

Crosstalk between Methylation and ncRNAs in Breast Cancer: Therapeutic and Diagnostic Implications
Abstract
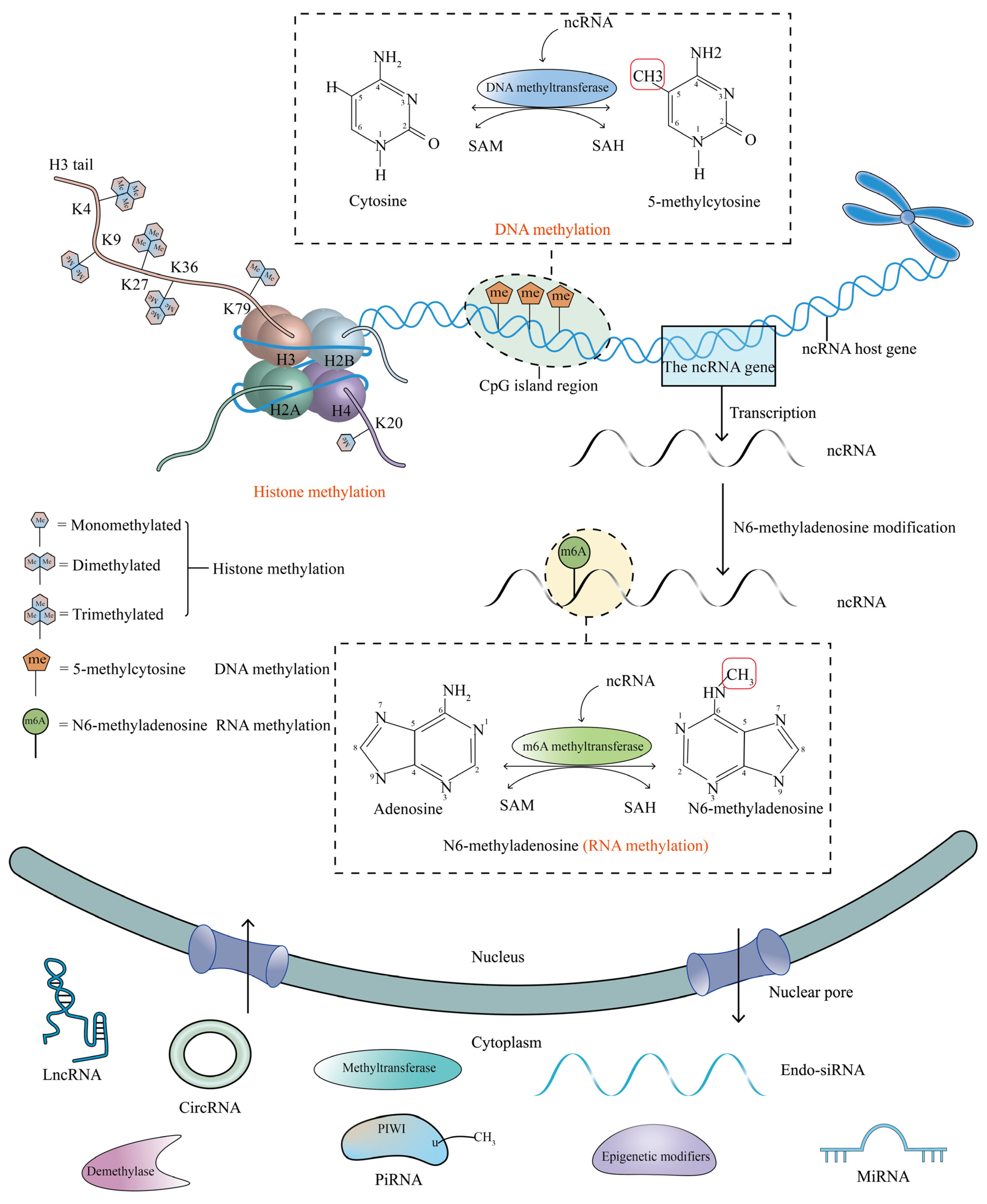
:1. Introduction
1.1. DNA Methylation
1.2. N6-Methyladenosine (m6A)
1.3. Histone Methylation
1.4. Non-Coding RNAs (ncRNAs)
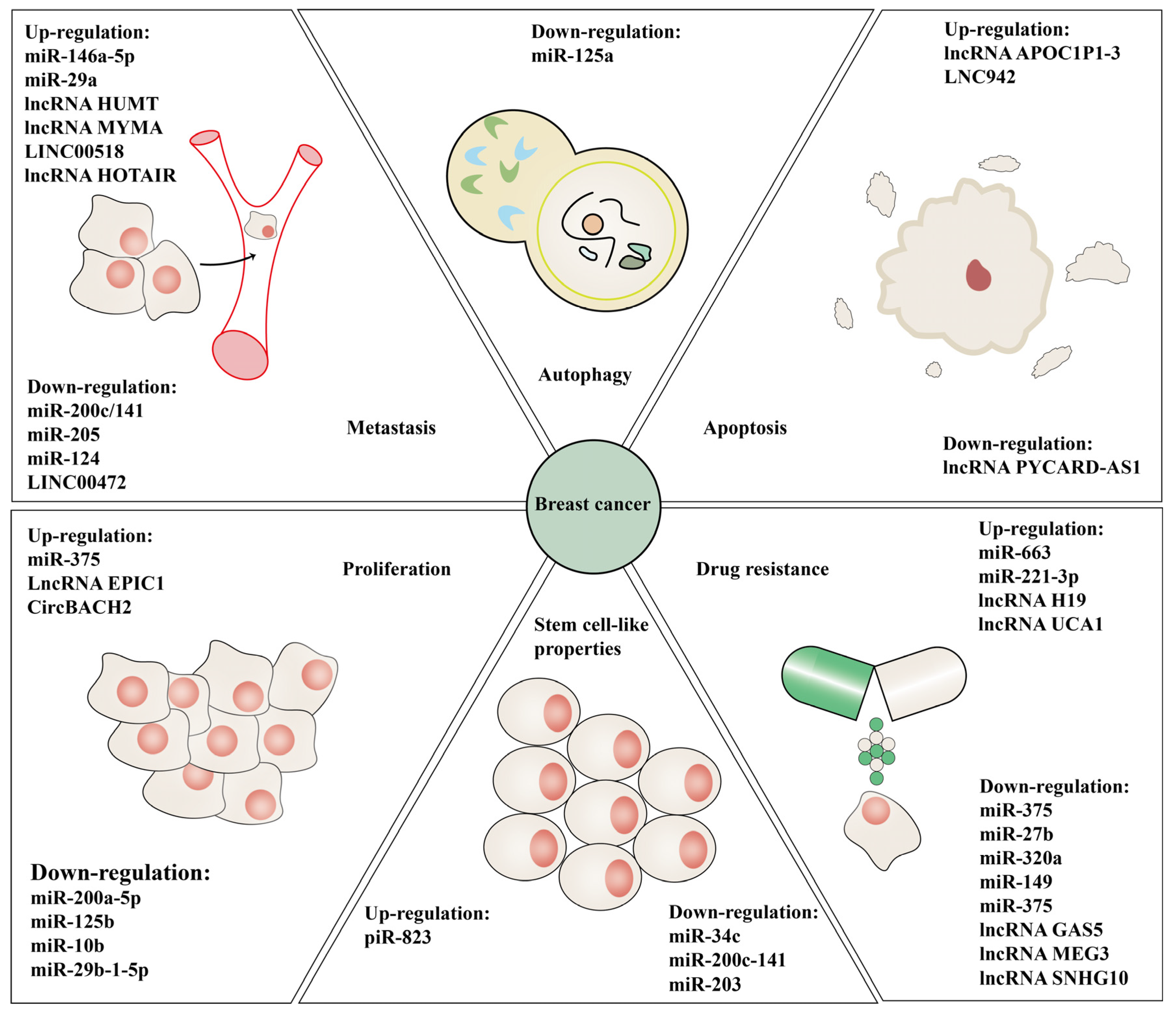
2. Crosstalk between Methylation and ncRNAs in BC
2.1. MiRNA
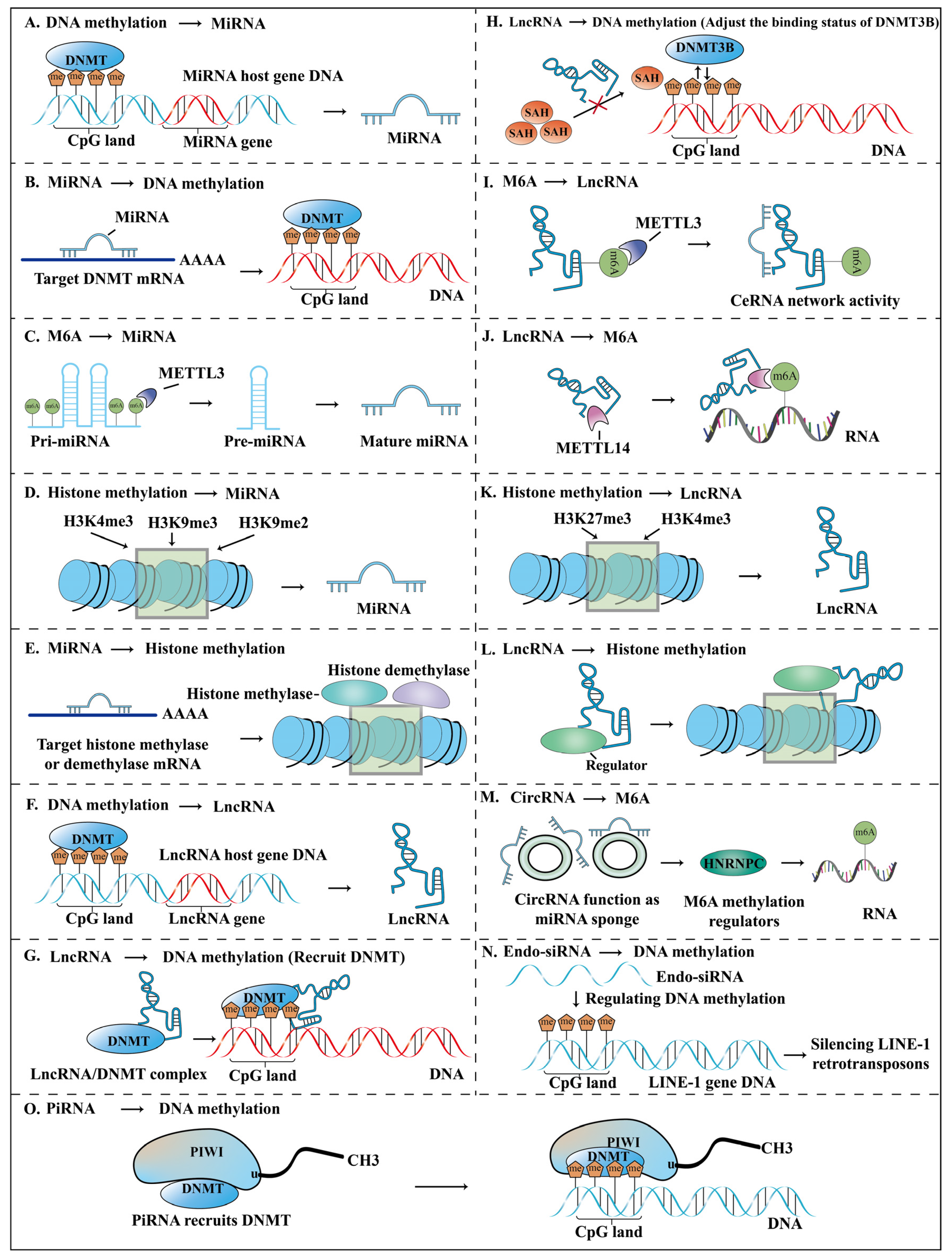
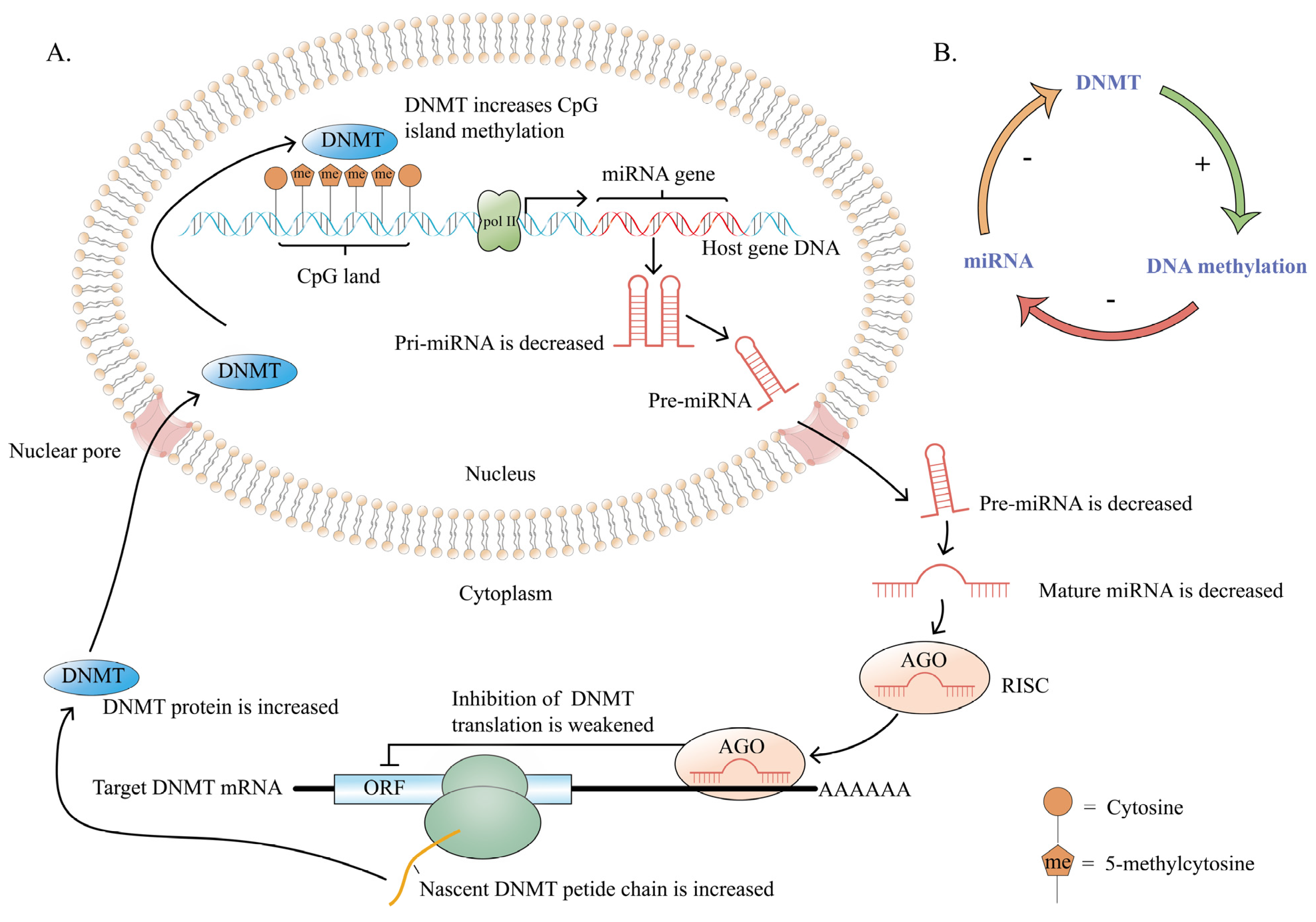
2.1.1. Crosstalk between DNA Methylation and miRNA
- miRNA regulation by DNA methylation
- 2.
- Regulation of DNA methylation by miRNA
2.1.2. Crosstalk between m6A and miRNA
2.1.3. Crosstalk between Histone Methylation and miRNA
- miRNA regulation by histone methylation
- 2.
- Regulation of histone methylation by miRNA
2.2. LncRNA
2.2.1. DNA Methylation and lncRNA
- lncRNA regulation by DNA methylation
- 2.
- Regulation of DNA methylation by lncRNA
2.2.2. m6A and lncRNA
2.2.3. Histone Methylation and lncRNA
- Regulation of lncRNA by histone methylation
- 2.
