
METTL3 Regulates the Inflammatory Response in CPB2 Toxin-Exposed IPEC-J2 Cells through the TLR2/NF-κB Signaling Pathway
 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Total m6A Contents and METTL3 Expression Levels Are Upregulated in IPEC-J2 Cells
2.2. METTL3 Promotes the Overall Level of m6A Methylation in IPEC-J2 Cells
2.3. Overexpressing METTL3 in IPEC-J2 Cells Induces CPB2-Induced Inflammation
2.4. Overexpression of METTL3 Decreases Cell Viability of CPB2-Exposed IPEC-J2
2.5. METTL3 Knockdown Promotes Expression of Tight Junction-Related Proteins
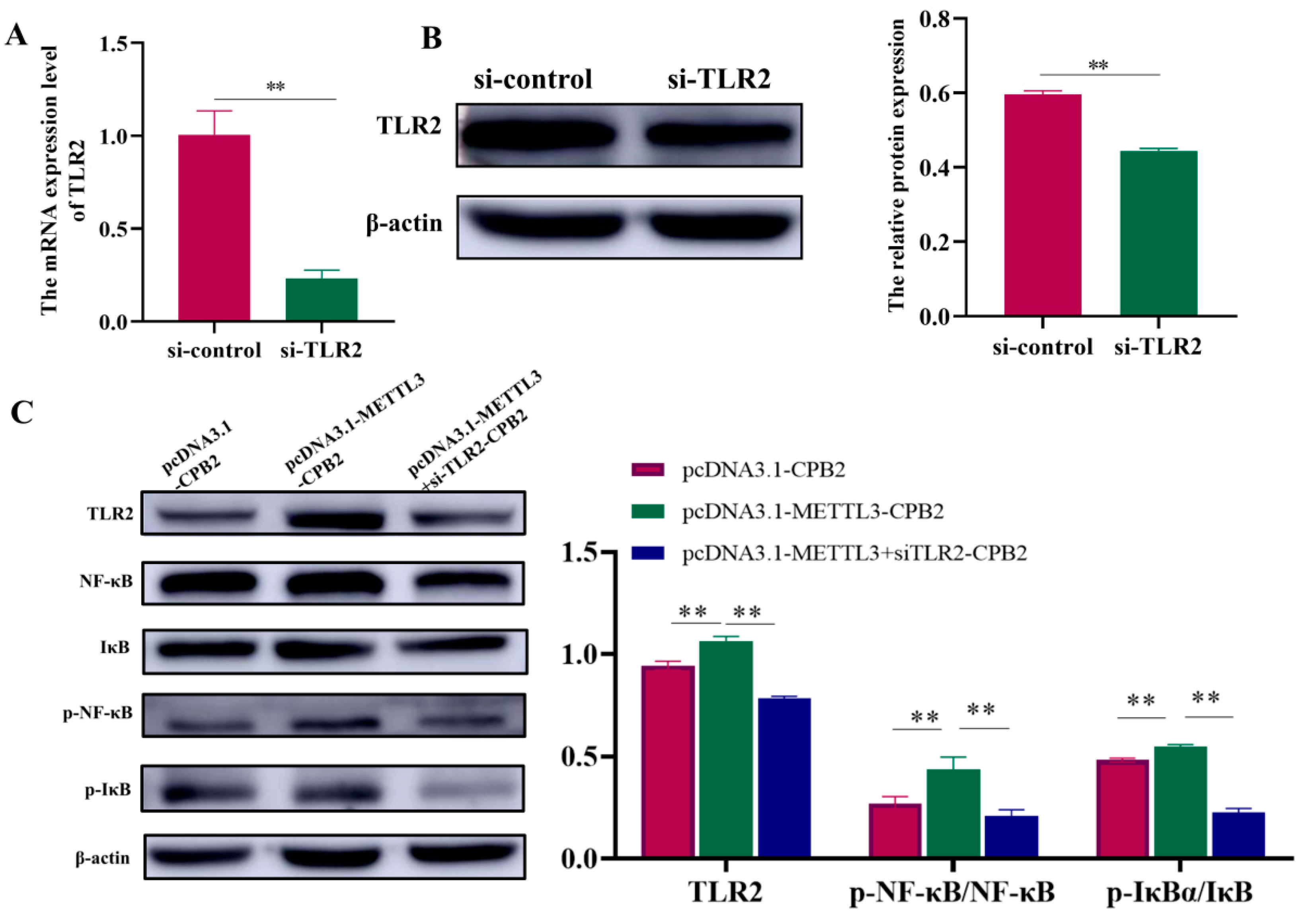
2.6. Overexpression of METTL3 in CPB2-Induced IPEC-J2 Cells Triggers Inflammation through TLR2/NF-κB
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. CPB2 Treatment and Total m6A Measurement
4.3. METTL3 Overexpression, Knockdown Transfection
4.4. Detection of Cell Viability and Lactate Dehydrogenase (LDH)
4.5. Detection of Enzyme-Linked Immunosorbent Assays (ELISA) for Cytokine Detection
4.6. Quantitative Reverse-Transcription PCR (RT-qPCR)
4.7. Western Blotting
4.8. Immunofluorescence Assay
4.9. Methylated RNA Immunoprecipitation-Quantitative Polymerase Chain Reaction (MeRIP-qPCR)
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Allaart, J.G.; van Asten, A.J.; Vernooij, J.C.; Gröne, A. Beta2 toxin is not involved in in vitro cell cytotoxicity caused by human and porcine cpb2-harbouring clostridium perfringens. Vet. Microbiol. 2014, 171, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Allaart, J.G.; van Asten, A.J.; Vernooij, J.C.M.; Gröne, A. Effect of lactobacillus fermentum on beta2 toxin production by clostridium perfringens. Appl. Environ. Microbiol. 2011, 77, 4406–4411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia, J.P.; Beingesser, J.; Fisher, D.J.; Sayeed, S.; Sayeed, S.; McClane, B.A.; Posthaus, H.; Posthaus, H.; Uzal, F.A. The effect of clostridium perfringens type c strain cn3685 and its isogenic beta toxin null mutant in goats. Vet. Microbiol. 2012, 157, 412–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertin, A.M.; Duchet-Suchaux, M.F. Relationship between virulence and adherence of various enterotoxigenic Escherichia coli strains to isolated intestinal epithelial cells from Chinese meishan and European large white pigs. Am. J. Vet. Res. 1991, 52, 45–49. [Google Scholar] [PubMed]
- Songer, J.G.; Uzal, F.A. Clostridial enteric infections in pigs. J. Vet. Diagn. Investig. 2005, 17, 528–536. [Google Scholar] [CrossRef] [Green Version]
- Petit, L.; Gibert, M.; Popoff, M.R. Clostridium perfringens: Toxinotype and genotype. Trends Microbiol. 1999, 7, 104–110. [Google Scholar] [CrossRef]
