
Effect of Bladder Injection of OnabotulinumtoxinA on the Central Expression of Genes Associated with the Control of the Lower Urinary Tract: A Study in Normal Rats
 , , and
, , and
Abstract
:1. Introduction
2. Results
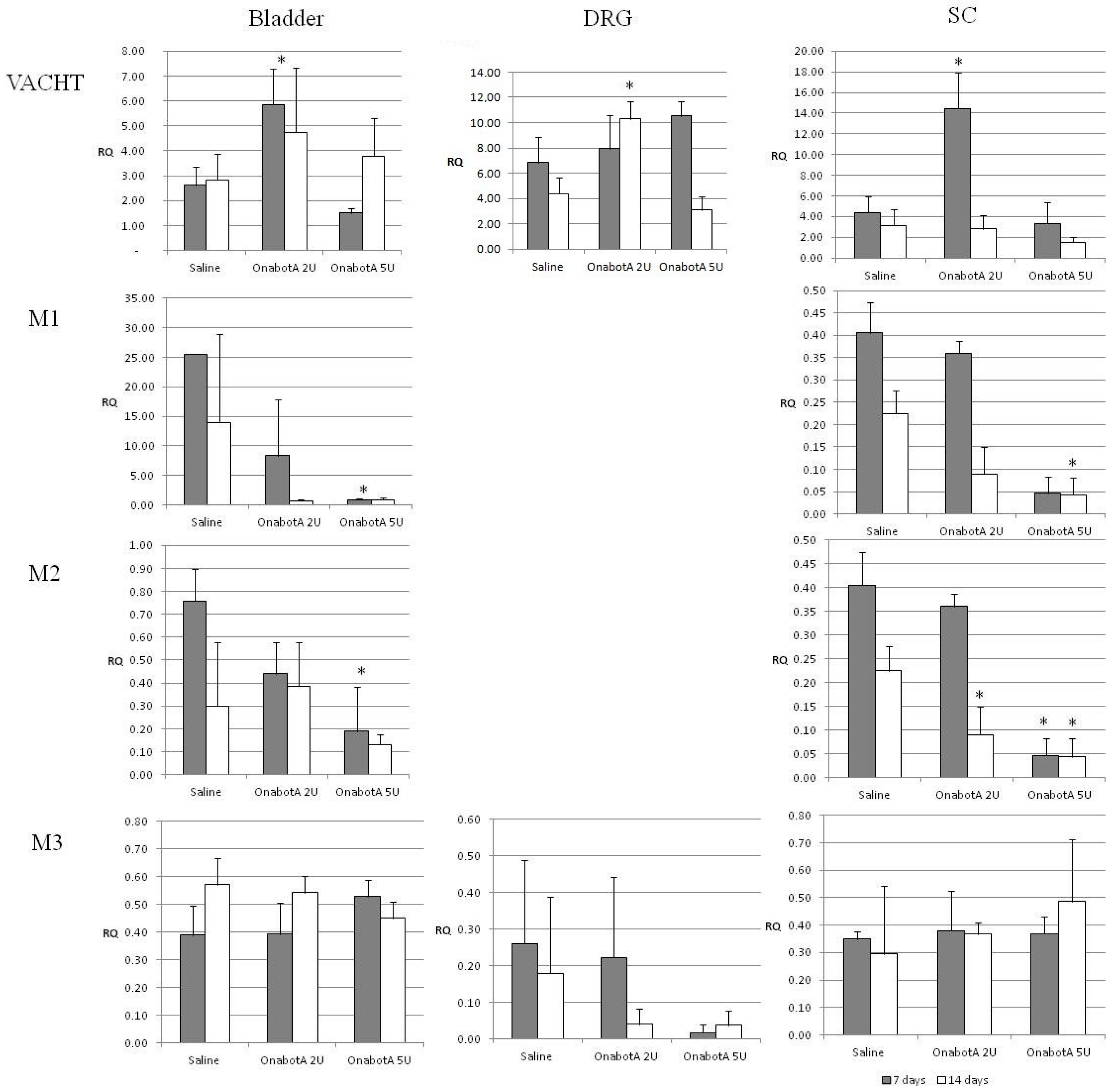
2.1. Real-Time PCR Gene Expression Results
2.2. Protein Expression Results
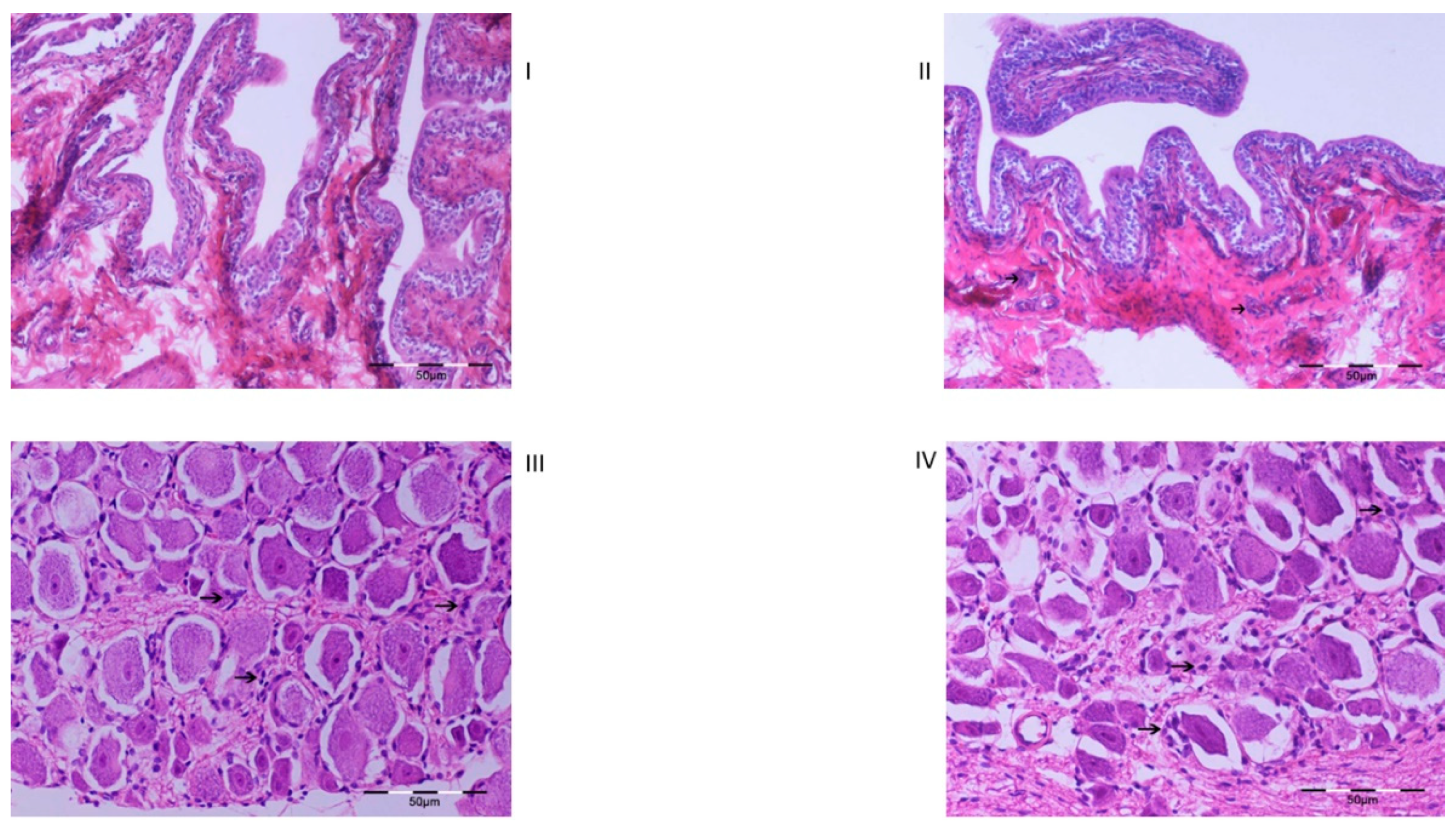
2.3. Histology
3. Discussion
4. Materials & Methods
4.1. Animal Experiments
4.2. Real-Time Polymerase Chain Reaction
4.3. Protein Extraction—Immunoblotting
4.4. Histology
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Apostolidis, A.; Dasgupta, P.; Denys, P.; Elneil, S.; Fowler, C.J.; Giannantoni, A.; Karsenty, G.; Schulte-Baukloh, H.; Schurch, B.; Wyndaele, J.-J.; et al. Recommendations on the use of botulinum toxin in the treatment of lower urinary tract disorders and pelvic floor dysfunctions: A European consensus panel report. Eur. Urol. 2009, 55, 100–120. [Google Scholar] [CrossRef] [PubMed]
- Dolly, O. Synaptic Transmission: Inhibition of Neurotransmitter Release by Botulinum Toxins. Headache: J. Head Face Pain 2003, 43, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Coelho, A.; Dinis, P.; Pinto, R.; Gorgal, T.; Silva, C.; Silva, A.; Silva, J.; Cruz, C.D.; Cruz, F.; Avelino, A. Distribution of the High-Affinity Binding Site and Intracellular Target of Botulinum Toxin Type A in the Human Bladder. Eur. Urol. 2010, 57, 884–890. [Google Scholar] [CrossRef]
- Smith, C.P.; Franks, M.E.; McNeil, B.K.; Ghosh, R.; De Groat, W.C.; Chancellor, M.B.; Somogyi, G.T. Effect of Botulinum Toxin A on the Autonomic Nervous System of the Rat Lower Urinary Tract. J. Urol. 2003, 169, 1896–1900. [Google Scholar] [CrossRef] [PubMed]
- Schulte-Baukloh, H.; Priefert, J.; Knispel, H.H.; Lawrence, G.W.; Miller, K.; Neuhaus, J. Botulinum Toxin A Detrusor Injections Reduce Postsynaptic Muscular M2, M3, P2X2, and P2X3 Receptors in Children and Adolescents Who Have Neurogenic Detrusor Overactivity: A Single-blind Study. Urology 2013, 81, 1052–1057. [Google Scholar] [CrossRef]
