

Gonadal Transcriptome Analysis and Sequence Characterization of Sex-Related Genes in Cranoglanis bouderius
Abstract
:1. Introduction
2. Results
2.1. Sequencing Quality Assessment
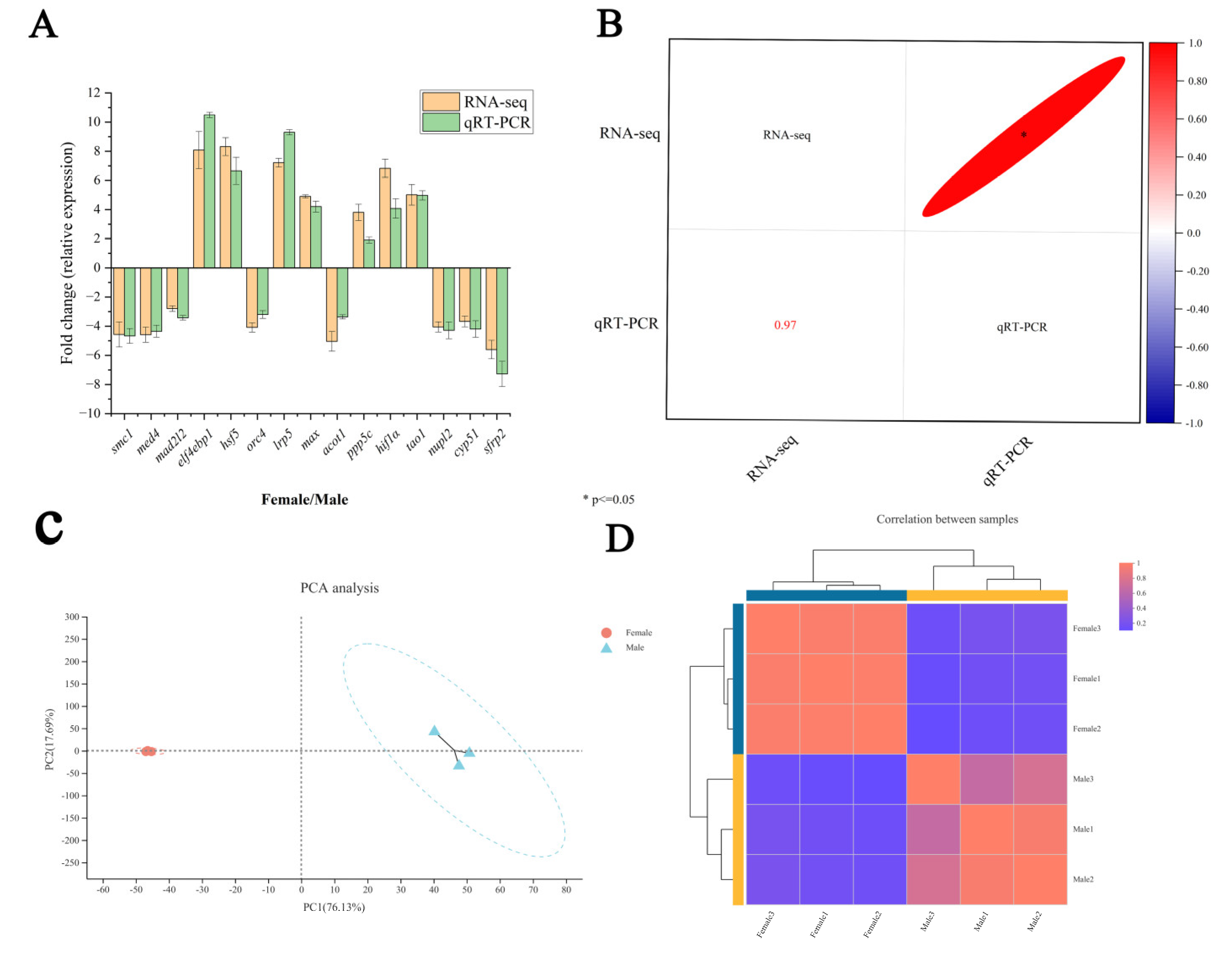
2.2. Transcriptome Data Validation
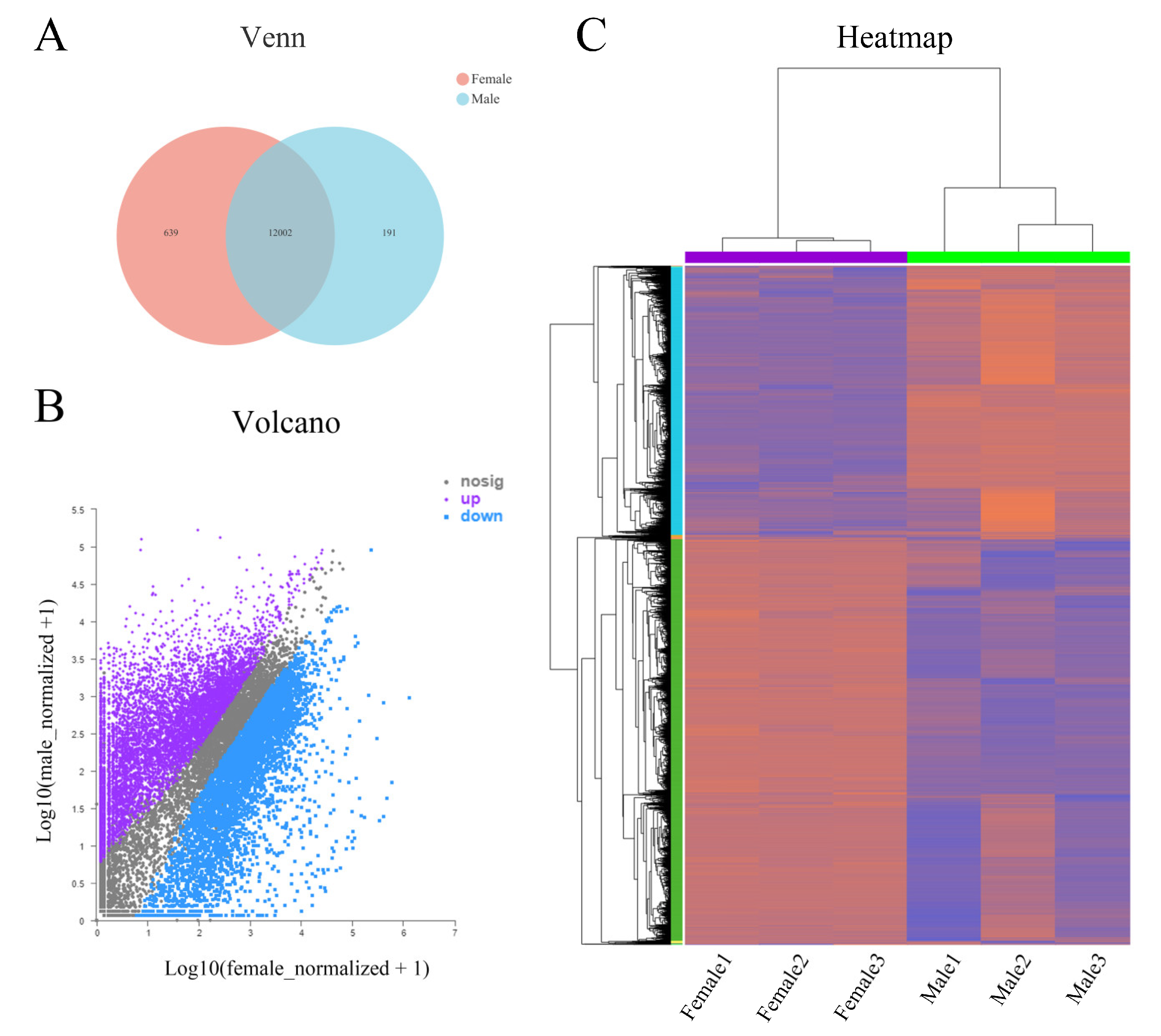
2.3. Analysis of Differentially Expressed Genes (DEGs)
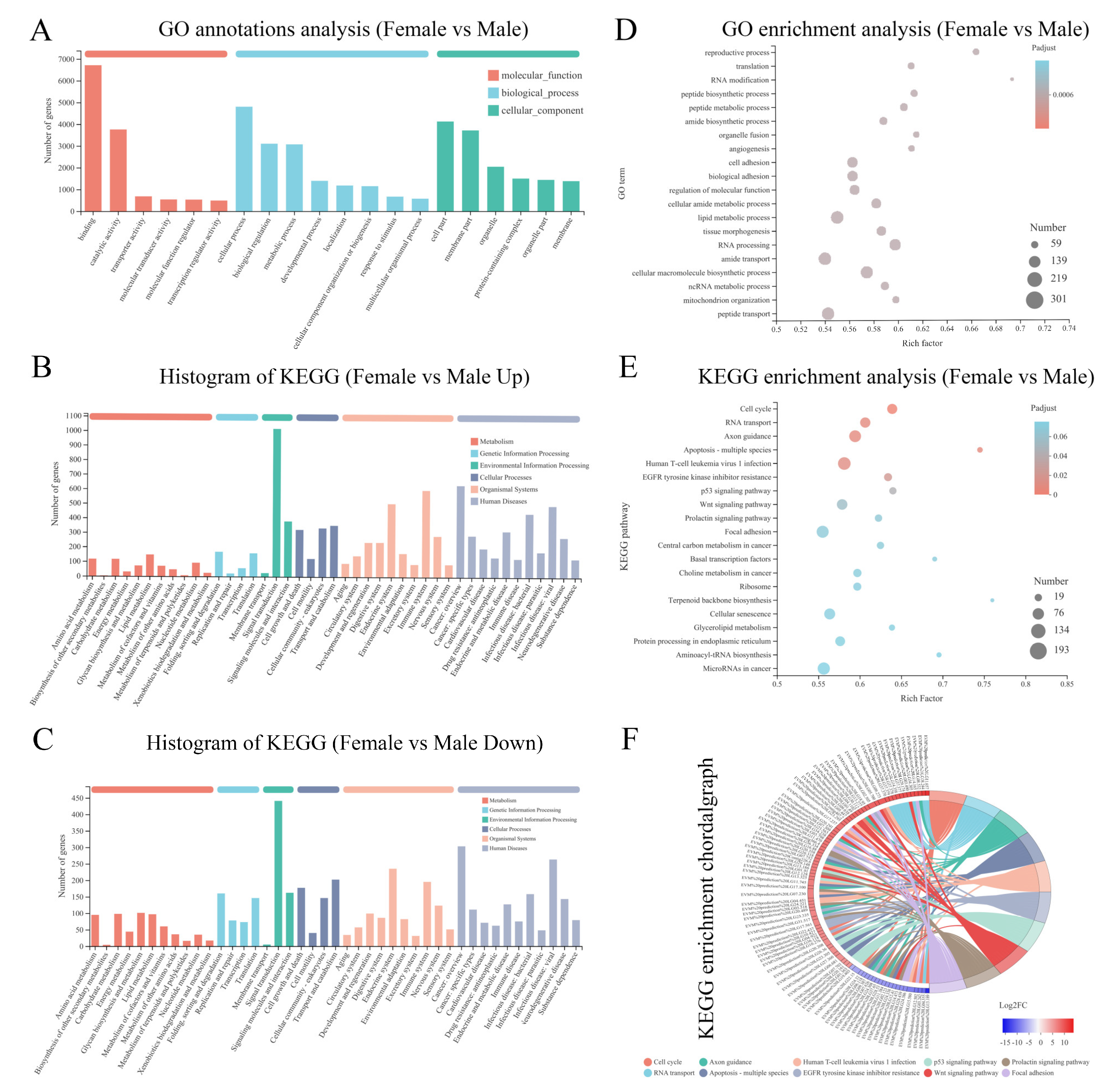
2.4. Identification of Sex-Related DEGs
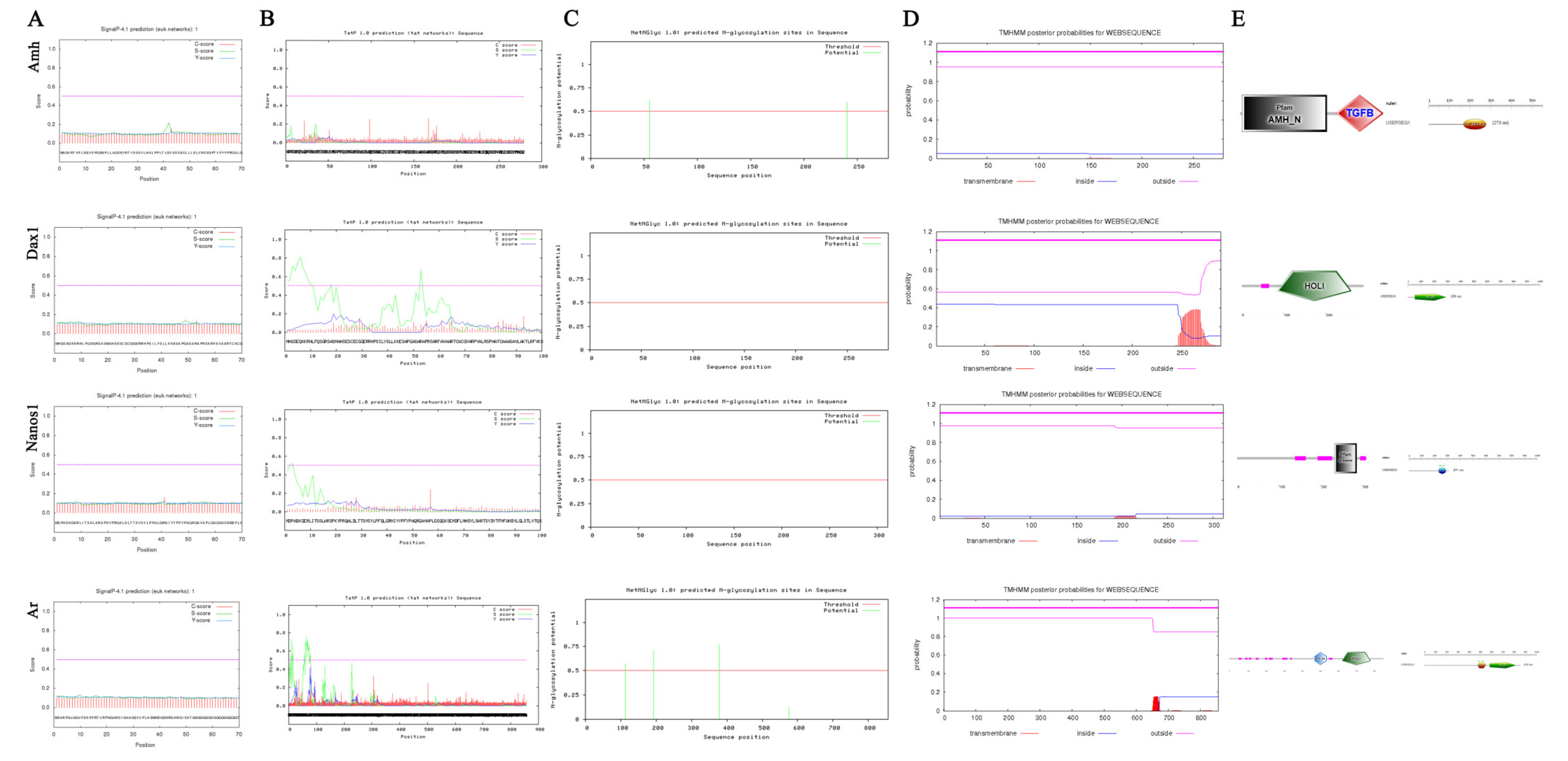
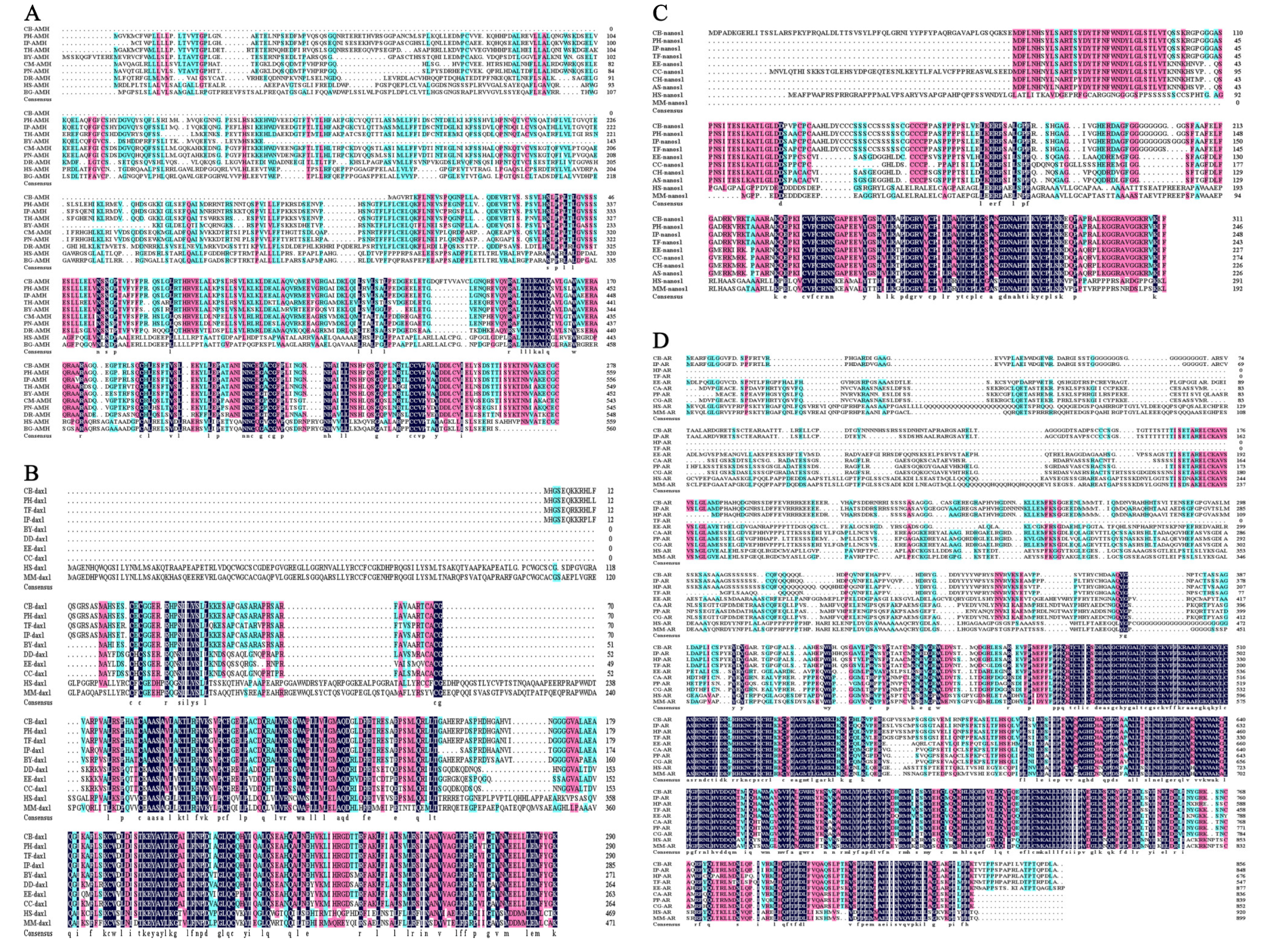
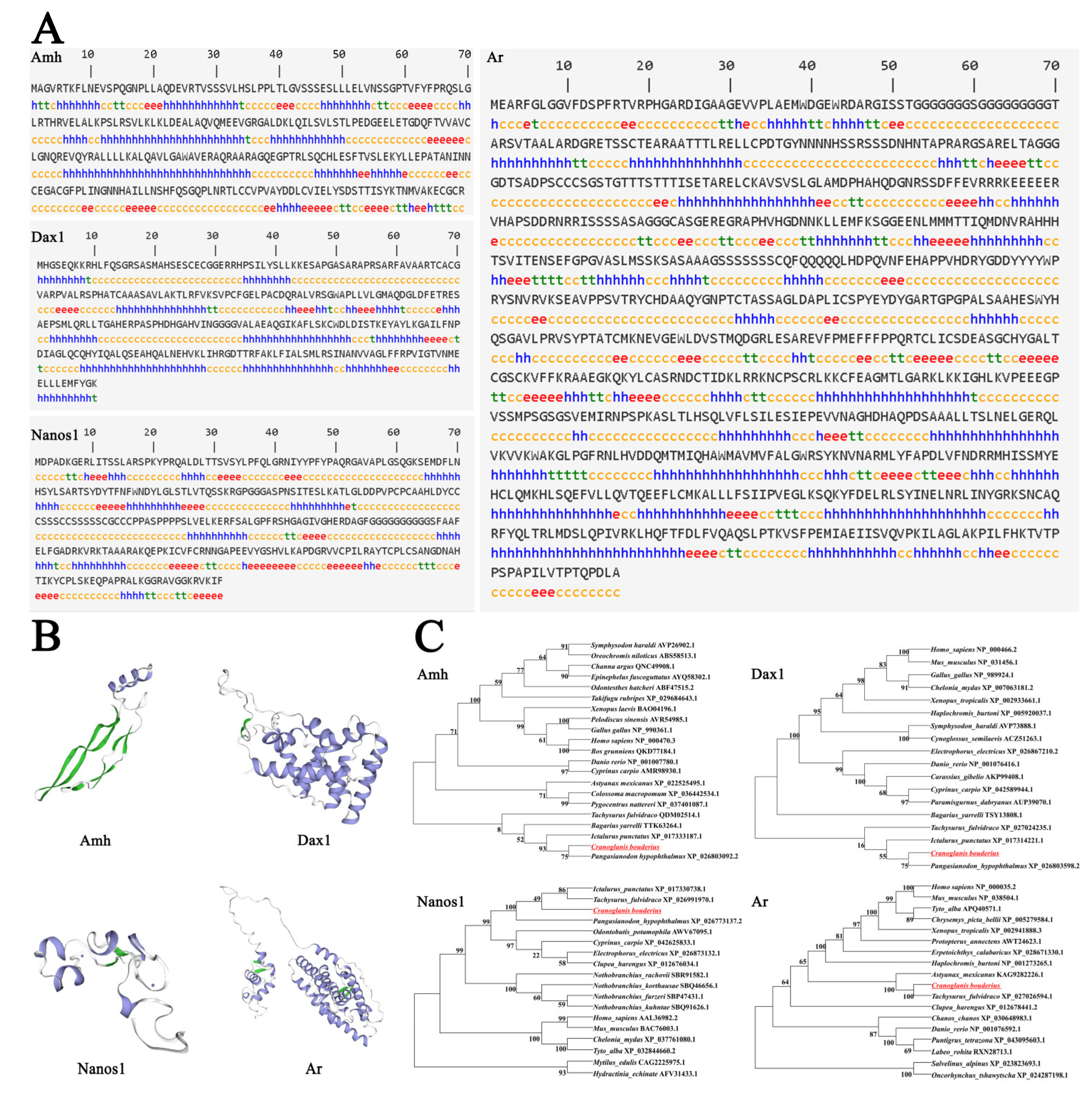
2.5. Sequence Characterization of Sex-Related Genes
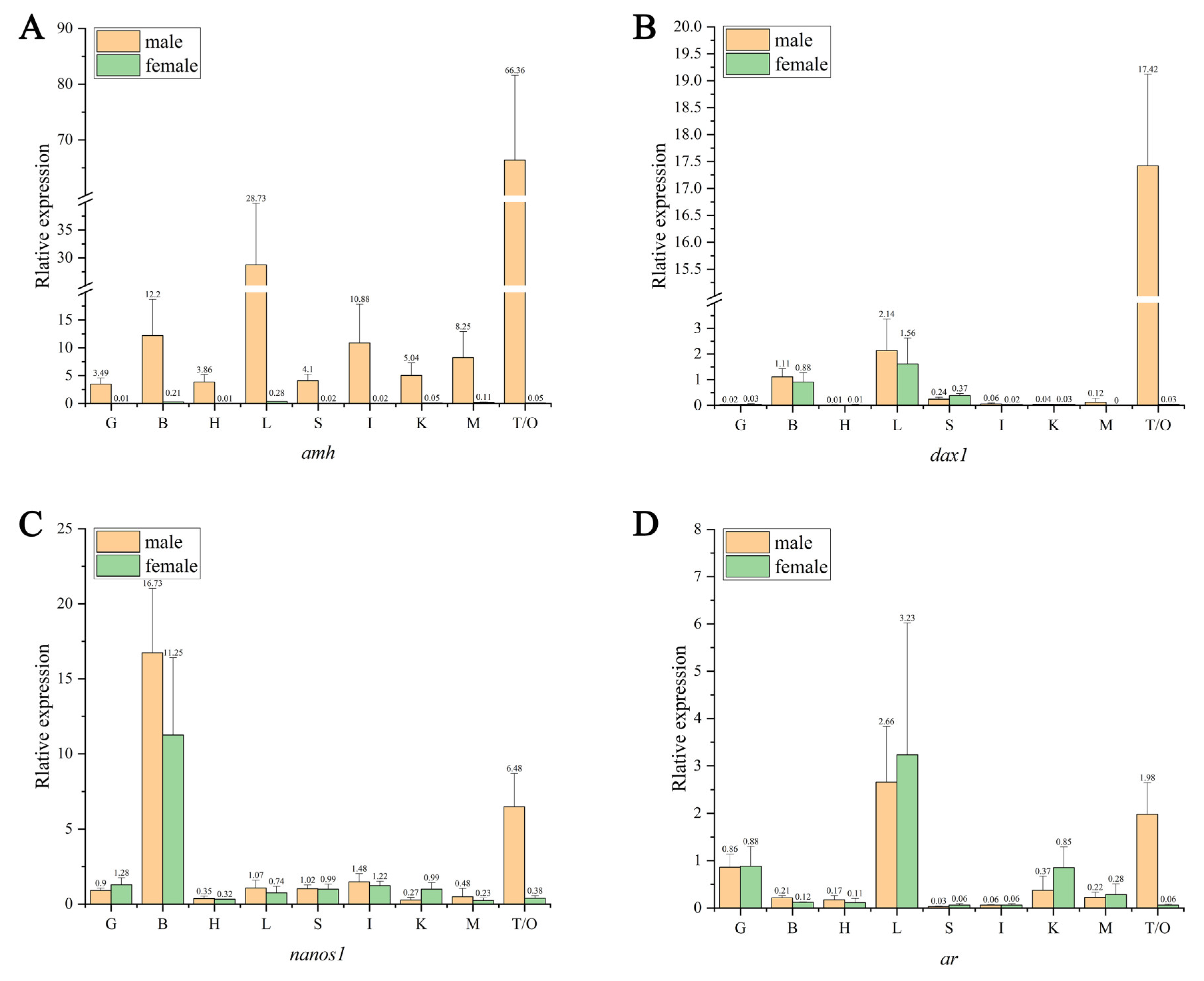
2.6. Sex-Related Gene Tissue Expression
3. Discussion
4. Materials and Methods
4.1. Experimental Fish and Sample Collection
4.2. RNA Extraction, Library Building and Sequencing
4.3. Transcriptome Data Validation
4.4. Identification and Functional Annotation of Differentially Expressed Genes (DEGs)
4.5. Sequence Characterization of Sex-Related Gene
4.6. Sex-Related Gene Tissue Expression Analysis
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Strussmann, C.A.; Yamamoto, Y.; Hattori, R.S.; Fernandino, J.I.; Somoza, G.M. Where the Ends Meet: An Overview of Sex Determination in Atheriniform Fishes. Sex. Dev. 2021, 15, 80–92. [Google Scholar] [CrossRef] [PubMed]
- Bokony, V.; Milne, G.; Pipoly, I.; Szekely, T.; Liker, A. Sex ratios and bimaturism differ between temperature-dependent and genetic sex-determination systems in reptiles. BMC Evol. Biol. 2019, 19, 57. [Google Scholar] [CrossRef] [PubMed]
- Muralidhar, P.; Veller, C. Sexual antagonism and the instability of environmental sex determination. Nat. Ecol. Evol. 2018, 2, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M. Is a Sex-Determining Gene(s) Necessary for Sex-Determination in Amphibians? Steroid Hormones May Be the Key Factor. Sex. Dev. 2013, 7, 104–114. [Google Scholar] [CrossRef]