- Rood, J.I.; Adams, V.; Lacey, J.; Lyras, D.; McClane, B.A.; Melville, S.B.; Moore, R.J.; Popoff, M.R.; Sarker, M.R.; Songer, J.G.; et al. Expansion of the clostridium perfringens toxin-based typing scheme. Anaerobe 2018, 53, 5–10. [Google Scholar] [CrossRef]
- Miclard, J.; Jäggi, M.; Sutter, E.; Wyder, M.; Grabscheid, B.; Posthaus, H. Clostridium perfringens beta-toxin targets endothelial cells in necrotizing enteritis in piglets. Vet. Microbiol. 2009, 137, 320–325. [Google Scholar] [CrossRef]
- Nagahama, M.; Kihara, A.; Kintoh, H.; Oda, M.; Sakurai, J. Involvement of tumour necrosis factor-alpha in clostridium perfringens beta-toxin-induced plasma extravasation in mice. Br. J. Pharmacol. 2010, 153, 1296–1302. [Google Scholar] [CrossRef] [Green Version]
- Nagahama, M.; Morimitsu, S.; Kihara, A.; Akita, M.; Setsu, K.; Sakurai, J. Involvement of tachykinin receptors in clostridium perfringens beta-toxin-induced plasma extravasation. Br. J. Pharmacol. 2003, 138, 23–30. [Google Scholar] [CrossRef]
- Gibert, M.; Jolivet-Reynaud, C.; Popoff, M.R. Beta2 toxin, a novel toxin produced by clostridium perfringens. Gene 1997, 203, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Bueschel, D.M.; Jost, B.H.; Billington, S.J.; Trinh, H.T.; Songer, J.G. Prevalence of cpb2, encoding beta2 toxin, in clostridium perfringens field isolates: Correlation of genotype with phenotype. Vet. Microbiol. 2003, 94, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Yang, Q.; Huang, X.; Yan, Z.; Zhang, S.; Luo, R.; Wang, P.; Wang, W.; Xie, K.; Jiang, T.; et al. Effects of clostridium perfringens beta2 toxin on apoptosis, inflammation, and barrier function of intestinal porcine epithelial cells. Microb. Pathog. 2020, 147, 104379. [Google Scholar] [CrossRef] [PubMed]
- Luo, R.; Yang, Q.; Huang, X.; Yan, Z.; Gao, X.; Wang, W.; Xie, K.; Wang, P.; Gun, S. Clostridium perfringens beta2 toxin induced in vitro oxidative damage and its toxic assessment in porcine small intestinal epithelial cell lines. Gene 2020, 759, 144999. [Google Scholar] [CrossRef] [PubMed]
- Cantara, W.A.; Crain, P.F.; Rozenski, J.; McCloskey, J.A.; Harris, K.A.; Zhang, X.; Vendeix, F.A.P.; Fabris, D.; Agris, P.F. The rna modification database, rnamdb: 2011 update. Nucleic Acids Res. 2011, 39, D195–D201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Y.; Hou, G.; Zhang, H.; Dou, J.; He, J.; Guo, Y.; Li, L.; Chen, R.; Wang, Y.; Deng, R.; et al. Sumoylation of the m6A-RNA methyltransferase mettl3 modulates its function. Nucleic Acids Res. 2018, 46, 5195–5208. [Google Scholar] [CrossRef]
