
Current Insight into the Role of IL-35 and Its Potential Involvement in the Pathogenesis and Therapy of Atopic Dermatitis

Abstract
:1. Introduction
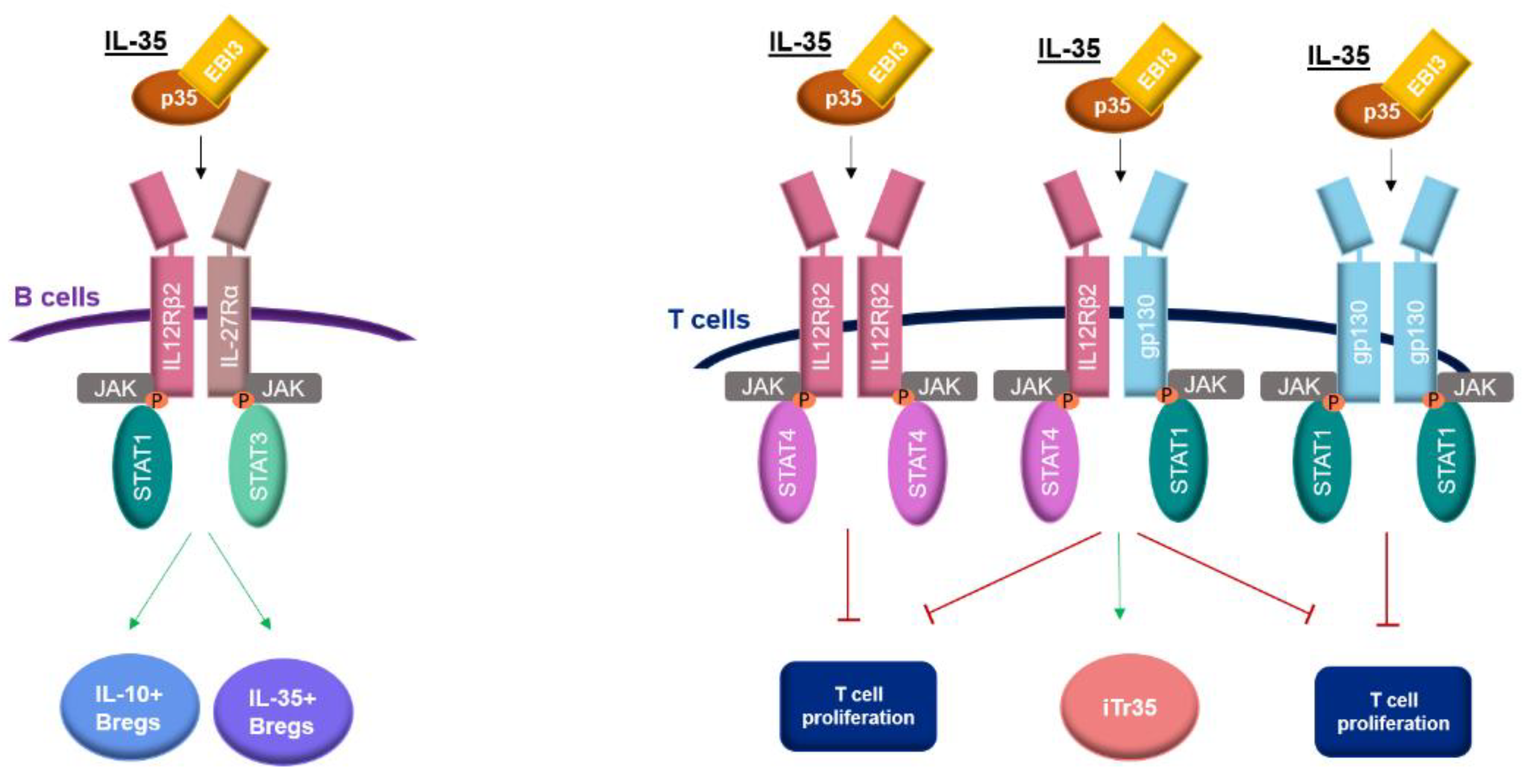
2. Receptor and Signaling Pathway of IL-35
3. Expression of IL-35 in Human Diseases
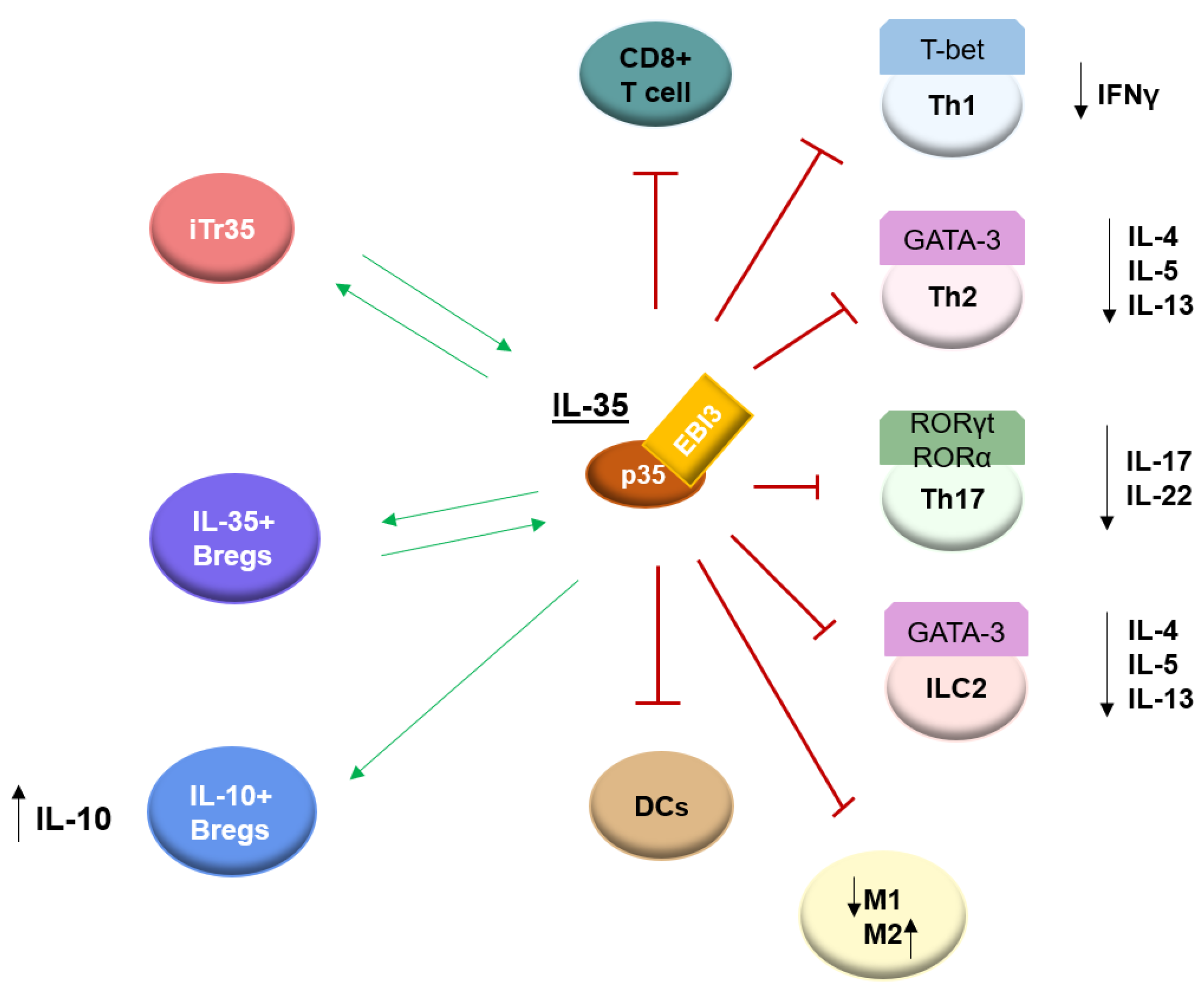
4. Biological Function of IL-35
5. IL-35-Induced Regulatory T Cells (iTr35)
6. Regulatory B Cells (Bregs)
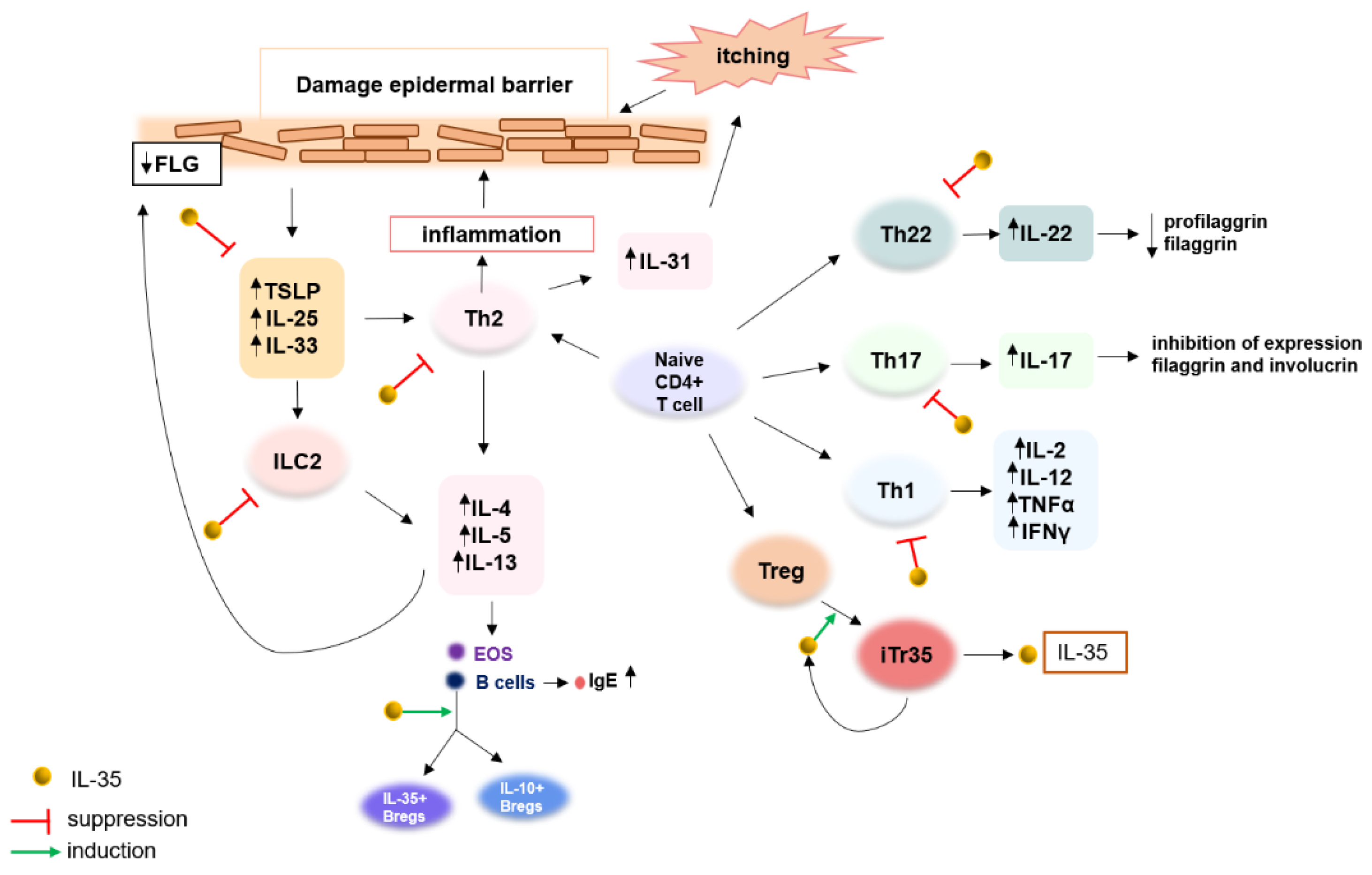
7. Immunological Imbalance in Atopic Dermatitis
8. IL-35 as a Novel Regulatory Cytokine and Its Potential Involvement in Pathogenesis and Therapeutic Effects in Atopic Dermatitis
9. Regulatory T Cells in Atopic Dermatitis
10. Regulatory B Cells in Atopic Dermatitis
11. Discussion
12. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vignali, D.A.A.; Kuchroo, V.K. IL-12 family cytokines: Immunological playmakers. Nat. Immunol. 2012, 13, 722–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collison, L.W.; Workman, C.J.; Kuo, T.T.; Boyd, K.; Wang, Y.; Vignali, K.M.; Cross, R.; Sehy, D.; Blumberg, R.S.; Vignali, D.A.A. The inhibitory cytokine IL-35 contributes to regulatory T-cell function. Nature 2007, 450, 566–569. [Google Scholar] [CrossRef] [PubMed]
- Su, L.C.; Liu, X.Y.; Huang, A.F.; Xu, W.D. Emerging role of IL-35 in inflammatory autoimmune diseases. Autoimmun. Rev. 2018, 17, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Behzadi, P.; Behzadi, E.; Ranjbar, R. IL-12 Family Cytokines: General Characteristics, Pathogenic Microorganisms, Receptors, and Signalling Pathways. Acta Microbiol. Immunol. Hung. 2016, 63, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Bardel, E.; Larousserie, F.; Charlot-Rabiega, P.; Coulomb-L’Herminé, A.; Devergne, O. Human CD4+ CD25+ Foxp3+ regulatory T cells do not constitutively express IL-35. J. Immunol. Baltim. 2008, 181, 6898–6905. [Google Scholar] [CrossRef] [Green Version]
- Guttek, K.; Reinhold, D. Stimulated human peripheral T cells produce high amounts of IL-35 protein in a proliferation-dependent manner. Cytokine 2013, 64, 46–50. [Google Scholar] [CrossRef]
- Collison, L.W.; Pillai, M.R.; Chaturvedi, V.; Vignali, D.A.A. Regulatory T cell suppression is potentiated by target T cells in a cell contact, IL-35- and IL-10-dependent manner. J. Immunol. Baltim. 2009, 182, 6121–6128. [Google Scholar] [CrossRef] [Green Version]
- Collison, L.W.; Chaturvedi, V.; Henderson, A.L.; Giacomin, P.R.; Guy, C.; Bankoti, J.; Finkelstein, D.I.; Forbes, K.; Workman, C.J.; Brown, S.A.; et al. IL-35-mediated induction of a potent regulatory T cell population. Nat. Immunol. 2010, 11, 1093–1101. [Google Scholar] [CrossRef] [Green Version]
- Shen, P.; Roch, T.; Lampropoulou, V.; O’Connor, R.A.; Stervbo, U.; Hilgenberg, E.; Ries, S.; Dang, V.D.; Jaimes, Y.; Daridon, C.; et al. IL-35-producing B cells are critical regulators of immunity during autoimmune and infectious diseases. Nature 2014, 507, 366–370. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.K.; Egwuagu, C.E. Interleukin 35 Regulatory B Cells. J. Mol. Biol. 2021, 433, 166607. [Google Scholar] [CrossRef]
- Li, X.; Mai, J.; Virtue, A.; Yin, Y.; Gong, R.; Sha, X.; Gutchigian, S.; Frisch, A.; Hodge, I.; Jiang, X.; et al. IL-35 is a novel responsive anti-inflammatory cytokine—A new system of categorizing anti-inflammatory cytokines. PLoS ONE 2012, 7, e33628. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.; Jiang, H.; Li, Y.; Yan, M.X. IL-35 expression is increased in laryngeal squamous cell carcinoma and in the peripheral blood of patients. Oncol. Lett. 2017, 13, 3303–3308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Hao, S.; Chen, X.; Zhao, H.; Du, L.; Ren, H.; Wang, C.; Mao, H. Human placental trophoblast cells contribute to maternal-fetal tolerance through expressing IL-35 and mediating iTR35 conversion. Nat. Commun. 2019, 10, 4601. [Google Scholar] [CrossRef] [Green Version]
