
Biological Functions of Hydrogen Sulfide in Plants

Abstract
:1. Introduction
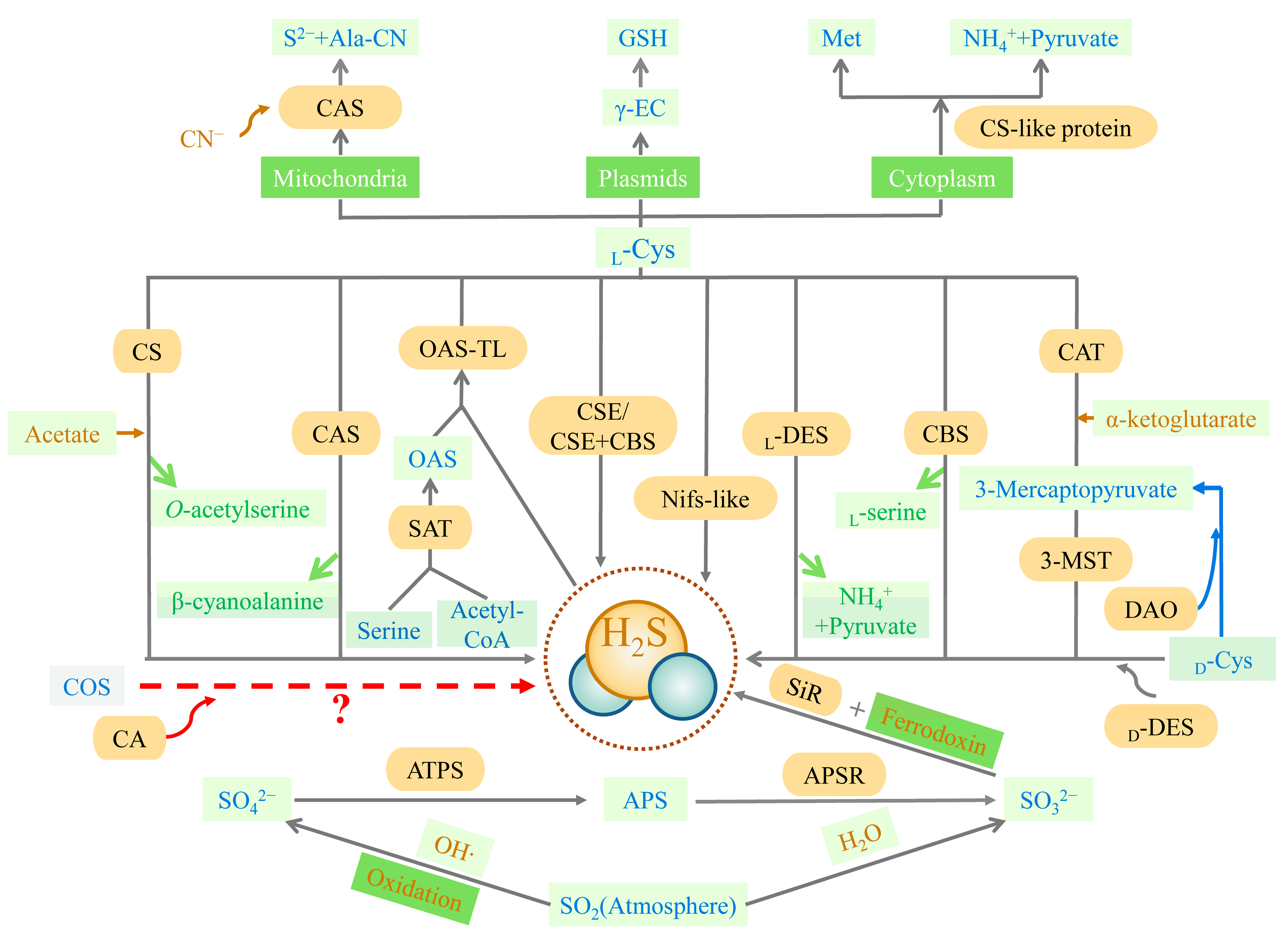
2. Biosynthesis of H2S in Organisms
2.1. Biosynthesis Pathway of H2S
2.1.1. Non-Enzymatic Pathway
2.1.2. Enzymatic Pathway
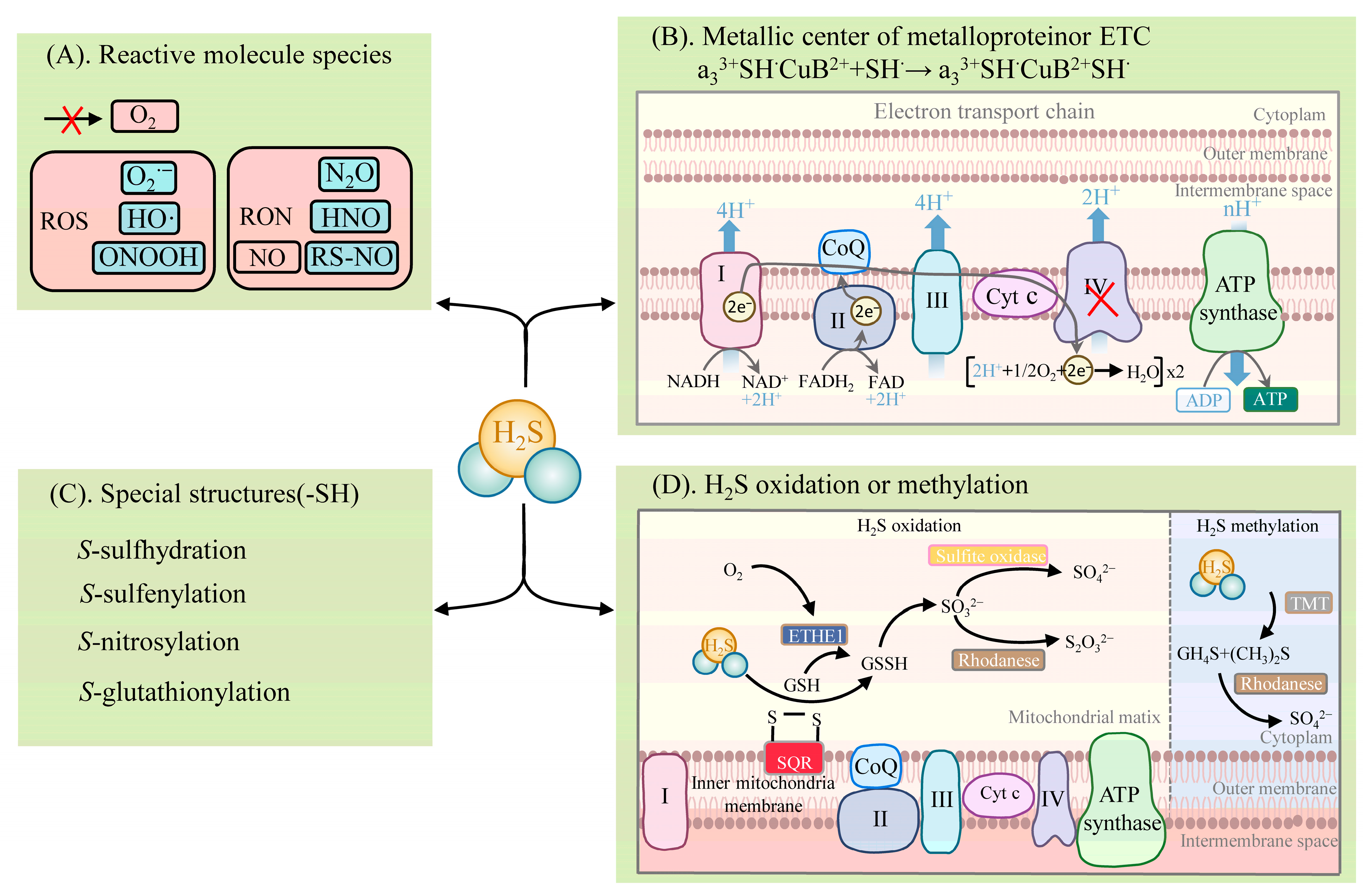
2.2. Metabolic Pathways of H2S
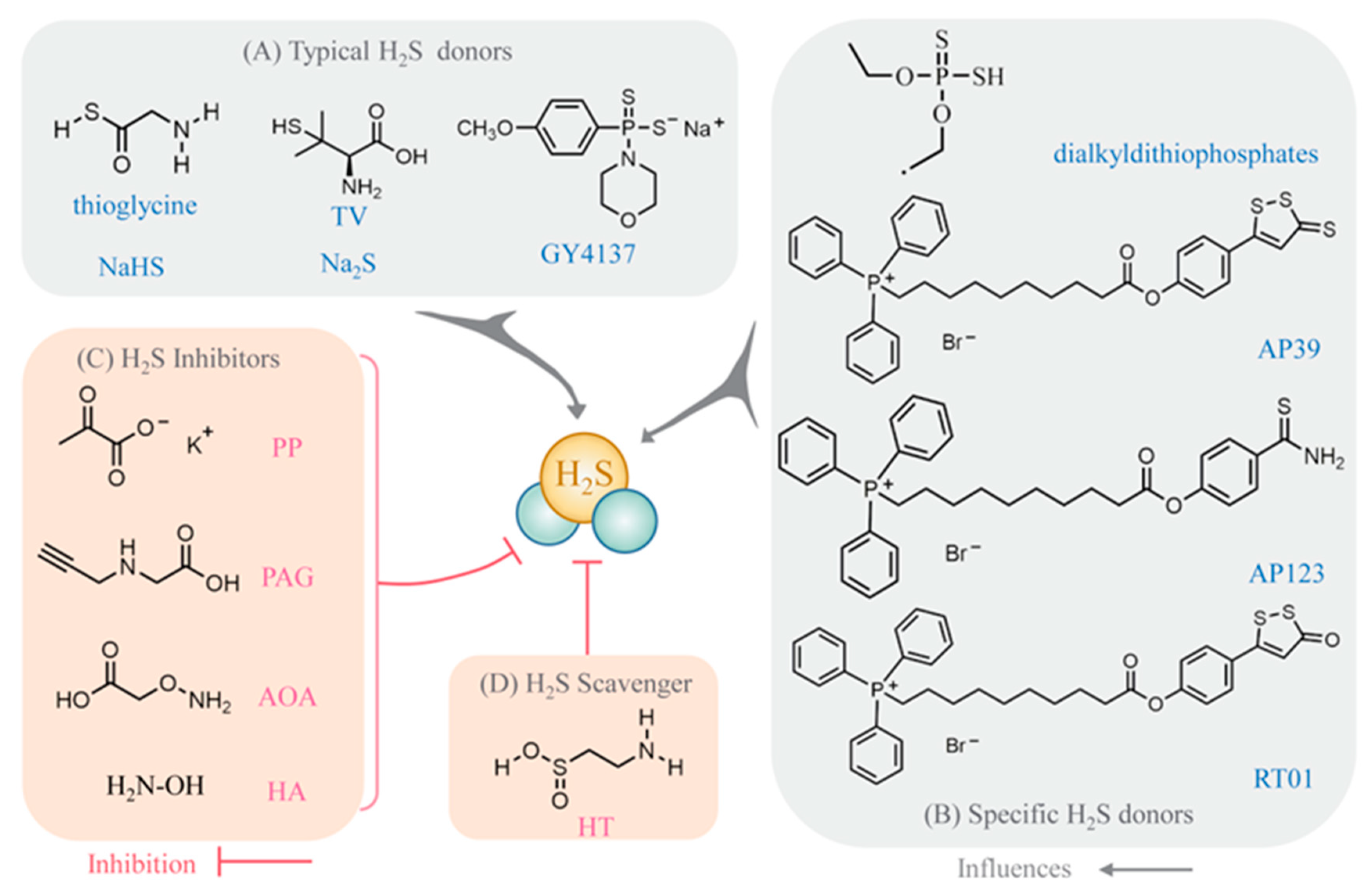
3. H2S Donors, Inhibitors (Including Scavengers), and Detection Methods
3.1. Selection of Pharmacological Agents for H2S Pharmacology Experiments
3.2. H2S Detection Methods
3.3. Detection of Protein S-Sulfhydration Modifications by H2S
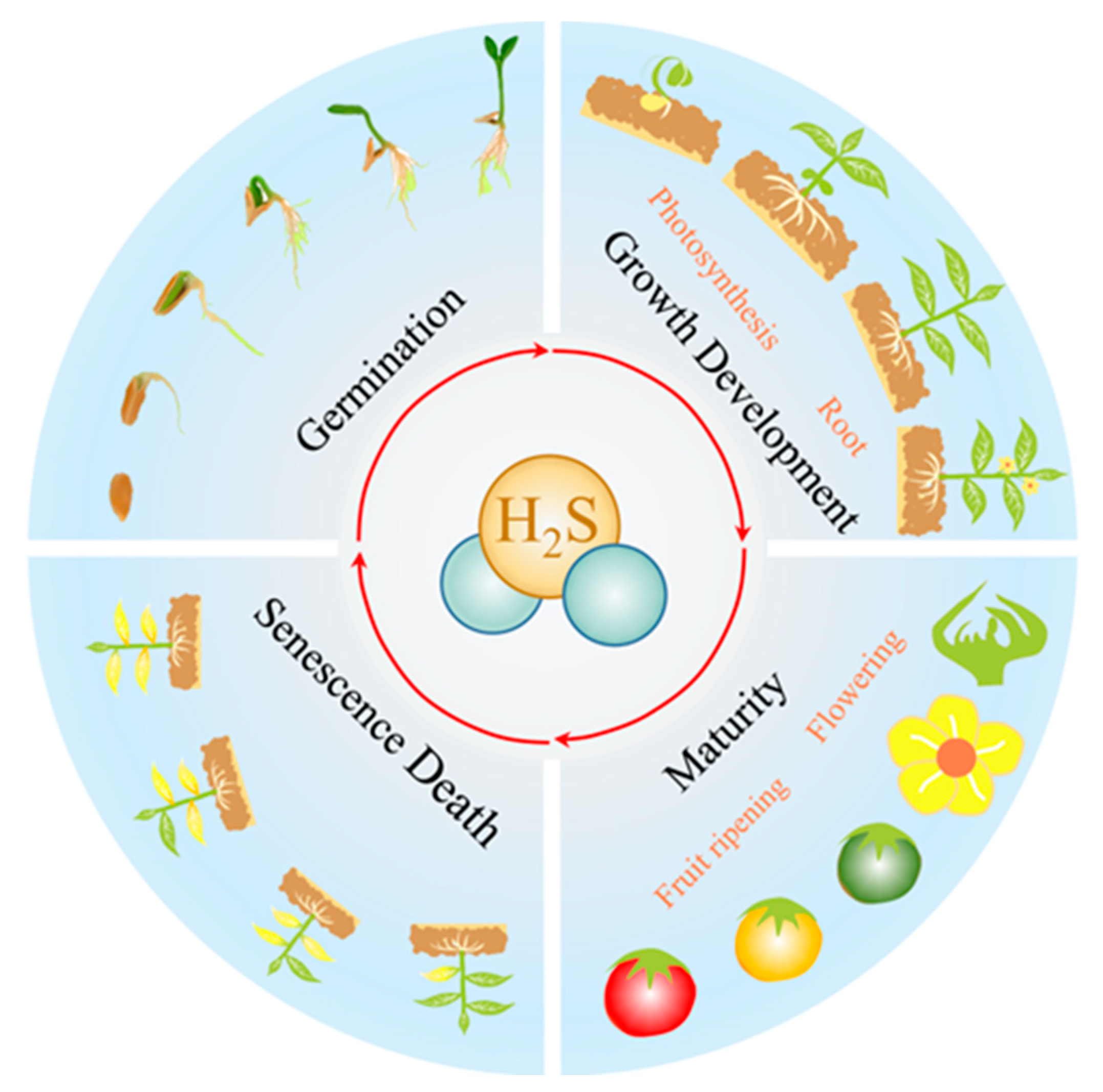
4. H2S Is Involved in Plant Growth and Development
4.1. H2S and Germination
4.2. H2S and Growth Development
4.2.1. H2S and Root Development
4.2.2. H2S and Photosynthesis Photomorphogenesis
4.3. H2S and Maturity
4.3.1. H2S and Flowering
4.3.2. H2S and Fruit Ripening
4.4. H2S and Senescence

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Development | Plant Species | H2S Doses | Effects | References |
|---|---|---|---|---|
| Germination | Arabidopsis thaliana | 12 μmol L−1 NaHS | Activated AOX Mediated cyanide-resistant respiration pathway | [83] |
| Arabidopsis thaliana | 0.1 mmol L−1 NaHS | Maintained the protein stability of ABSCISIC ACID-INSENSITIVE 4 | [1] | |
| Root development | Peach (Prunus persica (L.) Batsch) | 0.2 mmol L−1 NaHS | Increased the concentration of endogenous auxin Upregulated the expression of LATERAL ORGAN BOUNDARIES DOMAIN 16 | [145] |
| Cucumber (Cucumis sativus L.) | 100 μmol L−1 NaHS | Promoted the occurrence of adventitious roots Regulated osmotic substance content (proline) and enhanced antioxidant ability | [106] | |
| Photosynthesis | Wheat (Triticum aestivum L.) | 200 µmol L−1 NaHS | Increased photosynthesis and carbohydrate metabolism | [109] |
| Garlic (Allium sativum) | 200 µmol L−1 NaHS | Increased the values of net photosynthetic rate, transpiration rate and stomatal conductance | [108] | |
| Flowering | Heading Chinese cabbage (B. rapa L. syn. B. campestris L. ssp. chinensis Makino var. pekinensis (Rupr.) J. Cao et Sh. Cao) | 100 µmol L−1 NaHS | Promoted plant flowering by weakening or eliminating the binding abilities of BraFLCs to downstream promoters via S-sulfhydration | [120] |
| Maturity and senescence | Strawberry (Fragaria chiloensis (L.) Mill.) | 0.2 mmol L−1 NaHS | Maintained fruit firmness Delayed pectin degradation Downregulated the expression of polygalacturonase, pectate lyase, and expansin | [130] |
| Goji berry (Lycium barbarum L.) | 1.4 mmol L−1 NaHS | Improved postharvest quality Increased bioactive compounds accumulation Boosted antioxidant capacity | [128] |
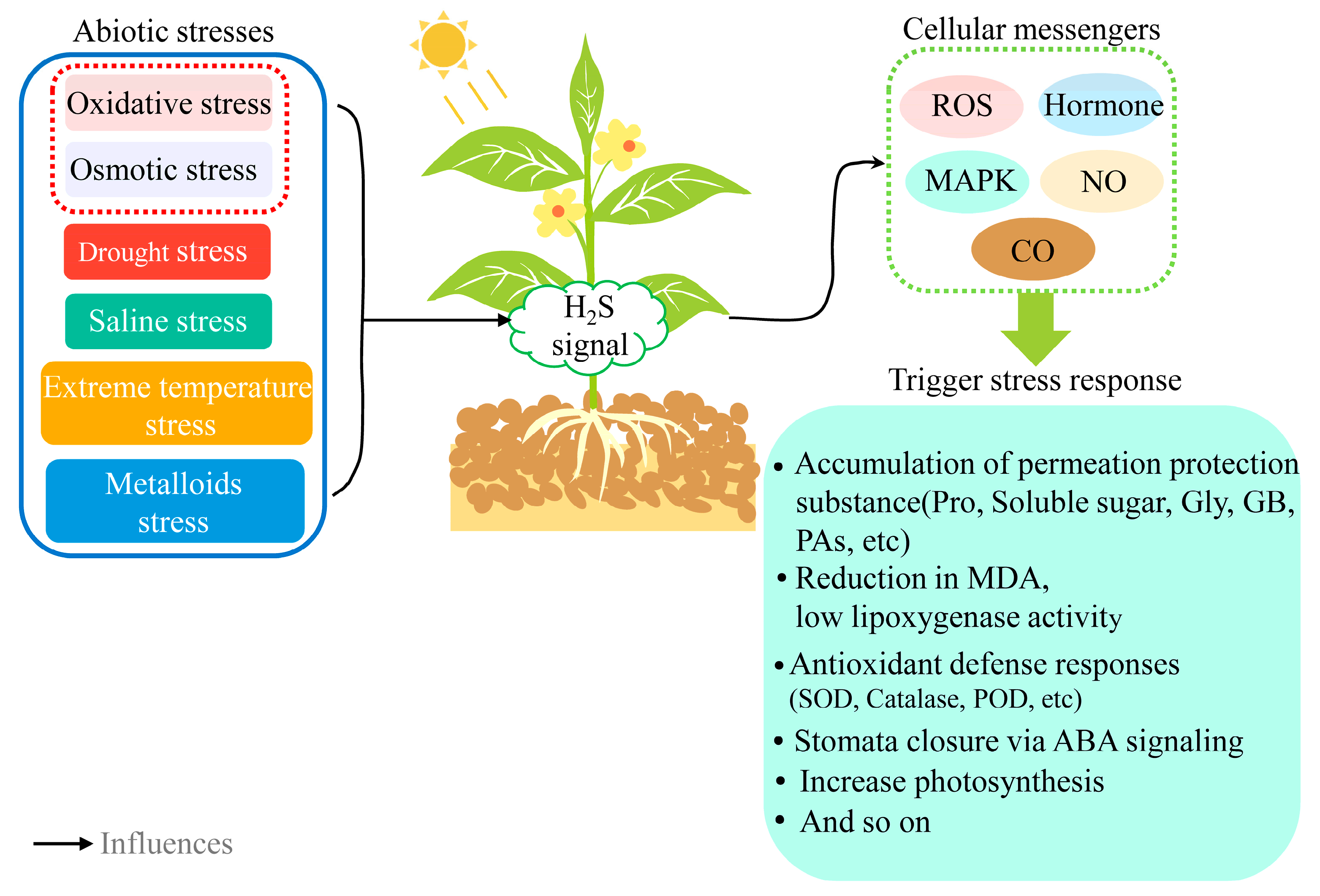
5. H2S Improves Plant Stress Tolerance
5.1. H2S and Oxidative Stress
5.2. H2S and Osmotic Stress
5.3. H2S and Drought Stress
5.4. H2S and Saline Stress
5.5. H2S and Extreme Temperature Stress
5.6. H2S and Metalloids Stress
| Plant Species | Stressors | H2S Doses | Protective Effects | References |
|---|---|---|---|---|
| Arabidopsis thaliana | Oxidative | 0.5 mmol L−1 NaHS | Repressed glycolate oxidase activities | [152] |
| Arabidopsis thaliana | Osmotic | 150 mmol L−1 NaHS | Involved in osmotic stress-triggered stomatal closure | [168] |
| Safflower | Drought: 70 and 50% field capacity | 0.5 and 1.0 mmol L−1 NaHS | Increased the accumulation of secondary metabolites Strengthened the antioxidant capacity Regulated elemental uptake | [171] |
| Wheat (Triticum aestivum L.) | Drought: 30% field capacity | 10 mg m−3 SO2 | Triggered proline accumulation Activated antioxidant enzymes Changed expression level of transcription factors Increased H2S content | [172] |
| Cyclocarya paliurus | Salinity: 100 mmol L−1 NaCl | 0.5 mmol L−1 NaHS | Maintained chlorophyll fluorescence Regulating nitric oxide level Improved antioxidant capacity | [194] |
| Wheat (Triticum aestivum L.) | Heat: 40 °C | 200 µmol L−1 NaHS | Reduced glucose sensitivity Increased the activities of SOD, catalase, and the AsA-GSH cycle | [211] |
| Pepper (Capsicum annuum L.) | Chilling: 10 °C/5 °C day/night | 1 mmol L−1 NaHS | Enhanced the antioxidant capacity Increased the enzyme transcription levels Reduced the contents of O2•−, H2O2, and MDA | [210] |
| Wheat (Triticum aestivum L.) Rice (Oryza sativa L. var.) | Metalloids: 20 μmol L−1 Cr(VI) | 15 μmol L−1 NaHS | Maintained fruit firmness Delayed pectin degradation Downregulated the expression of polygalacturonase, pectate lyase, and expansin | [221] |
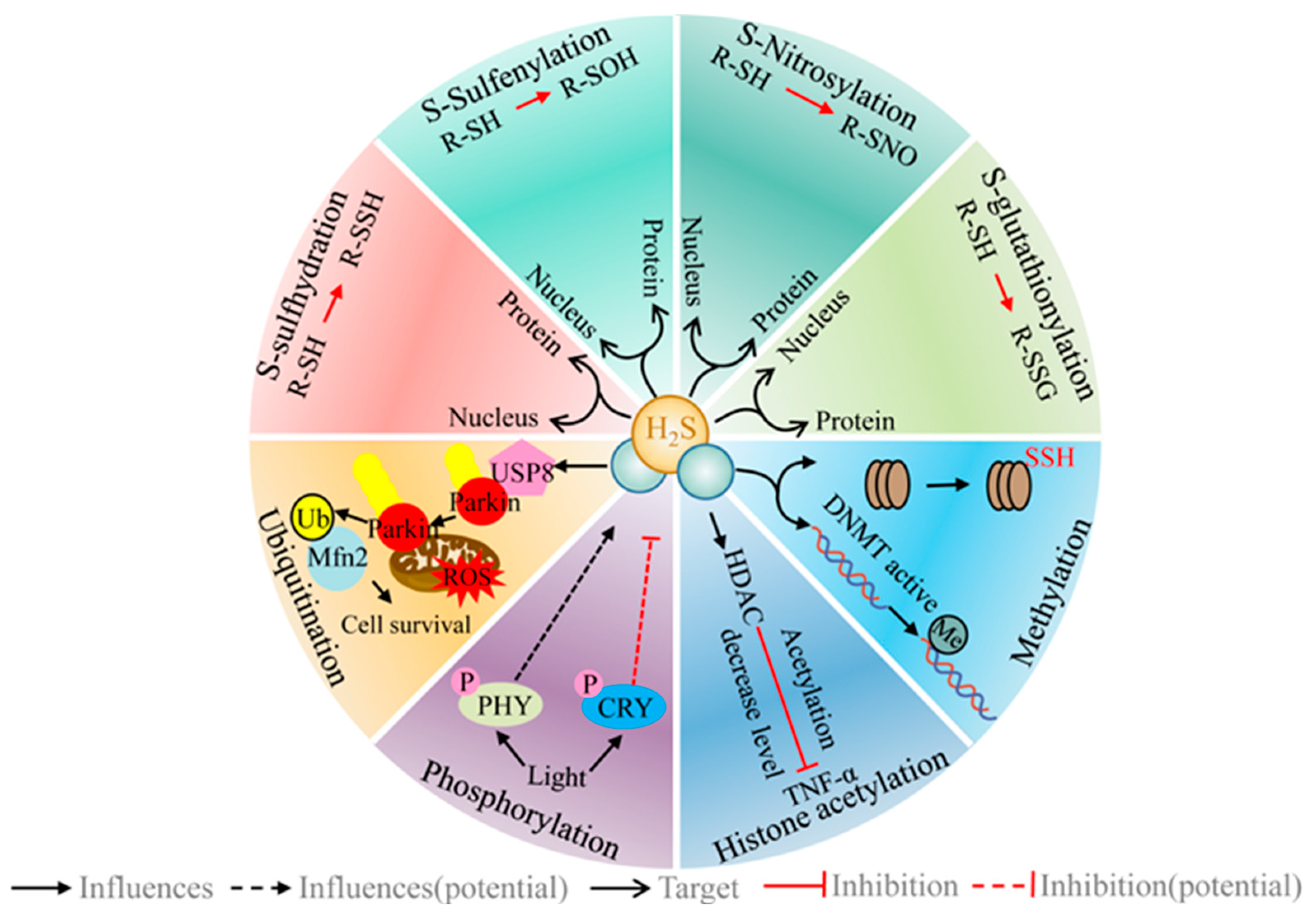
6. H2S Regulates a Range of Different PTMs/Signaling Systems
6.1. H2S Regulates the Post-Translational Modification of Protein Cysteine Residues (R-SH)
6.1.1. H2S and S-Sulfhydration
6.1.2. H2S and S-Sulfenylation
6.1.3. H2S and S-Nitrosylation
6.1.4. H2S and S-Glutathionylation
6.2. H2S and Phosphorylation
6.3. H2S and Ubiquitination (Ub)
6.4. H2S and Histone Acetylation
6.5. H2S and Methylation
7. Conclusions and Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhou, M.; Zhang, J.; Zhou, H.; Zhao, D.; Duan, T.; Wang, S.; Yuan, X.; Xie, Y. Hydrogen sulfide-linked persulfidation maintains protein stability of ABSCISIC ACID-INSENSITIVE 4 and delays seed germination. Int. J. Mol. Sci. 2022, 23, 1389. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.L.; Wang, L.X.; Shao, L.H.; Niu, J.; Zheng, F.X. Hydrogen sulfide generated by hydrogen peroxide mediates darkness-induced stomatal closure in Arabidopsis thaliana. Pak. J. Bot. 2022, 54, 57–64. [Google Scholar] [CrossRef]
- Mathur, P.; Roy, S.; Nasir Khan, M.; Mukherjee, S. Hydrogen sulphide (H2S) in the hidden half: Role in root growth, stress signalling and rhizospheric interactions. BMC Plant Biol. 2022, 24, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Feng, Y.X.; Lin, Y.J.; Yu, X.Z. Comparative effects of sodium hydrosulfide and proline on functional repair in rice chloroplast through the D1 protein and thioredoxin system under simulated thiocyanate pollution. Chemosphere 2021, 284, 131389. [Google Scholar] [CrossRef]
- Liu, D.; Pei, Y. The secret of H2S to keep plants young and fresh and its products. BMC Plant Biol. 2022, 24, 587–593. [Google Scholar] [CrossRef] [PubMed]
- Yao, G.F.; Li, C.; Sun, K.K.; Tang, J.; Huang, Z.Q.; Yang, F.; Huang, G.G.; Hu, L.Y.; Jin, P.; Hu, K.D.; et al. Hydrogen sulfide maintained the good appearance and nutrition in post-harvest tomato fruits by antagonizing the effect of ethylene. Front. Plant Sci. 2020, 11, 584. [Google Scholar] [CrossRef]
- Lindahl, T. An N-glycosidase from Escherichia coli that releases free uracil from DNA containing deaminated cytosine residues. Proc. Natl Acad. Sci. USA 1974, 71, 3649–3653. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, A.L.; Schaer, P. DNA glycosylases: In DNA repair and beyond. Chromosoma 2012, 121, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Rose, P.; Moore, P.K. Hydrogen sulfide and cell signaling. Annu. Rev. Pharmacol. Toxicol. 2011, 51, 169–187. [Google Scholar] [CrossRef] [Green Version]
- Gotor, C.; Garcia, I.; Aroca, A.; Laureano-Marin, A.M.; Arenas-Alfonseca, L.; Jurado-Flores, A.; Moreno, I.; Romero, L.C. Signaling by hydrogen sulfide and cyanide through post-translational modification. J. Exp. Bot. 2019, 70, 4251–4265. [Google Scholar] [CrossRef]
- Jin, Z.; Sun, L.; Yang, G.; Pei, Y. Hydrogen sulfide regulates energy production to delay leaf senescence induced by drought stress in Arabidopsis. Front. Plant Sci. 2018, 9, 1722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, G.Q.; Zhang, W.W.; Cao, H.; Yue, S.S.; Li, P.; Yang, H.Q. Effects hydrogen sulfide on the antioxidant system and membrane stability in mitochondria of Malus hupehensis under NaCl stress. Biol. Plant. 2019, 63, 228–236. [Google Scholar] [CrossRef]
- Hao, X.; Jin, Z.; Wang, Z.; Qin, W.; Pei, Y. Hydrogen sulfide mediates DNA methylation to enhance osmotic stress tolerance in Setaria italica L. Plant Soil 2020, 453, 355–370. [Google Scholar] [CrossRef]
- Thakur, M.; Anand, A. Hydrogen sulfide: An emerging signaling molecule regulating drought stress response in plants. Physio. Plant. 2021, 172, 1227–1243. [Google Scholar] [CrossRef]
- Li, H.; Yu, T.T.; Ning, Y.S.; Li, H.; Zhang, W.W.; Yang, H.Q. Hydrogen sulfide alleviates alkaline salt stress by regulating the expression of microRNAs in Malus hupehensis rehd. roots. Front. Plant Sci. 2021, 12, 663519. [Google Scholar] [CrossRef]
- Mishra, S.; Chowdhary, A.A.; Bhau, B.S.; Srivastava, V. Hydrogen sulphide-mediated alleviation and its interplay with other signalling molecules during temperature stress. BMC Plant Biol. 2022, 24, 569–575. [Google Scholar] [CrossRef]
- Arif, M.S.; Yasmeen, T.; Abbas, Z.; Ali, S.; Rizwan, M.; Aljarba, N.H.; Alkahtani, S.; Abdel-Daim, M.M. Role of exogenous and endogenous hydrogen sulfide (H2S) on functional traits of plants under heavy metal stresses: A recent perspective. Front. Plant Sci. 2021, 11, 545453. [Google Scholar] [CrossRef]
- Filipovic, M.R. Persulfidation (S-sulfhydration) and H2S. Handb. Exp. Pharmacol. 2015, 230, 29–59. [Google Scholar] [CrossRef]
- Wang, P.; Fang, H.; Gao, R.; Liao, W. Protein persulfidation in plants: Function and mechanism. Antioxidants 2021, 10, 1631. [Google Scholar] [CrossRef]
