
A Thiosemicarbazone Derivative as a Booster in Photodynamic Therapy—A Way to Improve the Therapeutic Effect

Abstract
:1. Introduction
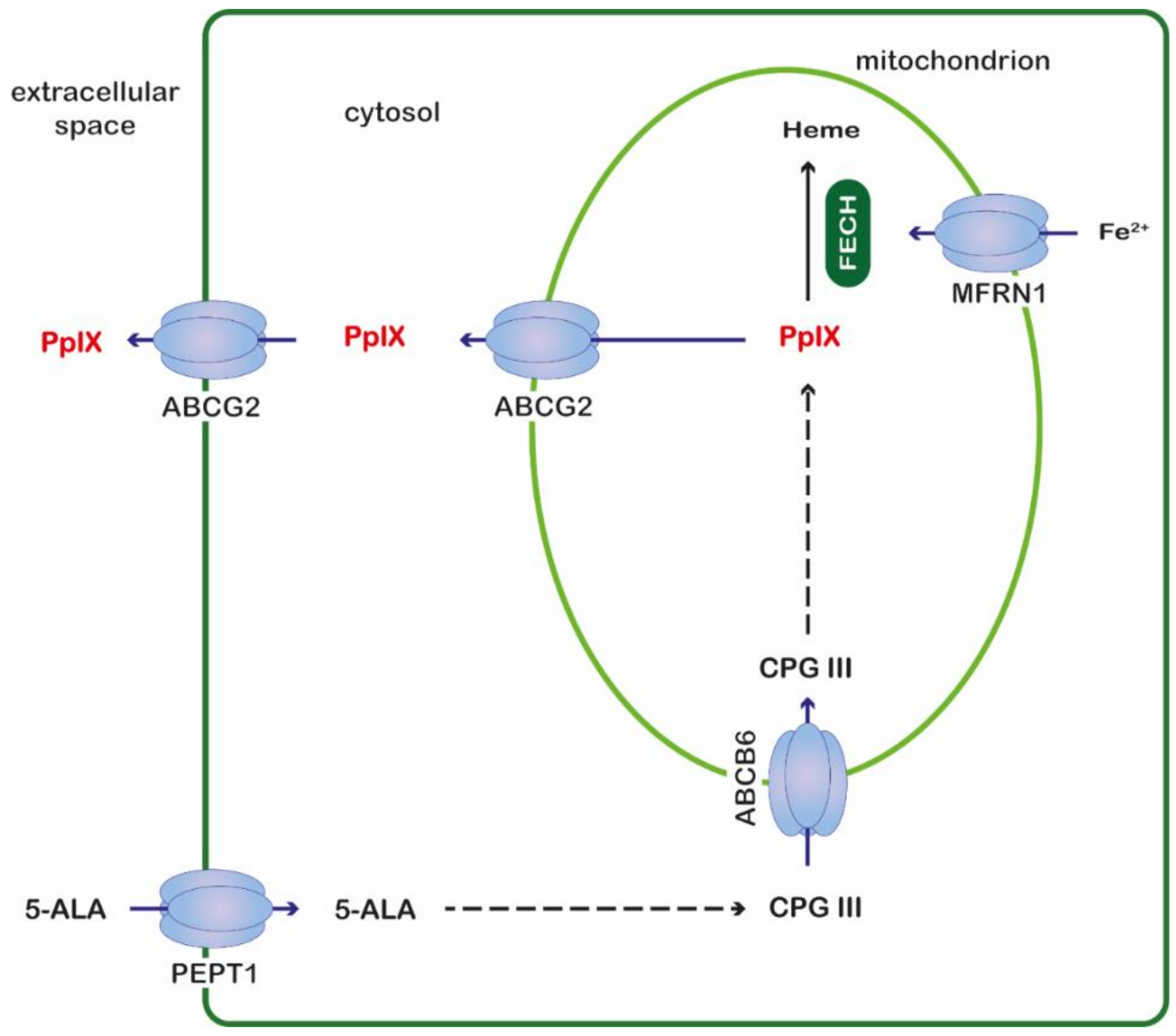
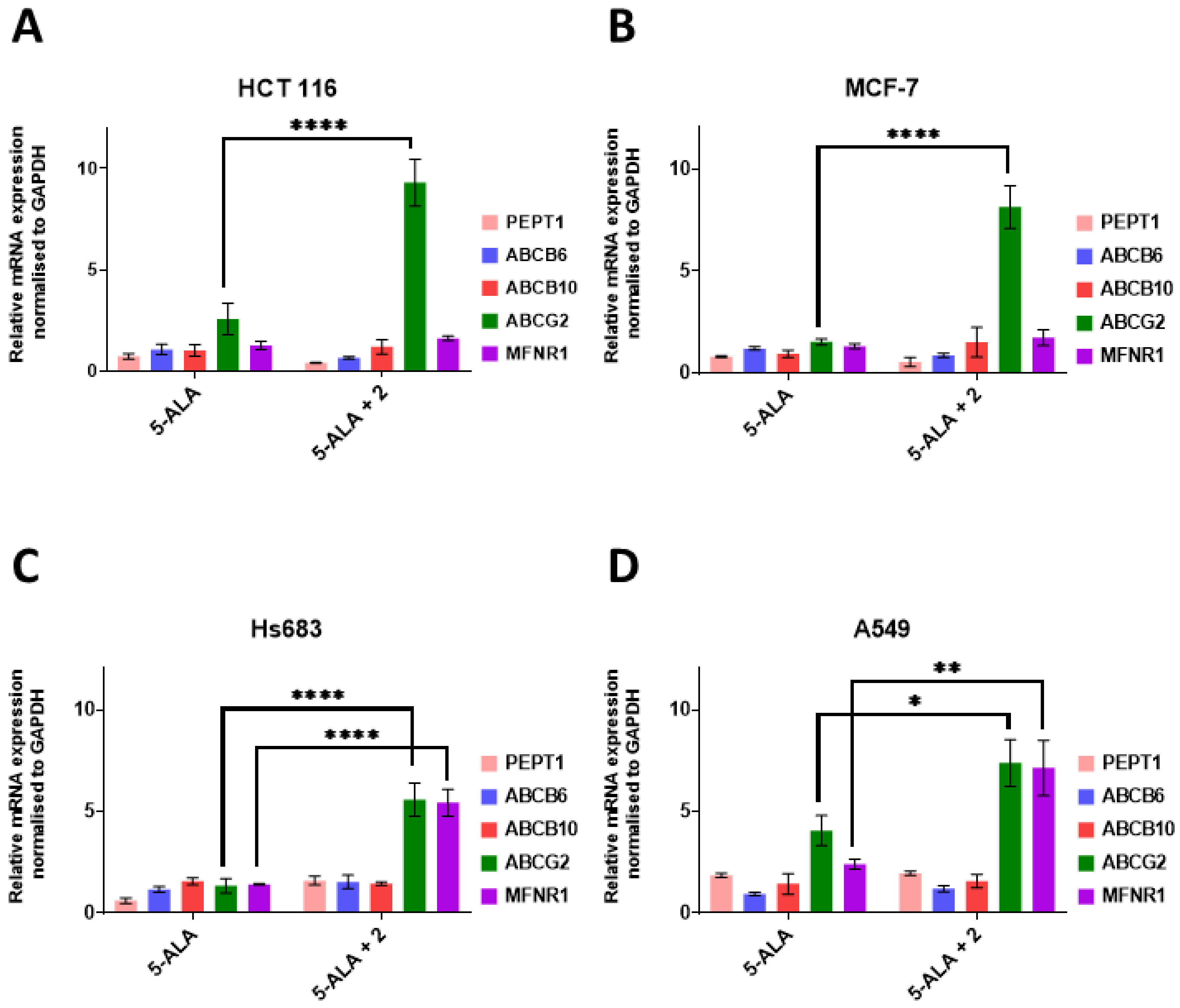
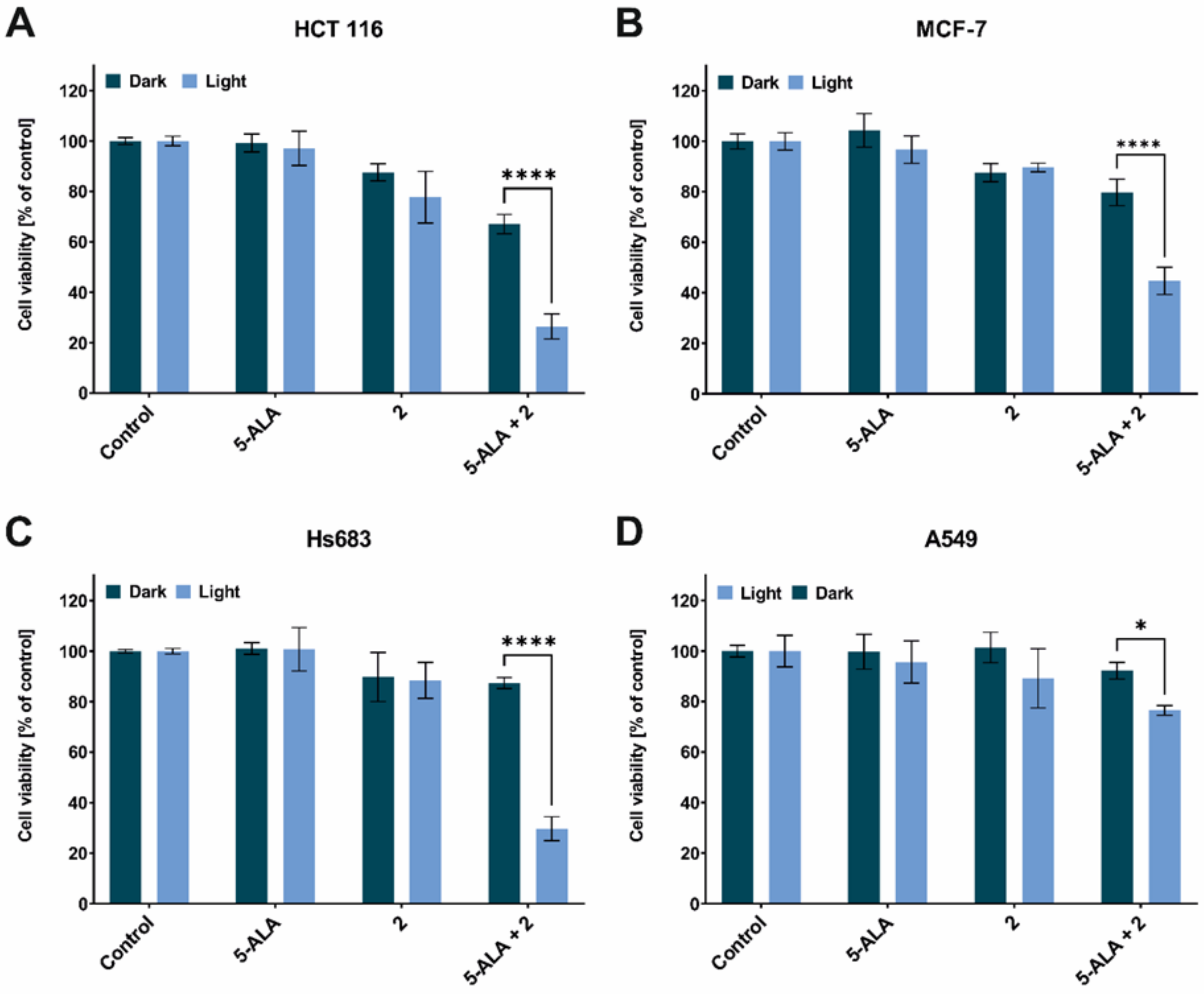
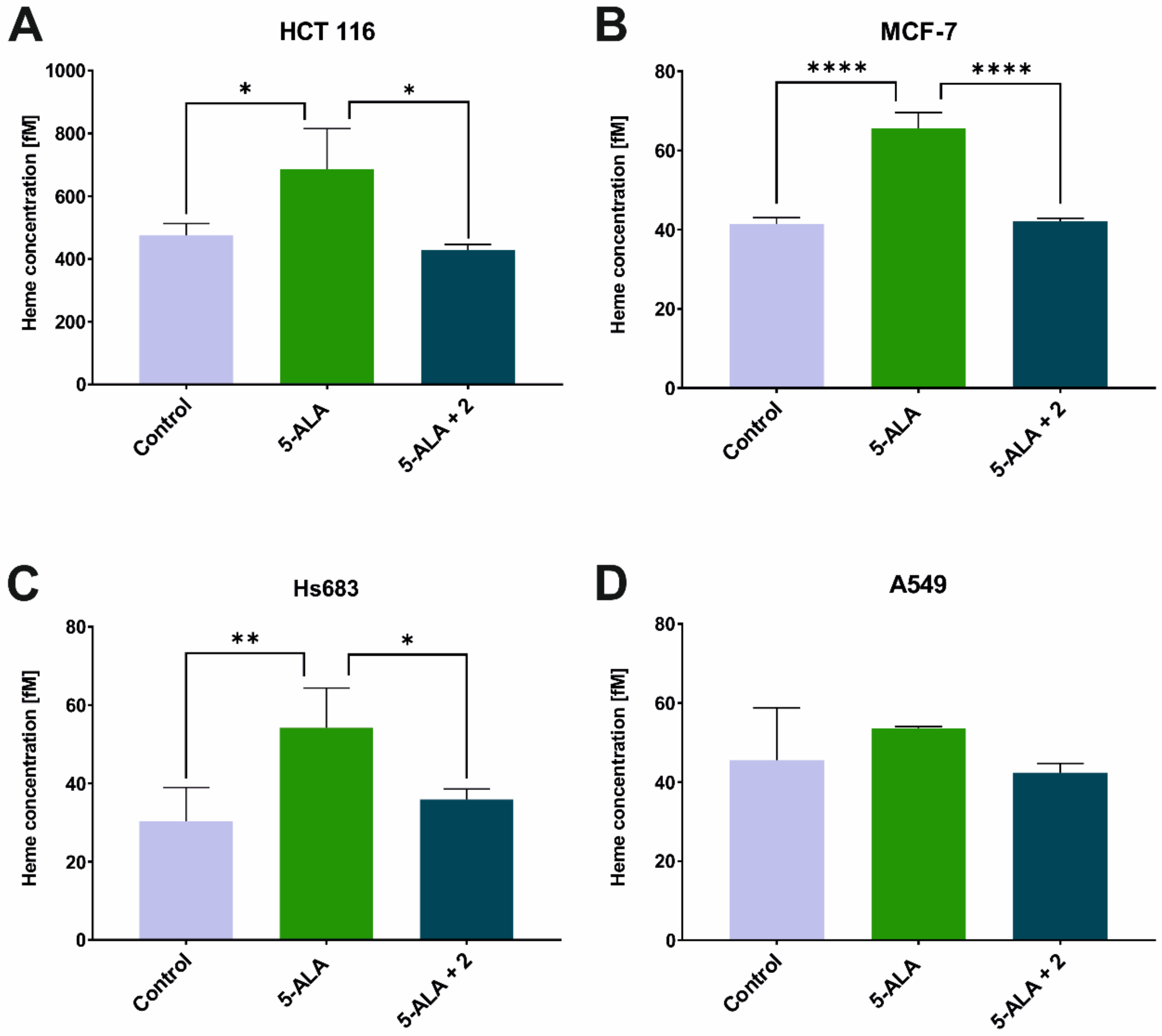
2. Results and Discussion
3. Materials and Methods
3.1. Cell Lines and Cell Culture
3.2. Transporter Genes Expression
3.3. Heme Concentration Measurement
3.4. Phototoxic Effect
3.5. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gunaydin, G.; Gedik, M.E.; Ayan, S. Photodynamic Therapy—Current Limitations and Novel Approaches. Front. Chem. 2021, 9, 691697. [Google Scholar] [CrossRef] [PubMed]
- Benov, L. Photodynamic Therapy: Current Status and Future Directions. Med. Princ. Pract. 2015, 24, 14–28. [Google Scholar] [CrossRef] [PubMed]
- Jori, G. Tumour photosensitizers: Approaches to enhance the selectivity and efficiency of photodynamic therapy. J. Photochem. Photobiol. B Biol. 1996, 36, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Castano, A.P.; Demidova, T.N.; Hamblin, M.R. Mechanisms in photodynamic therapy: Part three—Photosensitizer pharmacokinetics, biodistribution, tumor localization and modes of tumor destruction. Photodiagnosis Photodyn. Ther. 2005, 2, 91–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, N.; Squire, J.; Guirguis, M.; Saha, D.; Hoyt, K.; Wang, K.K.-H.; Agarwal, V.; Obaid, G. Deep-Tissue Activation of Photonanomedicines: An Update and Clinical Perspectives. Cancers 2022, 14, 2004. [Google Scholar] [CrossRef]