- Wiggins, J.M.; Santoyo-Brito, E.; Scales, J.B.; Fox, S.F. Gene Dose Indicates Presence of Sex Chromosomes in Collared Lizards (Crotaphytus collaris), a Species with Temperature-Influenced Sex Determination. Herpetologica 2020, 76, 27–30. [Google Scholar] [CrossRef]
- Corona-Herrera, G.A.; Arranz, S.E.; Martinez-Palacios, C.A.; Navarrete-Ramirez, P.; Toledo-Cuevas, E.M.; Valdez-Alarcon, J.J.; Martinez-Chavez, C.C. Experimental evidence of masculinization by continuous illumination in a temperature sex determination teleost (Atherinopsidae) model: Is oxidative stress involved? J. Fish Biol. 2018, 93, 229–237. [Google Scholar] [CrossRef]
- Kanaiwa, M.; Harada, Y. Collapse of one-locus two-allele sex determining system by releasing sex-reversed hatchery fish. Rev. Fish. Sci. 2008, 16, 296–302. [Google Scholar] [CrossRef]
- Andreata, A.A.; do Almeida-Toledo, L.; Oliveira, C.; de Toledo Filho, S. Chromosome studies in hypoptopomatinae (Pisces, Siluriformes, Loricariidae). II. ZZ/ZW sex-chromosome system, B chromosomes, and constitutive heterochromatin differentiation in Microlepidogaster leucofrenatus. Cytogenet. Cell Genet. 1993, 63, 215–220. [Google Scholar] [CrossRef]
- Nakamura, D.; Wachtel, S.S.; Kallman, K. H-Y antigen and the evolution of heterogamety. J. Hered. 1984, 75, 353–358. [Google Scholar] [CrossRef]
- Tave, D. Genetics for Fish Hatchery Managers; AVI Publishing Co., Inc.: Westport, CT, USA, 1986. [Google Scholar]
- Han, C.; Zhu, Q.Y.; Zhou, X.N.; Ouyang, H.F.; Han, L.Q.; Chen, J.H.; Li, S.S.; Li, G.F.; Lin, H.R.; Zhang, Y. A PCR-based genetic sex identification method in spotted mandarin fish (Siniperca scherzeri) and big eye mandarin fish (Siniperca kneri). Aquac. Rep. 2020, 18, 100552. [Google Scholar] [CrossRef]
