
Genome-Wide Characterization and Expression Profiling of the GRAS Gene Family in Salt and Alkali Stresses in Miscanthus sinensis
 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Genome-Wide Identification of GRAS Genes in M. sinensis
2.2. Chromosomal Locations of MsGRAS Genes
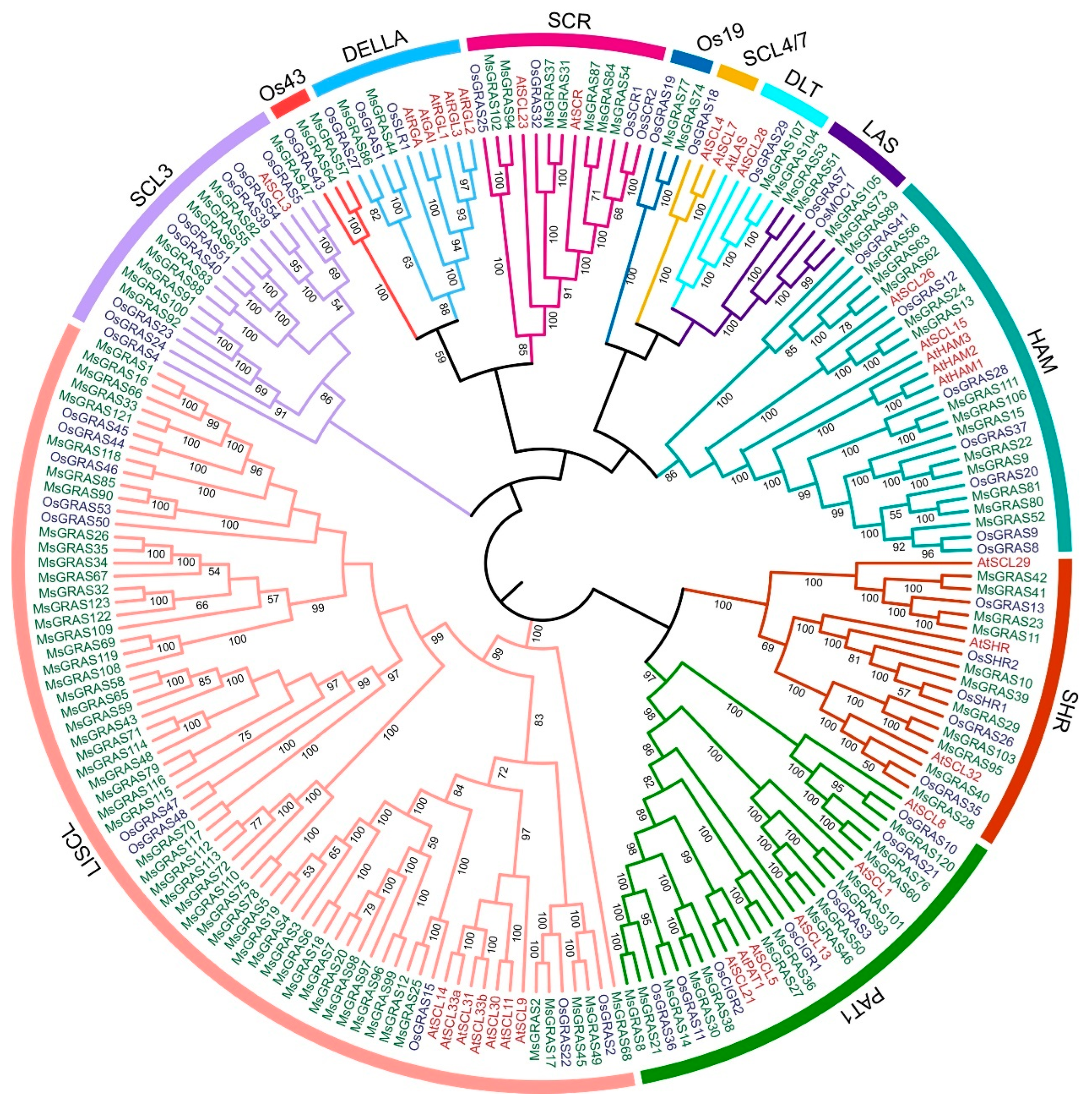
2.3. Phylogenetic Relationships, Gene Structure, and Conserved Motifs of MsGRAS
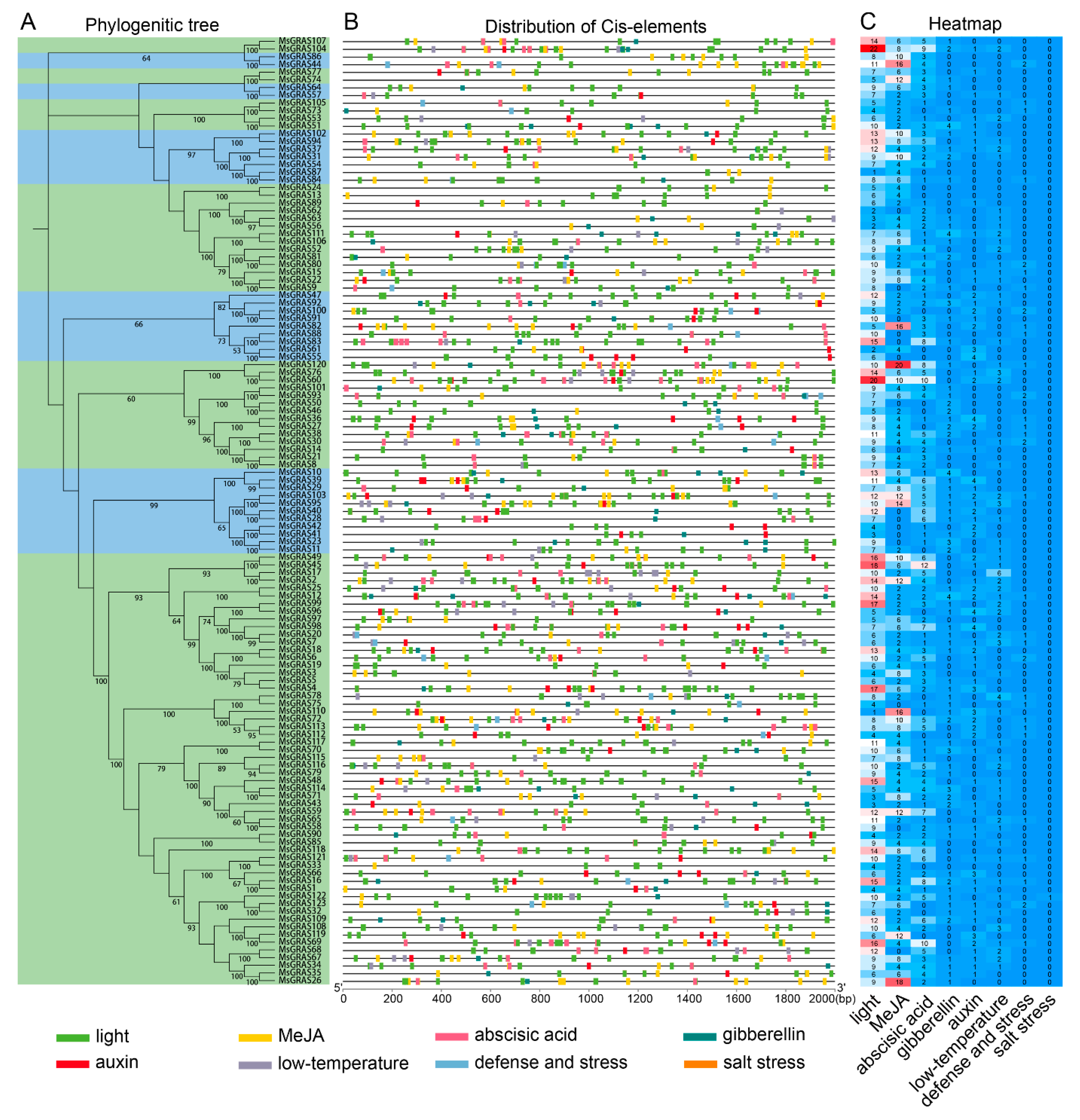
2.4. Cis-Elements in MsGRAS Promoters
2.5. Expression Profiling of MsGRAS Genes in Salt and Alkali Stress
2.6. MsGRAS60 Overexpression Confers Compromised Salt and Alkali Tolerance in Arabidopsis
2.7. MsGRAS60 Overexpression Alleviates ABA Sensitivity in Arabidopsis
3. Discussion
3.1. MsGRAS Gene Duplication and Phylogenetic Relationship
3.2. MsGRAS Protein Structure Characteristic and Function Prediction
3.3. Overexpression of MsGRAS Gene and Abiotic Stress Response
4. Materials and Methods
4.1. The Identification of GRAS Genes in M. sinensis
4.2. Phylogenetic Analysis
4.3. Gene Structure and Conserved Motif Analysis
4.4. Chromosomal Mapping and Segmental Duplication Analysis
4.5. Promoter Cis-Element Analysis
4.6. Plant Materials and Abiotic Stress Treatment
4.7. RNA Isolation and RT-qPCR
4.8. Generation of MsGRAS60 Overexpression Lines in Arabidopsis
4.9. Germination Rate and Fresh Weight Measurements under Salt and Alkali Stresses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Haj Amor, Z.; Araya, T.; Kim, D.G.; Bouri, S.; Lee, J.; Ghilou, W.; Yang, Y.; Kang, H.; Jhariya, M.K.; Banerjee, A.; et al. Soil salinity and its associated effects on soil microorganisms, greenhouse gas emissions, crop yield, biodiversity and desertification: A review. Sci. Total Environ. 2022, 843, 156946. [Google Scholar] [CrossRef] [PubMed]
- FAQ. Global Map of Salt Affected Soils Version 1.0. In 2021. Available online: https://www.fao.org/soils-portal/data-hub/soil-maps-and-databases/global-map-of-salt-affected-soils/en/ (accessed on 28 September 2022).
- Hassani, A.; Azapagic, A.; Shokri, N. Global predictions of primary soil salinization under changing climate in the 21st century. Nat. Commun. 2021, 12, 6663. [Google Scholar] [CrossRef] [PubMed]
- Jesus, J.M.; Danko, A.S.; Fiuza, A.; Borges, M.T. Phytoremediation of salt-affected soils: A review of processes, applicability, and the impact of climate change. Environ. Sci. Pollut. Res. 2015, 22, 6511–6525. [Google Scholar] [CrossRef]
- Shah, S.H.H.; Wang, J.Y.; Hao, X.Y.; Thomas, B. Modelling soil salinity effects on salt water uptake and crop growth using a modified denitrification-decomposition model: A phytoremediation approach. J. Environ. Manag. 2022, 301, 113820. [Google Scholar] [CrossRef] [PubMed]
- Hanin, M.; Ebel, C.; Ngom, M.; Laplaze, L.; Masmoudi, K. New insights on plant salt tolerance mechanisms and their potential use for breeding. Front. Plant Sci. 2016, 7, 1787. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Kong, Y.; Hu, R.; Zhou, G. Miscanthus: A fast-growing crop for environmental remediation and biofuel production. Glob. Chang. Biol. Bioenergy 2021, 13, 58–69. [Google Scholar] [CrossRef]
- Beale, C.V.; Long, S.P. Can perennial C4 grasses attain high efficiencies of radiant energy conversion in cool climates? Plant Cell Environ. 1995, 18, 641–650. [Google Scholar] [CrossRef]
- Wang, D.; Portis, A.R.; Moose, S.P.; Long, S.P. Cool C4 photosynthesis: Pyruvate Pi dikinase expression and activity corresponds to the exceptional cold tolerance of carbon assimilation in Miscanthus × giganteus. Plant Physiol. 2008, 148, 557–567. [Google Scholar] [CrossRef] [Green Version]