- Siodmak, A.; Hirt, H. Stomatal regulation: Role of H2S-induced persulfidation in ABA signaling. Mol. Plant. 2021, 14, 858–860. [Google Scholar] [CrossRef]
- Liu, D.; Guo, T.; Li, J.; Hao, Y.; Zhao, D.; Wang, L.; Liu, Z.; Zhang, L.; Jin, Z.; Pei, Y. Hydrogen sulfide inhibits the abscission of tomato pedicel through reconstruction of a basipetal auxin gradient. Plant Sci. 2022, 318, 111219. [Google Scholar] [CrossRef] [PubMed]
- Kaya, C.; Sarioglu, A.; Ashraf, M.; Alyemeni, M.N.; Ahmad, P. Gibberellic acid-induced generation of hydrogen sulfide alleviates boron toxicity in tomato (Solanum lycopersicum L.) plants. Plant Physiol. Biochem. 2020, 153, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Kaur, H.; Hussain, S.J.; Al-Huqail, A.A.; Siddiqui, M.H.; Al-Huqail, A.A.; Khan, M.I.R. Hydrogen sulphide and salicylic acid regulate antioxidant pathway and nutrient balance in mustard plants under cadmium stress. BMC Plant Biol. 2022, 24, 660–669. [Google Scholar] [CrossRef]
- Tian, B.; Zhang, Y.; Jin, Z.; Liu, Z.; Pei, Y. Role of hydrogen sulfide in the methyl jasmonate response to cadmium stress in foxtail millet. Front. Biosci. 2017, 22, 530–538. [Google Scholar] [CrossRef] [Green Version]
- Hu, D.; Wei, L.; Liao, W. Brassinosteroids in plants: Crosstalk with small-molecule compounds. Biomolecules 2021, 11, 1800. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Zhang, J.; Zhou, M.; Zhou, H.; Cui, B.; Gotor, C.; Romero, L.C.; Fu, L.; Yang, J.; Foyer, C.H.; et al. Persulfidation-based modification of cysteine desulfhydrase and the NADPH oxidase RBOHD controls guard cell abscisic acid signaling. Plant Cell 2020, 32, 1000–1017. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Jia, H.; Wang, X.; Shi, C.; Wang, X.; Ma, P.; Wang, J.; Ren, M.; Li, J. Hydrogen sulfide positively regulates abscisic acid signaling through persulfidation of SnRK2.6 in guard cells. Mol. Plant. 2020, 13, 732–744. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Zhang, J.; Shen, J.; Zhou, H.; Zhao, D.; Gotor, C.; Romero, L.C.; Fu, L.; Li, Z.; Yang, J.; et al. Hydrogen sulfide-linked persulfidation of ABI4 controls ABA responses through the transactivation of MAPKKK18 in Arabidopsis. Mol. Plant. 2021, 14, 921–936. [Google Scholar] [CrossRef]
- Benavides, G.A.; Squadrito, G.L.; Mills, R.W.; Patel, H.D.; Isbell, T.S.; Patel, R.P.; Darley-Usmar, V.M.; Doeller, J.E.; Kraus, D.W. Hydrogen sulfide mediates the vasoactivity of garlic. Proc. Natl Acad. Sci. USA 2007, 104, 17977–17982. [Google Scholar] [CrossRef] [Green Version]
- DeLeon, E.R.; Stoy, G.F.; Olson, K.R. Passive loss of hydrogen sulfide in biological experiments. Anal. Biochem. 2012, 421, 203–207. [Google Scholar] [CrossRef]
- Powell, C.R.; Dillon, K.M.; Matson, J.B. A review of hydrogen sulfide (H2S) donors: Chemistry and potential therapeutic applications. Biochem. Pharmacol. 2018, 149, 110–123. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Minkler, P.; Grove, D.; Wang, R.; Willard, B.; Dweik, R.; Hine, C. Non-enzymatic hydrogen sulfide production from cysteine in blood is catalyzed by iron and vitamin B6. Commun. Biol. 2019, 2019, 194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, B.; Bhattacharya, R.; Mukherjee, P. Hydrogen sulfide signaling in mitochondria and disease. FASEB J. 2019, 33, 13098–13125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kabil, O.; Banerjee, R. Enzymology of H2S biogenesis, decay and signaling. Antioxid. Redox Signal. 2014, 20, 770–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sbodio, J.I.; Snyder, S.H.; Paul, B.D. Regulators of the transsulfuration pathway. Br. J. Pharmacol. 2019, 176, 583–593. [Google Scholar] [CrossRef] [Green Version]
- Rose, P.; Moore, P.K.; Zhu, Y.Z. H2S biosynthesis and catabolism: New insights from molecular studies. Cell. Mol. Life Sci. 2017, 74, 1391–1412. [Google Scholar] [CrossRef] [Green Version]
- Shibuya, N.; Tanaka, M.; Yoshida, M.; Ogasawara, Y.; Togawa, T.; Ishii, K.; Kimura, H. 3-Mercaptopyruvate sulfurtransferase produces hydrogen sulfide and bound sulfane sulfur in the brain. Antioxid. Redox Signal. 2009, 11, 703–714. [Google Scholar] [CrossRef]
- Yadav, P.K.; Yamada, K.; Chiku, T.; Koutmos, M.; Banerjee, R. Structure and kinetic analysis of H2S production by human mercaptopyruvate sulfurtransferase. J. Biol. Chem. 2013, 288, 20002–20013. [Google Scholar] [CrossRef] [Green Version]
- Nagahara, N. Multiple role of 3-mercaptopyruvate sulfurtransferase: Antioxidative function, H2S and polysulfide production and possible SOx production. Br. J. Pharmacol. 2018, 175, 577–589. [Google Scholar] [CrossRef] [Green Version]
- Kimura, H. Physiological role of hydrogen sulfide and polysulfide in the central nervous system. Neurochem. Int. 2013, 63, 492–497. [Google Scholar] [CrossRef]
- da Costa Marques, L.A.; Teixeira, S.A.; de Jesus, F.N.; Wood, M.E.; Torregrossa, R.; Whiteman, M.; Costa, S.K.P.; Muscara, M.N. Vasorelaxant activity of AP39, a mitochondria-targeted H2S donor, on mouse mesenteric artery rings in vitro. Biomolecules 2022, 12, 280. [Google Scholar] [CrossRef] [PubMed]
- Aroca, A.; Gotor, C.; Romero, L.C. Hydrogen sulfide signaling in plants: Emerging roles of protein persulfidation. Front. Plant Sci. 2018, 9, 1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youssefian, S.; Nakamura, M.; Sano, H. Tobacco plants transformed with the O-acetylserine (thiol) lyase gene of wheat are resistant to toxic levels of hydrogen sulphide gas. Plant J. Cell Mol. Biol. 1993, 4, 759–769. [Google Scholar] [CrossRef]
- Nakayama, M.; Akashi, T.; Hase, T. Plant sulfite reductase: Molecular structure, catalytic function and interaction with ferredoxin. J. Inorg. Biochem. 2000, 82, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Kurmanbayeva, A.; Brychkova, G.; Bekturova, A.; Khozin, I.; Standing, D.; Yarmolinsky, D.; Sagi, M. Determination of total sulfur, sulfate, sulfite, thiosulfate, and sulfolipids in plants. Methods Mol. Biol. 2017, 1631, 253–271. [Google Scholar] [CrossRef] [PubMed]
- Pilon-Smits, E.A.H.; Garifullina, G.F.; Abdel-Ghany, S.; Kato, S.-I.; Mihara, H.; Hale, K.L.; Burkhead, J.L.; Esaki, N.; Kurihara, T.; Pilon, M. Characterization of a NifS-like chloroplast protein from Arabidopsis. Implications for its role in sulfur and selenium metabolism. Plant Physiol. 2002, 130, 1309–1318. [Google Scholar] [CrossRef] [Green Version]
- Hoewyk, D.V.; Pilon, M.; Pilon-Smits, E.A.H. The functions of NifS-like proteins in plant sulfur and selenium metabolism. Plant Sci. 2008, 174, 117–123. [Google Scholar] [CrossRef]
- Wirtz, M.; Hell, R. Functional analysis of the cysteine synthase protein complex from plants: Structural, biochemical and regulatory properties. J. Plant Physiol. 2006, 163, 273–286. [Google Scholar] [CrossRef]
- Frazzon, A.P.G.; Ramirez, M.V.; Warek, U.; Balk, J.; Frazzon, J.; Dean, D.R.; Winkel, B.S.J. Functional analysis of Arabidopsis genes involved in mitochondrial iron-sulfur cluster assembly. Plant Mol. Biol. 2007, 64, 225–240. [Google Scholar] [CrossRef]
- Bloem, E.; Rubekin, K.; Haneklaus, S.; Banfalvi, Z.; Hesse, H.; Schnug, E. H2S and COS gas exchange of transgenic potato lines with modified expression levels of enzymes involved in sulphur metabolism. J. Agron. Crop Sci. 2011, 197, 311–321. [Google Scholar] [CrossRef]
- Yamasaki, H.; Cohen, M.F. Biological consilience of hydrogen sulfide and nitric oxide in plants: Gases of primordial earth linking plant, microbial and animal physiologies. Nitric Oxide 2016, 55–56, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Nagy, P.; Winterbourn, C.C. Rapid reaction of hydrogen sulfide with the neutrophil oxidant hypochlorous acid to generate polysulfides. Chem. Res. Toxicol. 2010, 23, 1541–1543. [Google Scholar] [CrossRef]
- Sun, H.J.; Wu, Z.Y.; Cao, L.; Zhu, M.Y.; Nie, X.W.; Huang, D.J.; Sun, M.T.; Bian, J.S. Role of nitroxyl (HNO) in cardiovascular system: From biochemistry to pharmacology. Pharmacol. Res. 2020, 159, 10. [Google Scholar] [CrossRef] [PubMed]
- Filipovic, M.R.; Miljkovic, J.L.; Nauser, T.; Royzen, M.; Klos, K.; Shubina, T.; Koppenol, W.H.; Lippard, S.J.; Ivanovic-Burmazovic, I. Chemical characterization of the smallest S-nitrosothiol, HSNO; cellular cross-talk of H2S and S-nitrosothiols. J. Am. Chem. Soc. 2012, 134, 12016–12027. [Google Scholar] [CrossRef] [PubMed]
- Nicholls, P.; Marshall, D.C.; Cooper, C.E.; Wilson, M.T. Sulfide inhibition of and metabolism by cytochrome c oxidase. Biochem. Soc. Trans. 2013, 41, 1312–1316. [Google Scholar] [CrossRef] [Green Version]
- Hill, B.C.; Woon, T.C.; Nicholls, P.; Peterson, J.; Greenwood, C.; Thomson, A.J. Interactions of sulphide and other ligands with cytochrome c oxidase. An electron-paramagnetic-resonance study. Biochem. J. 1984, 224, 591–600. [Google Scholar] [CrossRef] [Green Version]
- Modis, K.; Coletta, C.; Erdelyi, K.; Papapetropoulos, A.; Szabo, C. Intramitochondrial hydrogen sulfide production by 3-mercaptopyruvate sulfurtransferase maintains mitochondrial electron flow and supports cellular bioenergetics. FASEB J. 2013, 27, 601–611. [Google Scholar] [CrossRef]
- Corpas, F.J.; Gonzalez-Gordo, S.; Palma, J.M. Nitric oxide and hydrogen sulfide modulate the NADPH-generating enzymatic system in higher plants. J. Exp. Bot. 2021, 72, 830–847. [Google Scholar] [CrossRef]
