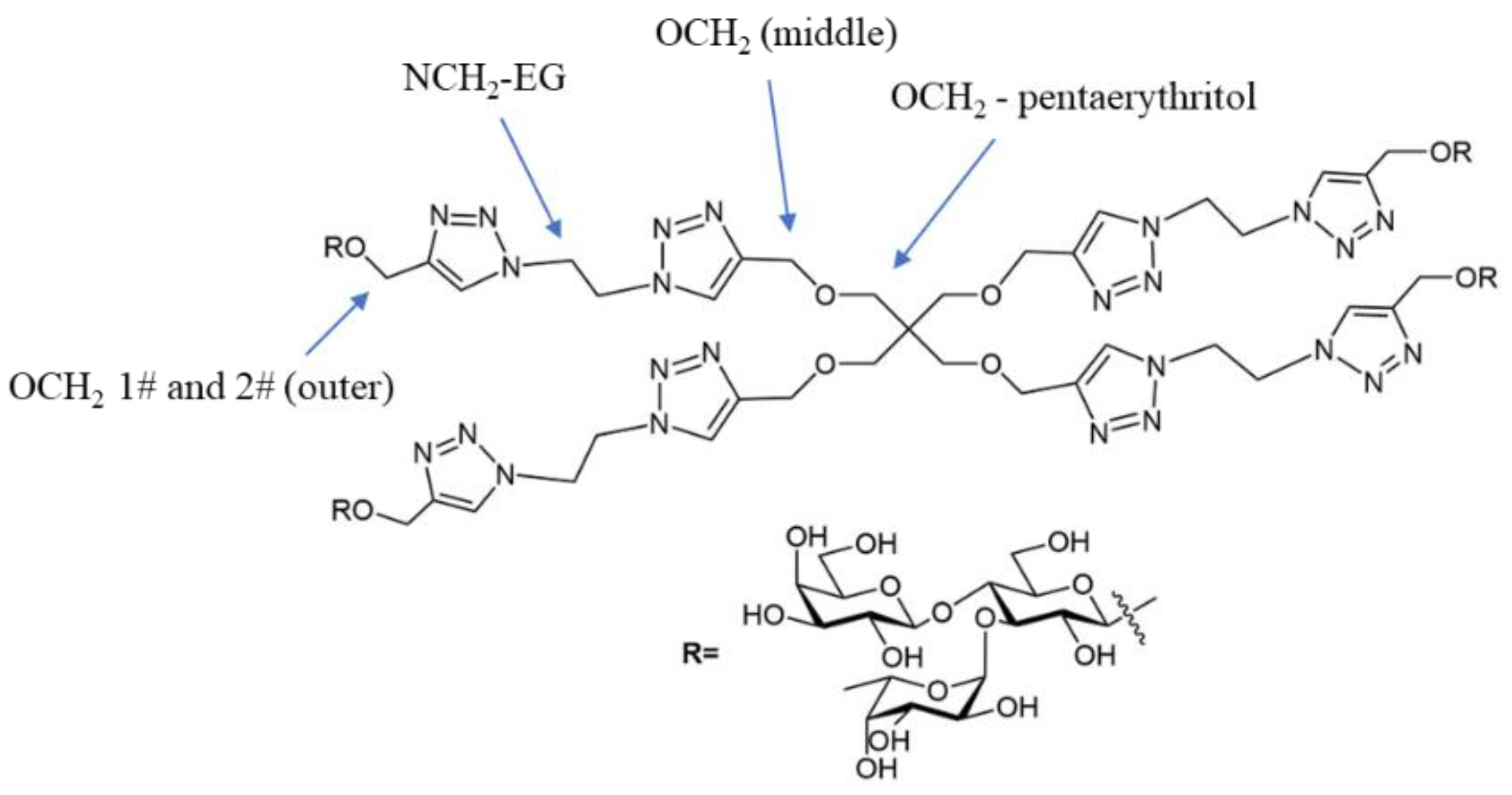
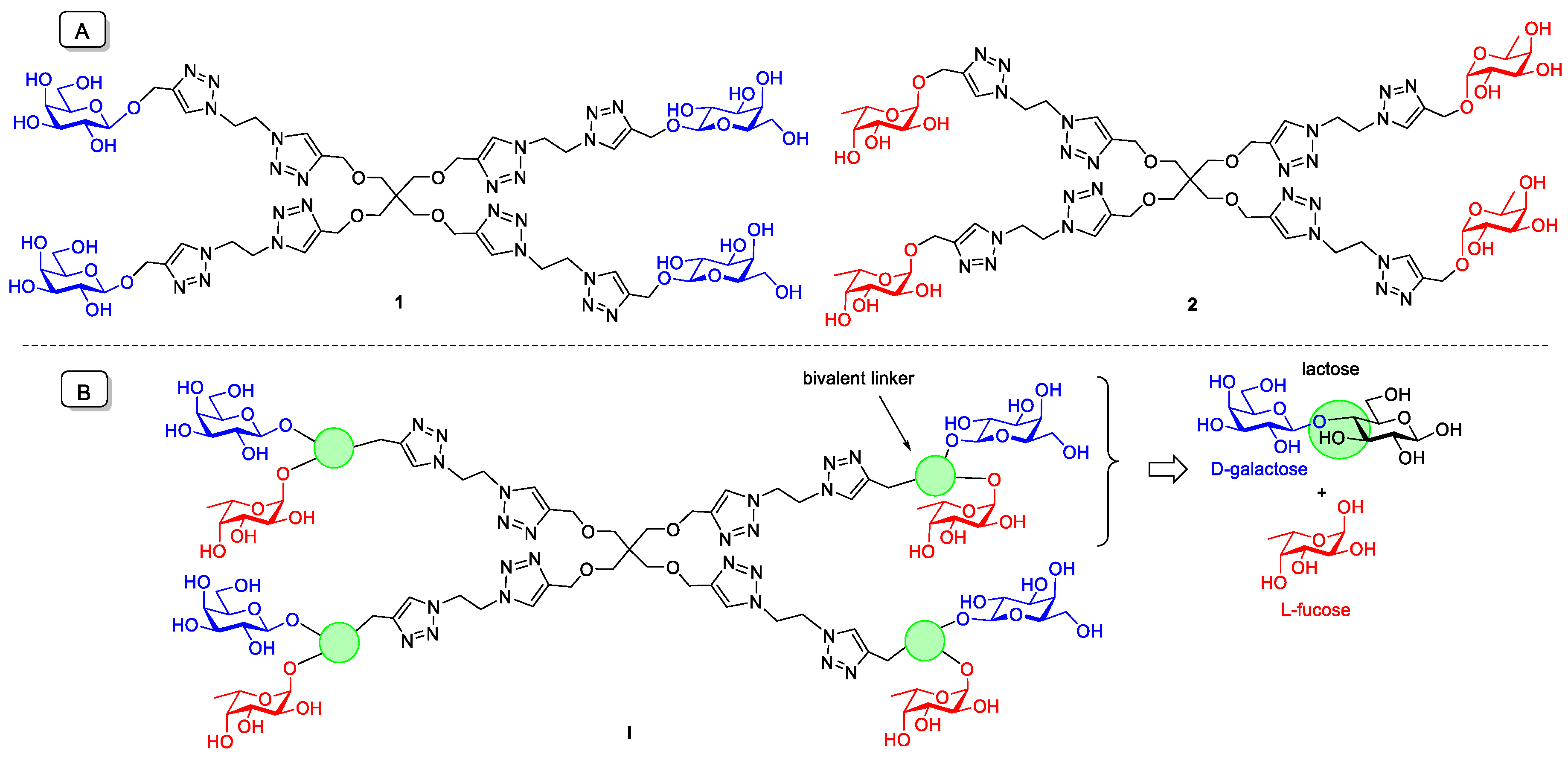
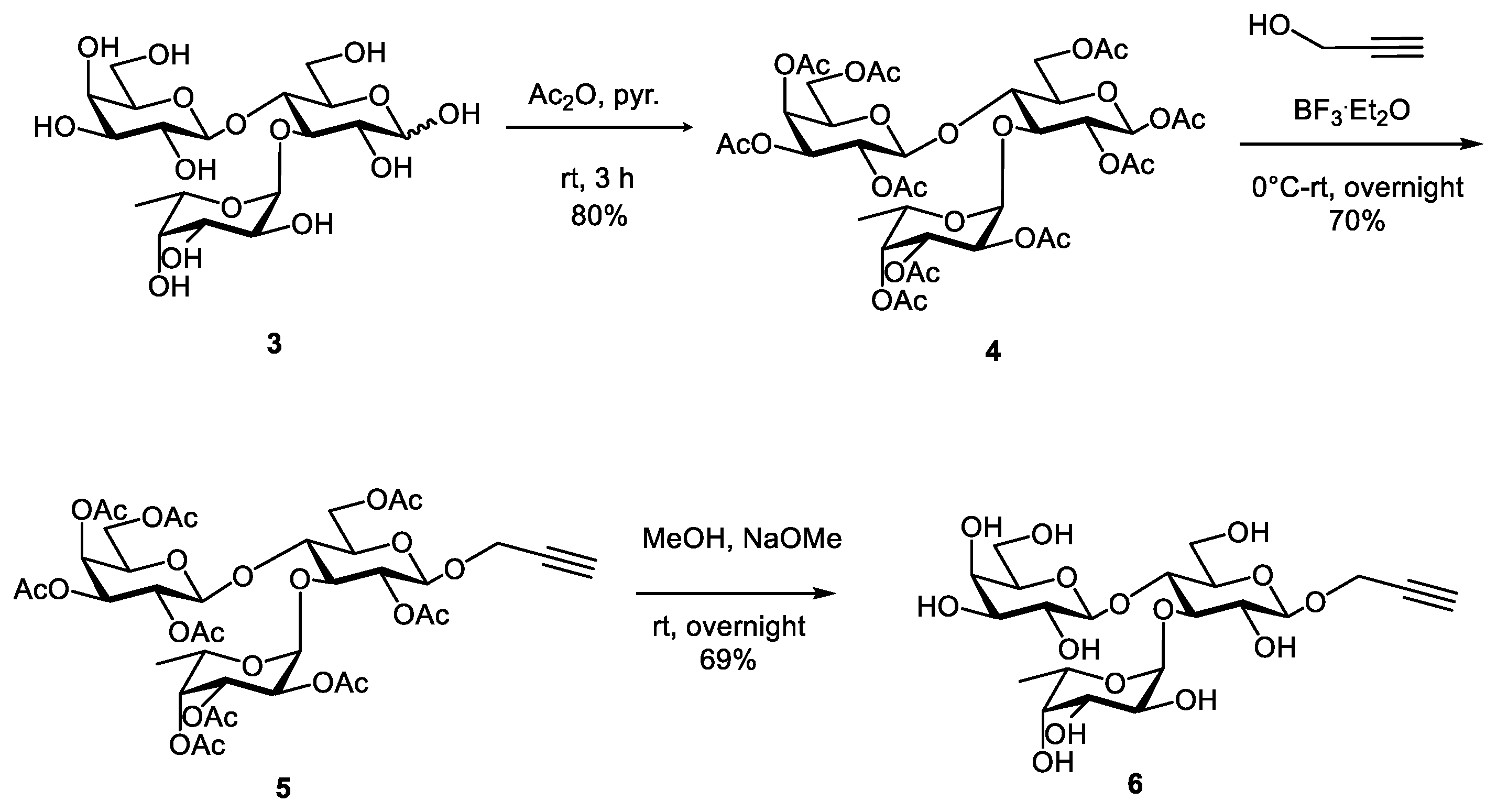
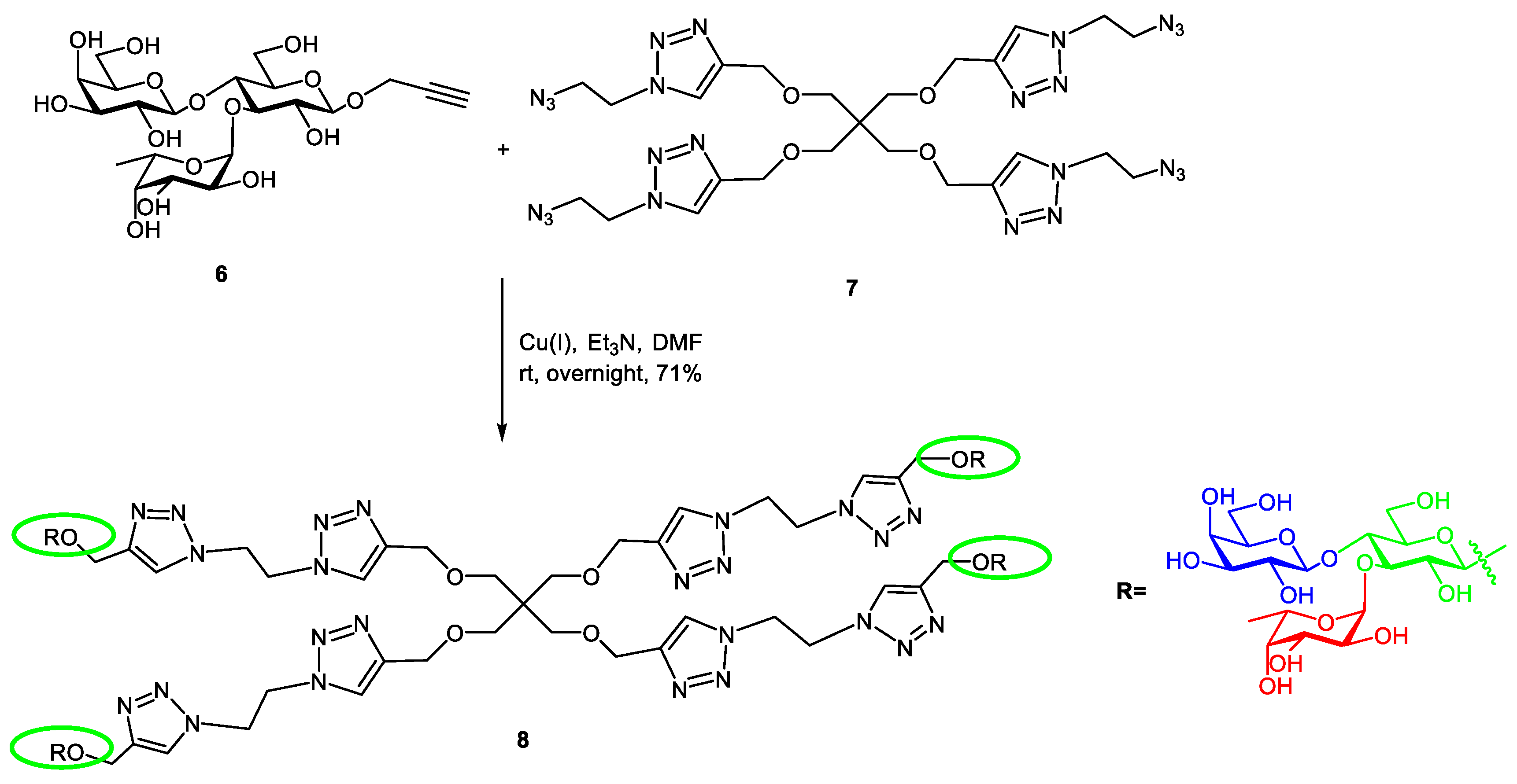
2.1. Synthesis of Tetravalent Fucosylated Lactoside
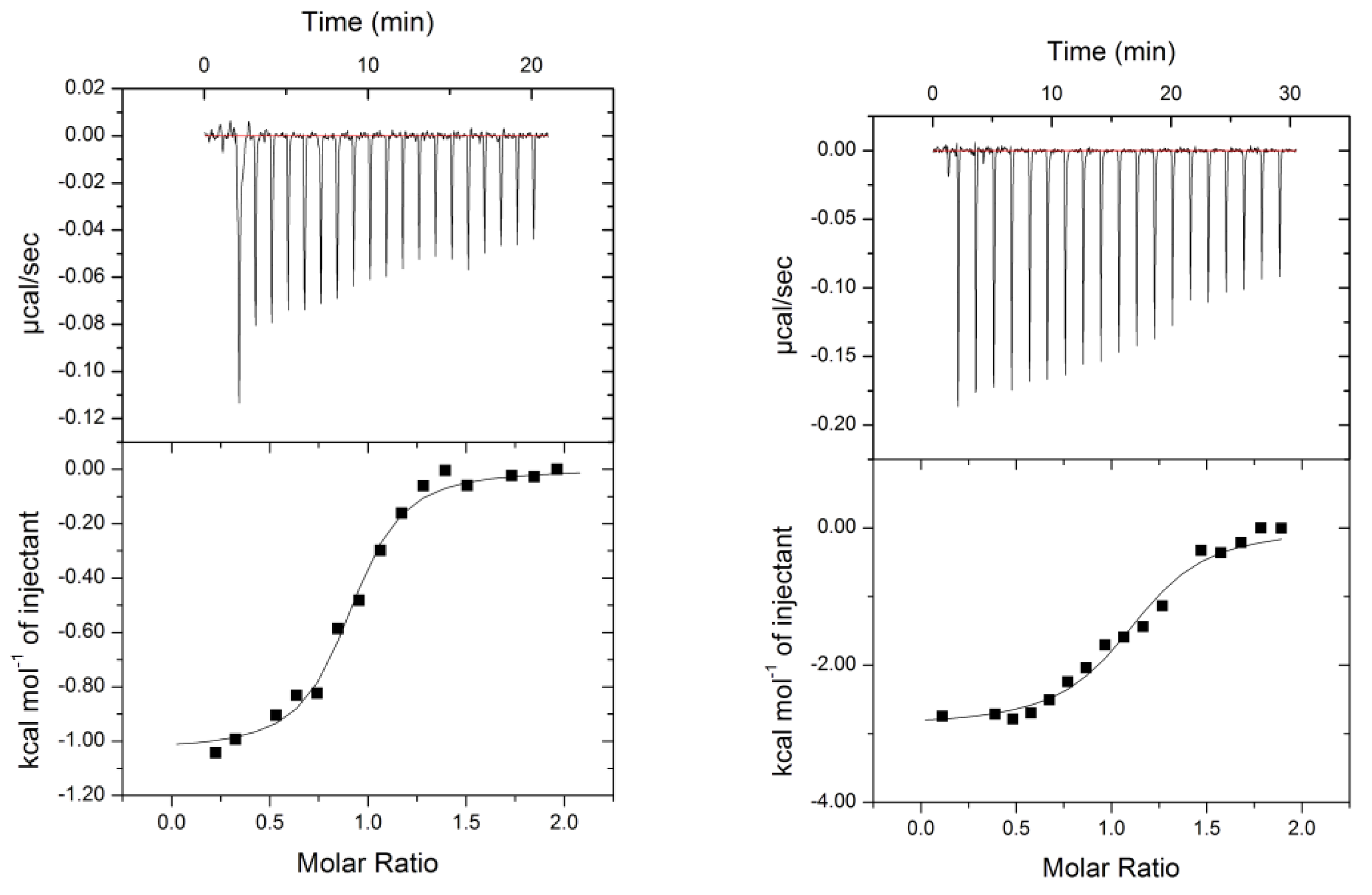
2.2. Thermodynamics of the Interactions
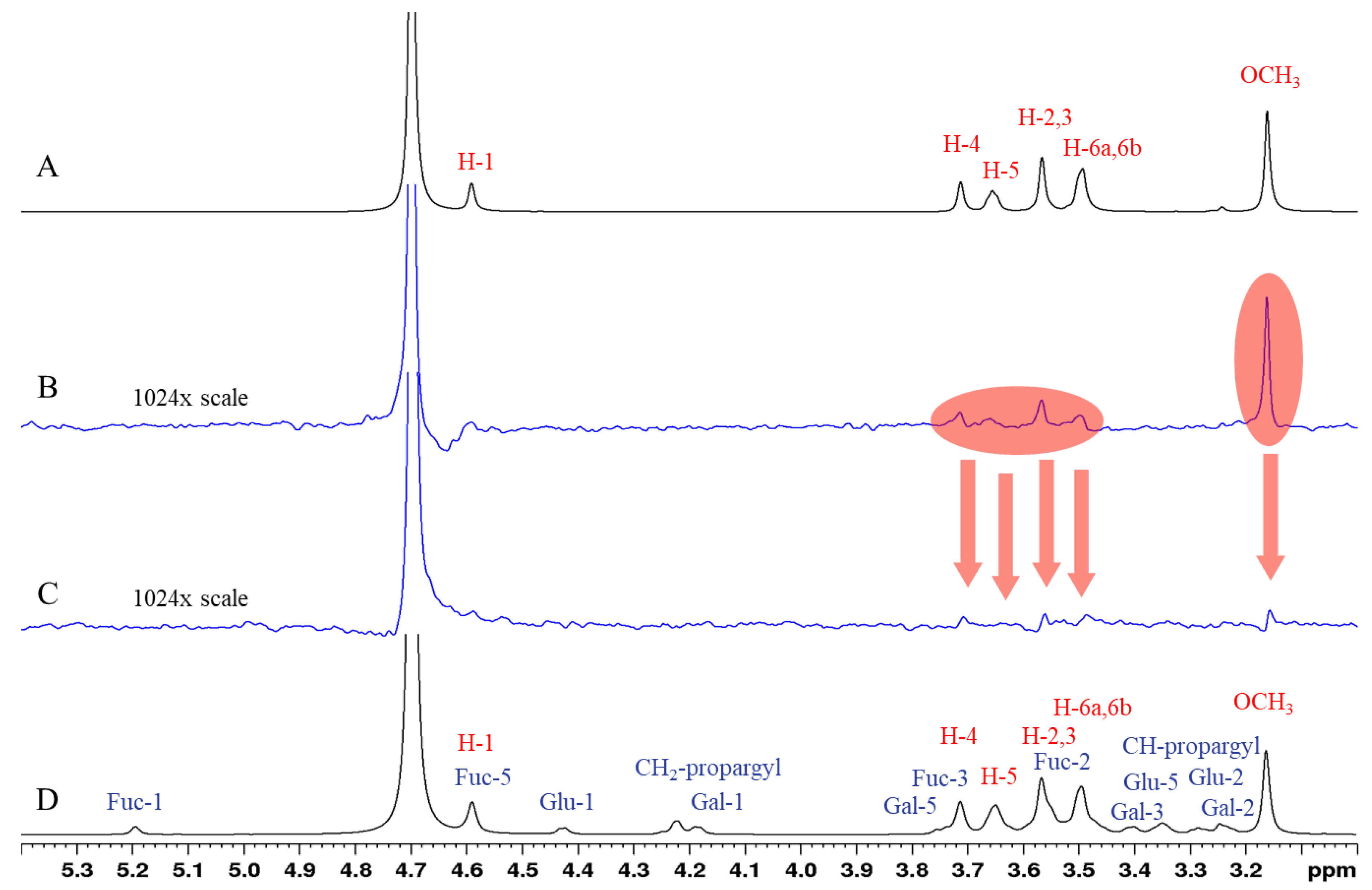
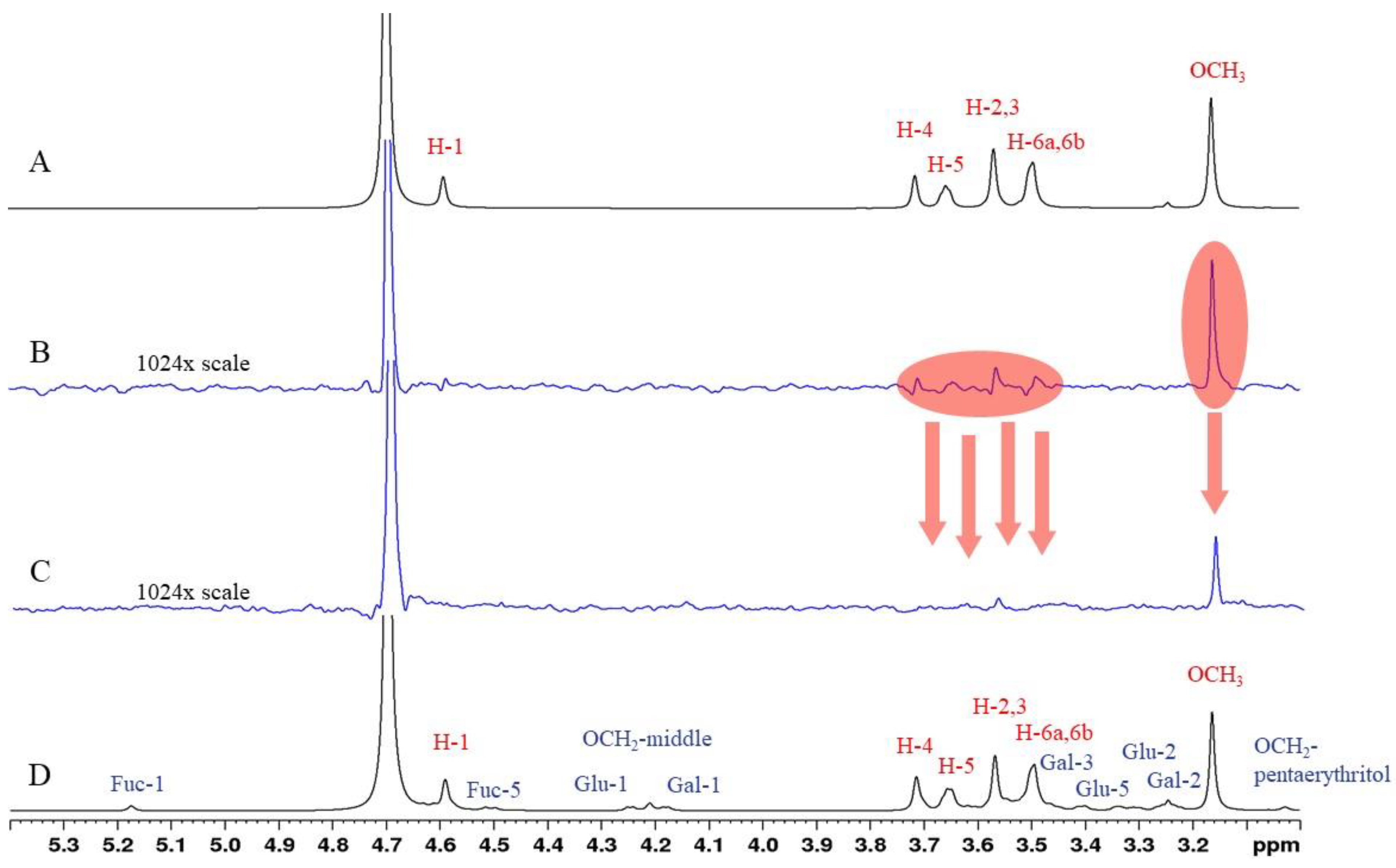
2.3. STD-NMR Studies: Binding of Compounds 6 and 8 to PA-IL and PA-IIL Lectins Characterized by 1H STD Competition NMR Experiments
3. Discussion
4. Materials and Methods
4.1. General Methods
4.1.1. Compound 4
4.1.2. Compound 5
4.1.3. Compound 6
4.1.4. Compound 8
4.2. Isothermal Microcalorimetry
4.3. 1H STD Competition NMR Experiments
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sharon, N.; Lis, H. Lectins: Carbohydrate-specific proteins that mediate cellular recognition. Chem. Rev. 1998, 98, 637–674. [Google Scholar] [CrossRef]
- Sharon, N. Carbohydrates as future anti-adhesion drugs for infectious diseases. Biochim. Biophys. Acta 2006, 1760, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Cecioni, S.; Imberty, A.; Vidal, S. Glycomimetics versus Multivalent Glycoconjugates for the Design of High Affinity Lectin Ligands. Chem. Rev. 2015, 115, 525–561. [Google Scholar] [CrossRef] [PubMed]
- Gibson, R.L.; Burns, J.L.; Ramsey, B.W. Pathophysiology and management of pulmonary infections in cystic fibrosis. Am. J. Respir. Crit. Care Med. 2003, 168, 918–951. [Google Scholar] [CrossRef]
- Folkesson, A.; Jelsbak, L.; Yang, L.; Johansen, H.K.; Ciofu, O.; Høiby, N.; Molin, S. Adaptation of Pseudomonas aeruginosa to the cystic fibrosis airway: An evolutionary perspective. Nat. Rev. Microbiol. 2012, 10, 841–851. [Google Scholar] [CrossRef]