- Liu, X.Y.; Dai, S.F.; Wu, J.H.; Wei, X.Y.; Zhou, X.; Chen, M.M.; Tan, D.J.; Pu, D.Y.; Li, M.H.; Wang, D.S. Roles of anti-Mullerian hormone and its duplicates in sex determination and germ cell proliferation of Nile tilapia. Genetics 2022, 220, iyab237. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.L.; Zhao, M.; Wang, L.; Yu, Z.S.; Wang, J.; Yu, Q.; Xiao, L.; Lu, M.W.; Li, S.S.; Zhang, Y.; et al. Overexpression of Anti-mullerian Hormone Gene in vivo Affects Gonad Sex Differentiation in Undifferentiated Orange-Spotted Groupers (Epinephelus coioides). Front. Endocrinol. 2019, 10, 210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, J.; Li, J.; Hu, J.; Sun, C.; Tian, Y.; Li, W.; Yan, N.; Sun, C.; Sheng, X.; Yang, S.; et al. Comparative Genomics Studies on the dmrt Gene Family in Fish. Front. Genet. 2020, 11, 563947. [Google Scholar] [CrossRef] [PubMed]
- Farhadi, A.; Fang, S.; Zhang, Y.; Cui, W.; Fang, H.; Ikhwanuddin, M.; Ma, H. The significant sex-biased expression pattern of Sp-Wnt4 provides novel insights into the ovarian development of mud crab (Scylla Paramamosain). Int. J. Biol. Macromol. 2021, 183, 490–501. [Google Scholar] [CrossRef]
- Xia, X.; Huo, W.; Wan, R.; Wang, P.; Chang, Z. Cloning, characterization and function analysis of DAX1 in Chinese loach (Paramisgurnus dabryanus). Genetica 2018, 146, 487–496. [Google Scholar] [CrossRef]
- Tan, X.G.; Sui, Y.L.; Li, M.J.; Jiao, S.; Wu, Z.H.; You, F. Characterization of nanos1 Homolog in the Olive Flounder, Paralichthys olivaceus (Temminck & Schlegel, 1846). Turk. J. Fish Quat. Sci. 2020, 20, 421–429. [Google Scholar] [CrossRef]
- Golshan, M.; Alavi, S.M.H. Androgen signaling in male fishes: Examples of anti-androgenic chemicals that cause reproductive disorders. Theriogenology 2019, 139, 58–71. [Google Scholar] [CrossRef]
- Hattori, R.S.; Murai, Y.; Oura, M.; Masuda, S.; Majhi, S.K.; Sakamoto, T.; Fernandino, J.I.; Somoza, G.M.; Yokota, M.; Strussmann, C.A. A Y-linked anti-Mullerian hormone duplication takes over a critical role in sex determination. Proc. Natl. Acad. Sci. USA 2012, 109, 2955–2959. [Google Scholar] [CrossRef] [Green Version]