- Beiki, D.; Eggleston, I.M.; Pourzand, C. Daylight-PDT: Everything under the sun. Biochem. Soc. Trans. 2022, 50, 975–985. [Google Scholar] [CrossRef]
- Curnow, A.; Perry, A.; Wood, M. Improving in vitro photodynamic therapy through the development of a novel iron chelating aminolaevulinic acid prodrug. Photodiagnosis Photodyn. Ther. 2018, 25, 157–165. [Google Scholar] [CrossRef]
- Gawecki, R.; Malarz, K.; Rejmund, M.; Polanski, J.; Mrozek-Wilczkiewicz, A. Impact of thiosemicarbazones on the accumulation of PpIX and the expression of the associated genes. J. Photochem. Photobiol. B Biol. 2019, 199, 111585. [Google Scholar] [CrossRef]
- Anderson, C.M.H.; Jevons, M.; Thangaraju, M.; Edwards, N.; Conlon, N.J.; Woods, S.; Ganapathy, V.; Thwaites, D.T. Transport of the Photodynamic Therapy Agent 5-Aminolevulinic Acid by Distinct H+-Coupled Nutrient Carriers Coexpressed in the Small Intestine. J. Pharmacol. Exp. Ther. 2010, 332, 220–228. [Google Scholar] [CrossRef] [Green Version]
- Döring, F.; Walter, J.; Will, J.; Föcking, M.; Boll, M.; Amasheh, S.; Clauss, W.; Daniel, H. Delta-aminolevulinic acid transport by intestinal and renal peptide transporters and its physiological and clinical implications. J. Clin. Investig. 1998, 101, 2761–2767. [Google Scholar] [CrossRef]