- Gilboa-Garber, N. Inhibition of broad spectrum hemagglutinin from Pseudomonas aeruginosa by D-galactose and its derivatives. FEBS Lett. 1972, 20, 242–244. [Google Scholar] [CrossRef] [Green Version]
- Gilboa-Garber, N. Purification and properties of hemagglutinin from Pseudomonas aeruginosa and its reaction with human blood cells. Biochim. Biophys. Acta 1972, 273, 165–173. [Google Scholar] [CrossRef]
- Garber, N.C.; Guempel, V.; Guempel, N.; Gilboa-Garber, N.; Doyle, R.J. Specificity of the fucose-binding lectin of Pseudomonas aeruginosa. FEMS Microbiol. Lett. 1987, 48, 331–334. [Google Scholar] [CrossRef]
- Cioci, G.; Mitchell, E.P.; Gautier, C.; Wimmerová, M.; Sudakevitz, D.; Pérez, S.; Gilboa-Garber, N.; Imberty, A. Structural basis of calcium and galactose recognition by the lectin PA-IL of Pseudomonas aeruginosa. FEBS Lett. 2003, 555, 297–301. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, E.P.; Sabin, C.; Šnajdrová, L.; Pokorná, M.; Perret, S.; Gautier, C.; Hofr, C.; Gilboa-Garber, N.; Koča, J.; Wimmerová, M.; et al. High affinity fucose binding of Pseudomonas aeruginosa lectin PA-IIL: 1.0 A resolution crystal structure of the complex combined with thermodynamics and computational chemistry approaches. Proteins 2005, 58, 735–746. [Google Scholar] [CrossRef]
- Chemani, C.; Imberty, A.; de Bentzmann, S.; Pierre, M.; Wimmerová, M.; Guery, B.P.; Faure, K. Role of LecA and LecB lectins in Pseudomonas aeruginosa-induced lung injury and effect of carbohydrate ligands. Infect. Immun. 2009, 77, 2065–2075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diggle, S.P.; Stacey, R.E.; Dodd, C.; Cámara, M.; Williams, P.; Winzer, K. The galactophilic lectin, LecA, contributes to biofilm development in Pseudomonas aeruginosa. Environ. Microbiol. 2006, 8, 1095–1104. [Google Scholar] [CrossRef] [PubMed]