- Kurokawa, H.; Saito, D.; Nakamura, S.; Katoh-Fukui, Y.; Ohta, K.; Baba, T.; Morohashi, K.I.; Tanaka, M. Germ cells are essential for sexual dimorphism in the medaka gonad. Proc. Natl. Acad. Sci. USA 2007, 104, 16958–16963. [Google Scholar] [CrossRef] [Green Version]
- Koprunner, M.; Thisse, C.; Thisse, B.; Raz, E. A zebrafish nanos-related gene is essential for the development of primordial germ cells. Genes Dev. 2001, 15, 2877–2885. [Google Scholar] [CrossRef]
- Black, B.E.; Paschal, B.M. Intranuclear organization and function of the androgen receptor. Trends Endocrinol. Metab. 2004, 15, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Liu, C.X.; He, S.P. Threatened fishes of the world: Cranoglanis bouderius (Richardson, 1846) (Cranoglanididae). Environ. Biol. Fishes 2009, 84, 157–158. [Google Scholar] [CrossRef] [Green Version]
- Diogo, R.; Chardon, M.; Vandewalle, P. Osteology and myology of the cephalic region and pectoral girdle of the Chinese catfish Cranoglanis bouderius, with a discussion on the autapomorphies and phylogenetic relationships of the Cranoglanididae (Teleostei:Siluriformes). J. Morphol. 2002, 253, 229–242. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.G.; Wang, J.; He, S.P. The complete mitochondrial genome of the helmet catfish Cranoglanis bouderius (Silurifonnes: Cranoglanididae) and the phylogeny of otophysan fishes. Gene 2006, 376, 290–297. [Google Scholar] [CrossRef]
- Yue, P.; Chen, Y. Pisces. In China Red Data Book of Endangered Animals; Wang, S., Ed.; Science Press: Beijing, China, 1998; pp. 222–223. [Google Scholar]
- Xie, S.L.; Zhou, A.G.; Feng, Y.Y.; Wang, Z.L.; Fan, L.F.; Zhang, Y.; Zeng, F.; Zou, J.X. Effects of fasting and re-feeding on mstn and mstnb genes expressions in Cranoglanis bouderius. Gene 2019, 682, 1–12. [Google Scholar] [CrossRef]