- Bello, R.O.; Chin, V.K.; Isnadi, M.F.A.R.; Majid, R.A.; Abdullah, M.A.; Lee, T.Y.; Zakaria, Z.A.; Hussain, M.K.; Basir, R. The Role, Involvement and Function(s) of Interleukin-35 and Interleukin-37 in Disease Pathogenesis. Int. J. Mol. Sci. 2018, 19, 1149. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.R.; Yadav, M.K.; Kang, M.; Jittayasothorn, Y.; Dong, L.; Egwuagu, C.E. Photoreceptor Cells Constitutively Express IL-35 and Promote Ocular Immune Privilege. Int. J. Mol. Sci. 2022, 23, 8156. [Google Scholar] [CrossRef]
- Collison, L.W.; Delgoffe, G.M.; Guy, C.S.; Vignali, K.M.; Chaturvedi, V.; Fairweather, D.; Satoskar, A.R.; Garcia, K.C.; Hunter, C.A.; Drake, C.G.; et al. The composition and signaling of the IL-35 receptor are unconventional. Nat. Immunol. 2012, 13, 290–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szabo, S.J.; Dighe, A.S.; Gubler, U.; Murphy, K.M. Regulation of the interleukin (IL)-12R beta 2 subunit expression in developing T helper 1 (Th1) and Th2 cells. J. Exp. Med. 1997, 185, 817–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietrich, C.; Candon, S.; Ruemmele, F.M.; Devergne, O. A soluble form of IL-27Rα is a natural IL-27 antagonist. J. Immunol. Baltim. 2014, 192, 5382–5389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silver, J.S.; Hunter, C.A. gp130 at the nexus of inflammation, autoimmunity, and cancer. J. Leukoc. Biol. 2010, 88, 1145–1156. [Google Scholar] [CrossRef] [Green Version]
- Qiu, F.; Song, L.; Yang, N.; Li, X. Glucocorticoid downregulates expression of IL-12 family cytokines in systemic lupus erythematosus patients. Lupus 2013, 22, 1011–1016. [Google Scholar] [CrossRef]
- Šenolt, L.; Šumová, B.; Jandová, R.; Hulejová, H.; Mann, H.; Pavelka, K.; Vencovsky, J.; Filkova, M. Interleukin 35 Synovial Fluid Levels Are Associated with Disease Activity of Rheumatoid Arthritis. PLoS ONE 2015, 10, e0132674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abushouk, A.; Alkhalaf, H.; Aldamegh, M.; Bin Shigair, S.; Mahabbat, N.; Hakami, M.; Abu-Jaffal, A.S.; Nasr, A. IL-35 and IL-37 are negatively correlated with high IgE production among children with asthma in Saudi Arabia. J. Asthma. 2022, 59, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.K.; Leung, T.F.; Chu, I.M.T.; Dong, J.; Lam, Y.Y.O.; Lam, C.W.K. Aberrant expression of regulatory cytokine IL-35 and pattern recognition receptor NOD2 in patients with allergic asthma. Inflammation 2015, 38, 348–360. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Li, P.; Yang, J. Decreased Circulating Interleukin-35 Levels Are Related to Interleukin-4-Producing CD8+ T Cells in Patients with Allergic Asthma. Iran. J. Allergy Asthma Immunol. 2015, 14, 379–385. [Google Scholar] [PubMed]
- Khoshkhui, M.; Alyasin, S.; Sarvestani, E.K.; Amin, R.; Ariaee, N. Evaluation of serum interleukin- 35 level in children with persistent asthma. Asian Pac. J. Allergy Immunol. 2017, 35, 91–95. [Google Scholar] [CrossRef]
- Ma, Y.; Liu, X.; Wei, Z.; Wang, X.; Xu, D.; Dai, S.; Li, Y.; Gao, M.; Ji, C.; Guo, C.; et al. The expression of a novel anti-inflammatory cytokine IL-35 and its possible significance in childhood asthma. Immunol Lett. 2014, 162, 11–17. [Google Scholar] [CrossRef]
- Wang, W.; Wei, C.; Cheng, Z.; Yang, J. Aberrant Th2 Immune Responses Are Associated With a Reduced Frequency of IL-35-Induced Regulatory T Cells After Allergen Exposure in Patients With Allergic Asthma. Allergy Asthma Immunol. Res. 2020, 12, 1029–1045. [Google Scholar] [CrossRef]
- Xie, F.; Hu, Q.; Cai, Q.; Yao, R.; Ouyang, S. IL-35 Inhibited Th17 Response in Children with Allergic Rhinitis. ORL 2020, 82, 47–52. [Google Scholar] [CrossRef]