- Bajolet-Laudinat, O.; Girod-de Bentzmann, S.; Tournier, J.M.; Madoulet, C.; Plotkowski, M.C.; Chippaux, C.; Puchelle, E. Cytotoxicity of Pseudomonas aeruginosa internal lectin PA-I to respiratory epithelial cells in primary culture. Infect. Immun. 1994, 62, 4481–4487. [Google Scholar] [CrossRef] [Green Version]
- Novoa, A.; Eierhoff, T.; Topin, J.; Varrot, A.; Barluenga, S.; Imberty, A.; Römer, W.; Winssinger, N. A LecA Ligand Identified from a Galactoside-Conjugate Array Inhibits Host Cell Invasion by Pseudomonas aeruginosa. Angew. Chem. Int. Ed. 2014, 53, 8885–8889. [Google Scholar] [CrossRef] [PubMed]
- Cecioni, S.; Oerthel, V.; Iehl, J.; Holler, M.; Goyard, D.; Praly, J.P.; Imberty, A.; Nierengarten, J.F.; Vidal, S. Synthesis of dodecavalent fullerene-based glycoclusters and evaluation of their binding properties towards a bacterial lectin. Chemistry 2011, 17, 3252–3261. [Google Scholar] [CrossRef] [PubMed]
- Chabre, Y.M.; Giguère, D.; Blanchard, B.; Rodrigue, J.; Rocheleau, S.; Neault, M.; Rauthu, S.; Papadopoulos, A.; Arnold, A.A.; Imberty, A.; et al. Combining Glycomimetic and Multivalent Strategies toward Designing Potent Bacterial Lectin Inhibitors. Chem. Eur. J. 2011, 17, 6545–6562. [Google Scholar] [CrossRef]
- Kadam, R.U.; Bergmann, M.; Hurley, M.; Garg, D.; Cacciarini, M.; Swiderska, M.A.; Nativi, C.; Sattler, M.; Smyth, A.R.; Williams, P.; et al. A glycopeptide dendrimer inhibitor of the galactose-specific lectin LecA and of Pseudomonas aeruginosa biofilms. Angew. Chem. Int. Ed. 2011, 50, 10631–10635. [Google Scholar] [CrossRef] [Green Version]
