
Molecular and Morphological Characteristics of the De-Obstructed Rat Urinary Bladder—An Update

Abstract
:1. Introduction
2. Results
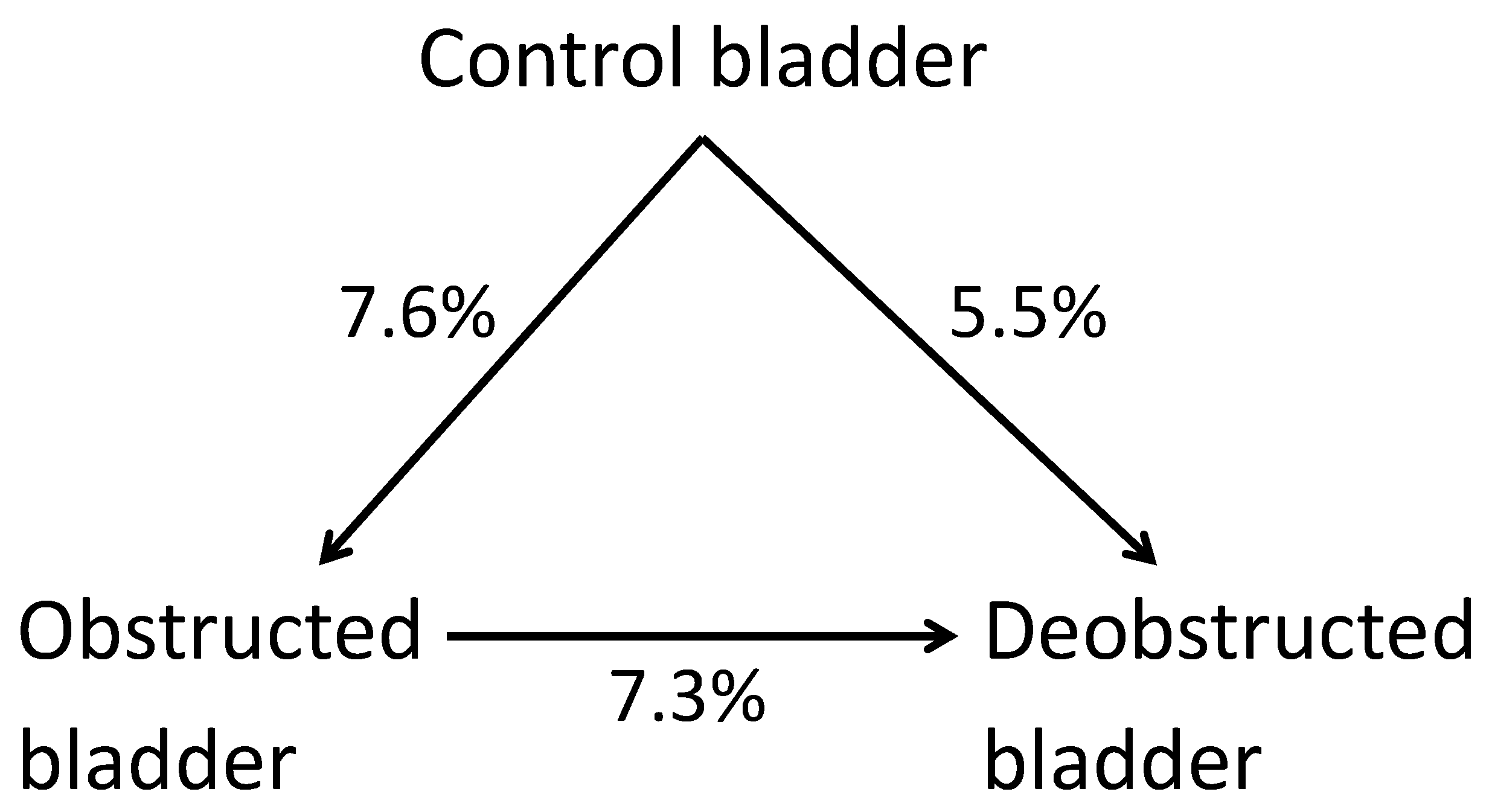
2.1. mRNA Expression in Control, Obstructed, and De-Obstructed Bladders
2.2. Marked mRNA Differences between Obstructed, De-Obstructed, and Control Bladders
2.2.1. mRNAs for Structural Proteins
2.2.2. mRNAs for Proteins Involved in Metabolism
2.2.3. Neurotrophins
2.2.4. Connexins
2.2.5. Cell Surface Receptors
2.2.6. Apoptosis
2.2.7. Inflammation
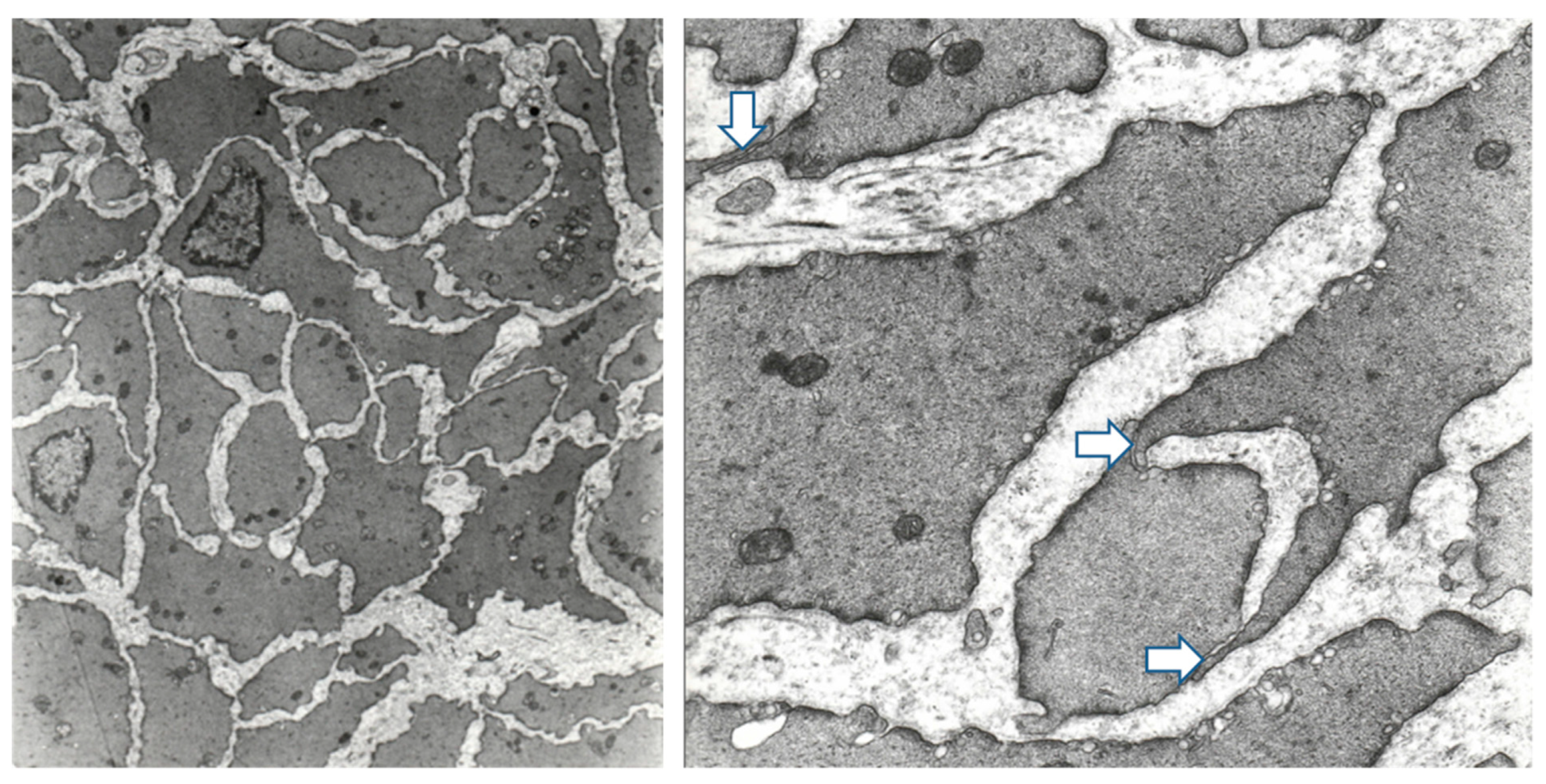
2.3. Structural Differences between Obstructed, De-Obstructed, and Control Detrusors
3. Discussion
3.1. Structural Changes
3.2. Functional Changes
3.3. Structural and Functional Considerations, and Size of Dynamic Motor Units
4. Materials and Methods
4.1. Surgery
4.2. Electron Microscopy
4.3. Microarrays
4.4. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abrams, P.H.; Farrar, D.J.; Turner-Warwick, R.T.; Whiteside, C.G.; Feneley, R.C. The Results of Prostatectomy: A Symptomatic and Urodynamic Analysis of 152 Patients. J. Urol. 1979, 121, 640–642. [Google Scholar] [CrossRef]