- Huang, L.; Zhao, M.; Luo, Q.; Liang, K.; Iu, C.L. The role and correlation of IL-35 and type II intrinsic lymphocytes in children with allergic rhinitis. Cell. Mol. Biol. 2021, 67, 127–131. [Google Scholar] [CrossRef]
- Chen, T.; Fu, L.X.; Sun, Q.M.; Zhou, P.M.; Guo, Z.P. Decreased interleukin-35 serum levels in patients with chronic spontaneous urticaria. Ann. Allergy Asthma Immunol. 2018, 121, 503–504. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Y.; Liu, Y.; Wang, Y.; Zuo, X.; Li, Y.; Lu, X. The possible role of the novel cytokines il-35 and il-37 in inflammatory bowel disease. Mediat. Inflamm. 2014, 2014, 136329. [Google Scholar] [CrossRef] [PubMed]
- LLi, T.; Gu, M.; Liu, P.; Liu, Y.; Guo, J.; Zhang, W.; Qian, C.; Deng, A. Clinical Significance of Decreased Interleukin-35 Expression in Patients with Psoriasis. Microbiol. Immunol. 2018. [Google Scholar] [CrossRef] [Green Version]
- Elbana, A.M.; Elgamal, E.; Hashim, O.; Emran, T.M.; Alkhrsawy, A.A. Pro-inflammatory versus anti-inflammatory cytokines in psoriatic patients (case-control study). J. Cosmet. Dermatol. 2022. Online ahead of print. [Google Scholar] [CrossRef]
- Ye, Z.; Jiang, Y.; Sun, D.; Zhong, W.; Zhao, L.; Jiang, Z. The Plasma Interleukin (IL)-35 Level and Frequency of Circulating IL-35+ Regulatory B Cells are Decreased in a Cohort of Chinese Patients with New-onset Systemic Lupus Erythematosus. Sci. Rep. 2019, 9, 13210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Yao, L.; Liu, S.; Wu, J.; Xia, L.; Shen, H.; Lu, J. Elevated serum IL-35 levels in rheumatoid arthritis are associated with disease activity. J. Investig. Med. 2019, 67, 707–710. [Google Scholar] [CrossRef] [PubMed]
- Kiwan, A.H.; Mohamed, H.A.K.; Hashim, O.A.E.A.; Abd-Elraheem, S.I.; Alkhrsawy, A.M. Pro-Inflammatory versus Anti-Inflammatory cytokines in atopic dermatitis patients: A case control study. J. Cosmet. Dermatol. 2022. Online ahead of print. [Google Scholar] [CrossRef]
- Liu, Y.; Wu, Y.; Wang, Y.; Cai, Y.; Hu, B.; Bao, G.; Fang, H.; Zhao, L.; Ma, S.; Cheng, Q.; et al. IL-35 mitigates murine acute graft-versus-host disease with retention of graft-versus-leukemia effects. Leukemia 2015, 29, 939–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, H.; Zhang, T.; Yan, M.X.; Wu, W. IL-35 inhibits CD8+ T cells activity by suppressing expression of costimulatory molecule CD28 and Th1 cytokine production. Transl. Cancer Res. 2019, 8, 1319–1325. [Google Scholar] [CrossRef]
- Zhu, J. GATA3 Regulates the Development and Functions of Innate Lymphoid Cell Subsets at Multiple Stages. Front. Immunol. 2017, 8, 1571. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Y.; Gu, W.; Sun, B. TH1/TH2 cell differentiation and molecular signals. Adv. Exp. Med. Biol. 2014, 841, 15–44. [Google Scholar] [CrossRef]
- Liu, W.; Zeng, Q.; Wen, Y.; Tang, Y.; Yan, S.; Li, Y.; Zhou, L.; Luo, R. Inhibited interleukin 35 expression and interleukin 35-induced regulatory T cells promote type II innate lymphoid cell response in allergic rhinitis. Ann. Allergy Asthma Immunol. 2021, 126, 152–161.e1. [Google Scholar] [CrossRef]
- Nie, M.; Zeng, Q.; Xi, L.; Tang, Y.; Luo, R.; Liu, W. The Effect of IL-35 on the Expression of Nasal Epithelial-Derived Proinflammatory Cytokines. Mediators Inflamm. 2021, 2021, 1110671. [Google Scholar] [CrossRef]
- Huang, C.H.; Loo, E.X.; Kuo, I.C.; Soh, G.H.; Goh, D.L.; Lee, B.W.; Chua, K.Y. Airway inflammation and IgE production induced by dust mite allergen-specific memory/effector Th2 cell line can be effectively attenuated by IL-35. J. Immunol. 2011, 187, 462–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Yang, M.; Htut, T.M.; Ouyang, X.; Hanidu, A.; Li, X.; Sellati, R.; Jiang, H.; Zhang, S.; Li, H.; et al. Epstein-Barr virus-induced gene 3 negatively regulates IL-17, IL-22 and RORgamma t. Eur. J. Immunol. 2008, 38, 1204–1214. [Google Scholar] [CrossRef] [PubMed]