- Regulation of histone methylation by lncRNA
2.3. CircRNA
2.4. Endo-siRNA
2.5. PiRNA
2.6. Housekeeping ncRNAs
3. Therapeutic and Diagnostic Implications
3.1. Treatment Strategies Based on Methylation of ncRNA
3.2. Diagnostic Markers Based on Methylation of ncRNA
4. Summary
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Brane, A.C.; Tollefsbol, T.O. MicroRNAs and Epigenetics Strategies to Reverse Breast Cancer. Cells 2019, 8, 1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanwal, R.; Gupta, S. Epigenetics and cancer. J. Appl. Physiol. 2010, 109, 598–605. [Google Scholar] [CrossRef] [PubMed]
- Rose, M.; Kloten, V.; Noetzel, E.; Gola, L.; Ehling, J.; Heide, T.; Meurer, S.K.; Gaiko-Shcherbak, A.; Sechi, A.S.; Huth, S.; et al. ITIH5 mediates epigenetic reprogramming of breast cancer cells. Mol. Cancer 2017, 16, 44. [Google Scholar] [CrossRef] [Green Version]
- Jeong, G.-Y.; Park, M.K.; Choi, H.-J.; An, H.W.; Park, Y.-U.; Choi, H.-J.; Park, J.; Kim, H.-Y.; Son, T.; Lee, H.; et al. NSD3-Induced Methylation of H3K36 Activates NOTCH Signaling to Drive Breast Tumor Initiation and Metastatic Progression. Cancer Res. 2021, 81, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, R.J.; Lewis, Z.A.; Goll, M.G. DNA Methylation: Shared and Divergent Features across Eukaryotes. Trends Genet. 2019, 35, 818–827. [Google Scholar] [CrossRef]
- Nishiyama, A.; Nakanishi, M. Navigating the DNA methylation landscape of cancer. Trends Genet. 2021, 37, 1012–1027. [Google Scholar] [CrossRef]
- Hazra, A.; Bose, P.; Sunita, P.; Pattanayak, S.P. Molecular epigenetic dynamics in breast carcinogenesis. Arch. Pharmacal Res. 2021, 44, 741–763. [Google Scholar] [CrossRef]
- Wu, H.-J.; Chu, P.-Y. Epigenetic Regulation of Breast Cancer Stem Cells Contributing to Carcinogenesis and Therapeutic Implications. Int. J. Mol. Sci. 2021, 22, 8113. [Google Scholar] [CrossRef]
- He, L.; Li, H.; Wu, A.; Peng, Y.; Shu, G.; Yin, G. Functions of N6-methyladenosine and its role in cancer. Mol. Cancer 2019, 18, 176. [Google Scholar] [CrossRef]
- Ma, S.; Chen, C.; Ji, X.; Liu, J.; Zhou, Q.; Wang, G.; Yuan, W.; Kan, Q.; Sun, Z. The interplay between m6A RNA methylation and noncoding RNA in cancer. J. Hematol. Oncol. 2019, 12, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; Lv, J.; Yu, H.; Han, J.; Yang, X.; Feng, D.; Wu, Q.; Yuan, B.; Lu, Q.; Yang, H. Mechanism of RNA modification N6-methyladenosine in human cancer. Mol. Cancer 2020, 19, 104. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Su, K.; Sun, X.; Jiang, Y.; Wang, L.; Hu, C.; Zhang, C.; Lu, M.; Du, X.; Xing, B. Sec62 promotes stemness and chemoresistance of human colorectal cancer through activating Wnt/β-catenin pathway. J. Exp. Clin. Cancer Res. 2021, 40, 132. [Google Scholar] [CrossRef]
- Niu, Y.; Lin, Z.; Wan, A.; Chen, H.; Liang, H.; Sun, L.; Wang, Y.; Li, X.; Xiong, X.-F.; Wei, B.; et al. RNA N6-methyladenosine demethylase FTO promotes breast tumor progression through inhibiting BNIP3. Mol. Cancer 2019, 18, 46. [Google Scholar] [CrossRef] [Green Version]
- Audia, J.E.; Campbell, R.M. Histone Modifications and Cancer. Cold Spring Harb. Perspect. Biol. 2016, 8, a019521. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Wu, F.; Wu, J. Targeting histone methylation for cancer therapy: Enzymes, inhibitors, biological activity and perspectives. J. Hematol. Oncol. 2016, 9, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyun, K.; Jeon, J.; Park, K.; Kim, J. Writing, erasing and reading histone lysine methylations. Exp. Mol. Med. 2017, 49, e324. [Google Scholar] [CrossRef] [Green Version]
- Yoo, K.H.; Hennighausen, L. EZH2 methyltransferase and H3K27 methylation in breast cancer. Int. J. Biol. Sci. 2012, 8, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Slack, F.J.; Chinnaiyan, A.M. The Role of Non-coding RNAs in Oncology. Cell 2019, 179, 1033–1055. [Google Scholar] [CrossRef]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef]
- Chipman, L.B.; Pasquinelli, A.E. miRNA Targeting: Growing beyond the Seed. Trends Genet. 2019, 35, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Goodall, G.J.; Wickramasinghe, V.O. RNA in cancer. Nat. Rev. Cancer 2021, 21, 22–36. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, L.S.; Andersen, M.S.; Stagsted, L.V.W.; Ebbesen, K.K.; Hansen, T.B.; Kjems, J. The biogenesis, biology and characterization of circular RNAs. Nat. Rev. Genet. 2019, 20, 675–691. [Google Scholar] [CrossRef] [PubMed]
- Shang, Q.; Yang, Z.; Jia, R.; Ge, S. The novel roles of circRNAs in human cancer. Mol. Cancer 2019, 18, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okamura, K.; Lai, E.C. Endogenous small interfering RNAs in animals. Nat. Rev. Mol. Cell Biol. 2008, 9, 673–678. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Pan, Y.; Fang, Y.; Zhang, J.; Xie, M.; Yang, F.; Yu, T.; Ma, P.; Li, W.; Shu, Y. The Biogenesis and Functions of piRNAs in Human Diseases. Mol. Ther. Nucleic Acids 2020, 21, 108–120. [Google Scholar] [CrossRef]
- Chen, M.; Zhang, C.; Liu, W.; Du, X.; Liu, X.; Xing, B. Long noncoding RNA LINC01234 promotes hepatocellular carcinoma progression through orchestrating aspartate metabolic reprogramming. Mol. Ther. 2022, 30, 2354–2369. [Google Scholar] [CrossRef]
- Gao, X.; Qin, T.; Mao, J.; Zhang, J.; Fan, S.; Lu, Y.; Sun, Z.; Zhang, Q.; Song, B.; Li, L. PTENP1/miR-20a/PTEN axis contributes to breast cancer progression by regulating PTEN via PI3K/AKT pathway. J. Exp. Clin. Cancer Res. 2019, 38, 256. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.; Kim, C.; Lee, H.; Rho, J.G.; Seo, J.W.; Nam, J.W.; Song, W.K.; Nam, S.W.; Kim, W.; Lee, E.K. Downregulation of microRNA-362-3p and microRNA-329 promotes tumor progression in human breast cancer. Cell Death Differ. 2016, 23, 484–495. [Google Scholar] [CrossRef]
- Li, D.; Zhao, Y.; Liu, C.; Chen, X.; Qi, Y.; Jiang, Y.; Zou, C.; Zhang, X.; Liu, S.; Wang, X.; et al. Analysis of MiR-195 and MiR-497 expression, regulation and role in breast cancer. Clin. Cancer Res. 2011, 17, 1722–1730. [Google Scholar] [CrossRef]
- Jiang, L.; Yu, L.; Zhang, X.; Lei, F.; Wang, L.; Liu, X.; Wu, S.; Zhu, J.; Wu, G.; Cao, L.; et al. miR-892b Silencing Activates NF-kappaB and Promotes Aggressiveness in Breast Cancer. Cancer Res. 2016, 76, 1101–1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Wu, J.; Guan, L.; Qi, L.; Tang, Y.; Ma, B.; Zhan, J.; Wang, Y.; Fang, W.; Zhang, H. Kindlin 2 promotes breast cancer invasion via epigenetic silencing of the microRNA200 gene family. Int. J. Cancer 2013, 133, 1368–1379. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Qu, X.; Zhao, C.; Xu, L.; Hou, K.; Liu, Y.; Zhang, N.; Feng, J.; Shi, S.; Zhang, L.; et al. FEN1 mediates miR-200a methylation and promotes breast cancer cell growth via MET and EGFR signaling. FASEB J. 2019, 33, 10717–10730. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Yang, S.; Wang, M.; Liu, D.; Liu, Y.; Zhang, Y.; Zhang, Q. Epigenetically altered miR193a3p promotes HER2 positive breast cancer aggressiveness by targeting GRB7. Int. J. Mol. Med. 2019, 43, 2352–2360. [Google Scholar] [CrossRef]
- Lee, J.Y.; Park, M.K.; Park, J.H.; Lee, H.J.; Shin, D.H.; Kang, Y.; Lee, C.H.; Kong, G. Loss of the polycomb protein Mel-18 enhances the epithelial-mesenchymal transition by ZEB1 and ZEB2 expression through the downregulation of miR-205 in breast cancer. Oncogene 2014, 33, 1325–1335. [Google Scholar] [CrossRef] [Green Version]