- Xu, C.A.-O.; Wang, X.; Liu, K.; Roundtree, I.A.; Tempel, W.; Li, Y.; Lu, Z.; He, C.; Min, J. Structural basis for selective binding of m6A RNA by the YTHDC1 YTH domain. Nat. Chem. Biol. 2014, 10, 927–929. [Google Scholar] [CrossRef]
- Liu, J.; Yue, Y.; Han, D.; Wang, X.; Fu, Y.; Zhang, L.; Jia, G.; Yu, M.; Lu, Z.; Deng, X.; et al. A mettl3-mettl14 complex mediates mammalian nuclear rna n6-adenosine methylation. Nat. Chem. Biol. 2014, 10, 93–95. [Google Scholar] [CrossRef] [Green Version]
- Ping, X.L.; Sun, B.F.; Wang, L.; Xiao, W.; Yang, X.; Wang, W.J.; Adhikari, S.; Shi, Y.; Lv, Y.; Chen, Y.S.; et al. Mammalian wtap is a regulatory subunit of the rna n6-methyladenosine methyltransferase. Cell Res. 2014, 24, 177–189. [Google Scholar] [CrossRef] [Green Version]
- Schumann, U.; Shafik, A.; Preiss, T. Mettl3 gains r/w access to the epitranscriptome. Mol. Cell 2016, 62, 323–324. [Google Scholar] [CrossRef]
- Zheng, G.; Dahl, J.A.; Niu, Y.; Fedorcsak, P.; Huang, C.-M.; Li, C.J.; Vågbø, C.B.; Shi, Y.; Wang, W.-L.; Song, S.-H.; et al. Alkbh5 is a mammalian rna demethylase that impacts rna metabolism and mouse fertility. Mol. Cell 2013, 49, 18–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, G.; Fu, Y.; Zhao, X.; Dai, Q.; Zheng, G.; Yang, Y.; Yi, C.; Lindahl, T.; Pan, T.; Yang, Y.-G.; et al. N6-methyladenosine in nuclear rna is a major substrate of the obesity-associated fto. Nat. Chem. Biol. 2011, 7, 885–887. [Google Scholar] [CrossRef] [PubMed]
- Meyer, K.D.; Jaffrey, S.R. Rethinking m(6)a readers, writers, and erasers. Annu. Rev. Cell Dev. Biol. 2017, 33, 319–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, H.; Wang, X.; Lu, Z.; Zhao, B.S.; Ma, H.; Hsu, P.J.; Liu, C.; He, C. Ythdf3 facilitates translation and decay of n(6)-methyladenosine-modified rna. Cell Res. 2017, 27, 315–328. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Lu, Z.; Gomez, A.; Hon, G.C.; Yue, Y.; Han, D.; Fu, Y.; Parisien, M.; Dai, Q.; Jia, G.; et al. N6-methyladenosine-dependent regulation of messenger rna stability. Nature 2014, 505, 117–120. [Google Scholar] [CrossRef] [Green Version]
- Aderem, A.; Ulevitch, R.J. Toll-like receptors in the induction of the innate immune response. Nature 2000, 406, 782–787. [Google Scholar] [CrossRef]
- Lee, M.S.; Kim, Y.J. Signaling pathways downstream of pattern-recognition receptors and their cross talk. Annu. Rev. Biochem. 2007, 76, 447–480. [Google Scholar] [CrossRef]
- Cario, E.; Gerken, G.; Podolsky, D.K. Toll-like receptor 2 controls mucosal inflammation by regulating epithelial barrier function. Gastroenterology 2007, 132, 1359–1374. [Google Scholar] [CrossRef]