- Chen, J.T.; Zhou, A.G.; Xie, S.L.; Wang, C.; Lv, Z.J.; Zou, J.X. Comparative Proteomic Identification of Mature and Immature Sperm in the Catfish Cranoglanis bouderius. PLoS ONE 2016, 11, e0151254. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Chen, J.Y.; Ding, G.H.; Lin, Z.H. Analyzing the gonadal transcriptome of the frog Hoplobatrachus rugulosus to identify genes involved in sex development. BMC Genom. 2021, 22, 552. [Google Scholar] [CrossRef]
- Zhang, Y.; Waiho, K.; Ikhwanuddin, M.; Ma, H.Y. Identification of Sex-Related Genes from the Three-Spot Swimming Crab Portunus sanguinolentus and Comparative Analysis with the Crucifix Crab Charybdis feriatus. Animals 2021, 11, 1946. [Google Scholar] [CrossRef]
- Wu, Y.P.; Zhao, X.Y.; Chen, L.; Wang, J.H.; Duan, Y.Q.; Li, H.Y.; Lu, L.Z. Transcriptomic Analyses of the Hypothalamic-Pituitary-Gonadal Axis Identify Candidate Genes Related to Egg Production in Xinjiang Yili Geese. Animals 2020, 10, 90. [Google Scholar] [CrossRef] [Green Version]
- Cui, W.X.; Yang, Q.; Zhang, Y.; Farhadi, A.; Fang, H.; Zheng, H.P.; Li, S.K.; Zhang, Y.L.; Ikhwanuddin, M.; Ma, H.Y. Integrative Transcriptome Sequencing Reveals the Molecular Difference of Maturation Process of Ovary and Testis in Mud Crab Scylla paramamosain. Front. Mar. Sci. 2021, 8, 658091. [Google Scholar] [CrossRef]
- Hu, Y.C.; Wang, B.Z.; Du, H.J. A review on sox genes in fish. Rev. Aquac. 2021, 13, 1986–2003. [Google Scholar] [CrossRef]
- Xu, S.; Zhang, S.; Zhang, W.; Liu, H.; Wang, M.; Zhong, L.; Bian, W.; Chen, X. Genome-Wide Identification, Phylogeny, and Expression Profile of the Dmrt (Doublesex and Mab-3 Related Transcription Factor) Gene Family in Channel Catfish (Ictalurus punctatus). Front. Genet. 2022, 13, 891204. [Google Scholar] [CrossRef] [PubMed]