- Qadir, M.; Oster, J.D.; Schubert, S.; Noble, A.D.; Sahrawat, K.L. Phytoremediation of sodic and saline-sodic soils. In Advances in Agronomy; Sparks, D.L., Ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2007; Volume 96, pp. 197–247. [Google Scholar]
- Xu, Y.; Zheng, C.; Liang, L.; Yi, Z.; Xue, S. Quantitative assessment of the potential for soil improvement by planting Miscanthus on saline-alkaline soil and the underlying microbial mechanism. Glob. Chang. Biol. Bioenergy 2021, 13, 1191–1205. [Google Scholar] [CrossRef]
- Bevan, M.W.; Uauy, C.; Wulff, B.B.H.; Zhou, J.; Krasileva, K.; Clark, M.D. Genomic innovation for crop improvement. Nature 2017, 543, 346–354. [Google Scholar] [CrossRef]
- Miao, J.; Feng, Q.; Li, Y.; Zhao, Q.; Zhou, C.; Lu, H.; Fan, D.; Yan, J.; Lu, Y.; Tian, Q.; et al. Chromosome-scale assembly and analysis of biomass crop Miscanthus lutarioriparius genome. Nat. Commun. 2021, 12, 2458. [Google Scholar] [CrossRef] [PubMed]
- Mitros, T.; Session, A.M.; James, B.T.; Wu, G.A.; Belaffif, M.B.; Clark, L.V.; Shu, S.; Dong, H.; Barling, A.; Holmes, J.R.; et al. Genome biology of the paleotetraploid perennial biomass crop Miscanthus. Nat. Commun. 2020, 11, 5442. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Ge, C.; Xu, P.; Wang, S.; Cheng, S.; Han, Y.; Wang, Y.; Zhuang, Y.; Hou, X.; Yu, T.; et al. The reference genome of Miscanthus floridulus illuminates the evolution of Saccharinae. Nat. Plants 2021, 7, 608–618. [Google Scholar] [CrossRef] [PubMed]
- Pysh, L.D.; Wysocka-Diller, J.W.; Camilleri, C.; Bouchez, D.; Benfey, P.N. The GRAS gene family in Arabidopsis: Sequence characterization and basic expression analysis of the SCARECROW-LIKE genes. Plant J. 1999, 18, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Tian, C.G.; Wan, P.; Sun, S.H.; Li, J.Y.; Chen, M.S. Genome-wide analysis of the GRAS gene family in rice and Arabidopsis. Plant Mol. Biol. 2004, 54, 519–532. [Google Scholar] [CrossRef] [PubMed]
- Laskar, P.; Bhattacharya, S.; Chaudhuri, A.; Kundu, A. Exploring the GRAS gene family in common bean (Phaseolus vulgaris L.): Characterization, evolutionary relationships, and expression analyses in response to abiotic stresses. Planta 2021, 254, 84. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.P.; Gubler, F. Molecular mechanism of gibberellin signaling in plants. Annu. Rev. Plant Biol. 2004, 55, 197–223. [Google Scholar] [CrossRef] [Green Version]
- Colebrook, E.H.; Thomas, S.G.; Phillips, A.L.; Hedden, P. The role of gibberellin signalling in plant responses to abiotic stress. J. Exp. Biol. 2014, 217, 67–75. [Google Scholar] [CrossRef] [Green Version]
- Augstein, F.; Carlsbecker, A. Salinity induces discontinuous protoxylem via a DELLA-dependent mechanism promoting salt tolerance in Arabidopsis seedlings. New Phytol. 2022, 236, 195–209. [Google Scholar] [CrossRef]
- D’Alessandro, S.; Mizokami, Y.; Legeret, B.; Havaux, M. The apocarotenoid beta-Cyclocitric acid elicits drought tolerance in plants. Iscience 2019, 19, 461–473. [Google Scholar] [CrossRef]
- Fode, B.; Siemsen, T.; Thurow, C.; Weigel, R.; Gatz, C. The Arabidopsis GRAS protein SCL14 interacts with class II TGA transcription factors and is essential for the activation of stress-inducible promoters. Plant Cell 2008, 20, 3122–3135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, K.; Chen, S.; Li, T.; Ma, X.; Liang, X.; Ding, X.; Liu, H.; Luo, L. OsGRAS23, a rice GRAS transcription factor gene, is involved in drought stress response through regulating expression of stress-responsive genes. BMC Plant Biol. 2015, 15, 141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Huang, W.; Xian, Z.; Hu, N.; Lin, D.; Ren, H.; Chen, J.; Su, D.; Li, Z. Overexpression of SIGRAS40 in tomato enhances tolerance to abiotic stresses and influences auxin and gibberellin signaling. Front. Plant Sci. 2017, 8, 1659. [Google Scholar] [CrossRef] [PubMed]