- Mudduluru, G.; Ceppi, P.; Kumarswamy, R.; Scagliotti, G.V.; Papotti, M.; Allgayer, H. Regulation of Axl receptor tyrosine kinase expression by miR-34a and miR-199a/b in solid cancer. Oncogene 2011, 30, 2888–2899. [Google Scholar] [CrossRef]
- Png, K.J.; Yoshida, M.; Zhang, X.H.; Shu, W.; Lee, H.; Rimner, A.; Chan, T.A.; Comen, E.; Andrade, V.P.; Kim, S.W.; et al. MicroRNA-335 inhibits tumor reinitiation and is silenced through genetic and epigenetic mechanisms in human breast cancer. Genes Dev. 2011, 25, 226–231. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yang, P.; Sun, T.; Li, D.; Xu, X.; Rui, Y.; Li, C.; Chong, M.; Ibrahim, T.; Mercatali, L.; et al. miR-126 and miR-126* repress recruitment of mesenchymal stem cells and inflammatory monocytes to inhibit breast cancer metastasis. Nat. Cell Biol. 2013, 15, 284–294. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Sun, Y.; Yuan, Y.; Han, Z.; Zhang, P.; Zhang, J.; You, M.J.; Teruya-Feldstein, J.; Wang, M.; Gupta, S.; et al. miR-100 induces epithelial-mesenchymal transition but suppresses tumorigenesis, migration and invasion. PLoS Genet. 2014, 10, e1004177. [Google Scholar] [CrossRef]
- Zhang, Y.; Yan, L.X.; Wu, Q.N.; Du, Z.M.; Chen, J.; Liao, D.Z.; Huang, M.Y.; Hou, J.H.; Wu, Q.L.; Zeng, M.S.; et al. miR-125b is methylated and functions as a tumor suppressor by regulating the ETS1 proto-oncogene in human invasive breast cancer. Cancer Res. 2011, 71, 3552–3562. [Google Scholar] [CrossRef]
- Biagioni, F.; Bossel Ben-Moshe, N.; Fontemaggi, G.; Canu, V.; Mori, F.; Antoniani, B.; Di Benedetto, A.; Santoro, R.; Germoni, S.; De Angelis, F.; et al. miR-10b*, a master inhibitor of the cell cycle, is down-regulated in human breast tumours. EMBO Mol. Med. 2012, 4, 1214–1229. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Jiao, Y.; Zhu, Y.; Wang, Y.; Zhu, J.; Cui, X.; Liu, Y.; He, Y.; Park, E.Y.; Zhang, H.; et al. MicroRNA 34c gene down-regulation via DNA methylation promotes self-renewal and epithelial-mesenchymal transition in breast tumor-initiating cells. J. Biol. Chem. 2012, 287, 465–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, Y.Y.; Wright, J.A.; Attema, J.L.; Gregory, P.A.; Bert, A.G.; Smith, E.; Thomas, D.; Lopez, A.F.; Drew, P.A.; Khew-Goodall, Y.; et al. Epigenetic modulation of the miR-200 family is associated with transition to a breast cancer stem-cell-like state. J. Cell Sci. 2013, 126, 2256–2266. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.; Li, S.; Cui, X.; Lv, X.; Jiao, Y.; Yu, F.; Yao, H.; Song, E.; Chen, Y.; Wang, M.; et al. The overexpression of hypomethylated miR-663 induces chemotherapy resistance in human breast cancer cells by targeting heparin sulfate proteoglycan 2 (HSPG2). J. Biol. Chem. 2013, 288, 10973–10985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, D.X.; Gu, X.T.; Jiang, L.; Jin, J.; Ma, X. A methylation-based regulatory network for microRNA 320a in chemoresistant breast cancer. Mol. Pharmacol. 2014, 86, 536–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, D.X.; Gu, X.T.; Li, Y.R.; Jiang, L.; Jin, J.; Ma, X. Methylation-regulated miR-149 modulates chemoresistance by targeting GlcNAc N-deacetylase/N-sulfotransferase-1 in human breast cancer. FEBS J. 2014, 281, 4718–4730. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.M.; Zhu, H.Y.; Bai, W.D.; Wang, T.; Wang, L.; Chen, Y.; Yang, A.G.; Jia, L.T. Epigenetic silencing of miR-375 induces trastuzumab resistance in HER2-positive breast cancer by targeting IGF1R. BMC Cancer 2014, 14, 134. [Google Scholar] [CrossRef] [Green Version]
- Poli, E.; Zhang, J.; Nwachukwu, C.; Zheng, Y.; Adedokun, B.; Olopade, O.I.; Han, Y.J. Molecular Subtype-Specific Expression of MicroRNA-29c in Breast Cancer Is Associated with CpG Dinucleotide Methylation of the Promoter. PLoS ONE 2015, 10, e0142224. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.G.; Gao, M.Q.; Kang, S.; Choi, Y.P.; Lee, J.H.; Kim, J.E.; Han, H.H.; Mun, S.G.; Cho, N.H. Mechanical compression induces VEGFA overexpression in breast cancer via DNMT3A-dependent miR-9 downregulation. Cell Death Dis. 2017, 8, e2646. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Qin, J.F.; Han, X.; Jin, F.J.; Zhang, J.H.; Lan, L.; Wang, Y. miR-21a negatively modulates tumor suppressor genes PTEN and miR-200c and further promotes the transformation of M2 macrophages. Immunol. Cell Biol. 2018, 96, 68–80. [Google Scholar] [CrossRef]
- Ahmadpour, F.; Igder, S.; Babaahmadi-Rezaei, H.; Khalili, E.; Kanani, M.; Soleimani, V.; Mohammadzadeh, G. Methylation-mediated silencing of miR-125a-5p facilitates breast cancer progression by inducing autophagy. Mol. Biol. Rep. 2022, 49, 6325–6339. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, R.; Rivenbark, A.G.; Coleman, W.B. Loss of post-transcriptional regulation of DNMT3b by microRNAs: A possible molecular mechanism for the hypermethylation defect observed in a subset of breast cancer cell lines. Int. J. Oncol. 2012, 41, 721–732. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Yi, J.; Zheng, X.; Liu, S.; Fu, W.; Ren, L.; Li, L.; Hoon, D.S.B.; Wang, J.; Du, G. miR-29c plays a suppressive role in breast cancer by targeting the TIMP3/STAT1/FOXO1 pathway. Clin. Epigenetics 2018, 10, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Blasio, A.; Di Fiore, R.; Pratelli, G.; Drago-Ferrante, R.; Saliba, C.; Baldacchino, S.; Grech, G.; Scerri, C.; Vento, R.; Tesoriere, G. A loop involving NRF2, miR-29b-1-5p and AKT, regulates cell fate of MDA-MB-231 triple-negative breast cancer cells. J. Cell. Physiol. 2020, 235, 629–637. [Google Scholar] [CrossRef] [PubMed]
- Noyan, S.; Andac Ozketen, A.; Gurdal, H.; Gur Dedeoglu, B. miR-770-5p regulates EMT and invasion in TNBC cells by targeting DNMT3A. Cell. Signal. 2021, 83, 109996. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Jiang, Y.; Yin, Y.; Li, Q.; He, J.; Jing, Y.; Qi, Y.T.; Xu, Q.; Li, W.; Lu, B.; et al. A regulatory circuit of miR-148a/152 and DNMT1 in modulating cell transformation and tumor angiogenesis through IGF-IR and IRS1. J. Mol. Cell Biol. 2013, 5, 3–13. [Google Scholar] [CrossRef] [Green Version]
- Pang, Y.; Liu, J.; Li, X.; Xiao, G.; Wang, H.; Yang, G.; Li, Y.; Tang, S.C.; Qin, S.; Du, N.; et al. MYC and DNMT3A-mediated DNA methylation represses microRNA-200b in triple negative breast cancer. J. Cell. Mol. Med. 2018, 22, 6262–6274. [Google Scholar] [CrossRef] [Green Version]
- Shi, W.; Tang, T.; Li, X.; Deng, S.; Li, R.; Wang, Y.; Wang, Y.; Xia, T.; Zhang, Y.; Zen, K.; et al. Methylation-mediated silencing of miR-133a-3p promotes breast cancer cell migration and stemness via miR-133a-3p/MAML1/DNMT3A positive feedback loop. J. Exp. Clin. Cancer Res. 2019, 38, 429. [Google Scholar] [CrossRef] [Green Version]
- Tan, Y.; Zhang, B.; Wu, T.; Skogerbo, G.; Zhu, X.; Guo, X.; He, S.; Chen, R. Transcriptional inhibiton of Hoxd4 expression by miRNA-10a in human breast cancer cells. BMC Mol. Biol. 2009, 10, 12. [Google Scholar] [CrossRef] [Green Version]
- Yi, D.; Wang, R.; Shi, X.; Xu, L.; Yilihamu, Y.; Sang, J. METTL14 promotes the migration and invasion of breast cancer cells by modulating N6methyladenosine and hsamiR146a5p expression. Oncol. Rep. 2020, 43, 1375–1386. [Google Scholar] [CrossRef]
- Pan, X.; Hong, X.; Li, S.; Meng, P.; Xiao, F. METTL3 promotes adriamycin resistance in MCF-7 breast cancer cells by accelerating pri-microRNA-221-3p maturation in a m6A-dependent manner. Exp. Mol. Med. 2021, 53, 91–102. [Google Scholar] [CrossRef] [PubMed]