- Okada, K.; Fujimura, T.; Kikuchi, T.; Aino, M.; Kamiya, Y.; Izawa, A.; Iwamura, Y.; Goto, H.; Okabe, I.; Miyake, E.; et al. Effect of interleukin (IL)-35 on IL-17 expression and production by human CD4+ T cells. PeerJ 2017, 5, e2999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, A.; Zhang, Y.; Wang, X.; Cui, Y.; Tan, W. Interleukin 35 regulates interleukin 17 expression and T helper 17 in patients with proliferative diabetic retinopathy. Bioengineered 2022, 13, 13293–13299. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Hao, S.; Zhao, Z.; Liu, J.; Shao, Q.; Wang, F.; Sun, D.; He, Y.; Gao, W.; Mao, H. Interleukin 35: Inhibitory regulator in monocyte-derived dendritic cell maturation and activation. Cytokine 2018, 108, 43–52. [Google Scholar] [CrossRef]
- Luo, Z.; Soläng, C.; Larsson, R.; Singh, K. Interleukin-35 Prevents the Elevation of the M1/M2 Ratio of Macrophages in Experimental Type 1 Diabetes. Int. J. Mol. Sci. 2022, 23, 7970. [Google Scholar] [CrossRef]
- Zhang, J.; Lin, Y.; Li, C.; Zhang, X.; Cheng, L.; Dai, L.; Wang, Y.; Wang, F.; Shi, G.; Li, Y.; et al. IL-35 Decelerates the Inflammatory Process by Regulating Inflammatory Cytokine Secretion and M1/M2 Macrophage Ratio in Psoriasis. J. Immunol. Baltim. 2016, 197, 2131–2144. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.; Huang, A.; Nie, J.; Tan, J.; Xing, S.; Qu, Y.; Jiang, K. IL-35 Regulates the Function of Immune Cells in Tumor Microenvironment. Front. Immunol. 2021, 12, 683332. [Google Scholar] [CrossRef]
- Tao, N.-N.; Gong, R.; Chen, X.; He, L.; Ren, F.; Yu, H.-B.; Chen, J.; Ren, J.-H. Interleukin-35 stimulates hepatitis B virus transcription and replication by targeting transcription factor HNF4α. J. Gen. Virol. 2018, 99, 645–654. [Google Scholar] [CrossRef]
- Sakaguchi, S.; Yamaguchi, T.; Nomura, T.; Ono, M. Regulatory T cells and immune tolerance. Cell 2008, 133, 775–787. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Jin, W.; Hardegen, N.; Lei, K.-J.; Li, L.; Marinos, N.; McGrady, G.; Wahl, S.M. Conversion of peripheral CD4+CD25- naive T cells to CD4+CD25+ regulatory T cells by TGF-beta induction of transcription factor Foxp3. J. Exp. Med. 2003, 198, 1875–1886. [Google Scholar] [CrossRef] [PubMed]
- Asseman, C.; Powrie, F. Interleukin 10 is a growth factor for a population of regulatory T cells. Gut 1998, 42, 157–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groux, H.; O’Garra, A.; Bigler, M.; Rouleau, M.; Antonenko, S.; De Vries, J.E.; Roncarolo, M.G. A CD4+ T-cell subset inhibits antigen-specific T-cell responses and prevents colitis. Nature 1997, 389, 737–742. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.G.; Wang, J.H.; Gray, J.D.; Soucier, H.; Horwitz, D.A. Natural and induced CD4+CD25+ cells educate CD4+CD25- cells to develop suppressive activity: The role of IL-2, TGF-beta, and IL-10. J. Immunol. 2004, 172, 5213–5221. [Google Scholar] [CrossRef] [Green Version]
- Shamji, M.H.; Layhadi, J.A.; Achkova, D.; Kouser, L.; Perera-Webb, A.; Couto-Francisco, N.C.; Parkin, R.V.; Matsuoka, T.; Scadding, G.; Ashton-Rickardt, P.G.; et al. Role of IL-35 in sublingual allergen immunotherapy. J. Allergy Clin. Immunol. 2019, 143, 1131–1142. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Zhang, H.; Huang, J.; Zhang, Q. Interleukin-35 in autoimmune dermatoses: Current concepts. Open Med. Wars Pol. 2022, 17, 589–600. [Google Scholar] [CrossRef]
- Chekol Abebe, E.; Asmamaw Dejenie, T.; Mengie Ayele, T.; Dagnew Baye, N.; Agegnehu Teshome, A.; Tilahun Muche, Z. The Role of Regulatory B Cells in Health and Diseases: A Systemic Review. J. Inflamm. Res. 2021, 14, 75–84. [Google Scholar] [CrossRef]