- Pascual-Anaya, J.; Sato, I.; Sugahara, F.; Higuchi, S.; Paps, J.; Ren, Y.D.; Takagi, W.; Ruiz-Villalba, A.; Ota, K.G.; Wang, W.; et al. Hagfish and lamprey Hox genes reveal conservation of temporal colinearity in vertebrates. Nat. Ecol. Evol. 2018, 2, 859–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safian, D.; Bogerd, J.; Schulz, R.W. Regulation of spermatogonial development by Fsh: The complementary roles of locally produced Igf and Wnt signaling molecules in adult zebrafish testis. Gen. Comp. Endocrinol. 2019, 284, 113244. [Google Scholar] [CrossRef]
- Chao, Q.; Shen, F.; Xue, Y.; Wu, J.; Zhang, J. Cbx2, a PcG Family Gene, Plays a Regulatory Role in Medaka Gonadal Development. Int. J. Mol. Sci. 2020, 21, 1288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castaneda-Cortes, D.C.; Fernandino, J.I. Stress and sex determination in fish: From brain to gonads. Int. J. Dev. Biol. 2021, 65, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Chae, M.; Bae, I.H.; Lim, S.; Jung, K.; Roh, J.; Kim, W. AP Collagen Peptides Prevent Cortisol-Induced Decrease of Collagen Type I in Human Dermal Fibroblasts. Int. J. Mol. Sci. 2021, 22, 4788. [Google Scholar] [CrossRef]
- He, Z.; Deng, F.Q.; Yang, D.Y.; He, Z.D.; Hu, J.X.; Ma, Z.J.; Zhang, Q.; He, J.Y.; Ye, L.J.; Chen, H.J.; et al. Crosstalk between sex-related genes and apoptosis signaling reveals molecular insights into sex change in a protogynous hermaphroditic teleost fish, ricefield eel Monopterus albus. Aquaculture 2022, 552, 737918. [Google Scholar] [CrossRef]
- Zohar, Y.; Zmora, N.; Trudeau, V.L.; Munoz-Cueto, J.A.; Golan, M. A half century of fish gonadotropin-releasing hormones: Breaking paradigms. J. Neuroendocrinol. 2022, 34, e13069. [Google Scholar] [CrossRef]
- Munsterberg, A.; Lovell-Badge, R. Expression of the mouse anti-mullerian hormone gene suggests a role in both male and female sexual differentiation. Development 1991, 113, 613–624. [Google Scholar] [CrossRef]
- Hinck, A.P.; Archer, S.J.; Qian, S.W.; Roberts, A.B.; Sporn, M.B.; Weatherbee, J.A.; Tsang, M.L.; Lucas, R.; Zhang, B.L.; Wenker, J.; et al. Transforming growth factor beta 1: Three-dimensional structure in solution and comparison with the X-ray structure of transforming growth factor beta 2. Biochemistry 1996, 35, 8517–8534. [Google Scholar] [CrossRef] [PubMed]