- Habib, S.; Waseem, M.; Li, N.; Yang, L.; Li, Z. Overexpression of SlGRAS7 affects multiple behaviors leading to confer abiotic stresses tolerance and impacts gibberellin and auxin signaling in tomato. Int. J. Genom. 2019, 2019, 4051981. [Google Scholar] [CrossRef] [Green Version]
- Habib, S.; Lwin, Y.Y.; Li, N. Down-regulation of SlGRAS10 in tomato confers abiotic stress tolerance. Genes 2021, 12, 623. [Google Scholar] [CrossRef]
- Ma, H.-S.; Liang, D.; Shuai, P.; Xia, X.-L.; Yin, W.-L. The salt- and drought-inducible poplar GRAS protein SCL7 confers salt and drought tolerance in Arabidopsis thaliana. J. Exp. Bot. 2010, 61, 4011–4019. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Zhang, B.; Su, T.; Li, P.; Xin, X.; Wang, W.; Zhao, X.; Yu, Y.; Zhang, D.; Yu, S.; et al. BrLAS, a GRAS transcription factor from Brassica rapa, is involved in drought stress tolerance in transgenic Arabidopsis. Front. Plant Sci. 2018, 9, 1792. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Yu, T.; Fu, J.; Su, H.; Chen, J.; Zhou, Y.; Chen, M.; Guo, J.; Ma, Y.; Wei, W.; et al. Genome-wide analysis of the GRAS gene family and functional identification of GmGRAS37 in drought and salt tolerance. Front. Plant Sci. 2020, 11, 604690. [Google Scholar] [CrossRef]
- Liu, X.; Widmer, A. Genome-wide comparative analysis of the GRAS gene family in Populus, Arabidopsis and Rice. Plant Mol. Biol. Report. 2014, 32, 1129–1145. [Google Scholar] [CrossRef]
- Xu, W.; Chen, Z.; Ahmed, N.; Han, B.; Cui, Q.; Liu, A. Genome-wide identification, evolutionary analysis, and stress responses of the GRAS gene family in Castor beans. Int. J. Mol. Sci. 2016, 17, 1004. [Google Scholar] [CrossRef]
- Huang, W.; Xian, Z.; Kang, X.; Tang, N.; Li, Z. Genome-wide identification, phylogeny and expression analysis of GRAS gene family in tomato. BMC Plant Biol. 2015, 15, 209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; We, H.; Li, X.; Li, Q.; Zhao, X.; Duan, X.; An, Y.; Lv, W.; An, H. Identification and expression of GRAS family genes in maize (Zea mays L.). PLoS ONE 2017, 12, e0185418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Y.; Yan, J.; Lai, D.; Yang, H.; Xue, G.; He, A.; Guo, T.; Chen, L.; Cheng, X.-B.; Xiang, D.-B.; et al. Genome-wide identification, expression analysis, and functional study of the GRAS transcription factor family and its response to abiotic stress in sorghum Sorghum bicolor (L.) Moench. BMC Genom. 2021, 22, 509. [Google Scholar] [CrossRef]
- Zhang, B.; Liu, J.; Yang, Z.E.; Chen, E.Y.; Zhang, C.J.; Zhang, X.Y.; Li, F.G. Genome-wide analysis of GRAS transcription factor gene family in Gossypium hirsutum L. BMC Genom. 2018, 19, 348. [Google Scholar] [CrossRef] [PubMed]
- Song, X.M.; Liu, T.K.; Duan, W.K.; Ma, Q.H.; Ren, J.; Wang, Z.; Li, Y.; Hou, X.L. Genome-wide analysis of the GRAS gene family in Chinese cabbage (Brassica rapa ssp pekinensis). Genomics 2014, 103, 135–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.; Wang, T.; Xu, Z.; Sun, L.; Zhang, Q. Genome-wide analysis of the GRAS gene family in Prunus mume. Mol. Genet. Genom. 2015, 290, 303–317. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, X.; Wang, X.; Sun, M.; Song, R.; Mao, P.; Jia, S. Genome-wide identification of GRAS gene family and their responses to abiotic stress in Medicago sativa. Int. J. Mol. Sci. 2021, 22, 7729. [Google Scholar] [CrossRef]