- Kottra, G.; Daniel, H. The proton oligopeptide cotransporter family SLC15 in physiology and pharmacology. Pflugers. Arch. 2004, 447, 610–618. [Google Scholar] [CrossRef]
- Hagiya, Y.; Fukuhara, H.; Matsumoto, K.; Endo, Y.; Nakajima, M.; Tanaka, T.; Okura, I.; Kurabayashi, A.; Furihata, M.; Inoue, K.; et al. Expression levels of PEPT1 and ABCG2 play key roles in 5-aminolevulinic acid (ALA)-induced tumor-specific protoporphyrin IX (PpIX) accumulation in bladder cancer. Photodiagnosis Photodyn. Ther. 2013, 10, 288–295. [Google Scholar] [CrossRef]
- Nakayama, T.; Nozawa, N.; Kawada, C.; Yamamoto, S.; Ishii, T.; Ishizuka, M.; Namikawa, T.; Ogura, S.-I.; Hanazaki, K.; Inoue, K.; et al. Mitomycin C-induced cell cycle arrest enhances 5-aminolevulinic acid-based photodynamic therapy for bladder cancer. Photodiagnosis Photodyn. Ther. 2020, 31, 101893. [Google Scholar] [CrossRef]
- Nakayama, T.; Otsuka, S.; Kobayashi, T.; Okajima, H.; Matsumoto, K.; Hagiya, Y.; Inoue, K.; Shuin, T.; Nakajima, M.; Tanaka, T.; et al. Dormant cancer cells accumulate high protoporphyrin IX levels and are sensitive to 5-aminolevulinic acid-based photodynamic therapy. Sci. Rep. 2016, 6, 36478. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, K.; Hagiya, Y.; Endo, Y.; Nakajima, M.; Ishizuka, M.; Tanaka, T.; Ogura, S.-I. Effects of plasma membrane ABCB6 on 5-aminolevulinic acid (ALA)-induced porphyrin accumulation in vitro: Tumor cell response to hypoxia. Photodiagnosis Photodyn. Ther. 2015, 12, 45–51. [Google Scholar] [CrossRef]