- Gerush, I.V.; Ferenchuk, Y.O. Hydrogen sulfide and mitochondria. Biopolym. Cell 2019, 35, 3–15. [Google Scholar] [CrossRef] [Green Version]
- Chatzianastasiou, A.; Bibli, S.I.; Andreadou, I.; Efentakis, P.; Kaludercic, N.; Wood, M.E.; Whiteman, M.; Di Lisa, F.; Daiber, A.; Manolopoulos, V.G.; et al. Cardioprotection by H2S donors: Nitric oxide-dependent and independent mechanisms. J. Pharmacol. Exp. Ther. 2016, 358, 431–440. [Google Scholar] [CrossRef]
- Zhou, Z.; von Wantoch Rekowski, M.; Coletta, C.; Szabo, C.; Bucci, M.; Cirino, G.; Topouzis, S.; Papapetropoulos, A.; Giannis, A. Thioglycine and l-thiovaline: Biologically active H2S-donors. Bioorg. Med. Chem. 2012, 20, 2675–2678. [Google Scholar] [CrossRef] [PubMed]
- Latorre, E.; Torregrossa, R.; Wood, M.E.; Whiteman, M.; Harries, L.W. Mitochondria-targeted hydrogen sulfide attenuates endothelial senescence by selective induction of splicing factors HNRNPD and SRSF2. Aging 2018, 10, 1666–1681. [Google Scholar] [CrossRef] [PubMed]
- Carter, J.M.; Brown, E.M.; Irish, E.E.; Bowden, N.B. Characterization of dialkyldithiophosphates as slow hydrogen sulfide releasing chemicals and their effect on the growth of maize. J. Agric. Food Chem. 2019, 67, 11883–11892. [Google Scholar] [CrossRef] [PubMed]
- Santisree, P.; Adimulam, S.S.; Bommineni, P.; Bhatnagar-Mathur, P.; Sharma, K.K. Hydrogen sulfide in plant abiotic stress tolerance. In Reactive Oxygen, Nitrogen and Sulfur Species in Plants: Production, Metabolism, Signaling and Defense Mechanisms, 1st ed.; Hasanuzzaman, M., Fotopoulos, V., Nahar, K., Fujita, M., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2019; Volume 2, pp. 743–775. [Google Scholar]
- Guo, H.; Xiao, T.; Zhou, H.; Xie, Y.; Shen, W. Hydrogen sulfide: A versatile regulator of environmental stress in plants. Acta Physiol. Plant. 2016, 38, 1–13. [Google Scholar] [CrossRef]
- Gao, X.H.; Krokowski, D.; Guan, B.-J.; Bederman, I.; Majumder, M.; Parisien, M.; Diatchenko, L.; Kabil, O.; Willard, B.; Banerjee, R.; et al. Quantitative H2S-mediated protein sulfhydration reveals metabolic reprogramming during the integrated stress response. Elife 2015, 4, e10067. [Google Scholar] [CrossRef]
- Chen, Y.; Zhu, C.; Yang, Z.; Chen, J.; He, Y.; Jiao, Y.; He, W.; Qiu, L.; Cen, J.; Guo, Z. A ratiometric fluorescent probe for rapid detection of hydrogen sulfide in mitochondria. Angew. Chem. 2013, 52, 1688–1691. [Google Scholar] [CrossRef]
- Yu, F.; Li, P.; Song, P.; Wang, B.; Zhao, J.; Han, K. An ICT-based strategy to a colorimetric and ratiometric fluorescence probe for hydrogen sulfide in living cells. Chem. Commun. 2012, 48, 2852–2854. [Google Scholar] [CrossRef]
- Feng, X.; Zhang, T.; Liu, J.T.; Miao, J.Y.; Zhao, B.X. A new ratiometric fluorescent probe for rapid, sensitive and selective detection of endogenous hydrogen sulfide in mitochondria. Chem. Commun. 2016, 52, 3131–3134. [Google Scholar] [CrossRef]
- Yuan, L.; Zuo, Q.P. FRET-Based mitochondria-targetable dual-excitation ratiometric fluorescent probe for monitoring hydrogen sulfide in living cells. Chem.-Asian J. 2014, 9, 1544–1549. [Google Scholar] [CrossRef]
- Liu, X.L.; Du, X.J.; Dai, C.G.; Song, Q.H. Ratiometric two-photon fluorescent probes for mitochondria hydrogen sulfide in living cells. J. Org. Chem. 2014, 79, 9481–9489. [Google Scholar] [CrossRef]
- Szczesny, B.; Modis, K.; Yanagi, K.; Coletta, C.; Le Trionnaire, S.; Perry, A.; Wood, M.E.; Whiteman, M.; Szabo, C. AP39 [10-oxo-10-(4-(3-thioxo-3H-1,2-dithiol-5yl)phenoxy)decyl) triphenylphosphonium bromide], a mitochondrially targeted hydrogen sulfide donor, stimulates cellular bioenergetics, exerts cytoprotective effects and protects against the loss of mitochondrial DNA integrity in oxidatively stressed endothelial cells in vitro. Nitric Oxide 2014, 41, 120–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gero, D.; Torregrossa, R.; Perry, A.; Waters, A.; Le-Trionnaire, S.; Whatmore, J.L.; Wood, M.; Whiteman, M. The novel mitochondria-targeted hydrogen sulfide (H2S) donors AP123 and AP39 protect against hyperglycemic injury in microvascular endothelial cells in vitro. Pharmacol. Res. 2016, 113, 186–198. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, A.K.; Gadalla, M.M.; Snyder, S.H. Signaling by gasotransmitters. Sci. Signal. 2009, 2, re2. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, N.; Fu, C.; Pappin, D.J.; Tonks, N.K. H2S-induced sulfhydration of the phosphatase PTP1B and its role in the endoplasmic reticulum stress response. Sci. Signal. 2011, 4, ra86. [Google Scholar] [CrossRef] [Green Version]
- Sen, N.; Paul, B.D.; Gadalla, M.M.; Mustafa, A.K.; Sen, T.; Xu, R.; Kim, S.; Snyder, S.H. Hydrogen sulfide-linked sulfhydration of NF-κB mediates its antiapoptotic actions. Mol. Cell 2012, 45, 13–24. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Macinkovic, I.; Devarie-Baez, N.O.; Pan, J.; Park, C.-M.; Carroll, K.S.; Filipovic, M.R.; Xian, M. Detection of protein S-sulfhydration by a tag-switch technique. Angew. Chem. 2014, 53, 575–581. [Google Scholar] [CrossRef] [Green Version]
- Zivanovic, J.; Kouroussis, E.; Kohl, J.B.; Adhikari, B.; Bursac, B.; Schott-Roux, S.; Petrovic, D.; Miljkovic, J.L.; Thomas-Lopez, D.; Jung, Y.; et al. Selective persulfide detection reveals evolutionarily conserved antiaging effects of S-sulfhydration. Cell Metab. 2019, 30, 1152–1170. [Google Scholar] [CrossRef]
- Filipovic, M.R.; Zivanovic, J.; Alvarez, B.; Banerjee, R. Chemical biology of H2S signaling through persulfidation. Chem. Rev. 2018, 118, 1253–1337. [Google Scholar] [CrossRef]
- Baudouin, E.; Poilevey, A.; Hewage, N.I.; Cochet, F.; Puyaubert, J.; Bailly, C. The significance of hydrogen sulfide for Arabidopsis seed germination. Front. Plant Sci. 2016, 7, 930. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.G.; Gong, M.; Liu, P. Hydrogen sulfide is a mediator in H2O2-induced seed germination in Jatropha curcas. Acta Physiol. Plant. 2012, 34, 2207–2213. [Google Scholar] [CrossRef]
- Li, Z.G.; He, Q.Q. Hydrogen peroxide might be a downstream signal molecule of hydrogen sulfide in seed germination of mung bean (Vigna radiata). Biologia 2015, 70, 753–759. [Google Scholar] [CrossRef]
- Fang, H.; Liu, R.; Yu, Z.; Wu, G.; Pei, Y. Gasotransmitter H2S accelerates seed germination via activating AOX mediated cyanide-resistant respiration pathway. Plant Physiol. Biochem. 2021, 190, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Dou, W.; Jiang, C.X.; Wei, Z.J.; Liu, J.; Jones, R.L. Hydrogen sulfide stimulates ß-amylase activity during early stages of wheat grain germination. Plant Signal. Behav. 2010, 5, 1031–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.H.; Wang, Y.; Ye, X.Y.; Li, Z.G. Signaling molecule hydrogen sulfide improves seed germination and seedling growth of maize (Zea mays L.) under high temperature by inducing antioxidant system and osmolyte biosynthesis. Front. Plant Sci. 2018, 9, 1288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Li, L.; Cui, W.; Xu, S.; Shen, W.; Wang, R. Hydrogen sulfide enhances alfalfa (Medicago sativa) tolerance against salinity during seed germination by nitric oxide pathway. Plant Soil 2012, 351, 107–119. [Google Scholar] [CrossRef]
- Villordon, A.Q.; Ginzberg, I.; Firon, N. Root architecture and root and tuber crop productivity. Trends Plant Sci. 2014, 19, 419–425. [Google Scholar] [CrossRef]
- Mukherjee, S.; Corpas, F.J. Crosstalk among hydrogen sulfide (H2S), nitric oxide (NO) and carbon monoxide (CO) in root-system development and its rhizosphere interactions: A gaseous interactome. Plant Physiol. Biochem. 2020, 155, 800–814. [Google Scholar] [CrossRef]
- Motte, H.; Vanneste, S.; Beeckman, T. Molecular and environmental regulation of root development. Annu. Rev. Plant Biol. 2019, 70, 465–488. [Google Scholar] [CrossRef] [Green Version]
- Pagnussat, G.C.; Simontacchi, M.; Puntarulo, S.; Lamattina, L. Nitric oxide is required for root organogenesis. Plant Physiol. 2002, 129, 954–956. [Google Scholar] [CrossRef] [Green Version]
- Kopyra, M.; Gwód, E.w.A. Nitric oxide stimulates seed germination and counteracts the inhibitory effect of heavy metals and salinity on root growth of Lupinus luteus. Plant Physiol. Biochem. 2003, 41, 1011–1017. [Google Scholar] [CrossRef]
- Li, C.; Huang, D.; Wang, C.; Wang, N.; Yao, Y.; Li, W.; Liao, W. NO is involved in H2-induced adventitious rooting in cucumber by regulating the expression and interaction of plasma membrane H+-ATPase and 14-3-3. Planta 2020, 252, 9. [Google Scholar] [CrossRef]
- Wei, M.Y.; Li, H.; Zhong, Y.H.; Shen, Z.J.; Ma, D.N.; Gao, C.H.; Liu, Y.L.; Wang, W.H.; Zhang, J.Y.; You, Y.P.; et al. Transcriptomic analyses reveal the effect of nitric oxide on the lateral root development and growth of mangrove plant Kandelia obovata. Plant Soil 2022, 472, 543–564. [Google Scholar] [CrossRef]
- Sharma, S.; Singh, H.P.; Batish, D.R.; Kohli, R.K. Nitric oxide induced modulations in adventitious root growth, lignin content and lignin synthesizing enzymes in the hypocotyls of Vigna radiata. Plant Physiol. Biochem. 2019, 141, 225–230. [Google Scholar] [CrossRef]