- Soomro, Z.H.; Cecioni, S.; Blanchard, H.; Praly, J.P.; Imberty, A.; Vidal, S.; Matthews, S.E. CuAAC synthesis of resorcin[4]arene-based glycoclusters as multivalent ligands of lectins. Org. Biomol. Chem. 2011, 9, 6587–6597. [Google Scholar] [CrossRef]
- Cecioni, S.; Faure, S.; Darbost, U.; Bonnamour, I.; Parrot-Lopez, H.; Roy, O.; Taillefumier, C.; Wimmerová, M.; Praly, J.P.; Imberty, A.; et al. Selectivity among two lectins: Probing the effect of topology, multivalency and flexibility of “clicked” multivalent glycoclusters. Chem. Eur. J. 2011, 17, 2146–2159. [Google Scholar] [CrossRef]
- Otsuka, I.; Blanchard, B.; Borsali, R.; Imberty, A.; Kakuchi, T. Enhancement of plant and bacterial lectin binding affinities by three-dimensional organized cluster glycosides constructed on helical poly(phenylacetylene) backbones. ChemBioChem. 2010, 11, 2399–2408. [Google Scholar] [CrossRef]
- Kašaková, M.; Malinovská, L.; Klejch, T.; Hlaváčková, M.; Dvořáková, H.; Fujdiarová, E.; Rottnerová, Z.; Maťátková, O.; Lhoták, P.; Wimmerová, M.; et al. Selectivity of original C-hexopyranosyl calix[4]arene conjugates towards lectins of different origin. Carbohydr. Res. 2018, 469, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Palmioli, A.; Sperandeo, P.; Polissi, A.; Airoldi, C. Targeting bacterial biofilm: A new LecA multivalent ligand with inhibitory activity. ChemBioChem. 2019, 20, 2911–2915. [Google Scholar] [CrossRef]