- Liu, M.; Sun, W.; Li, C.; Yu, G.; Wang, X. A multilayered cross-species analysis of GRAS transcription factors uncovered their functional networks in plant adaptation to the environment. J. Adv. Res. 2020, 29, 191–205. [Google Scholar] [CrossRef]
- Moore, R.C.; Purugganan, M.D. The early stages of duplicate gene evolution. Proc. Natl. Acad. Sci. USA 2003, 100, 15682–15687. [Google Scholar] [CrossRef]
- Prince, V.E.; Pickett, F.B. Splitting pairs: The diverging fates of duplicated genes. Nat. Rev. Genet. 2002, 3, 827–837. [Google Scholar] [CrossRef] [PubMed]
- Lehti-Shiu, M.D.; Panchy, N.; Wang, P.; Uygun, S.; Shiu, S.-H. Diversity, expansion, and evolutionary novelty of plant DNA-binding transcription factor families. Biochim. Et Biophys. Acta-Gene Regul. Mech. 2017, 1860, 3–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N.; Yang, Y.; Moore, M.J.; Brockington, S.F.; Walker, J.F.; Brown, J.W.; Liang, B.; Feng, T.; Edwards, C.; Mikenas, J.; et al. Evolution of portulacineae marked by gene tree conflict and gene family expansion associated with adaptation to harsh environments. Mol. Biol. Evol. 2019, 36, 112–126. [Google Scholar] [CrossRef] [PubMed]
- Bolle, C. The role of GRAS proteins in plant signal transduction and development. Planta 2004, 218, 683–692. [Google Scholar] [CrossRef]
- Silverstone, A.L.; Ciampaglio, C.N.; Sun, T.P. The Arabidopsis RGA gene encodes a transcriptional regulator repressing the gibberellin signal transduction pathway. Plant Cell 1998, 10, 155–169. [Google Scholar] [CrossRef] [Green Version]
- Itoh, H.; Ueguchi-Tanaka, M.; Sato, Y.; Ashikari, M.; Matsuoka, M. The gibberellin signaling pathway is regulated by the appearance and disappearance of SLENDER RICE1 in nuclei. Plant Cell 2002, 14, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Zhang, J.; Qin, Q.; Yue, J.; Huang, B.; Xu, X.; Yan, L.; Hou, S. The six conserved serine/threonine sites of REPRESSOR OF ga1-3 protein are important for its functionality and stability in gibberellin signaling in Arabidopsis. Planta 2014, 240, 763–779. [Google Scholar] [CrossRef]
- Shan, Z.; Luo, X.; Wu, M.; Wei, L.; Fan, Z.; Zhu, Y. Genome-wide identification and expression of GRAS gene family members in Cassava. BMC Plant Biol. 2020, 20, 46. [Google Scholar] [CrossRef] [Green Version]
- Patthy, L. Intron-dependent evolution-preferred types of exons and introns. Febs Lett. 1987, 214, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Niu, Y.; Zhao, T.; Xu, X.; Li, J. Genome-wide identification and characterization of GRAS transcription factors in tomato (Solanum lycopersicum). PeerJ 2017, 5, e3955. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, N.; Zhu, X.; Yang, J.; Li, S.; Che, Y.; Liu, W.; Si, H. Identification and expression analysis of StGRAS gene family in potato (Solanum tuberosum L.). Comput. Biol. Chem. 2019, 80, 195–205. [Google Scholar] [CrossRef]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. Transcriptional regulatory networks in cellular responses and tolerance to dehydration and cold stresses. Annu. Rev. Plant Biol. 2006, 57, 781–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutta, M.; Saha, A.; Moin, M.; Kirti, P.B. Genome-wide identification, transcript profiling and bioinformatic analyses of GRAS transcription factor genes in Rice. Front. Plant Sci. 2021, 12, 2649. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Zhang, Z. KaKs_calculator 3.0: Calculating selective pressure on coding and non-coding sequences. Genom. Proteom. Bioinform. 2022; in press. [Google Scholar]