- Correa-Aragunde, N.; Graziano, M.; Chevalier, C.; Lamattina, L. Nitric oxide modulates the expression of cell cycle regulatory genes during lateral root formation in tomato. J. Exp. Bot. 2006, 57, 581–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, X.; Li, Y.; Lu, R.; Cheng, P.; Zhang, Y.; Li, L.; Wang, R.; Cui, J.; Shen, W. Methane-induced lateral root formation requires the participation of nitric oxide signaling. Plant Physiol. Biochem. 2020, 147, 262–271. [Google Scholar] [CrossRef] [PubMed]
- Qi, F.; Xiang, Z.; Kou, N.; Cui, W.; Xu, D.; Wang, R.; Zhu, D.; Shen, W. Nitric oxide is involved in methane-induced adventitious root formation in cucumber. Physio. Plant. 2017, 159, 366–377. [Google Scholar] [CrossRef] [Green Version]
- Cao, Z.Y.; Wei, X.; Liu, Z.Y.; Li, X.N.; Shen, W.B. Carbon monoxide promotes lateral root formation in rapeseed. J. Integr. Plant Biol. 2007, 49, 1070–1079. [Google Scholar] [CrossRef]
- Santa-Cruz, D.M.; Pacienza, N.A.; Polizio, A.H.; Balestrasse, K.B.; Tomaro, M.L.; Yannarelli, G.G. Nitric oxide synthase-like dependent NO production enhances heme oxygenase up-regulation in ultraviolet-B-irradiated soybean plants. Phytochemistry 2010, 71, 1700–1707. [Google Scholar] [CrossRef]
- Jia, H.; Hu, Y.; Fan, T.; Li, J. Hydrogen sulfide modulates actin-dependent auxin transport via regulating ABPs results in changing of root development in Arabidopsis. Sci. Rep. 2015, 5, 8251. [Google Scholar] [CrossRef] [Green Version]
- Kolluru, G.K.; Yuan, S.; Shen, X.; Kevil, C.G. H2S regulation of nitric oxide metabolism. In Methods in Enzymology; Cadenas, E., Packer, L., Eds.; Academic Press: New York, NY, USA, 2015; Volume 554, pp. 271–297. [Google Scholar]
- Stevenson, F.J. Humus chemistry: Genesis, composition, reactions. Soil Sci. 1994, 135, 129–130. [Google Scholar]
- Selvig, K.; Alspaugh, J.A. pH response pathways in fungi: Adapting to host-derived and environmental signals. Mycobiology 2011, 39, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Mora, V.; Bacaicoa, E.; Zamarreno, A.M.; Aguirre, E.; Garnica, M.; Fuentes, M.; Garcia-Mina, J.M. Action of humic acid on promotion of cucumber shoot growth involves nitrate-related changes associated with the root-to-shoot distribution of cytokinins, polyamines and mineral nutrients. J. Plant Physiol. 2010, 167, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, W.H.; Wu, F.H.; You, C.Y.; Liu, T.W.; Dong, X.J.; He, J.X.; Zheng, H.L. Hydrogen sulfide alleviates aluminum toxicity in barley seedlings. Plant Soil 2013, 362, 301–318. [Google Scholar] [CrossRef]
- Liu, Y.; Wei, L.; Feng, L.; Zhang, M.; Hu, D.; Tie, J.; Liao, W. Hydrogen sulfide promotes adventitious root development in cucumber under salt stress by enhancing antioxidant ability. Plants 2022, 11, 935. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Luo, Q.; Wang, R.; Xu, J. Hydrogen sulfide toxicity inhibits primary root growth through the ROS-NO pathway. Sci. Rep. 2017, 7, 868. [Google Scholar] [CrossRef] [Green Version]
- Gharehbaghli, N.; Sepehri, A. The ameliorative effect of hydrogen sulfide on cadmium toxicity and oxidative stress damage in garlic (Allium sativum) seedlings. S. Afr. J. Bot. 2022, 150, 161–170. [Google Scholar] [CrossRef]
- Iqbal, N.; Fatma, M.; Gautam, H.; Umar, S.; Sofo, A.; D’Ippolito, I.; Khan, N.A. The crosstalk of melatonin and hydrogen sulfide determines photosynthetic performance by regulation of carbohydrate metabolism in wheat under heat stress. Plants 2021, 10, 1778. [Google Scholar] [CrossRef]
- Liu, Y.H.; Zhang, X.H.; Liu, B.W.; Ao, B.; Liu, Q.; Wen, S.Y.; Xu, Y.F. Hydrogen sulfide regulates photosynthesis of tall fescue under low-light stress. Photosynthetica 2019, 57, 714–723. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Moreiras, A.M.; Reigosa, M.J. Advances in Plant Ecophysiology Techniques; Springer: Cham, Switzerland, 2018. [Google Scholar]
- Li, J.; Shi, C.; Wang, X.; Liu, C.; Ding, X.; Ma, P.; Wang, X.; Jia, H. Hydrogen sulfide regulates the activity of antioxidant enzymes through persulfidation and improves the resistance of tomato seedling to copper oxide nanoparticles (CuO NPs)-induced oxidative stress. Plant Physiol. Biochem. 2020, 156, 257–266. [Google Scholar] [CrossRef]
- Li, S.P.; Hu, K.D.; Hu, L.Y.; Li, Y.H.; Jiang, A.M.; Xiao, F.; Han, Y.; Liu, Y.S.; Zhang, H. Hydrogen sulfide alleviates postharvest senescence of broccoli by modulating antioxidant defense and senescence-related gene expression. J. Agric. Food Chem. 2014, 62, 1119–1129. [Google Scholar] [CrossRef]
- Chen, J.; Wu, F.H.; Wang, W.H.; Zheng, C.J.; Lin, G.H.; Dong, X.J.; He, J.X.; Pei, Z.M.; Zheng, H.L. Hydrogen sulphide enhances photosynthesis through promoting chloroplast biogenesis, photosynthetic enzyme expression, and thiol redox modification in Spinacia oleracea seedlings. J. Exp. Bot. 2011, 62, 4481–4493. [Google Scholar] [CrossRef] [PubMed]
- Honda, K.; Yamada, N.; Yoshida, R.; Ihara, H.; Sawa, T.; Akaike, T.; Iwai, S. 8-Mercapto-cyclic GMP mediates hydrogen sulfide-induced stomatal closure in Arabidopsis. Plant Cell Physiol. 2015, 56, 1481–1489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medeiros, D.B.; Barros, J.A.S.; Fernie, A.R.; Araujo, W.L. Eating away at ROS to regulate stomatal opening. Trends Plant Sci. 2020, 25, 220–223. [Google Scholar] [CrossRef] [PubMed]
- Scuffi, D.; Nietzel, T.; Di Fino, L.M.; Meyer, A.J.; Lamattina, L.; Schwarzlaender, M.; Laxalt, A.M.; Garcia-Mata, C. Hydrogen sulfide increases production of NADPH oxidase-dependent hydrogen peroxide and phospholipase D-derived phosphatidic acid in guard cell signaling. Plant Physiol. 2018, 176, 2532–2542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, H.; Chen, S.; Liu, D.; Liesche, J.; Shi, C.; Wang, J.; Ren, M.; Wang, X.; Yang, J.; Shi, W.; et al. Ethylene-induced hydrogen sulfide negatively regulates ethylene biosynthesis by persulfidation of ACO in tomato under osmotic stress. Front. Plant Sci. 2018, 9, 1517. [Google Scholar] [CrossRef] [Green Version]
- Busch, F.A. Opinion: The red-light response of stomatal movement is sensed by the redox state of the photosynthetic electron transport chain. Photosynth. Res. 2014, 119, 131–140. [Google Scholar] [CrossRef]
- Ma, X.; Zhang, L.; Pei, Z.; Zhang, L.; Liu, Z.; Liu, D.; Hao, X.; Jin, Z.; Pei, Y. Hydrogen sulfide promotes flowering in heading Chinese cabbage by S-sulfhydration of BraFLCs. Hortic. Res. 2021, 8, 19. [Google Scholar] [CrossRef]
- Peng, Z.; Liu, G.; Li, H.; Wang, Y.; Gao, H.; Jemric, T.; Fu, D. Molecular and genetic events determining the softening of fleshy fruits: A comprehensive review. Int. J. Mol. Sci. 2022, 23, 12482. [Google Scholar] [CrossRef]
- Brumos, J. Gene regulation in climacteric fruit ripening. Curr. Opin. Plant Biol. 2021, 63, 102042. [Google Scholar] [CrossRef]
- Zhu, L.; Wang, W.; Shi, J.; Zhang, W.; Shen, Y.; Du, H.; Wu, S. Hydrogen sulfide extends the postharvest life and enhances antioxidant activity of kiwifruit during storage. J. Sci. Food Agric. 2014, 94, 2699–2704. [Google Scholar] [CrossRef]
- Ni, Z.J.; Hu, K.D.; Song, C.B.; Ma, R.H.; Li, Z.R.; Zheng, J.L.; Fu, L.H.; Wei, Z.J.; Zhang, H. Hydrogen sulfide alleviates postharvest senescence of grape by modulating the antioxidant defenses. Oxid. Med. Cell. Longev. 2016, 2016, 4715651. [Google Scholar] [CrossRef] [PubMed]
- Mackenzie, S.; McIntosh, L. Higher plant mitochondria. Plant Cell 1999, 11, 571–586. [Google Scholar] [CrossRef] [PubMed]
- Jorge Leon-Morcillo, R.; Angel, J.; Martin, R.; Vierheilig, H.; Antonio Ocampo, J.; Manuel Garcia-Garrido, J. Late activation of the 9-oxylipin pathway during arbuscular mycorrhiza formation in tomato and its regulation by jasmonate signalling. J. Exp. Bot. 2012, 63, 3545–3558. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Liu, D.; Li, P.; Shen, W. Hydrogen sulfide delays leaf yellowing of stored water spinach (Ipomoea aquatica) during dark-induced senescence by delaying chlorophyll breakdown, maintaining energy status and increasing antioxidative capacity. Postharvest Biol. Technol. 2015, 108, 8–20. [Google Scholar] [CrossRef]
- Wang, W.; Ni, Z.J.; Song, C.B.; Ma, W.P.; Cao, S.Q.; Wei, Z.J. Hydrogen sulfide treatment improves quality attributes via regulating the antioxidant system in goji berry (Lycium barbarum L.). Food Chem. 2023, 405, 134858. [Google Scholar] [CrossRef]
- Ali, S.; Nawaz, A.; Ejaz, S.; Haider, S.T.A.; Alam, M.W.; Javed, H.U. Effects of hydrogen sulfide on postharvest physiology of fruits and vegetables: An overview. Sci. Hortic. 2019, 243, 290–299. [Google Scholar] [CrossRef]
- Molinett, S.A.; Alfaro, J.F.; Saez, F.A.; Elgueta, S.; Moya-Leon, M.A.; Figueroa, C.R. Postharvest treatment of hydrogen sulfide delays the softening of chilean strawberry fruit by downregulating the expression of key genes involved in pectin catabolism. Int. J. Mol. Sci. 2021, 22, 10008. [Google Scholar] [CrossRef]
- Marshall, R.S.; Vierstra, R.D. Autophagy: The master of bulk and selective recycling. Annu. Rev. Plant Biol. 2018, 69, 173–208. [Google Scholar] [CrossRef]
- Zhang, Y.M.; Guo, P.; Xia, X.; Guo, H.; Li, Z. Multiple layers of regulation on leaf senescence: New advances and perspectives. Front. Plant Sci. 2021, 12, 788996. [Google Scholar] [CrossRef]
- Jabs, T. Reactive oxygen intermediates as mediators of programmed cell death in plants and animals. Biochem. Pharmacol. 1999, 57, 231–245. [Google Scholar] [CrossRef]