- Flockton, T.R.; Schnorbus, L.; Araujo, A.; Adams, J.; Hammel, M.; Perez, L.J. Inhibition of Pseudomonas aeruginosa Biofilm Formation with Surface Modified Polymeric Nanoparticles. Pathogens 2019, 8, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Beshr, G.; Garvey, C.J.; Tabor, R.F.; Titz, A.; Wilkinson, B.L. Photoswitchable Janus glycodendrimer micelles as multivalent inhibitors of LecA and LecB from Pseudomonas aeruginosa. Colloids Surf. B Biointerfaces 2017, 159, 605–612. [Google Scholar] [CrossRef] [PubMed]
- Sommer, R.; Wagner, S.; Rox, K.; Varrot, A.; Hauck, D.; Wamhoff, E.C.; Schreiber, J.; Ryckmans, T.; Brunner, T.; Rademacher, C.; et al. Glycomimetic, orally bioavailable LecB inhibitors block biofilm formation of Pseudomonas aeruginosa. J. Am. Chem. Soc. 2018, 140, 2537–2545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauck, D.; Joachim, I.; Frommeyer, B.; Varrot, A.; Philipp, B.; Möller, H.M.; Imberty, A.; Exner, T.E.; Titz, A. Discovery of two classes of potent glycomimetic inhibitors of Pseudomonas aeruginosa LecB with distinct binding modes. ACS Chem. Biol. 2013, 8, 1775–1784. [Google Scholar] [CrossRef]
- Buffet, K.; Gillon, E.; Holler, M.; Nierengarten, J.F.; Imberty, A.; Vincent, S.P. Fucofullerenes as tight ligands of RSL and LecB, two bacterial lectins. Org. Biomol. Chem. 2015, 13, 6482–6492. [Google Scholar] [CrossRef]
- Reymond, J.L.; Bergmann, M.; Darbre, T. Glycopeptide dendrimers as Pseudomonas aeruginosa biofilm inhibitors. Chem. Soc. Rev. 2013, 42, 4814–4822. [Google Scholar] [CrossRef] [Green Version]
- Galanos, N.; Gillon, E.; Imberty, A.; Matthews, S.E.; Vidal, S. Pentavalent pillar[5]arene-based glycoclusters and their multivalent binding to pathogenic bacterial lectins. Org. Biomol. Chem. 2016, 14, 3476–3481. [Google Scholar] [CrossRef]