- Nie, G.; Yang, Z.; He, J.; Liu, A.; Chen, J.; Wang, S.; Wang, X.; Feng, G.; Li, D.; Peng, Y.; et al. Genome-wide investigation of the NAC transcription factor family in Miscanthus sinensis and expression analysis under various abiotic stress. Front. Plant Sci. 2021, 12, 766550. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, Y.; Du, J.; Pei, S.; Guo, S.; Hao, R.; Wang, D.; Zhou, G.; Li, S.; O’Neill, M.; et al. A DE1 BINDING FACTOR 1-GLABRA2 module regulates rhamnogalacturonan I biosynthesis in Arabidopsis seed coat mucilage. Plant Cell 2022, 34, 1396–1414. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Gene Symbol | Gene Loci | Arabidopsis Ortholog Loci | Arabidopsis Ortholog Symbol | Score | E-Value |
|---|---|---|---|---|---|---|
| 1 | MsGRAS1 | Misin01G020500.1 | AT2G37650.1 | 421 | 1 × 10−136 | |
| 2 | MsGRAS2 | Misin01G025400.1 | AT1G07530.1 | GRAS2/SCL14 | 499 | 7 × 10−162 |
| 3 | MsGRAS3 | Misin01G152800.1 | AT2G37650.1 | 427 | 2 × 10−139 | |
| 4 | MsGRAS4 | Misin01G152900.1 | AT2G37650.1 | 429 | 2 × 10−140 | |
| 5 | MsGRAS5 | Misin01G153100.1 | AT2G37650.1 | 307 | 3 × 10−95 | |
| 6 | MsGRAS6 | Misin01G153200.1 | AT2G29065.1 | 390 | 5 × 10−127 | |
| 7 | MsGRAS7 | Misin01G161800.1 | AT2G37650.1 | 397 | 3 × 10−130 | |
| 8 | MsGRAS8 | Misin01G246900.1 | AT5G48150.1 | PAT1 | 566 | 0 |
| 9 | MsGRAS9 | Misin01G287600.1 | AT4G00150.1 | HAM3/LOM3 | 271 | 7 × 10−83 |
| 10 | MsGRAS10 | Misin01G310300.1 | AT4G37650.1 | SHR/SGR7 | 340 | 5 × 10−110 |
| 11 | MsGRAS11 | Misin01G323700.1 | AT3G13840.1 | 234 | 4 × 10−70 | |
| 12 | MsGRAS12 | Misin01G403600.1 | AT2G37650.1 | 545 | 0 | |
| 13 | MsGRAS13 | Misin01G408100.1 | AT4G08250.1 | 297 | 2 × 10−94 | |
| 14 | MsGRAS14 | Misin01G449800.1 | AT5G48150.1 | PAT1 | 523 | 0 |
| 15 | MsGRAS15 | Misin01G513100.1 | AT4G00150.1 | HAM3/LOM3 | 244 | 4 × 10−74 |
| 16 | MsGRAS16 | Misin02G001900.1 | AT1G07530.1 | GRAS2/SCL14 | 409 | 3 × 10−131 |
| 17 | MsGRAS17 | Misin02G014100.1 | AT1G07530.1 | GRAS2/SCL14 | 496 | 7 × 10−167 |
| 18 | MsGRAS18 | Misin02G151300.1 | AT2G29060.1 | 324 | 6 × 10−102 | |
| 19 | MsGRAS19 | Misin02G151700.1 | AT2G37650.1 | 424 | 2 × 10−138 | |
| 20 | MsGRAS20 | Misin02G158600.1 | AT2G37650.1 | 457 | 7 × 10−153 | |
| 21 | MsGRAS21 | Misin02G234500.1 | AT5G48150.1 | PAT1 | 576 | 0 |
| 22 | MsGRAS22 | Misin02G277200.1 | AT4G00150.1 | HAM3/LOM3 | 258 | 1 × 10−77 |
| 23 | MsGRAS23 | Misin02G312300.1 | AT3G13840.1 | 239 | 3 × 10−72 | |
| 24 | MsGRAS24 | Misin02G400500.1 | AT4G08250.1 | 295 | 1 × 10−93 | |
| 25 | MsGRAS25 | Misin02G410600.1 | AT2G37650.1 | 542 | 0 | |
| 26 | MsGRAS26 | Misin03G277400.1 | AT1G07530.1 | GRAS2/SCL14 | 423 | 1 × 10−137 |
| 27 | MsGRAS27 | Misin03G288500.1 | AT5G48150.1 | PAT1 | 519 | 0 |
| 28 | MsGRAS28 | Misin03G304000.1 | AT3G49950.1 | 422 | 2 × 10−145 | |
| 29 | MsGRAS29 | Misin03G306000.1 | AT4G37650.1 | SHR/SGR7/ | 410 | 2 × 10−137 |
| 30 | MsGRAS30 | Misin03G309100.1 | AT1G50600.1 | SCL5 | 525 | 0 |
| 31 | MsGRAS31 | Misin03G322400.1 | AT5G41920.1 | AtSCL23/SCL23 | 428 | 3 × 10−148 |
| 32 | MsGRAS32 | Misin04G284300.1 | AT1G07530.1 | GRAS2/SCL14 | 421 | 4 × 10−137 |
| 33 | MsGRAS33 | Misin04G284400.1 | AT2G37650.1 | 442 | 4 × 10−148 | |
| 34 | MsGRAS34 | Misin04G299900.1 | AT2G37650.1 | 442 | 3 × 10−145 | |
| 35 | MsGRAS35 | Misin04G300000.1 | AT2G37650.1 | 357 | 1 × 10−113 | |
| 36 | MsGRAS36 | Misin04G313500.1 | AT5G48150.1 | PAT1 | 517 | 4 × 10−180 |
| 37 | MsGRAS37 | Misin04G325100.1 | AT5G41920.1 | SCL23 | 432 | 2 × 10−149 |