- Catalán, D.; Mansilla, M.A.; Ferrier, A.; Soto, L.; Oleinika, K.; Aguillón, J.C.; Aravena, O. Immunosuppressive Mechanisms of Regulatory B Cells. Front. Immunol. 2021, 12, 611795. [Google Scholar] [CrossRef]
- Wang, K.; Liu, J.; Li, J. IL-35-producing B cells in gastric cancer patients. Medicine 2018, 97, e0710. [Google Scholar] [CrossRef]
- van de Veen, W. The role of regulatory B cells in allergen immunotherapy. Curr. Opin. Allergy Clin. Immunol. 2017, 17, 447–452. [Google Scholar] [CrossRef] [Green Version]
- Czarnowicki, T.; He, H.; Krueger, J.G.; Guttman-Yassky, E. Atopic dermatitis endotypes and implications for targeted therapeutics. J. Allergy Clin. Immunol. 2019, 143, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Leung, D.Y.M.; Boguniewicz, M.; Howell, M.D.; Nomura, I.; Hamid, Q.A. New insights into atopic dermatitis. J. Clin. Investig. 2004, 113, 651–657. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Akahoshi, M.; Matsuda, A.; Ebe, K.; Inomata, N.; Obara, K.; Hirota, T.; Nakashima, K.; Shimizu, M.; Tamari, M.; et al. Association of the IL12RB1 promoter polymorphisms with increased risk of atopic dermatitis and other allergic phenotypes. Hum. Mol. Genet. 2005, 14, 3149–3159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Namkung, J.-H.; Lee, J.-E.; Kim, E.; Kim, S.; Kim, S.; Shin, E.-S.; Cho, E.-Y.; Yang, J.-M. Association of single nucleotide polymorphisms in the IL-12 (IL-12A and B) and IL-12 receptor (IL-12Rbeta1 and beta2) genes and gene-gene interactions with atopic dermatitis in Koreans. J. Dermatol. Sci. 2010, 57, 199–206. [Google Scholar] [CrossRef]
- Kim, J.; Kim, B.E.; Leung, D.Y.M. Pathophysiology of atopic dermatitis: Clinical implications. Allergy Asthma Proc. 2019, 40, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Salimi, M.; Barlow, J.L.; Saunders, S.P.; Xue, L.; Gutowska-Owsiak, D.; Wang, X.; Huang, L.-C.; Johnson, D.; Scanlon, S.T.; McKenzie, A.N.J.; et al. A role for IL-25 and IL-33-driven type-2 innate lymphoid cells in atopic dermatitis. J. Exp. Med. 2013, 210, 2939–2950. [Google Scholar] [CrossRef]
- Nakajima, S.; Kabata, H.; Kabashima, K.; Asano, K. Anti-TSLP antibodies: Targeting a master regulator of type 2 immune responses. Allergol. Int. 2020, 69, 197–203. [Google Scholar] [CrossRef]
- Yang, G.; Oh, J.W.; Lee, H.E.; Lee, B.H.; Lim, K.M.; Lee, J.Y. Topical Application of Dieckol Ameliorates Atopic Dermatitis in NC/Nga Mice by Suppressing Thymic Stromal Lymphopoietin Production. J. Investig. Dermatol. 2016, 136, 1062–1066. [Google Scholar] [CrossRef]
- Lee, H.E.; Yang, G.; Kim, K.B.; Lee, B.M.; Lee, J.Y. Phloxine O, a Cosmetic Colorant, Suppresses the Expression of Thymic Stromal Lymphopoietin and Acute Dermatitis Symptoms in Mice. Biomol. Ther. 2018, 26, 481–486. [Google Scholar] [CrossRef]
- Adhikary, P.P.; Tan, Z.; Page, B.D.G.; Hedtrich, S. TSLP as druggable target—A silver-lining for atopic diseases? Pharmacol. Ther. 2021, 217, 107648. [Google Scholar] [CrossRef]
- Kim, B.S.; Siracusa, M.C.; Saenz, S.A.; Noti, M.; Monticelli, L.A.; Sonnenberg, G.F.; Hepworth, M.R.; Van Voorhees, A.S.; Comeau, M.R.; Artis, D. TSLP elicits IL-33-independent innate lymphoid cell responses to promote skin inflammation. Sci. Transl. Med. 2013, 5, 170ra16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebbo, M.; Crinier, A.; Vély, F.; Vivier, E. Innate lymphoid cells: Major players in inflammatory diseases. Nat. Rev. Immunol. 2017, 17, 665–678. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, S.; Kitoh, A.; Egawa, G.; Natsuaki, Y.; Nakamizo, S.; Moniaga, C.S.; Otsuka, A.; Honda, T.; Hanakawa, S.; Amano, W.; et al. IL-17A as an inducer for Th2 immune responses in murine atopic dermatitis models. J. Investig. Dermatol. 