- Donnier-Maréchal, M.; Galanos, N.; Grandjean, T.; Pascal, Y.; Ji, D.; Dong, L.; Gillon, E.; He, X.; Imberty, A.; Kipnis, E.; et al. Perylenediimide-based glycoclusters as high affinity ligands of bacterial lectins: Synthesis, binding studies and anti-adhesive properties. Org. Biomol. Chem. 2017, 15, 10037–10043. [Google Scholar] [CrossRef]
- Michaud, G.; Visini, R.; Bergmann, M.; Salerno, G.; Bosco, R.; Gillon, E.; Richichi, B.; Nativi, C.; Imberty, A.; Stocker, A.; et al. Overcoming antibiotic resistance in Pseudomonas aeruginosa biofilms using glycopeptide dendrimers. Chem. Sci. 2016, 7, 166–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goyard, D.; Thomas, B.; Gillon, E.; Imberty, A.; Renauder, O. Heteroglycoclusters With Dual Nanomolar Affinities for the Lectins LecA and LecB From Pseudomonas aeruginosa. Front. Chem. 2019, 7, 666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Guo, Q.; Dai, X.; Wei, X.; Yu, Y.; Chen, X.; Li, C.; Cao, Z.; Zhang, X. A Biomimetic Non-Antibiotic Approach to Eradicate Drug-Resistant Infections. Adv. Mater. 2019, 31, e1806024. [Google Scholar] [CrossRef] [PubMed]
- Vincent, S.P.; Buffet, K.; Nierengarten, I.; Imberty, A.; Nierengarten, J.F. Biologically active heteroglycoclusters constructed on a pillar [5] arene-containing [2] rotaxane scaffold. Chem. Eur. J. 2016, 22, 88–92. [Google Scholar] [CrossRef] [Green Version]
- Gerland, B.; Goudot, A.; Pourceau, G.; Meyer, A.; Vidal, S.; Souteyrand, E.; Vasseur, J.-J.; Chevolot, Y.; Morvan, F. Synthesis of homo-and heterofunctionalized glycoclusters and binding to Pseudomonas aeruginosa lectins PA-IL and PA-IIL. J. Org. Chem. 2012, 77, 7620–7626. [Google Scholar] [CrossRef]