- Bayeva, M.; Khechaduri, A.; Wu, R.; Burke, M.A.; Wasserstrom, J.A.; Singh, N.; Liesa, M.; Shirihai, O.S.; Langer, N.B.; Paw, B.H.; et al. ATP-Binding Cassette B10 Regulates Early Steps of Heme Synthesis. Circ. Res. 2013, 113, 279–287. [Google Scholar] [CrossRef] [Green Version]
- Seguin, A.; Takahashi-Makise, N.; Yien, Y.Y.; Huston, N.C.; Whitman, J.C.; Musso, G.; Wallace, J.A.; Bradley, T.; Bergonia, H.A.; Kafina, M.D.; et al. Reductions in the mitochondrial ABC transporter Abcb10 affect the transcriptional profile of heme biosynthesis genes. J. Biol. Chem. 2017, 292, 16284–16299. [Google Scholar] [CrossRef] [Green Version]
- Hagiya, Y.; Endo, Y.; Yonemura, Y.; Takahashi, K.; Ishizuka, M.; Abe, F.; Tanaka, T.; Okura, I.; Nakajima, M.; Ishikawa, T.; et al. Pivotal roles of peptide transporter PEPT1 and ATP-binding cassette (ABC) transporter ABCG2 in 5-aminolevulinic acid (ALA)-based photocytotoxicity of gastric cancer cells in vitro. Photodiagnosis Photodyn. Ther. 2012, 9, 204–214. [Google Scholar] [CrossRef]
- Kim, J.H.; Park, J.M.; Roh, Y.J.; Kim, I.-W.; Hasan, T.; Choi, M.-G. Enhanced efficacy of photodynamic therapy by inhibiting ABCG2 in colon cancers. BMC Cancer 2015, 15, 504. [Google Scholar] [CrossRef] [Green Version]
- Paradkar, P.N.; Zumbrennen, K.B.; Paw, B.H.; Ward, D.M.; Kaplan, J. Regulation of Mitochondrial Iron Import through Differential Turnover of Mitoferrin 1 and Mitoferrin 2. Mol. Cell. Biol. 2009, 29, 1007–1016. [Google Scholar] [CrossRef]