- Daneva, A.; Gao, Z.; Van Durme, M.; Nowack, M.K. Functions and regulation of programmed cell death in plant development. Annu. Rev. Cell Dev. Biol. 2016, 32, 441–468. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, Y.; Kasa, S.; Sakamoto, M.; Aoki, N.; Kai, K.; Yuasa, T.; Hanada, A.; Yamaguchi, S.; Iwaya-Inoue, M. A role for reactive oxygen species produced by NADPH oxidases in the embryo and aleurone cells in barley seed germination. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Aoki, N.; Ishibashi, Y.; Kai, K.; Tomokiyo, R.; Yuasa, T.; Iwaya-Inoue, M. Programmed cell death in barley aleurone cells is not directly stimulated by reactive oxygen species produced in response to gibberellin. J. Plant Physiol. 2014, 171, 615–618. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, F.; Cejudo, F.J. Identification of a nuclear-localized nuclease from wheat cells undergoing programmed cell death that is able to trigger DNA fragmentation and apoptotic morphology on nuclei from human cells. Biochem. J. 2006, 397, 529–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.X.; Hu, K.D.; Lv, K.; Li, Y.H.; Hu, L.Y.; Zhang, X.Q.; Ruan, L.; Liu, Y.S.; Zhang, H. The hydrogen sulfide donor NaHS delays programmed cell death in barley aleurone layers by acting as an antioxidant. Oxid. Med. Cell. Longev. 2015, 2015, 714756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, M.; Zhou, H.; Shen, J.; Zhang, Z.; Gotor, C.; Romero, L.C.; Yuan, X.; Xie, Y. H2S action in plant life cycle. Plant Growth Regul. 2021, 94, 1–9. [Google Scholar] [CrossRef]
- Huo, J.; Huang, D.; Zhang, J.; Fang, H.; Wang, B.; Wang, C.; Liao, W. Hydrogen sulfide: A gaseous molecule in postharvest freshness. Front. Plant Sci. 2018, 9, 1172. [Google Scholar] [CrossRef]
- Ge, Y.; Hu, K.D.; Wang, S.S.; Hu, L.Y.; Chen, X.Y.; Li, Y.H.; Yang, Y.; Yang, F.; Zhang, H. Correction: Hydrogen sulfide alleviates postharvest ripening and senescence of banana by antagonizing the effect of ethylene. PLoS ONE 2018, 13, e0191351. [Google Scholar] [CrossRef]
- Liu, D.M.; Li, J.N.; Li, Z.W.; Pei, Y.X. Hydrogen sulfide inhibits ethylene-induced petiole abscission in tomato (Solanum lycopersicum L.). Hortic. Res. 2020, 7, 14. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, S. Recent advancements in the mechanism of nitric oxide signaling associated with hydrogen sulfide and melatonin crosstalk during ethylene-induced fruit ripening in plants. Nitric Oxide 2019, 82, 25–34. [Google Scholar] [CrossRef]
- Yang, G.; Zhao, K.; Ju, Y.; Mani, S.; Cao, Q.; Puukila, S.; Khaper, N.; Wu, L.; Wang, R. Hydrogen sulfide protects against cellular senescence via S-sulfhydration of Keap1 and activation of Nrf2. Antioxid. Redox Signal. 2013, 18, 1906–1919. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Du, A.; Zhang, S.; Wang, W.; Liang, J.; Peng, F.; Xiao, Y. Regulation of growth in peach roots by exogenous hydrogen sulfide based on RNA-Seq. Plant Physiol. Biochem. 2021, 159, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, J.; Liu, J.; Liu, T.; Xue, S. Hydrogen sulfide (H2S) signaling in plant development and stress responses. aBIOTECH 2021, 2, 32–63. [Google Scholar] [CrossRef]
- Moller, I.M.; Jensen, P.E.; Hansson, A. Oxidative modifications to cellular components in plants. Annu. Rev. Plant Biol. 2007, 58, 459–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittler, R.; Zandalinas, S.I.; Fichman, Y.; Van Breusegem, F. Reactive oxygen species signalling in plant stress responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 663–679. [Google Scholar] [CrossRef]
- Corpas, F.J.; Gonzalez-Gordo, S.; Palma, J.M. Plant peroxisomes: A factory of reactive species. Front. Plant Sci. 2020, 11, 853. [Google Scholar] [CrossRef]
- Corpas, F.J. Peroxisomes in higher plants: An example of metabolic adaptability. Bot. Lett. 2019, 166, 298–308. [Google Scholar] [CrossRef]
- Corpas, F.J.; Barroso, J.B.; Gonzalez-Gordo, S.; Munoz-Vargas, M.A.; Palma, J.M. Hydrogen sulfide: A novel component in Arabidopsis peroxisomes which triggers catalase inhibition. J. Integr. Plant Biol. 2019, 61, 871–883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Mu, X.; Chen, X.; Han, Y. Hydrogen sulfide attenuates intracellular oxidative stress via repressing glycolate oxidase activities in Arabidopsis thaliana. BMC Plant Biol. 2022, 22, 98. [Google Scholar] [CrossRef]
- Aroca, A.; Benito, J.M.; Gotor, C.; Romero, L.C. Persulfidation proteome reveals the regulation of protein function by hydrogen sulfide in diverse biological processes in Arabidopsis. J. Exp. Bot. 2017, 68, 4915–4927. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Tian, M.; Han, Y. Hydrogen sulfide: A multi-tasking signal molecule in the regulation of oxidative stress responses. J. Exp. Bot. 2020, 71, 2862–2869. [Google Scholar] [CrossRef]
- Bhuyan, M.H.M.B.; Hasanuzzaman, M.; Parvin, K.; Mohsin, S.M.; Al Mahmud, J.; Nahar, K.; Fujita, M. Nitric oxide and hydrogen sulfide: Two intimate collaborators regulating plant defense against abiotic stress. Plant Growth Regul. 2020, 90, 409–424. [Google Scholar] [CrossRef]
- Silva, C.J.D.; Batista Fontes, E.P.; Modolo, L.V. Salinity-induced accumulation of endogenous H2S and NO is associated with modulation of the antioxidant and redox defense systems in Nicotiana tabacum L. cv. Havana. Plant Sci. 2017, 256, 148–159. [Google Scholar] [CrossRef] [PubMed]
- Peng, R.; Bian, Z.; Zhou, L.; Cheng, W.; Hai, N.; Yang, C.; Yang, T.; Wang, X.; Wang, C. Hydrogen sulfide enhances nitric oxide-induced tolerance of hypoxia in maize (Zea mays L.). Plant Cell Rep. 2016, 35, 2325–2340. [Google Scholar] [CrossRef]
- Silva, C.J.D.; Mollica, D.C.F.; Vicente, M.H.; Peres, L.E.P.; Modolo, L.V. NO, hydrogen sulfide does not come first during tomato response to high salinity. Nitric Oxide 2018, 76, 164–173. [Google Scholar] [CrossRef]
- Kaya, C.; Ashraf, M.; Alyemeni, M.N.; Ahmad, P. Responses of nitric oxide and hydrogen sulfide in regulating oxidative defence system in wheat plants grown under cadmium stress. Physiol. Plant 2020, 168, 345–360. [Google Scholar] [CrossRef] [PubMed]
- Muneer, S.; Lee, J.H. Hazardous gases (CO, NOx, CH4 and C3H8) released from CO2 fertilizer unit lead to oxidative damage and degrades photosynthesis in strawberry plants. Sci. Rep. 2018, 8, 12291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, N.; Bhatla, S.C. Heme oxygenase-nitric oxide crosstalk-mediated iron homeostasis in plants under oxidative stress. Free. Radic. Biol. Med. 2022, 182, 192–205. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.T.; Li, M.Y.; Cui, W.T.; Lu, W.; Shen, W.B. Haem oxygenase-1 is involved in hydrogen sulfide-induced cucumber adventitious root formation. J. Plant Growth Regul. 2012, 31, 519–528. [Google Scholar] [CrossRef]
- Li, Z.G.; Gu, S.P. Hydrogen sulfide as a signal molecule in hematin-induced heat tolerance of tobacco cell suspension. Biol. Plant. 2016, 60, 595–600. [Google Scholar] [CrossRef]
- Alvarez, M.E.; Savoure, A.; Szabados, L. Proline metabolism as regulatory hub. Trends Plant Sci. 2022, 27, 39–55. [Google Scholar] [CrossRef] [PubMed]
- Patriarca, E.J.; Cermola, F.; D’Aniello, C.; Fico, A.; Guardiola, O.; De Cesare, D.; Minchiotti, G. The multifaceted roles of proline in cell behavior. Front. Cell Dev. Biol. 2021, 9, 728576. [Google Scholar] [CrossRef] [PubMed]
- Wang, X. Regulatory functions of phospholipase D and phosphatidic acid in plant growth, development, and stress responses. Plant Physiol. 2005, 139, 566–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Ding, B.; Guo, Y.; Li, M.; Chen, S.; Huang, G.; Xie, X. Overexpression of a wheat phospholipase D gene, TaPLDα, enhances tolerance to drought and osmotic stress in Arabidopsis thaliana. Planta 2014, 240, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Zhou, Y.P.; Li, H.; Liu, R.R.; Wang, W.; Wu, W.Z.; Yang, N.; Wang, S.Y. Osmotic stress-triggered stomatal closure requires phospholipase δ and hydrogen sulfide in Arabidopsis thaliana. Biochem. Biophy. Res. Commun. 2021, 534, 914–920. [Google Scholar] [CrossRef]
- Zhao, M.; Liu, Q.; Zhang, Y.; Yang, N.; Wu, G.; Li, Q.; Wang, W. Alleviation of osmotic stress by H2S is related to regulated PLD α 1 and suppressed ROS in Arabidopsis thaliana. J. Plant Res. 2020, 133, 393–407. [Google Scholar] [CrossRef]
- Zhang, Y.; Cheng, P.; Wang, Y.; Li, Y.; Su, J.; Chen, Z.; Yu, X.; Shen, W. Genetic elucidation of hydrogen signaling in plant osmotic tolerance and stomatal closure via hydrogen sulfide. Free Radic. Biol. Med. 2020, 161, 1–14. [Google Scholar] [CrossRef]
- Amir, S.B.; Rasheed, R.; Ashraf, M.A.; Hussain, I.; Iqbal, M. Hydrogen sulfide mediates defense response in safflower by regulating secondary metabolism, oxidative defense, and elemental uptake under drought. Physiol. Plant 2021, 172, 795–808. [Google Scholar] [CrossRef]
- Li, L.H.; Yi, H.L.; Li, X.P.; Qi, H.X. Sulfur dioxide enhance drought tolerance of wheat seedlings through H2S signaling. Ecotoxicol. Environ. Saf. 2021, 207, 111248. [Google Scholar] [CrossRef]
- Iqbal, M.J. Role of osmolytes and antioxidant enzymes for drought tolerance in wheat. In Global Wheat Production; Shah, F., Abdul, B., Muhammad, A., Eds.; IntechOpen: Rijeka, Croatia, 2018; pp. 51–65. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Shang, Y.T.; Wang, W.H.; Chen, X.Y.; He, E.M.; Zheng, H.L.; Shangguan, Z.P. Hydrogen sulfide-mediated polyamines and sugar changes are involved in hydrogen sulfide-induced drought tolerance in Spinacia oleracea seedlings. Front. Plant Sci. 2016, 7, 1173. [Google Scholar] [CrossRef]