- Apostolidis, A.; Dasgupta, P.; Fowler, C.J. Proposed Mechanism for the Efficacy of Injected Botulinum Toxin in the Treatment of Human Detrusor Overactivity. Eur. Urol. 2006, 49, 644–650. [Google Scholar] [CrossRef]
- Howles, S.; Curry, J.; MacKay, I.; Raynard, J.; Brading, A.F. Lack of effectiveness of botulinum toxin on isolated detrusor strips and whole bladders from small animals in vitro. BJU Int. 2009, 104, 1524–1530. [Google Scholar] [CrossRef]
- Roosen, A.; Datta, S.N.; Chowdhury, R.A.; Patel, P.M.; Kalsi, V.; Elneil, S.; Dasgupta, P.; Kessler, T.M.; Khan, S.; Panicker, J.; et al. Suburothelial Myofibroblasts in the Human Overactive Bladder and the Effect of Botulinum Neurotoxin Type A Treatment. Eur. Urol. 2009, 55, 1440–1449. [Google Scholar] [CrossRef] [Green Version]
- Kanai, A.; Wyndaele, J.J.; Andersson, K.E.; Fry, C.; Ikeda, Y.; Zabbarova, I.; De Wachter, S. Researching bladder afferents-determining the effects of beta(3)-adrenergic receptor agonists and botulinum toxin type-A. Neurourol. Urodyn. 2011, 30, 684–691. [Google Scholar] [CrossRef] [Green Version]
- Jung, H.; Lauterburg, T.; Burgunder, J.-M. Expression of neurotransmitter genes in rat spinal motoneurons after chemodenervation with botulinum toxin. Neuroscience 1997, 78, 469–479. [Google Scholar] [CrossRef]
- Humm, A.; Pabst, C.; Lauterburg, T.; Burgunder, J.-M. Enkephalin and aFGF Are Differentially Regulated in Rat Spinal Motoneurons after Chemodenervation with Botulinum Toxin. Exp. Neurol. 2000, 161, 361–372. [Google Scholar] [CrossRef]
- Wiegand, H.; Erdmann, G.; Wellhöner, H.H. 125I-Labelled botulinum a neurotoxin: Pharmacokinetics in cats after intramuscular injection. Naunyn-Schmiedebergs Arch. Pharmacol. 1976, 292, 161–165. [Google Scholar] [CrossRef] [PubMed]
- Papagiannopoulou, D.; Vardouli, L.; Dimitriadis, F.; Apostolidis, A. Retrograde transport of radiolabelled botulinum neurotoxin type A to the CNS after intradetrusor injection in rats. Br. J. Urol. 2015, 117, 697–704. [Google Scholar] [CrossRef] [PubMed]