- Rzycka-Korzec, R.; Malarz, K.; Gawecki, R.; Mrozek-Wilczkiewicz, A.; Małecki, J.G.; Schab-Balcerzak, E.; Korzec, M.; Polanski, J. Effect of the complex-formation ability of thiosemicarbazones containing (aza)benzene or 3-nitro-1,8-naphthalimide unit towards Cu(II) and Fe(III) ions on their anticancer activity. J. Photochem. Photobiol. A Chem. 2021, 415, 113314. [Google Scholar] [CrossRef]
- Teshigawara, T.; Mizuno, M.; Ishii, T.; Kitajima, Y.; Utsumi, F.; Sakata, J.; Kajiyama, H.; Shibata, K.; Ishizuka, M.; Kikkawa, F. Novel potential photodynamic therapy strategy using 5-Aminolevulinic acid for ovarian clear-cell carcinoma. Photodiagnosis Photodyn. Ther. 2017, 21, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Briel-Pump, A.; Beez, T.; Ebbert, L.; Remke, M.; Weinhold, S.; Sabel, M.C.; Sorg, R.V. Accumulation of protoporphyrin IX in medulloblastoma cell lines and sensitivity to subsequent photodynamic treatment. J. Photochem. Photobiol. B Biol. 2018, 189, 298–305. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.; Johnson, R.; Chung, C.-W.; Jeong, Y.-I.; Kang, D.H.; Suh, H. Poly(L-histidine)-tagged 5-aminolevulinic acid prodrugs: New photosensitizing precursors of protoporphyrin IX for photodynamic colon cancer therapy. Int. J. Nanomed. 2012, 7, 2497–2512. [Google Scholar] [CrossRef] [Green Version]