- Liu, H.; Xue, S. Interplay between hydrogen sulfide and other signaling molecules in the regulation of guard cell signaling and abiotic/biotic stress response. Plant Commun. 2021, 2, 100179. [Google Scholar] [CrossRef] [PubMed]
- Chater, C.; Peng, K.; Movahedi, M.; Dunn, J.A.; Walker, H.J.; Liang, Y.-K.; McLachlan, D.H.; Casson, S.; Isner, J.C.; Wilson, I.; et al. Elevated CO2-induced responses in stomata require ABA and ABA signaling. Curr. Biol. 2015, 25, 2709–2716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauer, H.; Ache, P.; Lautner, S.; Fromm, J.; Hartung, W.; Al-Rasheid, K.A.S.; Sonnewald, S.; Sonnewald, U.; Kneitz, S.; Lachmann, N.; et al. The stomatal response to reduced relative humidity requires guard cell-autonomous ABA synthesis. Curr. Biol. 2013, 23, 53–57. [Google Scholar] [CrossRef] [Green Version]
- Jin, Z.; Xue, S.; Luo, Y.; Tian, B.; Fang, H.; Li, H.; Pei, Y. Hydrogen sulfide interacting with abscisic acid in stomatal regulation responses to drought stress in Arabidopsis. Plant Physiol. Biochem. 2013, 62, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhou, H.; Zhou, M.; Ge, Z.; Zhang, F.; Foyer, C.H.; Yuan, X.; Xie, Y. The coordination of guard-cell autonomous ABA synthesis and DES1 function in situ regulates plant water deficit responses. J. Adv. Res. 2021, 27, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Wang, Z.; Ma, Q.; Sun, L.; Zhang, L.; Liu, Z.; Liu, D.; Hao, X.; Pei, Y. Hydrogen sulfide mediates ion fluxes inducing stomatal closure in response to drought stress in Arabidopsis thaliana. Plant Soil 2017, 419, 141–152. [Google Scholar] [CrossRef]
- Wang, P.T.; Song, C.P. Guard-cell signalling for hydrogen peroxide and abscisic acid. New Phytol. 2008, 178, 703–718. [Google Scholar] [CrossRef]
- Kwak, J.M.; Mori, I.C.; Pei, Z.M.; Leonhardt, N.; Torres, M.A.; Dangl, J.L.; Bloom, R.E.; Bodde, S.; Jones, J.D.G.; Schroeder, J.I. NADPH oxidase AtrbohD and AtrbohF genes function in ROS-dependent ABA signaling in Arabidopsis. EMBO J. 2003, 22, 2623–2633. [Google Scholar] [CrossRef]
- Fujii, H.; Chinnusamy, V.; Rodrigues, A.; Rubio, S.; Antoni, R.; Park, S.Y.; Cutler, S.R.; Sheen, J.; Rodriguez, P.L.; Zhu, J.K. In vitro reconstitution of an abscisic acid signalling pathway. Nature 2009, 462, 660–664. [Google Scholar] [CrossRef] [Green Version]
- Kollist, H.; Jossier, M.; Laanemets, K.; Thomine, S. Anion channels in plant cells. FEBS J. 2011, 278, 4277–4292. [Google Scholar] [CrossRef]
- Wang, L.; Wan, R.; Shi, Y.; Xue, S. Hydrogen sulfide activates S-type anion channel via OST1 and Ca2+ modules. Mol. Plant. 2016, 9, 489–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papanatsiou, M.; Scuffi, D.; Blatt, M.R.; Garcia-Mata, C. Hydrogen sulfide regulates inward-rectifying K+ channels in conjunction with stomatal closure. Plant Physiol. 2015, 168, 29–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carmo-Silva, A.E.; Gore, M.A.; Andrade-Sanchez, P.; French, A.N.; Hunsaker, D.J.; Salvucci, M.E. Decreased CO2 availability and inactivation of Rubisco limit photosynthesis in cotton plants under heat and drought stress in the field. Environ. Exp. Bot. 2012, 83, 1–11. [Google Scholar] [CrossRef]
- Li, H.; Gao, M.Q.; Xue, R.L.; Wang, D.; Zhao, H.J. Effect of hydrogen sulfide on D1 protein in wheat under drought stress. Acta Physiol. Plant. 2015, 37, 225. [Google Scholar] [CrossRef]
- Fischer, S.; Wilckens, R.; Jara, J.; Aranda, M. Variation in antioxidant capacity of quinoa (Chenopodium quinoa Will) subjected to drought stress. Ind. Crops Prod. 2013, 46, 341–349. [Google Scholar] [CrossRef]
- Singh, R.K.; Deshmukh, R.; Muthamilarasan, M.; Rani, R.; Prasad, M. Versatile roles of aquaporin in physiological processes and stress tolerance in plants. Plant Physiol. Biochem. 2020, 149, 178–189. [Google Scholar] [CrossRef] [PubMed]
- Zaffagnini, M.; Bedhomme, M.; Marchand, C.H.; Morisse, S.; Trost, P.; Lemaire, S.D. Redox regulation in photosynthetic organisms: Focus on glutathionylation. Antioxid. Redox Signal. 2012, 16, 567–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.C.; Xu, T.; Li, X.; Tang, J.; Ma, D.F.; Li, Z.Y.; Sun, J. NaCl-induced changes of ion homeostasis and nitrogen metabolism in two sweet potato (Ipomoea batatas L.) cultivars exhibit different salt tolerance at adventitious root stage. Environ. Exp. Bot. 2016, 129, 23–36. [Google Scholar] [CrossRef]
- Chen, J.; Wang, W.H.; Wu, F.H.; He, E.M.; Liu, X.; Shangguan, Z.P.; Zheng, H.L. Hydrogen sulfide enhances salt tolerance through nitric oxide-mediated maintenance of ion homeostasis in barley seedling roots. Sci. Rep. 2015, 5, 12156. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.; Yang, W.; Wen, M.X.; Jin, S.; Liu, Y. Hydrogen sulfide alleviates salinity stress in Cyclocarya paliurus by maintaining chlorophyll fluorescence and regulating nitric oxide level and antioxidant capacity. Plant Physiol. Biochem. 2021, 167, 738–747. [Google Scholar] [CrossRef]
- Jiang, J.L.; Tian, Y.; Li, L.; Yu, M.; Hou, R.P.; Ren, X.M. H2S alleviates salinity stress in cucumber by maintaining the Na+/K+ balance and regulating H2S metabolism and oxidative stress response. Front. Plant Sci. 2019, 10, 678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, G.; Zhou, L.; Wang, Y.; Zhang, G.; Chen, X. Hydrogen sulfide acts downstream of jasmonic acid to inhibit stomatal development in Arabidopsis. Planta 2020, 251, 42. [Google Scholar] [CrossRef]
- Kaya, C.; Higgs, D.; Ashraf, M.; Alyemeni, M.N.; Ahmad, P. Integrative roles of nitric oxide and hydrogen sulfide in melatonin-induced tolerance of pepper (Capsicum annuum L.) plants to iron deficiency and salt stress alone or in combination. Physio. Plant. 2020, 168, 256–277. [Google Scholar] [CrossRef] [Green Version]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Zhu, J.; Dong, C.H.; Zhu, J.K. Interplay between cold-responsive gene regulation, metabolism and RNA processing during plant cold acclimation. Curr. Opin. Plant Biol. 2007, 10, 290–295. [Google Scholar] [CrossRef]
- Gong, Z.; Xiong, L.; Shi, H.; Yang, S.; Herrera-Estrella, L.R.; Xu, G.; Chao, D.-Y.; Li, J.; Wang, P.-Y.; Qin, F.; et al. Plant abiotic stress response and nutrient use efficiency. Sci. China-Life Sci. 2020, 63, 635–674. [Google Scholar] [CrossRef] [PubMed]
- Ruelland, E.; Vaultier, M.-N.; Zachowski, A.; Hurry, V. Cold signalling and cold acclimation in plants. In Advances in Botanical Research, 1st ed.; Kader, J.C., Delseny, M., Eds.; Elsevier Ltd.: Amsterdam, The Netherlands, 2009; Volume 49, pp. 35–150. [Google Scholar]
- Fu, P.; Wang, W.; Hou, L.; Liu, X. Hydrogen sulfide is involved in the chilling stress response in Vitis vinifera L. Acta Soc. Bot. Pol. 2013, 82, 295–302. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.J. Cold and abiotic stress signaling in plants. In Improving Crop Resistance to Abiotic Stress; Tuteja, N., Gill, S.S., Tiburcio, A.F., Tuteja, R., Eds.; Wiley-VCH Verlag GmbH & Co. KgaA: Weinheim, Germany, 2012; pp. 97–132. [Google Scholar]
- Du, X.Z.; Jin, Z.P.; Liu, D.M.; Yang, G.D.; Pei, Y.X. Hydrogen sulfide alleviates the cold stress through MPK4 in Arabidopsis thaliana. Plant Physiol. Biochem. 2017, 120, 112–119. [Google Scholar] [CrossRef]
- Du, X.Z.; Jin, Z.P.; Liu, Z.Q.; Liu, D.M.; Zhang, L.P.; Ma, X.L.; Yang, G.D.; Liu, S.; Guo, Y.R.; Pei, Y.X. H2S persulfidated and increased kinase activity of MPK4 to response cold stress in Arabidopsis. Front. Mol. Biosci. 2021, 8, 635470. [Google Scholar] [CrossRef]
- Janicka, M.; Reda, M.; Czyzewska, K.; Kabala, K. Involvement of signalling molecules NO, H2O2 and H2S in modification of plasma membrane proton pump in cucumber roots subjected to salt or low temperature stress. Funct. Plant Biol. 2018, 45, 428–439. [Google Scholar] [CrossRef]
- Geng, B.; Huang, D.D.; Zhu, S.H. Regulation of hydrogen sulfide metabolism by nitric oxide inhibitors and the quality of peaches during cold storage. Antioxidants 2019, 8, 401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, X.D.; An, B.Y.; Cao, D.M.; Xu, R.; Wang, S.Y.; Zhang, Z.D.; Liu, X.J.; Sun, X.G. Improving photosynthetic capacity, alleviating photosynthetic inhibition and oxidative stress under low temperature stress with exogenous hydrogen sulfide in blueberry seedlings. Front. Plant Sci. 2020, 11, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.J.; Zhang, X.W.; Cai, B.B.; Pan, D.Y.; Fu, X.; Bi, H.G.; Ai, X.Z. Physiological response and transcription profiling analysis reveal the role of glutathione in H2S-induced chilling stress tolerance of cucumber seedlings. Plant Sci. 2020, 291, 110363. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liu, Z.; Li, J.; Luo, S.; Zhang, J.; Xie, J. Hydrogen sulfide interacts with 5-aminolevulinic acid to enhance the antioxidant capacity of pepper (Capsicum annuum L.) seedlings under chilling stress. Agronomy 2022, 12, 572. [Google Scholar] [CrossRef]