- Matak, I.; Bach-Rojecky, L.; Filipović, B.; Lacković, Z. Behavioral and immunohistochemical evidence for central antinociceptive activity of botulinum toxin A. Neuroscience 2011, 186, 201–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antonucci, F.; Rossi, C.; Gianfranceschi, L.; Rossetto, O.; Caleo, M. Long-Distance Retrograde Effects of Botulinum Neurotoxin A. J. Neurosci. 2008, 28, 3689–3696. [Google Scholar] [CrossRef]
- Datta, S.N.; Roosen, A.; Popat, R.; Elneil, S.; Dasgupta, P.; Fowler, C.J.; Apostolidis, A. Cholinergic signalling pathways in the superficial layers of the human bladder; Comparing health, disease and the effect of botulinum toxin type A. J. Urol. 2009, 181, 676–677. [Google Scholar] [CrossRef]
- Aoki, K.R. Evidence for Antinociceptive Activity of Botulinum Toxin Type A in Pain Management. Headache 2003, 43, 9–15. [Google Scholar] [CrossRef]
- Andersson, K.-E.; Wein, A.J. Pharmacology of the Lower Urinary Tract: Basis for Current and Future Treatments of Urinary Incontinence. Pharmacol. Rev. 2004, 56, 581–631. [Google Scholar] [CrossRef]
- Yoshimura, N. Bladder afferent pathway and spinal cord injury: Possible mechanisms inducing hyperreflexia of the urinary bladder. Prog. Neurobiol. 1999, 57, 583–606. [Google Scholar] [CrossRef]
- Bossowska, A.; Majewski, M. Botulinum toxin type A-induced changes in the chemical coding of dorsal root ganglion neurons supplying the porcine urinary bladder. Pol. J. Vet. Sci. 2012, 15, 345–353. [Google Scholar] [CrossRef]
- Van den Bergh, P.; De Beukelaer, M.; Deconinck, N. Effect of muscle denervation on the expression of substance P in the ventral raphe-spinal pathway of the rat. Brain Res. 1996, 707, 206–212. [Google Scholar] [CrossRef]
- Lucioni, A.; Bales, G.T.; Lotan, T.; McGehee, D.S.; Cook, S.P.; Rapp, D.E. Botulinum toxin type A inhibits sensory neuropeptide release in rat bladder models of acute injury and chronic inflammation. Br. J. Urol. 2008, 101, 366–370. [Google Scholar] [CrossRef] [PubMed]
- Apostolidis, A.; Popat, R.; Yiangou, Y.; Cockayne, D.; Ford, A.; Davis, J.; Dasgupta, P.; Fowler, C.; Anand, P. Decreased sensory receptors P2X 3 and TRPV1 in suburothelial nerve fibers following intradetrusor injections of botulinum toxin for human detrusor overactivity. J. Urol. 2005, 174, 977–983. [Google Scholar] [CrossRef] [PubMed]