- Bourré, L.; Giuntini, F.; Eggleston, I.M.; Wilson, M.; MacRobert, A.J. 5-Aminolaevulinic acid peptide prodrugs enhance photosensitization for photodynamic therapy. Mol. Cancer Ther. 2008, 7, 1720–1729. [Google Scholar] [CrossRef] [Green Version]
- Ishizuka, M.; Abe, F.; Sano, Y.; Takahashi, K.; Inoue, K.; Nakajima, M.; Kohda, T.; Komatsu, N.; Ogura, S.-I.; Tanaka, T. Novel development of 5-aminolevurinic acid (ALA) in cancer diagnoses and therapy. Int. Immunopharmacol. 2011, 11, 358–365. [Google Scholar] [CrossRef]
- Frank, J.; Lornejad-Schäfer, M.R.; Schöffl, H.; Flaccus, A.; Lambert, C.; Biesalski, H.K. Inhibition of heme oxygenase-1 increases responsiveness of melanoma cells to ALA-based photodynamic therapy. Int. J. Oncol. 2007, 31, 1539–1545. [Google Scholar] [CrossRef] [Green Version]
- Nowis, D.; Legat, M.; Grzela, T.; Niderla, J.; Wilczek, E.; Wilczynski, G.M.; Głodkowska, E.; Mrówka, P.; Issat, T.; Dulak, J.; et al. Heme oxygenase-1 protects tumor cells against photodynamic therapy-mediated cytotoxicity. Oncogene 2006, 25, 3365–3374. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell line | Increase in fluorescence of PpIX relative to 5-ALA after treatment with TSC derivative 2 in combination with 5-ALA. | Expression of mRNA after treatment with TSC derivative 2 in combination with 5-ALA relative to 5-ALA. | |
| FECH | HO-1 | ||
| HCT 116 | 4.2-fold change | Down-regulated | Down-regulated |
| MCF-7 | 1.6-fold change | Non-significant changes | Down-regulated |