- Bourguet, W.; Germain, P.; Gronemeyer, H. Nuclear receptor ligand-binding domains: Three-dimensional structures, molecular interactions and pharmacological implications. Trends Pharmacol. Sci. 2000, 21, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Tao, W.J.; Yuan, J.; Zhou, L.Y.; Sun, L.N.; Sun, Y.L.; Yang, S.J.; Li, M.H.; Zeng, S.; Huang, B.F.; Wang, D.H. Characterization of Gonadal Transcriptomes from Nile Tilapia (Oreochromis niloticus) Reveals Differentially Expressed Genes. PLoS ONE 2013, 8, e63604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claessens, F.; Gewirth, D.T. DNA recognition by nuclear receptors. In Essays in Biochemistry: Nuclear Receptor Superfamily; McEwan, I.J., Ed.; Portland Press Ltd.: London, UK, 2004; Volume 40, pp. 59–72. [Google Scholar]
- Zhang, Z.W.; Zhu, B.; Chen, W.T.; Ge, W. Anti-Mullerian hormone (Amh/amh) plays dual roles in maintaining gonadal homeostasis and gametogenesis in zebrafish. Mol. Cell. Endocrinol. 2020, 517, 15. [Google Scholar] [CrossRef] [PubMed]
- Burris, T.P.; Guo, W.; Le, T.; McCabe, E.R. Identification of a putative steroidogenic factor-1 response element in the DAX-1 promoter. Biochem. Biophys. Res. Commun. 1995, 214, 576–581. [Google Scholar] [CrossRef]
- Draper, B.W.; McCallum, C.M.; Moens, C.B. nanos1 is required to maintain oocyte production in adult zebrafish. Dev. Biol. 2007, 305, 589–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jorgensen, A.; Andersen, O.; Bjerregaard, P.; Rasmussen, L.J. Identification and characterisation of an androgen receptor from zebrafish Danio rerio. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2007, 146, 561–568. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Male 1 | Male 2 | Male 3 | Female 1 | Female 2 | Female 3 |