- Goering, J.; Pope, M.R.; Fleming, S.D. Tlr2 regulates complement-mediated inflammation induced by blood loss during hemorrhage. Shock 2016, 45, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.T.; Kral, J.G. The nf-kappab/ikappab signaling system: A molecular target in breast cancer therapy. J. Surg. Res. 2005, 123, 158–169. [Google Scholar] [CrossRef]
- Dinarello, C.A. Il-1: Discoveries, controversies and future directions. Eur. J. Immunol. 2010, 40, 595–653. [Google Scholar] [CrossRef] [PubMed]
- Garlanda, C.; Dinarello, C.A.; Mantovani, A. The interleukin-1 family: Back to the future. Immunity 2013, 39, 1003–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, M.H.; Yang, Y.L.; Chen, Y.-P.; Hua, K.-F.; Lu, C.-P.; Sheu, F.; Lin, G.-H.; Tsay, S.-S.; Liang, S.-M.; Wu, S.-H. A novel exopolysaccharide from the biofilm of thermus aquaticus yt-1 induces the immune response through toll-like receptor 2. J. Biol. Chem. 2011, 286, 17736–17745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheller, J.; Chalaris, A.; Schmidt-Arras, D.; Rose-John, S. The pro- and anti-inflammatory properties of the cytokine interleukin-6. Biochim. Et Biophys. Acta-Mol. Cell Res. 2011, 1813, 878–888. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Yan, S.A.-O.; Lu, H.; Wang, S.A.-O.; Xu, D.A.-O. Mettl3 attenuates lps-induced inflammatory response in macrophages via nf-κb signaling pathway. Mediat. Inflamm. 2019, 2019, 3120391. [Google Scholar] [CrossRef] [Green Version]
- Feng, Z.; Li, Q.; Meng, R.; Yi, B.; Xu, Q.A.-O. Mettl3 regulates alternative splicing of myd88 upon the lipopolysaccharide-induced inflammatory response in human dental pulp cells. J. Cell. Mol. Med. 2018, 22, 2558–2568. [Google Scholar] [CrossRef] [PubMed]
- Tong, J.; Wang, X.A.-O.; Liu, Y.A.-O.; Ren, X.A.-O.; Wang, A.; Chen, Z.A.-O.; Yao, J.; Mao, K.; Liu, T.; Meng, F.A.-O.; et al. Pooled crispr screening identifies m(6)a as a positive regulator of macrophage activation. Sci. Adv. 2021, 7, eabd4742. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, F. Rna methylation regulates hematopoietic stem/progenitor cell specification. Sci. China (Life Sci.) 2018, 61, 610–612. [Google Scholar] [CrossRef]
- Geula, S.; Moshitch-Moshkovitz, S.; Dominissini, D.; Mansour, A.A.; Kol, N.; Salmon-Divon, M.; Hershkovitz, V.; Peer, E.; Mor, N.; Manor, Y.S.; et al. m6A mRNA methylation facilitates resolution of naïve pluripotency toward differentiation. Science 2015, 347, 1002–1006. [Google Scholar] [CrossRef]
- Wen, L.; Sun, W.; Xia, D.; Wang, Y.; Li, J.; Yang, S. The m6A methyltransferase METTL3 promotes LPS-induced microglia inflammation through TRAF6/NF-κB pathway. NeuroReport 2020, 33, 243–251. [Google Scholar] [CrossRef]