- Iqbal, N.; Umar, S.; Khan, N.A.; Corpas, F.J. Nitric oxide and hydrogen sulfide coordinately reduce glucose sensitivity and decrease oxidative stress via ascorbate-glutathione cycle in heat-stressed wheat (Triticum aestivum L.) plants. Antioxidants 2021, 10, 108. [Google Scholar] [CrossRef]
- Li, Z.G. Synergistic effect of antioxidant system and osmolyte in hydrogen sulfide and salicylic acid crosstalk-induced heat tolerance in maize (Zea mays L.) seedlings. Plant Signal. Behav. 2015, 10, e1051278. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.G.; Jin, J.Z. Hydrogen sulfide partly mediates abscisic acid-induced heat tolerance in tobacco (Nicotiana tabacum L.) suspension cultured cells. Plant Cell Tissue Organ Cult. 2015, 125, 207–214. [Google Scholar] [CrossRef]
- Li, Z.G.; Yang, S.Z.; Long, W.B.; Yang, G.X.; Shen, Z.Z. Hydrogen sulphide may be a novel downstream signal molecule in nitric oxide-induced heat tolerance of maize (Zea mays L.) seedlings. Plant Cell Environ. 2013, 36, 1564–1572. [Google Scholar] [CrossRef]
- Cheng, T.L.; Shi, J.S.; Dong, Y.N.; Ma, Y.; Peng, Y.; Hu, X.Y.; Chen, J.H. Hydrogen sulfide enhances poplar tolerance to high-temperature stress by increasing S-nitrosoglutathione reductase (GSNOR) activity and reducing reactive oxygen/nitrogen damage. Plant Growth Regul. 2018, 84, 11–23. [Google Scholar] [CrossRef]
- Christou, A.; Filippou, P.; Manganaris, G.A.; Fotopoulos, V. Sodium hydrosulfide induces systemic thermotolerance to strawberry plants through transcriptional regulation of heat shock proteins and aquaporin. BMC Plant Biol. 2014, 14, 42. [Google Scholar] [CrossRef]
- Rascio, N.; Navari-Izzo, F. Heavy metal hyperaccumulating plants: How and why do they do it? And what makes them so interesting? Plant Sci. 2011, 180, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Dhiman, A. Nickel and cadmium toxicity in plants. J. Pharm. Sci. Innov. 2013, 2, 20–24. [Google Scholar] [CrossRef]
- Zulfiqar, F.; Hancock, J.T. Hydrogen sulfide in horticulture: Emerging roles in the era of climate change. Plant Physiol. Biochem. 2020, 155, 667–675. [Google Scholar] [CrossRef] [PubMed]
- Shivaraj, S.M.; Vats, S.; Bhat, J.A.; Dhakte, P.; Goyal, V.; Khatri, P.; Kumawat, S.; Singh, A.; Prasad, M.; Sonah, H.; et al. Nitric oxide and hydrogen sulfide crosstalk during heavy metal stress in plants. Physio. Plant. 2020, 168, 437–455. [Google Scholar] [CrossRef]
- Singh, S.K.; Suhel, M.; Husain, T.; Prasad, S.M.; Singh, V.P. Hydrogen sulfide manages hexavalent chromium toxicity in wheat and rice seedlings: The role of sulfur assimilation and ascorbate-glutathione cycle. Environ. Pollut. 2022, 307, 119509. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.; Roychoudhury, A. Regulation of physiological aspects in plants by hydrogen sulfide and nitric oxide under challenging environment. Physio. Plant. 2020, 168, 374–393. [Google Scholar] [CrossRef] [Green Version]
- Krzeslowska, M. The cell wall in plant cell response to trace metals: Polysaccharide remodeling and its role in defense strategy. Acta Physiol. Plant. 2011, 33, 35–51. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.Z.; Ling, Q.L.; Li, Y.H.; Lin, Y.J. mRNA analysis of genes encoded with phytochelatin synthase (PCS) in rice seedlings exposed to chromium: The role of phytochelatins in Cr detoxification. Bull. Environ. Contam. Toxicol. 2018, 101, 257–261. [Google Scholar] [CrossRef]
- Jia, H.; Wang, X.; Shi, C.; Guo, J.; Ma, P.; Ren, X.; Wei, T.; Liu, H.; Li, J. Hydrogen sulfide decreases Cd translocation from root to shoot through increasing Cd accumulation in cell wall and decreasing Cd2+ influx in Isatis indigotica. Plant Physiol. Biochem. 2020, 155, 605–612. [Google Scholar] [CrossRef]
- Alamri, S.; Ali, H.M.; Khan, M.I.R.; Singh, V.P.; Siddiqui, M.H. Exogenous nitric oxide requires endogenous hydrogen sulfide to induce the resilience through sulfur assimilation in tomato seedlings under hexavalent chromium toxicity. Plant Physiol. Biochem. 2020, 155, 20–34. [Google Scholar] [CrossRef]
- Qiao, Z.J.; Jing, T.; Jin, Z.P.; Liang, Y.L.; Zhang, L.P.; Liu, Z.Q.; Liu, D.M.; Pei, Y.X. CDPKs enhance Cd tolerance through intensifying H2S signal in Arabidopsis thaliana. Plant Soil 2016, 398, 99–110. [Google Scholar] [CrossRef]
- Zanganeh, R.; Jamei, R.; Rahmani, F. Impacts of seed priming with salicylic acid and sodium hydrosulfide on possible metabolic pathway of two amino acids in maize plant under lead stress. Mol. Biol. Res. Commun. 2018, 7, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, T.B.; Chapman, L.M. The importance of thiamine (vitamin B1) in plant health: From crop yield to biofortification. J. Biol. Chem. 2020, 295, 12002–12013. [Google Scholar] [CrossRef] [PubMed]
- Kaya, C.; Aslan, M. Hydrogen sulphide partly involves in thiamine-induced tolerance to cadmium toxicity in strawberry (Fragaria x ananassa Duch) plants. Environ. Sci. Pollut. Res. 2020, 27, 941–953. [Google Scholar] [CrossRef]
- Filipovic, M.R.; Jovanovic, V.M. More than just an intermediate: Hydrogen sulfide signalling in plants. J. Exp. Bot. 2017, 68, 4733–4736. [Google Scholar] [CrossRef]
- Marti, M.C.; Jimenez, A.; Sevilla, F. Thioredoxin network in plant mitochondria: Cysteine S-posttranslational modifications and stress conditions. Front. Plant Sci. 2020, 11, 571288. [Google Scholar] [CrossRef]
- Sevilla, F.; Camejo, D.; Ortiz-Espin, A.; Calderon, A.; Lazaro, J.J.; Jimenez, A. The thioredoxin/peroxiredoxin/sulfiredoxin system: Current overview on its redox function in plants and regulation by reactive oxygen and nitrogen species. J. Exp. Bot. 2015, 66, 2945–2955. [Google Scholar] [CrossRef] [Green Version]
- Begara-Morales, J.C.; López-Jaramillo, F.J.; Sánchez-Calvo, B.; Carreras, A.; Ortega-Muñoz, M.; Santoyo-González, F.; Corpas, F.J.; Barroso, a.J.B. Vinyl sulfone silica: Application of an open preactivated support to the study of transnitrosylation of plant proteins by S-nitrosoglutathione. BMC Plant Biol. 2013, 13, 61. [Google Scholar] [CrossRef] [Green Version]
- Aroca, A.; Serna, A.; Gotor, C.; Romero, L.C. S-sulfhydration: A cysteine posttranslational modification in plant systems. Plant Physiol. 2015, 168, 334–342. [Google Scholar] [CrossRef] [Green Version]
- Ju, Y.; Fu, M.; Stokes, E.; Wu, L.; Yang, G. H2S-mediated protein S-sulfhydration: A prediction for its formation and regulation. Molecules 2017, 22, 1334. [Google Scholar] [CrossRef]
- Mustafa, A.K.; Gadalla, M.M.; Sen, N.; Kim, S.; Mu, W.; Gazi, S.K.; Barrow, R.K.; Yang, G.; Wang, R.; Snyder, S.H. H2S signals through protein S-sulfhydration. Sci. Signal. 2009, 2, ra72. [Google Scholar] [CrossRef] [Green Version]
- Libiad, M.; Yadav, P.K.; Vitvitsky, V.; Martinov, M.; Banerjee, R. Organization of the human mitochondrial hydrogen sulfide oxidation pathway. J. Biol. Chem. 2014, 289, 30901–30910. [Google Scholar] [CrossRef] [Green Version]
- Aroca, A.; Gotor, C. Hydrogen sulfide: A key role in autophagy regulation from plants to mammalians. Antioxidants 2022, 11, 327. [Google Scholar] [CrossRef]
- Laureano-Marin, A.M.; Moreno, I.; Romero, L.C.; Gotor, C. Negative regulation of autophagy by sulfide is independent of reactive oxygen species. Plant Physiol. 2016, 171, 1378–1391. [Google Scholar] [CrossRef] [Green Version]
- Laureano-Marin, A.M.; Aroca, A.; Perez-Perez, M.E.; Yruela, I.; Jurado-Flores, A.; Moreno, I.; Crespo, J.L.; Romero, L.C.; Gotor, C. Abscisic acid-triggered persulfidation of the Cys protease ATG4 mediates regulation of autophagy by sulfide. Plant Cell 2020, 32, 3902–3920. [Google Scholar] [CrossRef]
- Yoshida, T.; Christmann, A.; Yamaguchi-Shinozaki, K.; Grill, E.; Fernie, A.R. Revisiting the basal role of ABA-roles outside of stress. Trends Plant Sci. 2019, 24, 625–635. [Google Scholar] [CrossRef]
- Munoz-Vargas, M.A.; Gonzalez-Gordo, S.; Canas, A.; Lopez-Jaramillo, J.; Palma, J.M.; Corpas, F.J. Endogenous hydrogen sulfide (H2S) is up-regulated during sweet pepper (Capsicum annuum L.) fruit ripening. In vitro analysis shows that NADP-dependent isocitrate dehydrogenase (ICDH) activity is inhibited by H2S and NO. Nitric Oxide 2018, 81, 36–45. [Google Scholar] [CrossRef]
- Munoz-Vargas, M.A.; Gonzalez-Gordo, S.; Palma, J.M.; Corpas, F.J. Inhibition of NADP-malic enzyme activity by H2S and NO in sweet pepper (Capsicum annuum L.) fruits. Physiol. Plant. 2020, 168, 278–288. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Chen, S.; Wang, X.; Shi, C.; Liu, H.; Yang, J.; Shi, W.; Guo, J.; Jia, H. Hydrogen sulfide disturbs actin polymerization via S-sulfhydration resulting in stunted root hair growth. Plant Physiol. 2018, 178, 936–949. [Google Scholar] [CrossRef] [Green Version]
- Aroca, A.; Zhang, J.; Xie, Y.; Romero, L.C.; Gotor, C. Hydrogen sulfide signaling in plant adaptations to adverse conditions: Molecular mechanisms. J. Exp. Bot. 2021, 72, 5893–5904. [Google Scholar] [CrossRef]
- Vasas, A.; Doka, E.; Fabian, I.; Nagy, P. Kinetic and thermodynamic studies on the disulfide-bond reducing potential of hydrogen sulfide. Nitric Oxide 2015, 46, 93–101. [Google Scholar] [CrossRef] [Green Version]
- Lo Conte, M.; Carroll, K.S. The redox biochemistry of protein sulfenylation and sulfinylation. J. Biol. Chem. 2013, 288, 26480–26488. [Google Scholar] [CrossRef] [Green Version]
- Astier, J.; Lindermayr, C. Nitric oxide-dependent posttranslational modification in plants: An update. Int. J. Mol. Sci. 2012, 13, 15193–15208. [Google Scholar] [CrossRef] [Green Version]