2014, 134, 2122–2130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kader, H.; Azeem, M.; Jwayed, S.; Al-Shehhi, A.; Tabassum, A.; Ayoub, M.; Hetta, H.; Waheed, Y.; Iratni, R.; Al-Dhaheri, A.; et al. Current Insights into Immunology and Novel Therapeutics of Atopic Dermatitis. Cells 2021, 10, 1392. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Ren, X.; Liu, C.; Wang, J.; Dong, J.; Xu, O. The regulatory effect of IL-35 on the balance of Treg/Th17 cells in allergic rhinitis patients. Lin Chuang Er Bi Yan Hou Tou Jing Wai Ke Za Zhi J. Clin. Otorhinolaryngol. Head Neck Surg. 2016, 30, 213–216. [Google Scholar]
- Wang, D.; Lei, L. Interleukin-35 regulates the balance of Th17 and Treg responses during the pathogenesis of connective tissue diseases. Int. J. Rheum. Dis. 2021, 24, 21–27. [Google Scholar] [CrossRef]
- Ma, L.; Xue, H.-B.; Guan, X.-H.; Shu, C.-M.; Wang, F.; Zhang, J.-H.; An, R.-Z. The Imbalance of Th17 cells and CD4(+) CD25(high) Foxp3(+) Treg cells in patients with atopic dermatitis. J. Eur. Acad. Dermatol. Venereol. 2014, 28, 1079–1086. [Google Scholar] [CrossRef]
- Verhagen, J.; Akdis, M.; Traidl-Hoffmann, C.; Schmid-Grendelmeier, P.; Hijnen, D.; Knol, E.; Behrendt, H.; Blaser, K.; Akdis, C.A. Absence of T-regulatory cell expression and function in atopic dermatitis skin. J. Allergy Clin. Immunol. 2006, 117, 176–183. [Google Scholar] [CrossRef]
- Szegedi, A.; Baráth, S.; Nagy, G.; Szodoray, P.; Gal, M.; Sipka, S.; Bagdi, E.; Banham, A.; Krenács, L. Regulatory T cells in atopic dermatitis: Epidermal dendritic cell clusters may contribute to their local expansion. Br. J. Dermatol. 2009, 160, 984–993. [Google Scholar] [CrossRef]
- Fujimura, T.; Okuyama, R.; Ito, Y.; Aiba, S. Profiles of Foxp3+ regulatory T cells in eczematous dermatitis, psoriasis vulgaris and mycosis fungoides. Br. J. Dermatol. 2008, 158, 1256–1263. [Google Scholar] [CrossRef]
- Roesner, L.M.; Floess, S.; Witte, T.; Olek, S.; Huehn, J.; Werfel, T. Foxp3(+) regulatory T cells are expanded in severe atopic dermatitis patients. Allergy 2015, 70, 1656–1660. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y.; Adachi, Y.; Makino, T.; Higashiyama, H.; Fuchizawa, T.; Shimizu, T.; Miyawaki, T. Expansion of FOXP3-positive CD4+CD25+ T cells associated with disease activity in atopic dermatitis. Ann. Allergy Asthma Immunol. 2009, 103, 160–165. [Google Scholar] [CrossRef] [PubMed]
- García, E.M.; Galicia-Carreón, J.; Novak, N. In vitro Conversion into CD4+CD25+Foxp3+ Induced Regulatory T Cells Is Reduced in Atopic Dermatitis Patients. Int. Arch. Allergy Immunol. 2020, 181, 353–356. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.J.; Hao, F.; Qian, T.; Cheng, H.X. Expression of Helper and Regulatory T Cells in Atopic Dermatitis: A Meta-Analysis. Front. Pediatr. 2022, 10, 777992. [Google Scholar] [CrossRef] [PubMed]
- Matsui, E.; Kaneko, H.; Fukao, T.; Teramoto, T.; Inoue, R.; Watanabe, M.; Kasahara, K.; Kondo, N. Mutations of the IL-12 receptor beta2 chain gene in atopic subjects. Biochem. Biophys. Res. Commun. 1999, 266, 551–555. [Google Scholar] [CrossRef]
- CChen, X.-F.; Zhang, L.-J.; Zhang, J.; Dou, X.; Shao, Y.; Jia, X.-J.; Zhang, W.; Yu, B. MiR-151a is involved in the pathogenesis of atopic dermatitis by regulating interleukin-12 receptor β2. Exp. Dermatol. 2018, 27, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Shen, C.; Liu, Y.; Li, Y.; Sun, L.; Jiao, L.; Jiao, W.; Xiao, J.; Shen, C.; Qi, H.; et al. Impaired Function of CD5+CD19+CD1dhi B10 Cells on IgE Secretion in an Atopic Dermatitis-Like Mouse Model. PLoS ONE 2015, 10, e0132173. [Google Scholar] [CrossRef]