- Liu, Q.; Li, M.; Jiang, L.; Jiang, R.; Fu, B. Mettl3 promotes experimental osteoarthritis development by regulating inflammatory response and apoptosis in chondrocyte. Biochem. Biophys. Res. Commun. 2019, 516, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Zong, X.; Zhao, J.; Wang, H.; Lu, Z.; Wang, F.; Du, H.; Wang, Y. Mettl3 deficiency sustains long-chain fatty acid absorption through suppressing traf6-dependent inflammation response. J. Immunol. 2019, 202, 567–578. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Gu, X.; Li, D.; Cai, L.; Xu, Q. Mettl3 regulates osteoblast differentiation and inflammatory response via smad signaling and mapk signaling. Int. J. Mol. Sci. 2020, 21, 199. [Google Scholar] [CrossRef] [Green Version]
- Brun, P.; Giron, M.C.; Qesari, M.; Porzionato, A.; Caputi, V.; Zoppellaro, C.; Banzato, S.; Grillo, A.R.; Spagnol, L.; De Caro, R.; et al. Toll-like receptor 2 regulates intestinal inflammation by controlling integrity of the enteric nervous system. Gastroenterology 2013, 145, 1323–1333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janeckova, L.; Pospichalova, V.; Fafilek, B.; Vojtechova, M.; Tureckova, J.; Dobes, J.; Dubuissez, M.; Leprince, D.; Baloghova, N.; Horazna, M.; et al. Hic1 tumor suppressor loss potentiates tlr2/nf-κb signaling and promotes tissue damage-associated tumorigenesis. Mol. Cancer Res. 2015, 13, 1139–1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Huang, X.; Xu, P.; Liu, X.; Zhou, Z.; Wang, F.; Li, J.; Wang, Y.; Xian, X.; Liu, G.; et al. Apolipoprotein c3 aggravates diabetic nephropathy in type 1 diabetes by activating the renal tlr2/nf-κb pathway. Metabolism 2021, 119, 154740. [Google Scholar] [CrossRef] [PubMed]
- Wirtz, S.; Becker, C.; Fantini, M.C.; Nieuwenhuis, E.E.; Tubbe, I.; Galle, P.R.; Schild, H.-J.; Birkenbach, M.; Blumberg, R.S.; Neurath, M.F. Ebv-induced gene 3 transcription is induced by tlr signaling in primary dendritic cells via nf-kappa b activation. J. Immunol. 2005, 174, 2814–2824. [Google Scholar] [CrossRef] [Green Version]
- Shon, W.J.; Lee, Y.K.; Shin, J.H.; Choi, E.Y.; Shin, D.M. Severity of dss-induced colitis is reduced in ido1-deficient mice with down-regulation of tlr-myd88-NF-kb transcriptional networks. Sci. Rep. 2015, 5, 17305. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Gan, Y.; Li, M.; Chen, L.; Liang, J.; Zhuo, J.; Luo, H.; Xu, N.; Wu, X.; Wu, Q.; et al. Patchouli alcohol attenuates 5-fluorouracil-induced intestinal mucositis via tlr2/myd88/nf-kb pathway and regulation of microbiota. Biomed. Pharmacother. 2020, 124, 109883. [Google Scholar] [CrossRef]