- Mukerji, G.; Yiangou, Y.; Agarwal, S.K.; Anand, P. Transient Receptor Potential Vanilloid Receptor Subtype 1 in Painful Bladder Syndrome and its Correlation with Pain. J. Urol. 2006, 176, 797–801. [Google Scholar] [CrossRef]
- Morenilla-Palao, C.; Planells-Cases, R.; García-Sanz, N.; Ferrer-Montiel, A. Regulated Exocytosis Contributes to Protein Kinase C Potentiation of Vanilloid Receptor Activity. J. Biol. Chem. 2004, 279, 25665–25672. [Google Scholar] [CrossRef] [Green Version]
- Min, J.-W.; Liu, W.-H.; He, X.-H.; Peng, B.-W. Different types of toxins targeting TRPV1 in pain. Toxicon 2013, 71, 66–75. [Google Scholar] [CrossRef]
- Shimizu, T.; Shibata, M.; Toriumi, H.; Iwashita, T.; Funakubo, M.; Sato, H.; Kuroi, T.; Ebine, T.; Koizumi, K.; Suzuki, N. Reduction of TRPV1 expression in the trigeminal system by botulinum neurotoxin type-A. Neurobiol. Dis. 2012, 48, 367–378. [Google Scholar] [CrossRef]
- Giannantoni, A.; Conte, A.; Farfariello, V.; Proietti, S.; Vianello, A.; Nardicchi, V.; Santoni, G.; Amantini, C. Onabotulinumtoxin-A intradetrusorial injections modulate bladder expression of NGF, TrkA, p75 and TRPV1 in patients with detrusor overactivity. Pharmacol. Res. 2013, 68, 118–124. [Google Scholar] [CrossRef]
- Bossowska, A.; Lepiarczyk, E.; Mazur, U.; Janikiewicz, P.; Markiewicz, W. Botulinum toxin type A induces changes in the chemical coding of substance P-immunoreactive dorsal root ganglia sensory neurons supplying the porcine urinary bladder. Toxins 2015, 7, 4797–4816. [Google Scholar] [CrossRef] [Green Version]
- Lepiarczyk, E.; Bossowska, A.; Kaleczyc, J.; Majewski, M. The influence of botulinum toxin type A (BTX) on the immunohistochemical characteristics of noradrenergic and cholinergic nerve fibers supplying the porcine urinary bladder wall. Pol. J. Vet. Sci. 2011, 14, 181–189. [Google Scholar] [CrossRef]
- Datta, S.N.; Roosen, A.; Pullen, A.; Popat, R.; Rosenbaum, T.P.; Elneil, S.; Dasgupta, P.; Fowler, C.J.; Apostolidis, A. Immunohistochemical Expression of Muscarinic Receptors in the Urothelium and Suburothelium of Neurogenic and Idiopathic Overactive Human Bladders, and Changes With Botulinum Neurotoxin Administration. J. Urol. 2010, 184, 2578–2585. [Google Scholar] [CrossRef] [PubMed]