- Yoshihara, Y.; Ishiuji, Y.; Yoshizaki, A.; Kurita, M.; Hayashi, M.; Ishiji, T.; Nakagawa, H.; Asahina, A.; Yanaba, K. IL-10-Producing Regulatory B Cells Are Decreased in Patients with Atopic Dermatitis. J. Investig. Dermatol. 2019, 139, 475–478. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Mao, Y.; Zhang, J.; Shi, G.; Cheng, L.; Lin, Y.; Li, Y.; Zhang, X.; Zhang, Y.; Chen, X.; et al. IL-35 recombinant protein reverses inflammatory bowel disease and psoriasis through regulation of inflammatory cytokines and immune cells. J. Cell. Mol. Med. 2018, 22, 1014–1025. [Google Scholar] [CrossRef]
- Cai, Z.; Wong, C.K.; Dong, J.; Chu, M.; Jiao, D.; Kam, N.W.; Lam, C.W.K.; Tam, L.S. Remission of systemic lupus erythematosus disease activity with regulatory cytokine interleukin (IL)-35 in Murphy Roths Large (MRL)/lpr mice. Clin. Exp. Immunol. 2015, 181, 253–266. [Google Scholar] [CrossRef] [Green Version]
- Nakashima, C.; Yanagihara, S.; Otsuka, A. Innovation in the treatment of atopic dermatitis: Emerging topical and oral Janus kinase inhibitors. Allergol. Int. 2022, 71, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Puar, N.; Chovatiya, R.; Paller, A.S. New treatments in atopic dermatitis. Ann. Allergy Asthma Immunol. 2021, 126, 21–31. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Country and Authors | Patients and Condition | IL-35 Level Measurement Method | Level in Blood | Additional Information |
|---|---|---|---|---|
| Saudi Arabia 2022; Abushouk et al. [22] | Children with Asthma vs. Healthy individuals | ELISA; serum | Increased; Asthma patients: 28.06 ± 8.39 pg/mL; Healthy control: 5.38 ± 5.54 pg/mL | IL-35 levels negatively correlated with levels of IgE. |
| China 2015; Wong et al. [23] | Adults and children with Asthma vs. Healthy individuals | ELISA; plasma | Increased; Asthma patients: 55.9 (6.6–419.0) ng/mL; Healthy control: 2.5 (0.1–16.1) ng/mL | IL-35 levels positively correlated with disease severity scores. |
| China 2015; Wang et al. [24] | Adults with Asthma vs. Healthy individuals | ELISA; plasma; qPCR; IL-35 mRNA expression levels in PBMCs | Decreased levels of IL-35 in plasma *. Decreased mRNA levels of the IL-35 subunits * | IL-35 levels negatively correlated with the frequency of IL-4-producing CD8+ T (Tc2) cells and with the IL-4 level. |
| Iran 2017; Khoshkhui et al. [25] | Children with Asthma vs. Healthy individuals | ELISA; serum | No significant difference; Asthma patients: 30.9 (3.8–110.7) pg/mL; Healthy control: 30.2 (6.1–239.7) pg/mL | |
| China 2014; Ma et al. [26] | Children with Asthma vs. Healthy individuals | ELISA; serum; qPCR; IL-35 mRNA expression levels in PBMCs | Decreased levels of IL-35 in serum *. Decreased mRNA levels of the IL-35 * | IL-35 levels negatively correlated with IL-4 levels and positively with IFN-y levels. |
| China 2020; Wang et al. [27] | Adults: Allergic asthmatic patients vs. Asymptomatic sensitized patients vs. Healthy individuals | Flow cytometry; the iTr35 cell frequency in PBMCs | Decreased iTr35 cell frequencies and IL-35 levels in allergic asthmatic patients * | 1. sIgE levels negatively correlated with the percentage of iTr35 cells in asthmatic individuals; 2. Th2 cytokines levels negatively correlated with the iTr35 cell frequency in asthmatic, asymptomatic, and healthy individuals; 3. Th2 cytokines and sIgE levels negatively correlated with IL-35 levels in asthmatic patients. |
| China 2020; Xie et al. [28] | Children with Allergic rhinitis vs. Healthy individuals | ELISA; serum | Decreased * | IL-35 levels negatively correlated with IL-17 and IL-23 levels. |
| China 2021; Huang et al. [29] | Children with Allergic rhinitis vs. Healthy individuals | ELISA; plasma | Decreased; Allergic rhinitis patients: 138.52 ± 50.13 ng/mL; Healthy control: 426.45 ± 80.15 ng/mL | IL-35 levels negatively correlated with ILC2s. |