| 38 | MsGRAS38 | Misin04G335700.1 | AT5G48150.1 | PAT1 | 524 | 0 |
| 39 | MsGRAS39 | Misin04G338800.1 | AT4G37650.1 | SHR/SGR7 | 393 | 1 × 10−133 |
| 40 | MsGRAS40 | Misin04G340300.1 | AT3G49950.1 | 427 | 2 × 10−147 | |
| 41 | MsGRAS41 | Misin04G350600.1 | AT3G13840.1 | 253 | 2 × 10−77 | |
| 42 | MsGRAS42 | Misin04G351200.1 | AT3G13840.1 | 262 | 9 × 10−81 | |
| 43 | MsGRAS43 | Misin05G054400.1 | AT2G37650.1 | 298 | 2 × 10−94 | |
| 44 | MsGRAS44 | Misin05G234500.1 | AT1G14920.1 | RGA2/GAI | 307 | 1 × 10−98 |
| 45 | MsGRAS45 | Misin05G342400.1 | AT2G37650.1 | 525 | 2 × 10−176 | |
| 46 | MsGRAS46 | Misin05G352700.1 | AT1G21450.1 | SCL1 | 556 | 0 |
| 47 | MsGRAS47 | Misin05G394400.1 | AT1G50420.1 | SCL3 | 378 | 3 × 10−127 |
| 48 | MsGRAS48 | Misin05G411500.1 | AT2G37650.1 | 201 | 8 × 10−59 | |
| 49 | MsGRAS49 | Misin06G319300.1 | AT2G37650.1 | 527 | 2 × 10−177 | |
| 50 | MsGRAS50 | Misin06G347800.1 | AT1G21450.1 | SCL1 | 550 | 0 |
| 51 | MsGRAS51 | Misin07G079700.1 | AT1G55580.1 | SCL18/LAS | 245 | 2 × 10−76 |
| 52 | MsGRAS52 | Misin07G432500.1 | AT4G00150.1 | HAM3/LOM3 | 261 | 7 × 10−81 |
| 53 | MsGRAS53 | Misin08G098200.1 | AT1G55580.1 | SCL18/LAS | 228 | 5 × 10−70 |
| 54 | MsGRAS54 | Misin09G001300.1 | AT3G54220.1 | SGR1/SCR | 553 | 0 |
| 55 | MsGRAS55 | Misin09G017500.1 | AT1G50420.1 | SCL3 | 264 | 1 × 10−82 |
| 56 | MsGRAS56 | Misin09G032300.1 | AT5G41920.1 | SCL23 | 130 | 4 × 10−32 |
| 57 | MsGRAS57 | Misin09G129300.1 | AT1G66350.1 | RGL1 | 312 | 2 × 10−97 |
| 58 | MsGRAS58 | Misin09G205000.1 | AT2G37650.1 | 333 | 5 × 10−105 | |
| 59 | MsGRAS59 | Misin09G205200.1 | AT2G29060.1 | 268 | 2 × 10−81 | |
| 60 | MsGRAS60 | Misin10G000200.1 | AT5G52510.1 | SCL8 | 249 | 7 × 10−74 |
| 61 | MsGRAS61 | Misin10G015400.1 | AT1G50420.1 | SCL3 | 254 | 1 × 10−78 |
| 62 | MsGRAS62 | Misin10G037000.1 | AT4G08250.1 | 115 | 4 × 10−28 | |
| 63 | MsGRAS63 | Misin10G037100.1 | AT5G41920.1 | SCL23 | 152 | 6 × 10−41 |
| 64 | MsGRAS64 | Misin10G088000.1 | AT1G66350.1 | RGL1/RGL | 307 | 1 × 10−95 |
| 65 | MsGRAS65 | Misin10G180800.1 | AT2G37650.1 | 317 | 7 × 10−99 | |
| 66 | MsGRAS66 | Misin10G200900.1 | AT2G37650.1 | 436 | 6 × 10−144 | |
| 67 | MsGRAS67 | Misin10G201300.1 | AT2G29060.1 | 430 | 1 × 10−142 | |
| 68 | MsGRAS68 | Misin10G201500.1 | AT2G37650.1 | 130 | 2.68 × 10−31 | |
| 69 | MsGRAS69 | Misin10G201800.1 | AT1G07530.1 | GRAS2/SCL14 | 434 | 3.88 × 10−142 |
| 70 | MsGRAS70 | Misin10G202000.1 | AT1G07530.1 | GRAS2/SCL14 | 399 | 3.77 × 10−130 |
| 71 | MsGRAS71 | Misin10G202100.1 | AT1G07530.1 | GRAS2/SCL14 | 303 | 1.14 × 10−93 |
| 72 | MsGRAS72 | Misin10G202300.1 | AT2G37650.1 | 242 | 4.29 × 10−74 | |
| 73 | MsGRAS73 | Misin11G037100.1 | AT1G55580.1 | SCL18/LAS | 225 | 2.42 × 10−68 |
| 74 | MsGRAS74 | Misin11G080600.1 | AT5G41920.1 | SCL23 | 120 | 9.17 × 10−30 |
| 75 | MsGRAS75 | Misin11G092500.1 | AT2G29065.1 | 333 | 6.82 × 10−106 | |
| 76 | MsGRAS76 | Misin11G188000.1 | AT5G52510.1 | SCL8 | 261 | 3 × 10−78 |
| 77 | MsGRAS77 | Misin12G082800.1 | AT3G03450.1 | RGL2 | 129 | 3.96 × 10−32 |
| 78 | MsGRAS78 | Misin12G093700.1 | AT2G37650.1 | 328 | 1.11 × 10−103 | |
| 79 | MsGRAS79 | Misin12G093800.1 | AT2G37650.1 | 335 | 6.9 × 10−105 | |
| 80 | MsGRAS80 | Misin12G179500.1 | AT4G00150.1 | HAM3/LOM3 | 286 | 5.04 × 10−88 |
| 81 | MsGRAS81 | Misin12G181200.1 | AT4G00150.1 | HAM3/LOM3 | 286 | 3.27 × 10−87 |
| 82 | MsGRAS82 | Misin14G012600.1 | AT1G50420.1 | SCL3 | 305 | 1 × 10−97 |
| 83 | MsGRAS83 | Misin14G013500.1 | AT1G50420.1 | SCL3 | 270 | 1.33 × 10−85 |
| 84 | MsGRAS84 | Misin14G050000.1 | AT3G54220.1 | SCR/SGR1 | 524 | 6.24 × 10−179 |