- De Souza Rocha Simonini, P.; Breiling, A.; Gupta, N.; Malekpour, M.; Youns, M.; Omranipour, R.; Malekpour, F.; Volinia, S.; Croce, C.M.; Najmabadi, H.; et al. Epigenetically deregulated microRNA-375 is involved in a positive feedback loop with estrogen receptor alpha in breast cancer cells. Cancer Res. 2010, 70, 9175–9184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damiano, V.; Brisotto, G.; Borgna, S.; di Gennaro, A.; Armellin, M.; Perin, T.; Guardascione, M.; Maestro, R.; Santarosa, M. Epigenetic silencing of miR-200c in breast cancer is associated with aggressiveness and is modulated by ZEB1. Genes Chromosom. Cancer 2017, 56, 147–158. [Google Scholar] [CrossRef]
- Mitra, D.; Das, P.M.; Huynh, F.C.; Jones, F.E. Jumonji/ARID1 B (JARID1B) protein promotes breast tumor cell cycle progression through epigenetic repression of microRNA let-7e. J. Biol. Chem. 2011, 286, 40531–40535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, N.; Huangyang, P.; Yang, X.; Han, X.; Yan, R.; Jia, H.; Shang, Y.; Sun, L. microRNA-7 suppresses the invasive potential of breast cancer cells and sensitizes cells to DNA damages by targeting histone methyltransferase SET8. J. Biol. Chem. 2013, 288, 19633–19642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hui, Z.; Yiling, C.; Wenting, Y.; XuQun, H.; ChuanYi, Z.; Hui, L. miR-491-5p functions as a tumor suppressor by targeting JMJD2B in ERalpha-positive breast cancer. FEBS Lett. 2015, 589, 812–821. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Shi, W.; Tang, T.; Wang, Y.; Yin, X.; Chen, Y.; Zhang, Y.; Xing, Y.; Shen, Y.; Xia, T.; et al. miR-29a contributes to breast cancer cells epithelial-mesenchymal transition, migration, and invasion via down-regulating histone H4K20 trimethylation through directly targeting SUV420H2. Cell Death Dis. 2019, 10, 176. [Google Scholar] [CrossRef] [Green Version]
- Aure, M.R.; Fleischer, T.; Bjørklund, S.; Ankill, J.; Castro-Mondragon, J.A.; OSBREAC; Børresen-Dale, A.-L.; Tost, J.; Sahlberg, K.K.; Mathelier, A.; et al. Crosstalk between microRNA expression and DNA methylation drives the hormone-dependent phenotype of breast cancer. Genome Med. 2021, 13, 72. [Google Scholar] [CrossRef]
- Gong, Z.; Chen, J.; Wang, J.; Liu, S.; Ambrosone, C.B.; Higgins, M.J. Differential methylation and expression patterns of microRNAs in relation to breast cancer subtypes among American women of African and European ancestry. PLoS ONE 2021, 16, e0249229. [Google Scholar] [CrossRef]
- Shi, F.; Chen, X.; Fu, A.; Hansen, J.; Stevens, R.; Tjonneland, A.; Vogel, U.B.; Zheng, T.; Zhu, Y. Aberrant DNA methylation of miR-219 promoter in long-term night shiftworkers. Environ. Mol. Mutagen. 2013, 54, 406–413. [Google Scholar] [CrossRef]
- Liu, R.; Jacobs, D.I.; Hansen, J.; Fu, A.; Stevens, R.G.; Zhu, Y. Aberrant methylation of miR-34b is associated with long-term shiftwork: A potential mechanism for increased breast cancer susceptibility. Cancer Causes Control. 2015, 26, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Hsu, P.Y.; Deatherage, D.E.; Rodriguez, B.A.; Liyanarachchi, S.; Weng, Y.I.; Zuo, T.; Liu, J.; Cheng, A.S.; Huang, T.H. Xenoestrogen-induced epigenetic repression of microRNA-9-3 in breast epithelial cells. Cancer Res. 2009, 69, 5936–5945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Zhang, Y. Role of Mammalian DNA Methyltransferases in Development. Annu. Rev. Biochem. 2020, 89, 135–158. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Lin, Y.; Shu, Y.; He, J.; Gao, W. Interaction between N(6)-methyladenosine (m(6)A) modification and noncoding RNAs in cancer. Mol. Cancer 2020, 19, 94. [Google Scholar] [CrossRef] [PubMed]
- Liao, X.H.; Wang, J.G.; Li, L.Y.; Zhou, D.M.; Ren, K.H.; Jin, Y.T.; Lv, L.; Yu, J.G.; Yang, J.Y.; Lu, Q.; et al. Long intergenic non-coding RNA APOC1P1-3 inhibits apoptosis by decreasing alpha-tubulin acetylation in breast cancer. Cell Death Dis. 2016, 7, e2236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Yang, B.; Zhang, M.; Guo, W.; Wu, Z.; Wang, Y.; Jia, L.; Li, S.; Cancer Genome Atlas Research, N.; Xie, W.; et al. lncRNA Epigenetic Landscape Analysis Identifies EPIC1 as an Oncogenic lncRNA that Interacts with MYC and Promotes Cell-Cycle Progression in Cancer. Cancer Cell 2018, 33, 706–720.e9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, S.; Yang, L.; Zou, Y.; Liang, J.Y.; Liu, P.; Gao, G.; Yang, A.; Tang, H.; Xie, X. Long non-coding RNA HUMT hypomethylation promotes lymphangiogenesis and metastasis via activating FOXK1 transcription in triple-negative breast cancer. J. Hematol. Oncol. 2020, 13, 17. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Li, L.; Yuan, H.; Huang, X.W.; Xiang, T.; Dai, S. Up-regulated lncRNA GAS5 promotes chemosensitivity and apoptosis of triple-negative breast cancer cells. Cell Cycle 2019, 18, 1965–1975. [Google Scholar] [CrossRef]
- Li, H.; Wang, P.; Liu, J.; Liu, W.; Wu, X.; Ding, J.; Kang, J.; Li, J.; Lu, J.; Pan, G. Hypermethylation of lncRNA MEG3 impairs chemosensitivity of breast cancer cells. J. Clin. Lab. Anal. 2020, 34, e23369. [Google Scholar] [CrossRef]
- Li, C.; Wang, S.; Xing, Z.; Lin, A.; Liang, K.; Song, J.; Hu, Q.; Yao, J.; Chen, Z.; Park, P.K.; et al. A ROR1-HER3-lncRNA signalling axis modulates the Hippo-YAP pathway to regulate bone metastasis. Nat. Cell Biol. 2017, 19, 106–119. [Google Scholar] [CrossRef]
- Vennin, C.; Spruyt, N.; Robin, Y.M.; Chassat, T.; Le Bourhis, X.; Adriaenssens, E. The long non-coding RNA 91H increases aggressive phenotype of breast cancer cells and up-regulates H19/IGF2 expression through epigenetic modifications. Cancer Lett. 2017, 385, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Miao, H.; Wang, L.; Zhan, H.; Dai, J.; Chang, Y.; Wu, F.; Liu, T.; Liu, Z.; Gao, C.; Li, L.; et al. A long noncoding RNA distributed in both nucleus and cytoplasm operates in the PYCARD-regulated apoptosis by coordinating the epigenetic and translational regulation. PLoS Genet. 2019, 15, e1008144. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.B.; Wei, H.; Wang, J.S.; Li, L.; Chen, A.Y.; Li, Z.G. Down-regulated expression of LINC00518 prevents epithelial cell growth and metastasis in breast cancer through the inhibition of CDX2 methylation and the Wnt signaling pathway. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 708–723. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Xie, S.; Yang, J.; Xiong, H.; Jia, Y.; Zhou, Y.; Chen, Y.; Ying, X.; Chen, C.; Ye, C.; et al. The long noncoding RNA H19 promotes tamoxifen resistance in breast cancer via autophagy. J. Hematol. Oncol. 2019, 12, 81. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Liu, H.; Wan, L.; Zhang, W.; Wang, Q.; Zhang, S.; Shang, S.; Zhang, Y.; Pang, D. The MS-lincRNA landscape reveals a novel lincRNA BCLIN25 that contributes to tumorigenesis by upregulating ERBB2 expression via epigenetic modification and RNA-RNA interactions in breast cancer. Cell Death Dis. 2019, 10, 920. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Hu, X.; Yu, S.; Deng, S.; Yan, M.; Sun, F.; Song, J.; Tang, L. Silence of lncRNA MIAT-mediated inhibition of DLG3 promoter methylation suppresses breast cancer progression via the Hippo signaling pathway. Cell. Signal. 2020, 73, 109697. [Google Scholar] [CrossRef]