- Deguise, I.; Lagnoux, D.; Roy, R. Synthesis of glycodendrimers containing both fucoside and galactoside residues and their binding properties to PA-IL and PA-IIL lectins from Pseudomonas aeruginosa. New J. Chem. 2007, 31, 1321–1331. [Google Scholar] [CrossRef]
- Malinovská, L.; Thai, L.S.; Herczeg, M.; Vašková, M.; Houser, J.; Fujdiarová, E.; Komárek, J.; Hodek, P.; Borbás, A.; Wimmerová, M.; et al. Synthesis of β-D-galactopyranoside-presenting glycoclusters, investigation of their interactions with Pseudomonas aeruginosa lectin A (PA-IL) and evaluation of their anti-adhesion potential. Biomolecules 2019, 9, 686. [Google Scholar] [CrossRef] [Green Version]
- Illyés, T.Z.; Malinovská, L.; Rőth, E.; Tóth, B.; Farkas, B.; Korsák, M.; Wimmerová, M.; Kövér, K.E.; Csávás, M. Synthesis of Tetravalent Thio- and Selenogalactoside-Presenting Galactoclusters and Their Interactions with Bacterial Lectin PA-IL from Pseudomonas aeruginosa. Molecules 2021, 26, 542. [Google Scholar] [CrossRef]
- Thai, L.S.; Malinovska, L.; Vašková, M.; Mező, E.; Kelemen, V.; Borbás, A.; Hodek, P.; Wimmerová, M.; Csávás, M. Investigation of the Binding Affinity of a Broad Array of l-Fucosides with Six Fucose-Specific Lectins of Bacterial and Fungal Origin. Molecules 2019, 24, 2262. [Google Scholar] [CrossRef] [Green Version]
- Jančaříková, G.; Herczeg, M.; Fujdiarová, E.; Houser, J.; Kövér, K.E.; Borbás, A.; Wimmerová, M.; Csávás, M. Synthesis of α-L-fucopyranoside-presenting glycoclusters and investigation of their interaction with recombinant Photorhabdus asymbiotica lectin (PHL). Chem. Eur. J. 2018, 24, 4055–4068. [Google Scholar] [CrossRef]
- Matwiejuk, M.; Dékány, G. Crystalline 3-O-fucosyllactose and manufacturing method. WO 2013-DK50300.