- Leitner, M.; Vandelle, E.; Gaupels, F.; Bellin, D.; Delledonne, M. NO signals in the haze: Nitric oxide signalling in plant defence. Curr. Opin. Plant Biol. 2009, 12, 451–458. [Google Scholar] [CrossRef]
- Fancy, N.N.; Bahlmann, A.K.; Loake, G.J. Nitric oxide function in plant abiotic stress. Plant Cell Environ. 2017, 40, 462–472. [Google Scholar] [CrossRef] [Green Version]
- Li, X.H.; Xue, W.L.; Wang, M.J.; Zhou, Y.; Zhang, C.C.; Sun, C.; Zhu, L.; Liang, K.; Chen, Y.; Tao, B.B.; et al. H2S regulates endothelial nitric oxide synthase protein stability by promoting microRNA-455-3p expression. Sci. Rep. 2017, 7, 44807. [Google Scholar] [CrossRef] [Green Version]
- Altaany, Z.; Ju, Y.; Yang, G.; Wang, R. The coordination of S-sulfhydration, S-nitrosylation, and phosphorylation of endothelial nitric oxide synthase by hydrogen sulfide. Sci. Signal. 2014, 7, ra87. [Google Scholar] [CrossRef]
- Chen, S.; Chang, Y.; Ding, Y. Roles of H2S and NO in regulating the antioxidant system of Vibrio alginolyticus under norfloxacin stress. PeerJ 2021, 9, e12255. [Google Scholar] [CrossRef]
- Bogdandi, V.; Ditroi, T.; Batai, I.Z.; Sandor, Z.; Minnion, M.; Vasas, A.; Galambos, K.; Buglyo, P.; Pinter, E.; Feelisch, M.; et al. Nitrosopersulfide (SSNO-) is a unique cysteine polysulfidating agent with reduction-resistant bioactivity. Antioxid. Redox Signal. 2020, 33, 1277–1294. [Google Scholar] [CrossRef]
- Eberhardt, M.; Dux, M.; Namer, B.; Miljkovic, J.; Cordasic, N.; Will, C.; Kichko, T.I.; de la Roche, J.; Fischer, M.; Suarez, S.A.; et al. H2S and NO cooperatively regulate vascular tone by activating a neuroendocrine HNO-TRPA1-CGRP signalling pathway. Nat. Commun. 2014, 5, 4381. [Google Scholar] [CrossRef] [Green Version]
- Marcolongo, J.P.; Venancio, M.F.; Rocha, W.R.; Doctorovich, F.; Olabe, J.A. NO/H2S “Crosstalk” reactions. The role of thionitrites (SNO-) and perthionitrites (SSNO-). Inorg. Chem. 2019, 58, 14981–14997. [Google Scholar] [CrossRef]
- Michelet, L.; Zaffagnini, M.; Marchand, C.; Collin, V.; Decottignies, P.; Tsan, P.; Lancelin, J.-M.; Trost, P.; Miginiac-Maslow, M.; Noctor, G.; et al. Glutathionylation of chloroplast thioredoxin f is a redox signaling mechanism in plants. Proc. Natl. Acad. Sci. USA 2005, 102, 16478–16483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, W.N.; Yadav, P.K.; Adamec, J.; Banerjee, R. S-glutathionylation enhances human cystathionine β-synthase activity under oxidative stress conditions. Antioxid. Redox Signal. 2015, 22, 350–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parsanathan, R.; Jain, S.K. Hydrogen sulfide increases glutathione biosynthesis, and glucose uptake and utilisation in C2C12 mouse myotubes. Free Radic. Res. 2018, 52, 288–303. [Google Scholar] [CrossRef]
- Niazi, A.K.; Bariat, L.; Riondet, C.; Carapito, C.; Mhamdi, A.; Noctor, G.; Reichheld, J.P. Cytosolic isocitrate dehydrogenase from Arabidopsis thaliana is regulated by glutathionylation. Antioxidants 2019, 8, 16. [Google Scholar] [CrossRef] [Green Version]
- Song, W.; Hu, L.; Ma, Z.; Yang, L.; Li, J. Importance of tyrosine phosphorylation in hormone-regulated plant growth and development. Int. J. Mol. Sci. 2022, 23, 6603. [Google Scholar] [CrossRef] [PubMed]
- Hoang, Q.T.N.; Han, Y.J.; Kim, J.I. Plant phytochromes and their phosphorylation. Int. J. Mol. Sci. 2019, 20, 3450. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Cao, C.; Li, Y.; Yang, G.; Pei, Y. Light regulates hydrogen sulfide signalling during skoto- and photo-morphogenesis in foxtail millet. Funct. Plant Biol. 2019, 46, 916–924. [Google Scholar] [CrossRef]
- Amooaghaie, R.; Zangene-Madar, F.; Enteshari, S. Role of two-sided crosstalk between NO and H2S on improvement of mineral homeostasis and antioxidative defense in Sesamum indicwn under lead stress. Ecotoxicol. Environ. Saf. 2017, 139, 210–218. [Google Scholar] [CrossRef]
- Chen, S.; Wang, X.; Jia, H.; Li, F.; Ma, Y.; Liesche, J.; Liao, M.; Ding, X.; Liu, C.; Chen, Y.; et al. Persulfidation-induced structural change in SnRK2.6 establishes intramolecular interaction between phosphorylation and persulfidation. Mol. Plant. 2021, 14, 1814–1830. [Google Scholar] [CrossRef]
- Swatek, K.N.; Komander, D. Ubiquitin modifications. Cell Res. 2016, 26, 399–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Lu, F.; Yu, X.; Wang, B.; Chen, J.; Lu, F.; Peng, S.; Sun, X.; Yu, M.; Chen, H.; et al. Exogenous H2S promoted USP8 sulfhydration to regulate mitophagy in the hearts of db/db mice. Aging Dis. 2020, 11, 269–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khago, D.; Fucci, I.J.; Byrd, R.A. The role of conformational dynamics in the recognition and regulation of ubiquitination. Molecules 2020, 25, 5933. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.R.M.; Wang, Y.; Zhang, X.; Cheng, H.; Sun, L.; He, S.; Hao, F. Redox components: Key regulators of epigenetic modifications in plants. Int. J. Mol. Sci. 2020, 21, 1419. [Google Scholar] [CrossRef] [Green Version]
- Schiedel, M.; Conway, S.J. Small molecules as tools to study the chemical epigenetics of lysine acetylation. Curr. Opin. Chem. Biol. 2018, 45, 166–178. [Google Scholar] [CrossRef]
- Pandey, R.; Muller, A.; Napoli, C.A.; Selinger, D.A.; Pikaard, C.S.; Richards, E.J.; Bender, J.; Mount, D.W.; Jorgensen, R.A. Analysis of histone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatin modification among multicellular eukaryotes. Nucleic Acids Res. 2002, 30, 5036–5055. [Google Scholar] [CrossRef] [Green Version]
- Ageeva-Kieferle, A.; Rudolf, E.E.; Lindermayr, C. Redox-dependent chromatin remodeling: A new function of nitric oxide as architect of chromatin structure in plants. Front. Plant Sci. 2019, 10, 625. [Google Scholar] [CrossRef] [Green Version]
- Rios, E.C.S.; Szczesny, B.; Soriano, F.G.; Olah, G.; Szabo, C. Hydrogen sulfide attenuates cytokine production through the modulation of chromatin remodeling. Int. J. Mol. Med. 2015, 35, 1741–1746. [Google Scholar] [CrossRef] [Green Version]
- Chi, Z.; Byeon, H.E.; Seo, E.; Nguyen, Q.A.T.; Lee, W.; Jeong, Y.; Choi, J.; Pandey, D.; Berkowitz, D.E.; Kim, J.H.; et al. Histone deacetylase 6 inhibitor tubastatin A attenuates angiotensin II-induced hypertension by preventing cystathionine γ-lyase protein degradation. Pharmacol. Res. 2019, 146, 104281. [Google Scholar] [CrossRef]
- Leucker, T.M.; Nomura, Y.; Kim, J.H.; Bhatta, A.; Wang, V.; Wecker, A.; Jandu, S.; Santhanam, L.; Berkowitz, D.; Romer, L.; et al. Cystathionine γ-lyase protects vascular endothelium: A role for inhibition of histone deacetylase 6. Am. J. Physiol.-Heart Circ. Physiol. 2017, 312, H711–H720. [Google Scholar] [CrossRef]
- Du, C.; Lin, X.; Xu, W.; Zheng, F.; Cai, J.; Yang, J.; Cui, Q.; Tang, C.; Cai, J.; Xu, G.; et al. Sulfhydrated sirtuin-1 increasing its deacetylation activity is an essential epigenetics mechanism of anti-atherogenesis by hydrogen sulfide. Antioxid. Redox Signal. 2019, 30, 184–197. [Google Scholar] [CrossRef] [PubMed]
- Currais, A.; Huang, L.; Goldberg, J.; Petrascheck, M.; Ates, G.; Pinto-Duarte, A.; Shokhirev, M.N.; Schubert, D.; Maher, P. Elevating acetyl-CoA levels reduces aspects of brain aging. eLife 2019, 8, e47866. [Google Scholar] [CrossRef] [PubMed]
- Mews, P.; Egervari, G.; Nativio, R.; Sidoli, S.; Donahue, G.; Lombroso, S.I.; Alexander, D.C.; Riesche, S.L.; Heller, E.A.; Nestler, E.J.; et al. Alcohol metabolism contributes to brain histone acetylation. Nature 2019, 574, 717–721. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Zhang, Y.; Fu, M.; Pei, Y.; Wu, L.; Wang, R.; Yang, G. Cystathionine gamma-lyase/H2S system suppresses hepatic acetyl-CoA accumulation and nonalcoholic fatty liver disease in mice. Life Sci. 2020, 252, 117661. [Google Scholar] [CrossRef]
- Wang, Y.; Yu, R.; Wu, L.; Yang, G. Hydrogen sulfide signaling in regulation of cell behaviors. Nitric Oxide 2020, 103, 9–19. [Google Scholar] [CrossRef]
- Zhu, C.Q.; Wei, Q.; Hu, W.J.; Kong, Y.L.; Xiang, X.J.; Zhang, H.; Cao, X.C.; Zhu, L.F.; Liu, J.; Tian, W.H.; et al. Unearthing the alleviatory mechanisms of hydrogen sulfide in aluminum toxicity in rice. Plant Physiol. Biochem. 2022, 182, 133–144. [Google Scholar] [CrossRef]
- Cao, H.; Liang, Y.; Zhang, L.; Liu, Z.; Liu, D.; Cao, X.; Deng, X.; Jin, Z.; Pei, Y. AtPRMT5-mediated AtLCD methylation improves Cd2+ tolerance via increased H2S production in Arabidopsis. Plant Physiol. 2022, 190, 2637–2650. [Google Scholar] [CrossRef]
- Li, S.S.; Yang, G.D. Hydrogen sulfide maintains mitochondrial DNA replication via demethylation of TFAM. Antioxid. Redox Signal. 2015, 23, 630–642. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Z.; Wang, X.; Feng, J.; Zhu, S. Biological Functions of Hydrogen Sulfide in Plants. Int. J. Mol. Sci. 2022, 23, 15107. https://doi.org/10.3390/ijms232315107
Yang Z, Wang X, Feng J, Zhu S. Biological Functions of Hydrogen Sulfide in Plants. International Journal of Molecular Sciences. 2022; 23(23):15107. https://doi.org/10.3390/ijms232315107
Chicago/Turabian StyleYang, Zhifeng, Xiaoyu Wang, Jianrong Feng, and Shuhua Zhu. 2022. "Biological Functions of Hydrogen Sulfide in Plants" International Journal of Molecular Sciences 23, no. 23: 15107. https://doi.org/10.3390/ijms232315107