|---|---|---|---|---|---|---|
| Raw reads | 50,755,254 | 44,882,700 | 54,264,414 | 51,980,490 | 59,555,972 | 56,712,898 |
| Raw bases (GB) | 7.66 | 6.77 | 8.19 | 7.84 | 8.99 | 8.56 |
| Clean reads | 50,230,124 | 44,382,438 | 53,706,854 | 51,603,750 | 59,081,800 | 56,248,822 |
| Clean bases (GB) | 7.38 | 6.50 | 7.89 | 7.64 | 8.76 | 8.34 |
| Error rate (%) | 0.0246 | 0.0246 | 0.0247 | 0.0241 | 0.0241 | 0.0244 |
| Q20 (%) | 98.11 | 98.07 | 98.05 | 98.34 | 98.34 | 98.21 |
| Q30 (%) | 94.64 | 94.63 | 94.56 | 95.11 | 95.11 | 94.77 |
| GC (%) | 49.04 | 49.1 | 49.98 | 50.19 | 50.26 | 50.45 |
| Mapped | 46,878,397 | 41,483,671 | 50,154,719 | 49,439,775 | 56,398,939 | 53,722,386 |
| Mapped (%) | 93.33 | 93.47 | 93.39 | 95.81 | 95.46 | 95.51 |
| Uniquely mapped | 43,477,040 | 38,131,053 | 45,239,980 | 44,625,697 | 51,147,482 | 48,902,184 |
| Uniquely mapped (%) | 86.56 | 85.91 | 84.24 | 86.48 | 86.57 | 86.94 |
| Clean reads | 50,230,124 | 44,382,438 | 53,706,854 | 51,603,750 | 59,081,800 | 56,248,822 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, D.; Pan, Z.; Wang, G.; Ye, B.; Wang, Q.; Zuo, Z.; Zou, J.; Xie, S. Gonadal Transcriptome Analysis and Sequence Characterization of Sex-Related Genes in Cranoglanis bouderius. Int. J. Mol. Sci. 2022, 23, 15840. https://doi.org/10.3390/ijms232415840
Wang D, Pan Z, Wang G, Ye B, Wang Q, Zuo Z, Zou J, Xie S. Gonadal Transcriptome Analysis and Sequence Characterization of Sex-Related Genes in Cranoglanis bouderius. International Journal of Molecular Sciences. 2022; 23(24):15840. https://doi.org/10.3390/ijms232415840
Chicago/Turabian StyleWang, Dongjie, Zhengkun Pan, Guoxia Wang, Bin Ye, Qiujie Wang, Zhiheng Zuo, Jixing Zou, and Shaolin Xie. 2022. "Gonadal Transcriptome Analysis and Sequence Characterization of Sex-Related Genes in Cranoglanis bouderius" International Journal of Molecular Sciences 23, no. 24: 15840. https://doi.org/10.3390/ijms232415840