- Cecioni, S.; Praly, J.P.; Matthews, S.E.; Wimmerová, M.; Imberty, A.; Vidal, S. Rational design and synthesis of optimized glycoclusters for multivalent lectin-carbohydrate interactions: Influence of the linker arm. Chem. Eur. J. 2012, 18, 6250–6263. [Google Scholar] [CrossRef] [PubMed]
- Sabin, C.; Mitchell, E.P.; Pokorná, M.; Gautier, C.; Utille, J.P.; Wimmerová, M.; Imberty, A. Binding of different monosaccharides by lectin PA-IIL from Pseudomonas aeruginosa: Thermodynamics data correlated with X-ray structures. FEBS Lett. 2006, 580, 982–987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, M.; Meyer, B. Characterization of Ligand Binding by Saturation Transfer Difference NMR Spectroscopy. Angew. Chem. Int. Ed. 1999, 38, 1784–1788. [Google Scholar] [CrossRef]
- Mayer, M.; Meyer, B. Group Epitope Mapping by Saturation Transfer Difference NMR to Identify Segments of a Ligand in Direct Contact with a Protein Receptor. J. Am. Chem. Soc. 2001, 123, 6108–6117. [Google Scholar] [CrossRef]
- Groves, P.; Kövér, K.E.; André, S.; Bandorowicz-Pikuła, J.; Batta, G.; Bruix, M.; Buchet, R.; Canales, Á.; Cañada, F.J.; Gabius, H.-J.; et al. Temperature dependence of ligand-protein complex formation as reflected by saturation transfer difference NMR experiments. J. Magn. Reson. Chem. 2007, 45, 745–748. [Google Scholar] [CrossRef] [Green Version]
- Adam, J.; Pokorná, M.; Sabin, C.; Mitchell, E.P.; Imberty, A.; Wimmerová, M. Engineering of PA-IIL lectin from Pseudomonas aeruginosa—Unravelling the role of the specificity loop for sugar preference. BMC Struct. Biol. 2007, 7, 36. [Google Scholar] [CrossRef] [Green Version]
- Hauber, H.P.; Schulz, M.; Pforte, A.; Mack, D.; Zabel, P.; Schumacher, U. Inhalation with fucose and galactose for treatment of Pseudomonas aeruginosa in cystic fibrosis patients. Int. J. Med. Sci. 2008, 5, 371–376. [Google Scholar] [CrossRef] [Green Version]
- Kubíčková, B.; Hadrabová, J.; Vašková, L.; Mandys, V.; Stiborová, M.; Hodek, P. Susceptibility of airways to Pseudomonas aeruginosa infection: Mouse neuraminidase model. Monatsh. Chem. 2017, 148, 1993–2002. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inhibitor | Valency | n | Ka [104 M−1] |
|---|---|---|---|
| d-galactose | 1 | 0.78 ± 0.05 | 0.68 ± 0.038 [37] |
| Me β−d-Gal | 1 | 1.4 [42] | |
| Compound 6 | 1 | 0.51 ± 0.08 | 59 ± 28.4 |
| Compound 1 | 4 | 0.21 ± 0.01 | 470 ± 54.5 [37] |
| Compound 8 | 4 | 0.89 ± 0.02 | 496 ± 104 |
| Inhibitor | ΔH [kJ/mol] | −TΔS [kJ/mol] | ΔG [kJ/mol] |
| d-galactose | −37.8 | 15.9 | −21.9 |
| Me β−d-Gal | −39.0 | 15.0 | −24.0 |
| Compound 6 | −3.48 | −29.3 | −32.8 |
| Compound 1 | −134 | 95.7 | −38.1 |
| Compound 8 | −4.3 | −33.8 | −38.1 |
| Inhibitor | Valency | n | Ka [104 M−1] |
|---|---|---|---|
| l-fucose | 1 | 0.96 ± 0.04 | 12 ± 1 [43] |
| Me α-L-Fuc | 1 | 0.43± 0.01 | 235 ± 8.0 [43] |
| Compound 6 | 1 | 1.12 ± 0.05 | 344 ± 124 |
| Compound 2 | 4 | 2.1 ± 0.1 | 310 ± 100 [37] |
| Compound 8 | 4 | 1.1 ± 0.03 | 257 ± 66.9 |
| Inhibitor | ΔH [kJ/mol] | −TΔS [kJ/mol] | ΔG [kJ/mol] |
| l-fucose | −27.7 | −1.3 | −29 |
| Me α-L-Fuc | −41.3 | 4.9 | −36.4 |
| Compound 6 | −36.3 | −0.8 | −37.1 |
| Compound 2 | −56.9 | −25.5 | −31.4 |
| Compound 8 | −12.0 | −24.3 | −36.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Csávás, M.; Kalmár, L.; Szőke, P.; Farkas, L.B.; Bécsi, B.; Kónya, Z.; Kerékgyártó, J.; Borbás, A.; Erdődi, F.; Kövér, K.E. A Fucosylated Lactose-Presenting Tetravalent Glycocluster Acting as a Mutual Ligand of Pseudomonas aeruginosa Lectins A (PA-IL) and B (PA-IIL)—Synthesis and Interaction Studies. Int. J. Mol. Sci. 2022, 23, 16194. https://doi.org/10.3390/ijms232416194
Csávás M, Kalmár L, Szőke P, Farkas LB, Bécsi B, Kónya Z, Kerékgyártó J, Borbás A, Erdődi F, Kövér KE. A Fucosylated Lactose-Presenting Tetravalent Glycocluster Acting as a Mutual Ligand of Pseudomonas aeruginosa Lectins A (PA-IL) and B (PA-IIL)—Synthesis and Interaction Studies. International Journal of Molecular Sciences. 2022; 23(24):16194. https://doi.org/10.3390/ijms232416194
Chicago/Turabian StyleCsávás, Magdolna, László Kalmár, Petronella Szőke, László Bence Farkas, Bálint Bécsi, Zoltán Kónya, János Kerékgyártó, Anikó Borbás, Ferenc Erdődi, and Katalin E. Kövér. 2022. "A Fucosylated Lactose-Presenting Tetravalent Glycocluster Acting as a Mutual Ligand of Pseudomonas aeruginosa Lectins A (PA-IL) and B (PA-IIL)—Synthesis and Interaction Studies" International Journal of Molecular Sciences 23, no. 24: 16194. https://doi.org/10.3390/ijms232416194