- Kim, S.; Karin, M. Role of tlr2-dependent inflammation in metastatic progression. Ann. N. Y. Acad. Sci. 2011, 1217, 191–206. [Google Scholar] [CrossRef]
- Tye, H.; Kennedy, C.L.; Najdovska, M.; McLeod, L.; McCormack, W.; Hughes, N.; Dev, A.; Sievert, W.; Ooi, C.H.; Ishikawa, T.-O.; et al. Stat3-driven upregulation of tlr2 promotes gastric tumorigenesis independent of tumor inflammation. Cancer Cell 2021, 22, 466–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oshima, T.; Miwa, H. Gastrointestinal mucosal barrier function and diseases. J. Gastroenterol. 2016, 51, 768–778. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Chatterjee, I.; Anbazhagan, A.N.; Jayawardena, D.; Priyamvada, S.; Alrefai, W.A.; Sun, J.A.-O.; Borthakur, A.A.-O.; Dudeja, P.K. Cryptosporidium parvum disrupts intestinal epithelial barrier function via altering expression of key tight junction and adherens junction proteins. Cell. Microbiol. 2018, 20, e12830. [Google Scholar] [CrossRef] [PubMed]
- Diesing, A.K.; Nossol, C.; Dänicke, S.; Walk, N.; Post, A.; Kahlert, S.; Rothkötter, H.-J.; Kluess, J. Vulnerability of polarised intestinal porcine epithelial cells to mycotoxin deoxynivalenol depends on the route of application. PLoS ONE 2011, 6, e17472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanello, G.; Berri, M.; Dupont, J.; Sizaret, P.-Y.; D’Inca, R.; Salmon, H.; Meurens, F. Saccharomyces cerevisiae modulates immune gene expressions and inhibits etec-mediated erk1/2 and p38 signaling pathways in intestinal epithelial cells. PLoS ONE 2011, 6, e18573. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative pcr and the 2(-delta delta c(t)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Yang, J.; Gao, X.; Huang, X.; Luo, R.; Yang, Q.; Yan, Z.; Wang, P.; Wang, W.; Xie, K.; et al. METTL3 Regulates the Inflammatory Response in CPB2 Toxin-Exposed IPEC-J2 Cells through the TLR2/NF-κB Signaling Pathway. Int. J. Mol. Sci. 2022, 23, 15833. https://doi.org/10.3390/ijms232415833
Zhang J, Yang J, Gao X, Huang X, Luo R, Yang Q, Yan Z, Wang P, Wang W, Xie K, et al. METTL3 Regulates the Inflammatory Response in CPB2 Toxin-Exposed IPEC-J2 Cells through the TLR2/NF-κB Signaling Pathway. International Journal of Molecular Sciences. 2022; 23(24):15833. https://doi.org/10.3390/ijms232415833
Chicago/Turabian StyleZhang, Juanli, Jiaojiao Yang, Xiaoli Gao, Xiaoyu Huang, Ruirui Luo, Qiaoli Yang, Zunqiang Yan, Pengfei Wang, Wei Wang, Kaihui Xie, and et al. 2022. "METTL3 Regulates the Inflammatory Response in CPB2 Toxin-Exposed IPEC-J2 Cells through the TLR2/NF-κB Signaling Pathway" International Journal of Molecular Sciences 23, no. 24: 15833. https://doi.org/10.3390/ijms232415833