- Pfaff, D.W.; Arnold, A.P.; Etgen, A.M.; Fahrbach, S.E.; Rubin, R.T. (Eds.) Hormones, Brain and Behavior; Academic Press: Cambridge, MA, USA, 2002. [Google Scholar]
- Andersson, K.-E. Detrusor myocyte activity and afferent signaling. Neurourol. Urodyn. 2010, 29, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Apostolidis, A.; Jacques, T.S.; Freeman, A.; Kalsi, V.; Popat, R.; Gonzales, G.; Datta, S.N.; Ghazi-Noori, S.; Elneil, S.; Dasgupta, P.; et al. Histological Changes in the Urothelium and Suburothelium of Human Overactive Bladder following Intradetrusor Injections of Botulinum Neurotoxin Type A for the Treatment of Neurogenic or Idiopathic Detrusor Overactivity. Eur. Urol. 2008, 53, 1245–1253. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.; Khanijou, S.; Rubino, J.; Aoki, K.R. Subcutaneous administration of botulinum toxin A reduces formalin-induced pain. Pain 2004, 107, 125–133. [Google Scholar] [CrossRef]
- Çayan, S.; Coşkun, B.; Bozlu, M.; Acar, D.; Akbay, E.; Ulusoy, E. Botulinum toxin type A may improve bladder function in a rat chemical cystitis model. Urol. Res. 2003, 30, 399–404. [Google Scholar] [CrossRef]
- Richardson, W.S.; Willis, G.W.; Smith, J.W. Evaluation of scar formation after botulinum toxin injection or Forced balloon dilation to the lower esophageal sphincter. Surg. Endosc. 2003, 17, 696–698. [Google Scholar] [CrossRef]
- Haferkamp, A.; Schurch, B.; Reitz, A.; Krengel, U.; Grosse, J.; Kramer, G.; Schumacher, S.; Bastian, P.; Büttner, R.; Müller, S.; et al. Lack of Ultrastructural Detrusor Changes Following Endoscopic Injection of Botulinum Toxin Type A in Overactive Neurogenic Bladder. Eur. Urol. 2004, 46, 784–791. [Google Scholar] [CrossRef]
- Coelho, A.; Oliveira, R.; Rossetto, O.; Cruz, C.; Cruz, F.; Avelino, A. Intrathecal administration of botulinum toxin type A improves urinary bladder function and reduces pain in rats with cystitis. Eur. J. Pain 2014, 18, 1480–1489. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Treatment | p-Values | ||