| China 2018; Chen et al. [30] | CSU patients vs. AD patients vs. Healthy individuals | ELISA; serum | Decreased in CSU patients: 73.46 ± 9.146 ng/mL; AD: 1264 ± 186.9 ng/mL; Healthy control: 1349 ± 170.7 ng/mL | |
| China 2014; Li et al. [31] | Patients with IBD: ulcerative colitis (UC) Crohn’s disease (CD) vs. Healthy individuals | ELISA; serum | Decreased in UC and CD *. | IL-35 levels negatively correlated with UC activity. |
| China 2018; Li et al. [32] | Adults with Psoriasis vs. Healthy individuals | ELISA; plasma; qPCR; PBMCs; Ebi3 and p35 mRNA levels | Decreased IL-35 levels: 2.67 (1.38–21.81) ng/mL vs. 7.92 (2.88–41.07) ng/mL; Decreased mRNA levels of EBI3 and p35 * | IL-35 levels negatively correlated with IFNy, TNF-a, levels of IL-23, -17, and -22, or the PASI and positively with TGF-β and IL-10 levels. |
| Egypt 2022; Elbana et al. [33] | Adults with Psoriasis vs. Healthy individuals | ELISA; serum | Decreased; Psoriasis patients: 72.65 ± 16.24 ng/L; Healthy control: 451.02 ± 117.16 ng/L | IL-35 levels negatively correlated with TNF-α, IL-17, IFN-γ. |
| China 2019; Ye et al. [34] | Adults with newly diagnosed SLE vs. Healthy individuals | ELISA; plasma | Decreased * | IL-35+B cells and IL-10+B cells decreased; The percentage of IL-35+Bregs and IL-35 levels inversely correlated with the SLE disease activity index. |
| China 2013; Qiu et al. [20] | Patients with newly diagnosed SLE; Pre-treatment SLE vs. post-treatment SLE vs. Healthy individuals | ELISA; serum | Increased; Pre-treatment: 9.94 (4.28–63.83) ng/L; Post-treatment: 5.78 (4.08–54.95) ng/L; Healthy control: 4.74 (3.38–12.45) ng/L | After prednisone treatment, the serum levels of IL-35 decreased significantly. |
| China 2019; Li et al. [35] | Adults with RA vs. Healthy individuals | ELISA; serum | Increased; RA patients: 6.3 (4.8–10.0) pg/mL; Healthy control: 1.3 (0.7–2.5) pg/mL | IL-35 levels negatively correlated with diseases activity based on ESR (DAS28-ESR). |
| Czech Republic 2015; Šenolt et al. [21] | Adults with treatment-naïve early RA vs. established RA vs. Control patients with osteoarthritis | ELISA; Serum and synovial fluid | Serum: Increased in patients with treatment-naïve early RA at baseline: 81.6 (20.7–564.4) pg/mL; Control patients: 10.4 (0.6–64.1); Established RA 22.8 (1.2–145.5) pg/mL; Synovial fluid: Increased Established RA: 445.0 (40.7–1908.0) pg/mL; Control patients: 125.5 (39.1–1062.0) pg/mL | IL-35 levels significantly decreased after treatment initiation to 36.5 (5.0–204.8) pg/mL; Synovial fluid IL-35 levels positively correlated with disease activity assessed by CRP and DAS28. |
| Egypt 2022; Kiwan et al. [36] | Patients with AD vs. Healthy individuals | ELISA; serum | Decreased; AD patients: 69.7 ± 14.8 ng/L; Healthy control: 415.96 ± 99.25 ng/L | IL-35 levels negatively correlated with diseases severity assessed by SCORAD, TNF- α, IL-17 and positively correlated with TGF-β. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zysk, W.; Gleń, J.; Trzeciak, M. Current Insight into the Role of IL-35 and Its Potential Involvement in the Pathogenesis and Therapy of Atopic Dermatitis. Int. J. Mol. Sci. 2022, 23, 15709. https://doi.org/10.3390/ijms232415709
Zysk W, Gleń J, Trzeciak M. Current Insight into the Role of IL-35 and Its Potential Involvement in the Pathogenesis and Therapy of Atopic Dermatitis. International Journal of Molecular Sciences. 2022; 23(24):15709. https://doi.org/10.3390/ijms232415709
Chicago/Turabian StyleZysk, Weronika, Jolanta Gleń, and Magdalena Trzeciak. 2022. "Current Insight into the Role of IL-35 and Its Potential Involvement in the Pathogenesis and Therapy of Atopic Dermatitis" International Journal of Molecular Sciences 23, no. 24: 15709. https://doi.org/10.3390/ijms232415709