- Shao, G.; Fan, X.; Zhang, P.; Liu, X.; Huang, L.; Ji, S. Methylation-dependent MCM6 repression induced by LINC00472 inhibits triple-negative breast cancer metastasis by disturbing the MEK/ERK signaling pathway. Aging 2021, 13, 4962–4975. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, J.; You, Z.; Yin, Y.; Liu, L.; Kang, Y.; Li, S.; Ning, S.; Li, H.; Gong, Y.; et al. LncRNA TINCR favors tumorigenesis via STAT3-TINCR-EGFR-feedback loop by recruiting DNMT1 and acting as a competing endogenous RNA in human breast cancer. Cell Death Dis. 2021, 12, 83. [Google Scholar] [CrossRef]
- Wang, Y.; Dong, T.; Wang, P.; Li, S.; Wu, G.; Zhou, J.; Wang, Z. LINC00922 regulates epithelial-mesenchymal transition, invasive and migratory capacities in breast cancer through promoting NKD2 methylation. Cell. Signal. 2021, 77, 109808. [Google Scholar] [CrossRef]
- Xu, X.; Yuan, X.; Ni, J.; Guo, J.; Gao, Y.; Yin, W.; Li, F.; Wei, L.; Zhang, J. MAGI2-AS3 inhibits breast cancer by downregulating DNA methylation of MAGI2. J. Cell. Physiol. 2021, 236, 1116–1130. [Google Scholar] [CrossRef]
- Aini, S.; Bolati, S.; Ding, W.; Liu, S.; Su, P.; Aili, S.; Naman, Y.; Xuekelaiti, K. LncRNA SNHG10 suppresses the development of doxorubicin resistance by downregulating miR-302b in triple-negative breast cancer. Bioengineered 2022, 13, 11430–11439. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Wu, Z.; Wang, X.; Wang, Y.; Hu, X.; Qin, W.; Lu, S.; Xu, D.; Wu, Y.; Chen, Q.; et al. LNC942 promoting METTL14-mediated m(6)A methylation in breast cancer cell proliferation and progression. Oncogene 2020, 39, 5358–5372. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.; Wang, L. N(6)-Methyladenosine-regulated LINC00675 suppress the proliferation, migration and invasion of breast cancer cells via inhibiting miR-513b-5p. Bioengineered 2021, 12, 10690–10702. [Google Scholar] [CrossRef] [PubMed]
- Pang, B.; Sui, S.; Wang, Q.; Wu, J.; Yin, Y.; Xu, S. Upregulation of DLEU1 expression by epigenetic modification promotes tumorigenesis in human cancer. J. Cell. Physiol. 2019, 234, 17420–17432. [Google Scholar] [CrossRef]
- Pang, B.; Wang, Q.; Ning, S.; Wu, J.; Zhang, X.; Chen, Y.; Xu, S. Landscape of tumor suppressor long noncoding RNAs in breast cancer. J. Exp. Clin. Cancer Res. 2019, 38, 79. [Google Scholar] [CrossRef] [Green Version]
- Bhan, A.; Hussain, I.; Ansari, K.I.; Bobzean, S.A.; Perrotti, L.I.; Mandal, S.S. Bisphenol-A and diethylstilbestrol exposure induces the expression of breast cancer associated long noncoding RNA HOTAIR in vitro and in vivo. J. Steroid Biochem. Mol. Biol. 2014, 141, 160–170. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.J.; Kim, M.; Kim, H.P. Epigenetic regulation of long noncoding RNA UCA1 by SATB1 in breast cancer. BMB Rep. 2016, 49, 578–583. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.A.; Shah, N.; Wang, K.C.; Kim, J.; Horlings, H.M.; Wong, D.J.; Tsai, M.C.; Hung, T.; Argani, P.; Rinn, J.L.; et al. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature 2010, 464, 1071–1076. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Yu, D.; Li, H.; Lv, Y.; Li, S. Long noncoding RNA UCA1 confers tamoxifen resistance in breast cancer endocrinotherapy through regulation of the EZH2/p21 axis and the PI3K/AKT signaling pathway. Int. J. Oncol. 2019, 54, 1033–1042. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Sui, S.; Wu, H.; Zhang, J.; Zhang, X.; Xu, S.; Pang, D. The transcriptional landscape of lncRNAs reveals the oncogenic function of LINC00511 in ER-negative breast cancer. Cell Death Dis. 2019, 10, 599. [Google Scholar] [CrossRef]
- Hu, A.; Hong, F.; Li, D.; Jin, Y.; Kon, L.; Xu, Z.; He, H.; Xie, Q. Long non-coding RNA ROR recruits histone transmethylase MLL1 to up-regulate TIMP3 expression and promote breast cancer progression. J. Transl. Med. 2021, 19, 95. [Google Scholar] [CrossRef] [PubMed]
- Chu, W.; Zhang, X.; Qi, L.; Fu, Y.; Wang, P.; Zhao, W.; Du, J.; Zhang, J.; Zhan, J.; Wang, Y.; et al. The EZH2-PHACTR2-AS1-Ribosome Axis induces Genomic Instability and Promotes Growth and Metastasis in Breast Cancer. Cancer Res. 2020, 80, 2737–2750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, L.; Zhu, G.; Zhang, C.; Deng, Q.; Katsaros, D.; Mayne, S.T.; Risch, H.A.; Mu, L.; Canuto, E.M.; Gregori, G.; et al. Association of large noncoding RNA HOTAIR expression and its downstream intergenic CpG island methylation with survival in breast cancer. Breast Cancer Res. Treat. 2012, 136, 875–883. [Google Scholar] [CrossRef]
- Huang, W.; Li, H.; Yu, Q.; Xiao, W.; Wang, D.O. LncRNA-mediated DNA methylation: An emerging mechanism in cancer and beyond. J. Exp. Clin. Cancer Res. 2022, 41, 100. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Ci, C.; Zhang, X.; Su, M.; Lv, W.; Chen, C.; Liu, H.; Zhang, D.; Zhang, S.; Zhang, Y. Prediction of circRNAs Based on the DNA Methylation-Mediated Feature Sponge Function in Breast Cancer. Front. Bioeng. Biotechnol. 2019, 7, 365. [Google Scholar] [CrossRef]
- Lv, W.; Tan, Y.; Xiong, M.; Zhao, C.; Wang, Y.; Wu, M.; Wu, Y.; Zhang, Q. Analysis and validation of m6A regulatory network: A novel circBACH2/has-miR-944/HNRNPC axis in breast cancer progression. J. Transl. Med. 2021, 19, 527. [Google Scholar] [CrossRef]
- Chen, L.; Dahlstrom, J.E.; Lee, S.H.; Rangasamy, D. Naturally occurring endo-siRNA silences LINE-1 retrotransposons in human cells through DNA methylation. Epigenetics 2012, 7, 758–771. [Google Scholar] [CrossRef]
- Liu, T.; Wang, J.; Sun, L.; Li, M.; He, X.; Jiang, J.; Zhou, Q. Piwi-interacting RNA-651 promotes cell proliferation and migration and inhibits apoptosis in breast cancer by facilitating DNMT1-mediated PTEN promoter methylation. Cell Cycle 2021, 20, 1603–1616. [Google Scholar] [CrossRef]
- Ding, X.; Li, Y.; Lu, J.; Zhao, Q.; Guo, Y.; Lu, Z.; Ma, W.; Liu, P.; Pestell, R.G.; Liang, C.; et al. piRNA-823 Is Involved in Cancer Stem Cell Regulation Through Altering DNA Methylation in Association With Luminal Breast Cancer. Front. Cell Dev. Biol. 2021, 9, 641052. [Google Scholar] [CrossRef]
- McKerrow, W.; Wang, X.; Mendez-Dorantes, C.; Mita, P.; Cao, S.; Grivainis, M.; Ding, L.; LaCava, J.; Burns, K.H.; Boeke, J.D.; et al. LINE-1 expression in cancer correlates with p53 mutation, copy number alteration, and S phase checkpoint. Proc. Natl. Acad. Sci. USA 2022, 119, e2115999119. [Google Scholar] [CrossRef]
- Ren, W.; Fan, H.; Grimm, S.A.; Kim, J.J.; Li, L.; Guo, Y.; Petell, C.J.; Tan, X.-F.; Zhang, Z.-M.; Coan, J.P.; et al. DNMT1 reads heterochromatic H4K20me3 to reinforce LINE-1 DNA methylation. Nat. Commun. 2021, 12, 2490. [Google Scholar] [CrossRef] [PubMed]
- Jia, D.D.; Jiang, H.; Zhang, Y.F.; Zhang, Y.; Qian, L.L.; Zhang, Y.F. The regulatory function of piRNA/PIWI complex in cancer and other human diseases: The role of DNA methylation. Int. J. Biol. Sci. 2022, 18, 3358–3373. [Google Scholar] [CrossRef] [PubMed]
- Fu, A.; Jacobs, D.I.; Zhu, Y. Epigenome-wide analysis of piRNAs in gene-specific DNA methylation. RNA Biol. 2014, 11, 1301–1312. [Google Scholar] [CrossRef] [Green Version]
- Fu, A.; Jacobs, D.I.; Hoffman, A.E.; Zheng, T.; Zhu, Y. PIWI-interacting RNA 021285 is involved in breast tumorigenesis possibly by remodeling the cancer epigenome. Carcinogenesis 2015, 36, 1094–1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hombach, S.; Kretz, M. Non-coding RNAs: Classification, Biology and Functioning. Adv. Exp. Med. Biol. 2016, 937, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Gupta, T.; Malkin, M.G.; Huang, S. tRNA Function and Dysregulation in Cancer. Front. Cell Dev. Biol. 2022, 10, 886642. [Google Scholar] [CrossRef]