| 85 | MsGRAS85 | Misin14G112500.1 | AT2G37650.1 | 459 | 1.56 × 10−152 | |
| 86 | MsGRAS86 | Misin14G164000.1 | AT2G01570.1 | RGA/RGA1/RGA24 | 246 | 9.53 × 10−75 |
| 87 | MsGRAS87 | Misin15G009500.1 | AT3G54220.1 | SCR/SGR1 | 553 | 0 |
| 88 | MsGRAS88 | Misin15G025900.1 | AT1G50420.1 | SCL3 | 266 | 6.14 × 10−84 |
| 89 | MsGRAS89 | Misin15G070400.1 | AT4G08250.1 | 116 | 6.37 × 10−28 | |
| 90 | MsGRAS90 | Misin15G148100.1 | AT2G37650.1 | 461 | 4.96 × 10−153 | |
| 91 | MsGRAS91 | Misin16G120300.1 | AT1G50420.1 | SCL3 | 164 | 6.3 × 10−45 |
| 92 | MsGRAS92 | Misin16G120400.1 | AT1G50420.1 | SCL3 | 223 | 9.28 × 10−68 |
| 93 | MsGRAS93 | Misin16G145600.1 | AT1G21450.1 | SCL1 | 523 | 0 |
| 94 | MsGRAS94 | Misin16G178000.1 | AT3G54220.1 | SCR/SGR1 | 269 | 7.51 × 10−83 |
| 95 | MsGRAS95 | Misin16G190800.1 | AT3G49950.1 | 293 | 8.04 × 10−96 | |
| 96 | MsGRAS96 | Misin17G020200.1 | AT2G37650.1 | 350 | 1.88 × 10−115 | |
| 97 | MsGRAS97 | Misin17G020300.1 | AT2G37650.1 | 479 | 3.77 × 10−161 | |
| 98 | MsGRAS98 | Misin17G020400.1 | AT2G37650.1 | 429 | 2.77 × 10−142 | |
| 99 | MsGRAS99 | Misin17G049900.1 | AT2G37650.1 | 409 | 1.46 × 10−132 | |
| 100 | MsGRAS100 | Misin17G120700.1 | AT1G50420.1 | SCL3 | 130 | 4.04 × 10−33 |
| 101 | MsGRAS101 | Misin17G146900.1 | AT1G21450.1 | SCL1 | 541 | 0 |
| 102 | MsGRAS102 | Misin17G169300.1 | AT3G54220.1 | SCR/SGR1 | 274 | 9.87 × 10−85 |
| 103 | MsGRAS103 | Misin17G191000.1 | AT3G49950.1 | 297 | 2.89 × 10−97 | |
| 104 | MsGRAS104 | Misin18G011200.1 | AT1G63100.1 | SCL28 | 374 | 1.49 × 10−121 |
| 105 | MsGRAS105 | Misin18G187000.1 | AT1G55580.1 | SCL18/LAS | 225 | 2.17 × 10−68 |
| 106 | MsGRAS106 | Misin19G008400.1 | AT4G00150.1 | HAM3/LOM3 | 242 | 2.74 × 10−73 |
| 107 | MsGRAS107 | Misin19G015600.1 | AT1G63100.1 | SCL28 | 375 | 1.19 × 10−121 |
| 108 | MsGRAS108 | MisinT086900.1 | AT4G00150.1 | HAM3/LOM3 | 233 | 1.74 × 10−69 |
| 109 | MsGRAS109 | MisinT072400.1 | AT1G07530.1 | GRAS2/SCL14 | 430 | 3.89 × 10−140 |
| 110 | MsGRAS110 | MisinT072600.1 | AT2G37650.1 | 323 | 2.94 × 10−101 | |
| 111 | MsGRAS111 | MisinT074500.1 | AT2G37650.1 | 393 | 2.46 × 10−127 | |
| 112 | MsGRAS112 | MisinT131300.1 | AT2G37650.1 | 375 | 9.91 × 10−121 | |
| 113 | MsGRAS113 | MisinT131400.1 | AT2G29065.1 | 305 | 5.61 × 10−95 | |
| 114 | MsGRAS114 | MisinT131700.1 | AT1G07530.1 | GRAS2/SCL14 | 310 | 7.58 × 10−96 |
| 115 | MsGRAS115 | MisinT131900.1 | AT2G29060.1 | 347 | 2.12 × 10−111 | |
| 116 | MsGRAS116 | MisinT132000.1 | AT2G37650.1 | 328 | 1.36 × 10−101 | |
| 117 | MsGRAS117 | MisinT132100.1 | AT1G07530.1 | GRAS2/SCL14 | 400 | 1.09 × 10−130 |
| 118 | MsGRAS118 | MisinT132200.1 | AT2G37650.1 | 294 | 3.81 × 10−90 | |
| 119 | MsGRAS119 | MisinT132300.1 | AT1G07530.1 | GRAS2/SCL14 | 342 | 2.5 × 10−108 |
| 120 | MsGRAS120 | MisinT322700.1 | AT5G52510.1 | SCL8 | 258 | 4.98 × 10−77 |
| 121 | MsGRAS121 | MisinT390500.1 | AT1G07530.1 | GRAS2/SCL14 | 453 | 4.6 × 10−148 |
| 122 | MsGRAS122 | MisinT390800.1 | AT2G37650.1 | 446 | 8.22 × 10−148 | |
| 123 | MsGRAS123 | MisinT390900.1 | AT1G07530.1 | GRAS2/SCL14 | 437 | 2.07 × 10−143 |
| No. | Locus 1 | Locus 2 | Protein Identity (%) | Ka | Ks | Ka/Ks | Duplication Type | Purify Selection | Divergence Time (Mya) |
|---|---|---|---|---|---|---|---|---|---|
| 1 | MsGRAS1 | MsGRAS16 | 93.39 | 0.035 | 0.089 | 0.392 | Segmental | Yes | 2.12 |
| 2 | MsGRAS2 | MsGRAS17 | 97.39 | 0.013 | 0.047 | 0.280 | Segmental | Yes | 1.12 |
| 3 | MsGRAS3 | MsGRAS18 | 71.10 | 0.169 | 0.288 | 0.588 | Segmental | Yes | 6.85 |
| 4 | MsGRAS9 | MsGRAS15 | 59.68 | 0.272 | 2.855 | 0.095 | Segmental | Yes | 67.97 |