|---|---|---|---|---|
| Bladder | Dorsal Root Ganglia | Spinal Cord | ||
| Tac1 | 0.000 0.042 | 0.006 - | 0.000 0.042 | |
| TRPV1 | - - | - 0.001 | 0.017 0.005 | |
| NPY | - 0.047 - - | 0.028 0.028 - 0.028 | - - 0.021 0.008 | |
| M1 | 0.030 - | - - | - 0.014 | |
| M2 | 0.002 - - | - - - | 0.000 0.052 0.003 | |
| PENK | - - | - 0.065 | 0.047 - | |
| Protein | Treatment | p-Values | ||
|---|---|---|---|---|
| Bladder | Dorsal Root Ganglia | Spinal Cord | ||
| TRPV1 | - - | 0.383 0.415 | - - | |
| NPY | - | 0.010 | - | |
| M1 | 0.000 0.000 0.000 0.000 | - - - - | - - - - | |
| M2 | - | - | 0.050 | |
| PENK | - - | - 0.054 | - - | |
| Gene | Taqman Assay ID | Thermal Cycling Profile |
|---|---|---|
| PENK | Rn00567566 | 95 °C for 20 min, 45 cycles consisting of 95 °C for 1 min, 60 °C for 20 min. |
| TRPV1 | Rn00583117 | |
| NPY | Rn01410145 | |
| Tac1 | Rn01500392 | |
| M1 | Rn00589936 | |
| M2 | Rn02532311 | |
| M3 | Rn00560986 | |
| GAPDH | Rn01775763 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Markopoulou, S.; Vardouli, L.; Dimitriadis, F.; Psalla, D.; Lambropoulos, A.; Apostolidis, A. Effect of Bladder Injection of OnabotulinumtoxinA on the Central Expression of Genes Associated with the Control of the Lower Urinary Tract: A Study in Normal Rats. Int. J. Mol. Sci. 2022, 23, 14419. https://doi.org/10.3390/ijms232214419
Markopoulou S, Vardouli L, Dimitriadis F, Psalla D, Lambropoulos A, Apostolidis A. Effect of Bladder Injection of OnabotulinumtoxinA on the Central Expression of Genes Associated with the Control of the Lower Urinary Tract: A Study in Normal Rats. International Journal of Molecular Sciences. 2022; 23(22):14419. https://doi.org/10.3390/ijms232214419
Chicago/Turabian StyleMarkopoulou, Soultana, Lina Vardouli, Fotios Dimitriadis, Dimitra Psalla, Alexandros Lambropoulos, and Apostolos Apostolidis. 2022. "Effect of Bladder Injection of OnabotulinumtoxinA on the Central Expression of Genes Associated with the Control of the Lower Urinary Tract: A Study in Normal Rats" International Journal of Molecular Sciences 23, no. 22: 14419. https://doi.org/10.3390/ijms232214419