| Hs683 | 1.7-fold change | Up-regulated | Up-regulated |
| A549 | Non-significant changes | Non-significant changes | Non-significant changes |
| Gene Name | Sequence 5′–3′ (F-Forward, R-Reverse) | |
|---|---|---|
| PEPT1 | F | AGGCAACAACTATGTCCGGG |
| R | CACAGCATCGAAGATCGGGA | |
| ABCB6 | F | CTGCGGTATGTGGTCTCTGG |
| R | CCAGGTAGACTGTTGGGCTG | |
| ABCB10 | F | GTACGGGTCGCACGCA |
| R | GTGAACGGCGATAGGGACC | |
| ABCG2 | F | GCACAGGAAGTTTACGCACAG |
| R | AAGGGGCTAGAAGAAGGGGG | |
| MFRN1 | F | ACTCGGTGAAGACACGAATGC |
| R | CAGCTATCCCGTTGGCTAGG | |
| GAPDH | F | GAGTCAACGGATTTGGTCGTA |
| R | GCCCCACTTGATTTTGGAG | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gawecki, R.; Polanski, J.; Mrozek-Wilczkiewicz, A. A Thiosemicarbazone Derivative as a Booster in Photodynamic Therapy—A Way to Improve the Therapeutic Effect. Int. J. Mol. Sci. 2022, 23, 15370. https://doi.org/10.3390/ijms232315370
Gawecki R, Polanski J, Mrozek-Wilczkiewicz A. A Thiosemicarbazone Derivative as a Booster in Photodynamic Therapy—A Way to Improve the Therapeutic Effect. International Journal of Molecular Sciences. 2022; 23(23):15370. https://doi.org/10.3390/ijms232315370
Chicago/Turabian StyleGawecki, Robert, Jaroslaw Polanski, and Anna Mrozek-Wilczkiewicz. 2022. "A Thiosemicarbazone Derivative as a Booster in Photodynamic Therapy—A Way to Improve the Therapeutic Effect" International Journal of Molecular Sciences 23, no. 23: 15370. https://doi.org/10.3390/ijms232315370