| 5 | MsGRAS9 | MsGRAS22 | 95.02 | 0.019 | 0.083 | 0.234 | Segmental | Yes | 1.97 |
| 6 | MsGRAS10 | MsGRAS39 | 85.31 | 0.075 | 1.661 | 0.045 | Segmental | Yes | 39.54 |
| 7 | MsGRAS11 | MsGRAS23 | 95.60 | 0.012 | 0.161 | 0.074 | Segmental | Yes | 3.84 |
| 8 | MsGRAS11 | MsGRAS42 | 61.64 | 0.231 | 2.591 | 0.089 | Segmental | Yes | 61.68 |
| 9 | MsGRAS12 | MsGRAS25 | 98.09 | 0.010 | 0.028 | 0.341 | Segmental | Yes | 0.67 |
| 10 | MsGRAS13 | MsGRAS24 | 97.16 | 0.010 | 0.071 | 0.136 | Segmental | Yes | 1.68 |
| 11 | MsGRAS15 | MsGRAS22 | 65.69 | 0.275 | 3.108 | 0.089 | Segmental | Yes | 73.99 |
| 12 | MsGRAS23 | MsGRAS41 | 62.29 | 0.231 | 3.089 | 0.075 | Segmental | Yes | 73.56 |
| 13 | MsGRAS27 | MsGRAS36 | 97.72 | 0.011 | 0.061 | 0.176 | Segmental | Yes | 1.45 |
| 14 | MsGRAS28 | MsGRAS40 | 97.82 | 0.006 | 0.230 | 0.026 | Segmental | Yes | 5.47 |
| 15 | MsGRAS29 | MsGRAS39 | 98.44 | 0.005 | 0.110 | 0.049 | Segmental | Yes | 2.61 |
| 16 | MsGRAS30 | MsGRAS38 | 95.73 | 0.022 | 0.109 | 0.200 | Segmental | Yes | 2.59 |
| 17 | MsGRAS31 | MsGRAS37 | 96.72 | 0.011 | 0.147 | 0.076 | Segmental | Yes | 3.49 |
| 18 | MsGRAS45 | MsGRAS49 | 98.25 | 0.010 | 0.028 | 0.360 | Segmental | Yes | 0.67 |
| 19 | MsGRAS46 | MsGRAS50 | 97.19 | 0.013 | 0.041 | 0.310 | Segmental | Yes | 0.97 |
| 20 | MsGRAS46 | MsGRAS93 | 73.04 | 0.158 | 1.327 | 0.119 | Segmental | Yes | 31.60 |
| 21 | MsGRAS50 | MsGRAS93 | 73.04 | 0.156 | 1.350 | 0.116 | Segmental | Yes | 32.14 |
| 22 | MsGRAS50 | MsGRAS101 | 72.31 | 0.158 | 1.383 | 0.114 | Segmental | Yes | 32.92 |
| 23 | MsGRAS51 | MsGRAS53 | 93.48 | 0.023 | 0.234 | 0.098 | Segmental | Yes | 5.57 |
| 24 | MsGRAS51 | MsGRAS105 | 63.73 | 0.200 | 4.361 | 0.046 | Segmental | Yes | 103.83 |
| 25 | MsGRAS52 | MsGRAS80 | 67.61 | 0.220 | 1.172 | 0.187 | Segmental | Yes | 27.91 |
| 26 | MsGRAS52 | MsGRAS81 | 77.60 | 0.225 | 1.152 | 0.195 | Segmental | Yes | 27.42 |
| 27 | MsGRAS57 | MsGRAS64 | 95.31 | 0.016 | 0.164 | 0.095 | Segmental | Yes | 3.92 |
| 28 | MsGRAS74 | MsGRAS77 | 91.53 | 0.038 | 0.143 | 0.264 | Segmental | Yes | 3.40 |
| 29 | MsGRAS75 | MsGRAS78 | 84.44 | 0.073 | 0.221 | 0.332 | Segmental | Yes | 5.26 |
| 30 | MsGRAS80 | MsGRAS81 | 97.61 | 0.011 | 0.060 | 0.185 | Segmental | Yes | 1.44 |
| 31 | MsGRAS91 | MsGRAS100 | 88.21 | 0.053 | 0.140 | 0.379 | Segmental | Yes | 3.32 |
| 32 | MsGRAS93 | MsGRAS101 | 96.80 | 0.015 | 0.058 | 0.266 | Segmental | Yes | 1.38 |
| 33 | MsGRAS94 | MsGRAS102 | 97.59 | 0.009 | 0.048 | 0.195 | Segmental | Yes | 1.15 |
| 34 | MsGRAS95 | MsGRAS103 | 93.87 | 0.018 | 0.181 | 0.098 | Segmental | Yes | 4.30 |
| 35 | MsGRAS104 | MsGRAS107 | 97.27 | 0.010 | 0.049 | 0.199 | Segmental | Yes | 1.16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, X.; Xu, Y.; He, G.; He, K.; Xiao, L.; Hu, R.; Li, S. Genome-Wide Characterization and Expression Profiling of the GRAS Gene Family in Salt and Alkali Stresses in Miscanthus sinensis. Int. J. Mol. Sci. 2022, 23, 14521. https://doi.org/10.3390/ijms232314521
Zhao X, Xu Y, He G, He K, Xiao L, Hu R, Li S. Genome-Wide Characterization and Expression Profiling of the GRAS Gene Family in Salt and Alkali Stresses in Miscanthus sinensis. International Journal of Molecular Sciences. 2022; 23(23):14521. https://doi.org/10.3390/ijms232314521
Chicago/Turabian StyleZhao, Xuhong, Yan Xu, Guo He, Kang He, Liang Xiao, Ruibo Hu, and Shengjun Li. 2022. "Genome-Wide Characterization and Expression Profiling of the GRAS Gene Family in Salt and Alkali Stresses in Miscanthus sinensis" International Journal of Molecular Sciences 23, no. 23: 14521. https://doi.org/10.3390/ijms232314521