- Martinez, A.; Yamashita, S.; Nagaike, T.; Sakaguchi, Y.; Suzuki, T.; Tomita, K. Human BCDIN3D monomethylates cytoplasmic histidine transfer RNA. Nucleic Acids Res. 2017, 45, 5423–5436. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, Y.; Yano, H.; Ogasawara, S.; Yoshioka, S.-I.; Imamura, H.; Okamoto, K.; Tsuneoka, M. Mild Glucose Starvation Induces KDM2A-Mediated H3K36me2 Demethylation through AMPK To Reduce rRNA Transcription and Cell Proliferation. Mol. Cell. Biol. 2015, 35, 4170–4184. [Google Scholar] [CrossRef] [Green Version]
- Shah, D.; Gandhi, M.; Kumar, A.; Cruz-Martins, N.; Sharma, R.; Nair, S. Current insights into epigenetics, noncoding RNA interactome and clinical pharmacokinetics of dietary polyphenols in cancer chemoprevention. Crit. Rev. Food Sci. Nutr. 2021. [Google Scholar] [CrossRef]
- Montgomery, M.; Srinivasan, A. Epigenetic Gene Regulation by Dietary Compounds in Cancer Prevention. Adv. Nutr. 2019, 10, 1012–1028. [Google Scholar] [CrossRef] [Green Version]
- Shankar, E.; Kanwal, R.; Candamo, M.; Gupta, S. Dietary phytochemicals as epigenetic modifiers in cancer: Promise and challenges. Semin. Cancer Biol. 2016, 40–41, 82–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fatima, N.; Baqri, S.S.R.; Bhattacharya, A.; Koney, N.K.-K.; Husain, K.; Abbas, A.; Ansari, R.A. Role of Flavonoids as Epigenetic Modulators in Cancer Prevention and Therapy. Front. Genet. 2021, 12, 758733. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, G.; Carcache, P.J.B.; Addo, E.M.; Kinghorn, A.D. Current status and contemporary approaches to the discovery of antitumor agents from higher plants. Biotechnol. Adv. 2020, 38, 107337. [Google Scholar] [CrossRef]
- Jiang, F.; Li, Y.; Mu, J.; Hu, C.; Zhou, M.; Wang, X.; Si, L.; Ning, S.; Li, Z. Glabridin inhibits cancer stem cell-like properties of human breast cancer cells: An epigenetic regulation of miR-148a/SMAd2 signaling. Mol. Carcinog. 2016, 55, 929–940. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Ji, H.W.; Kim, H.W.; Yun, S.H.; Park, J.E.; Kim, S.J. Ginsenoside Rg3 Prevents Oncogenic Long Noncoding RNA ATXN8OS from Inhibiting Tumor-Suppressive microRNA-424-5p in Breast Cancer Cells. Biomolecules 2021, 11, 118. [Google Scholar] [CrossRef]
- Han, B.; Peng, X.; Cheng, D.; Zhu, Y.; Du, J.; Li, J.; Yu, X. Delphinidin suppresses breast carcinogenesis through the HOTAIR/microRNA-34a axis. Cancer Sci. 2019, 110, 3089–3097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, X.; Chang, H.; Chen, J.; Zhang, Q.; Yu, X.; Mi, M. 3,6-Dihydroxyflavone regulates microRNA-34a through DNA methylation. BMC Cancer 2017, 17, 619. [Google Scholar] [CrossRef] [Green Version]
- Su, Z.; Wang, C.; Chang, D.; Zhu, X.; Sai, C.; Pei, J. Limonin attenuates the stemness of breast cancer cells via suppressing MIR216A methylation. Biomed. Pharmacother. 2019, 112, 108699. [Google Scholar] [CrossRef]
- Jeong, D.; Ham, J.; Park, S.; Kim, H.W.; Kim, H.; Ji, H.W.; Kim, S.J. Ginsenoside Rh2 Suppresses Breast Cancer Cell Proliferation by Epigenetically Regulating the Long Noncoding RNA C3orf67-AS1. Am. J. Chin. Med. 2019, 47, 1643–1658. [Google Scholar] [CrossRef]
- Golshan, M.; Khaleghi, S.; Shafiee, S.M.; Valaee, S.; Ghanei, Z.; Jamshidizad, A.; Dashtizad, M.; Shamsara, M. Metformin modulates oncogenic expression of HOTAIR gene via promoter methylation and reverses epithelial-mesenchymal transition in MDA-MB-231 cells. J. Cell. Biochem. 2021, 122, 385–393. [Google Scholar] [CrossRef]
- Si, L.; Jiang, F.; Li, Y.; Ye, X.; Mu, J.; Wang, X.; Ning, S.; Hu, C.; Li, Z. Induction of the mesenchymal to epithelial transition by demethylation- activated microRNA-200c is involved in the anti-migration/invasion effects of arsenic trioxide on human breast cancer cells. Mol. Carcinog. 2015, 54, 859–869. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Zhang, X.; Liu, X.; Wang, P.; Chu, W.; Zhao, W.; Wang, Y.; Zhou, G.; Yu, Y.; Zhang, H. The DNMT1-PAS1-PH20 axis drives breast cancer growth and metastasis. Signal Transduct. Target. Ther. 2022, 7, 81. [Google Scholar] [CrossRef] [PubMed]
- Dhillon, S. Decitabine/Cedazuridine: First Approval. Drugs 2020, 80, 1373–1378. [Google Scholar] [CrossRef] [PubMed]
- Cordero, F.; Ferrero, G.; Polidoro, S.; Fiorito, G.; Campanella, G.; Sacerdote, C.; Mattiello, A.; Masala, G.; Agnoli, C.; Frasca, G.; et al. Differentially methylated microRNAs in prediagnostic samples of subjects who developed breast cancer in the European Prospective Investigation into Nutrition and Cancer (EPIC-Italy) cohort. Carcinogenesis 2015, 36, 1144–1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manoochehri, M.; Jones, M.; Tomczyk, K.; Fletcher, O.; Schoemaker, M.J.; Swerdlow, A.J.; Borhani, N.; Hamann, U. DNA methylation of the long intergenic noncoding RNA 299 gene in triple-negative breast cancer: Results from a prospective study. Sci. Rep. 2020, 10, 11762. [Google Scholar] [CrossRef] [PubMed]
- Bermejo, J.L.; Huang, G.; Manoochehri, M.; Mesa, K.G.; Schick, M.; Silos, R.G.; Ko, Y.D.; Bruning, T.; Brauch, H.; Lo, W.Y.; et al. Long intergenic noncoding RNA 299 methylation in peripheral blood is a biomarker for triple-negative breast cancer. Epigenomics 2019, 11, 81–93. [Google Scholar] [CrossRef]
- Oltra, S.S.; Pena-Chilet, M.; Vidal-Tomas, V.; Flower, K.; Martinez, M.T.; Alonso, E.; Burgues, O.; Lluch, A.; Flanagan, J.M.; Ribas, G. Methylation deregulation of miRNA promoters identifies miR124-2 as a survival biomarker in Breast Cancer in very young women. Sci. Rep. 2018, 8, 14373. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Martinez, L.; Zhang, Y.; Nakata, Y.; Chan, H.L.; Morey, L. Epigenetic mechanisms in breast cancer therapy and resistance. Nat. Commun. 2021, 12, 1786. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MiRNAs | Epigenetic Mechanism | Expression in BC | Target Gene or Protein/Pathway Involved | Biological Function | Ref. |
|---|---|---|---|---|---|
| MiR-362-3p miRNA-329 | Regulation of miRNA expression by DNA methylation | ↓ | MiR-362-3p/miRNA-329- p130Cas | Inhibiting proliferation, migration, and invasion | [29] |
| MiR-195/497 | ↓ | MiR-195-497/Raf-1 | Suppressing cell proliferation and invasion | [30] | |
| MiR-892b | ↓ | MiR-892b/NF-κB | Inhibiting aggressiveness | [31] | |
| MiR-200b | ↓ | Kindlin 2/DNMT3A/miRNA-200b | Inhibiting invasion | [32] | |
| MiR-200a-5p | ↓ | FEN1/PCNA/DNMT3a/miR-200a-5p | Inhibiting proliferation | [33] | |
| MiR-193a-3p | ↓ | MiR-193a-3p/GRB7 | Inhibiting HER2 positive BC aggressiveness | [34] | |
| MiR-205 | ↓ | Mel-18/DNMT/miR-205 | Inhibiting EMT | [35] | |
| MiR-34 miR-199a/b | ↓ | MiR-34-miR-199a/b/Axl | Inhibiting migration and invasion | [36] | |
| MiR-335 | ↓ | Not applicable | Inhibiting tumor reinitiation | [37] | |
| MiR-126 miR-126 ∗ | ↓ | MiR-126-miR-126 ∗/SDF-1α | Inhibiting metastasis | [38] | |
| MiR-100 | ↓ | MiR-100/SMARCA5 MiR-100/HOXA1 | EMT-inducing and tumor-suppressing effects | [39] | |
| MiR-125b | ↓ | MiR-125b/ETS1 | Suppressing proliferation | [40] | |
| MiR-10b∗ | ↓ | MiR-10b∗/BUB1, PLK1 and CCNA2 | Inhibiting proliferation | [41] | |
| MiR-34c | ↓ | MiR-34c/Notch4 | Inhibiting tumor-initiating cells self-renewal | [42] | |
| MiR-200c-141 | ↓ | Not applicable | Decreasing stem-like properties | [43] | |
| MiR-663 | ↑ | MiR-663/HSPG2 | Inducing chemotherapy resistance | [44] | |
| MiR-320a | ↓ | MiR-320a/TRPC5 and NFATC3 | Decreasing chemoresistance | [45] | |
| MiR-149 | ↓ | MiR-149/NDST1 | Modulating chemoresistance | [46] | |
| MiR-375 | ↓ | MiR-375/IGF1R | Inhibiting trastuzumab resistance | [47] | |
| MiR-29c | ↓ | Not applicable | Involving in invasion and chemotherapy sensitivity | [48] | |
| MiR-9 | ↓ | DNMT3A/miR-9/VEGFA | Regulating mechanical compression | [49] | |
| MiR-200c | ↓ | MiR-21a/miR-200c/PTEN | Inhibiting the transformation of M2 macrophages | [50] | |
| MiR-125a | ↓ | Not applicable | Reducing autophagy | [51] | |
| MiR-148b MiR-29c MiR-26b | Regulation of DNA methylation by miRNA | ↓ | MiRNAs/DNMT3B | Not applicable | [52] |
| MiR-29c | ↓ | MiR-29c/DNMT3B/TIMP3/STAT1/FOXO1 | Inhibiting aggressiveness | [53] | |
| MiR-29b-1-5p | ↓ | MiR-29b-1-5p/DNMTs | Inhibiting cell proliferation | [54] | |
| MiR-770-5p | ↓ | MiR-770-5p/DNMT3A | Inhibiting EMT | [55] | |
| MiR-148a/152 | ↓ | MiR-148a/152/DNMT1 regulatory circuit | Inhibiting aggressiveness | [56] | |
| MiR-200b | ↓ | MiR-200b/DNMT3A regulatory circuit | Inhibiting aggressiveness | [57] | |
| MiR-133a-3p | ↓ | MiR-133a-3p/MAML1/DNMT3A regulatory circuit | Inhibiting migration and stemness | [58] | |
| MiR-10a | ↑ | MiR-10a/Hoxd4 | Not applicable | [59] | |
| MiR-146a-5p | M6A modification of miRNA | ↑ | METTL14/miR-146a-5p | Promoting migration and invasion | [60] |
| MiR-221-3p | ↑ | METTL3/miR-221-3p/HIPK2/Che-1 | Inducing chemotherapy resistance | [61] | |
| MiR-375 | Regulation of miRNA expression by histone methylation | ↑ | MiR-375/RASD1 | Promoting cell proliferation | [62] |
| MiR-200c | ↓ | MiR-200c/ZEB1 | Inhibiting lymph node metastasis | [63] | |
| MiR let-7e | ↓ | JARID1B/let-7e/cyclin D1 | Inhibiting proliferation | [64] | |
| MiR-7 | Regulation of histone methylation by miRNA | ↓ | MiR-7/SET8 | Inhibiting EMT and invasiveness | [65] |
| MiR-491-5p | ↓ | MiR-491-5p/JMJD2B | Inhibiting proliferation | [66] | |
| MiR-29a | ↑ | MiR-29a/SUV420H2 | Promoting EMT, migration, and invasion | [67] |
| LncRNAs | Epigenetic Mechanism | Expression in BC | Target Gene or Protein/Pathway Involved | Biological Function | Ref. |
|---|---|---|---|---|---|
| LncRNA APOC1P1-3 | Regulation of lncRNA expression by DNA methylation | ↑ | APOC1P1-3/α-tubulin acetylation | Inhibiting apoptosis | [75] |
| LncRNA EPIC1 | ↑ | LncRNA EPIC1/MYC | Promoting Cell-Cycle progression | [76] | |
| LncRNA HUMT | ↑ | HUMT/YBX1/FOXK1 axis | Promoting lymphangiogenesis and metastasis | [77] | |
| LncRNA GAS5 | ↓ | Not applicable | Promoting chemosensitivity and apoptosis | [78] | |
| LncRNA MEG3 | ↓ | Not applicable | Promoting chemosensitivity | [79] | |
| LncRNA MYMA | Regulation of DNA methylation by lncRNA | ↑ | ROR1/HER3/lncRNA MYMA axis | Promoting bone metastasis | [80] |
| LncRNA 91H | ↑ | LncRNA 91H -H19/IGF2 | Increasing aggressive phenotype | [81] | |
| LncRNA PYCARD-AS1 | Not applicable | LncRNA PYCARD-AS1/PYCARD | Regulating apoptosis | [82] | |
| LINC00518 | ↑ | LINC00518/CDX2/Wnt signaling | Promoting epithelial cell growth and metastasis | [83] | |
| LncRNA H19 | ↑ | H19/SAHH/DNMT3B axis | Promoting tamoxifen resistance | [84] | |
| LncRNA BCLIN25 | ↑ | BCLIN25/miR-125b/ERBB2 axis | Promoting tumorigenesis | [85] | |
| LncRNA MIAT | ↑ | LncRNA MIAT/DLG3/Hippo signaling pathway | Promoting tumorigenesis | [86] | |
| LINC00472 | ↓ | LINC00472/MCM6/MEK/ERK signaling pathway | Inhibiting metastasis | [87] | |
| LncRNA TINCR | ↑ | STAT3/TINCR/EGFR feedback loop | Promoting tumorigenesis | [88] | |
| LncRNA LINC00922 | ↑ | LINC00922/NKD2/Wnt signaling pathway | Promoting EMT, invasive and migratory capacities | [89] | |
| LncRNA MAGI2-AS3 | ↓ | MAGI2-AS3/MAGI2/Wnt/β-catenin pathway | Inhibiting cell proliferation and migration | [90] | |
| LncRNA SNHG10 | ↓ | LncRNA SNHG10/miR-302b | Promoting chemosensitivity of TNBC cells to doxorubicin. | [91] | |
| LNC942 | Regulation of m6A modification by lncRNA | ↑ | LNC942/METTL14/CXCR4 and CYP1B1 signaling axis | Promoting proliferation and inhibiting apoptosis | [92] |
| LINC00675 | M6A modification of lncRNA | ↓ | METTL3/LINC00675/miR-513b-5p | Suppressing proliferation, migration, and invasion | [93] |
| LncRNA DLEU1 | Regulation of lncRNA expression by histone methylation | ↑ | Not applicable | Promoting tumorigenesis | [94] |
| LncRNA EPB41L4A-AS2 | ↓ | ZNF217/EPB41L4A-AS2/RARRES1 | Inhibiting proliferation, migration, and invasion and inducing apoptosis | [95] | |
| LncRNA HOTAIR | ↑ | Not applicable | Associating with endocrine disruption | [96] | |
| LncRNA UCA1 | Not applicable | SATB1/lncRNA UCA1 | Promoting growth and metastasis | [97] | |
| LncRNA HOTAIR | Regulation of histone methylation by lncRNA | ↑ | Not applicable | Promoting invasiveness and metastasis | [98] |
| LncRNA UCA1 | ↑ | LncRNA UCA1/EZH2/p21 axis | Inducing tamoxifen resistance | [99] | |
| LINC00511 | ↑ | LINC00511/EZH2/CDKN1B | Promoting proliferation and inhibiting apoptosis | [100] | |
| LncRNA ROR | ↑ | LncRNA ROR/MLL1/TIMP3 | Promoting BC progression | [101] | |
| LncRNA PHACTR2-AS1 | ↓ | EZH2/lncRNA PHACTR2-AS1 | Inhibiting growth and metastasis | [102] |
| NcRNAs | Epigenetic Mechanism | Expression in BC | Target Gene or Protein/Pathway Involved | Biological Function | Ref. |
|---|---|---|---|---|---|
| CircBACH2 | CircRNA indirectly regulates m6A modification | ↑ | CircBACH2/hsa-miR-944/HNRNPC axis | Promoting cell proliferation | [106] |
| Endo-siRNA | Regulation of DNA methylation by endo-siRNA | ↓ | Endo-siRNA/LINE-1 | Maintaining a stable genome structure | [107] |
| PiRNA-651 | Regulation of DNA methylation by piRNA | ↑ | PIWI/piRNA-651/DNMT1/PTEN | Promoting cell proliferation and migration and inhibiting apoptosis | [108] |
| PiR-823 | ↑ | PiR-823/DNMT/APC/Wnt signaling | Promoting cell proliferation and being involved in stem cell regulation | [109] |
| ncRNA | Therapeutic Substance | Epigenetic Mechanism | Target Gene or Protein/Pathway Involved | Biological Function | Ref. |
|---|---|---|---|---|---|
| MiR-148a | Glabridin | DNA methylation | MiR-148a/TGFβ/SMAd2 | Stemness | [124] |
| LncRNA ATXN8OS | Ginsenoside Rg3 | DNA methylation | LncRNA ATXN8OS/miR-424-5p | Proliferation | [125] |
| MiR-34a | Delphinidin | H3K27me3 | HOTAIR/miRNA-34a | Carcinogenesis | [126] |
| MiR-34a | 3,6-Dihydroxyflavone | DNA methylation | PI3K/Akt/mTOR | Carcinogenesis | [127] |
| MiR-216a-3p | Limonin | DNA methylation | MiR-216a-3p/WNT3A | Stemness | [128] |
| LncRNA C3orf67-AS1 | Ginsenoside Rh2 | DNA methylation | Not applicable | Proliferation | [129] |
| LncRNA HOTAIR | Metformin | DNA methylation | Not applicable | EMT | [130] |
| miR-200c | Arsenic trioxide | DNA methylation | Not applicable | Migration and invasion | [131] |
| LncRNA PAS1 | Decitabine | DNA methylation | DNMT1/PAS1/PH20 | Growth and metastasis | [132] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Leng, P.; Liu, Y.; Guo, J.; Zhou, H. Crosstalk between Methylation and ncRNAs in Breast Cancer: Therapeutic and Diagnostic Implications. Int. J. Mol. Sci. 2022, 23, 15759. https://doi.org/10.3390/ijms232415759
Liu Y, Leng P, Liu Y, Guo J, Zhou H. Crosstalk between Methylation and ncRNAs in Breast Cancer: Therapeutic and Diagnostic Implications. International Journal of Molecular Sciences. 2022; 23(24):15759. https://doi.org/10.3390/ijms232415759
Chicago/Turabian StyleLiu, Yitong, Ping Leng, Yan Liu, Jinlin Guo, and Hao Zhou. 2022. "Crosstalk between Methylation and ncRNAs in Breast Cancer: Therapeutic and Diagnostic Implications" International Journal of Molecular Sciences 23, no. 24: 15759. https://doi.org/10.3390/ijms232415759