- Madersbacher, S.; Marberger, M. Is transurethral resection of the prostate still justified? Br. J. Urol. 1999, 83, 227–237. [Google Scholar] [CrossRef]
- Yang, Q.; Peters, T.J.; Donovan, J.L.; Wilt, T.J.; Abrams, P. Transurethral incision compared with transurethral resection of the prostate for bladder outlet obstruction: A systematic review and meta-analysis of randomized controlled trials. J. Urol. 2001, 165, 1526–1532. [Google Scholar] [CrossRef]
- Tubaro, A.; Carter, S.; Hind, A.; Vicentini, C.; Miano, L. A prospective study of the safety and efficacy of suprapubic transvesical prostatectomy in patients with benign prostatic hyperplasia. J. Urol. 2001, 166, 172–176. [Google Scholar] [CrossRef]
- Antunes, A.A.; Iscaife, A.; Reis, S.; Albertini, A.; Nunes, M.A.; Lucon, A.M.; Nahas, W.C.; Srougi, M. Can We Predict Which Patients will Experience Resolution of Detrusor Overactivity after Transurethral Resection of the Prostate? J. Urol. 2015, 193, 2028–2032. [Google Scholar] [CrossRef]
- Fusco, F.; Creta, M.; De Nunzio, C.; Iacovelli, V.; Mangiapia, F.; Marzi, V.L.; Agrò, E.F. Progressive bladder remodeling due to bladder outlet obstruction: A systematic review of morphological and molecular evidences in humans. BMC Urol. 2018, 18, 15. [Google Scholar] [CrossRef]
- Andersson, K.-E.; Soler, R.; Füllhase, C. Rodent models for urodynamic investigation. Neurourol. Urodynamics 2011, 30, 636–646. [Google Scholar] [CrossRef]
- Fraser, M.O.; Smith, P.P.; Sullivan, M.P.; Bjorling, D.E.; Campeau, L.; Andersson, K.E.; Yoshiyama, M. Best practices for cystometric evaluation of lower urinary tract function in muriform rodents. Neurourol. Urodynamics 2020, 39, 1868–1884. [Google Scholar] [CrossRef]
- Mattiasson, A.; Uvelius, B. Changes in Contractile Properties in Hypertrophic Rat Urinary Bladder. J. Urol. 1982, 128, 1340–1342. [Google Scholar] [CrossRef]
- Malmgren, A.; Sjögren, C.; Uvelius, B.; Mattiasson, A.; Andersson, K.-E.; Andersson, P.O. Cystometrical Evaluation of Bladder Instability in Rats with Infravesical Outflow Obstruction. J. Urol. 1987, 137, 1291–1294. [Google Scholar] [CrossRef]
- Gabella, G.; Uvelius, B. Urinary bladder of rat: Fine structure of normal and hypertrophic musculature. Cell Tissue Res. 1990, 262, 67–79. [Google Scholar] [CrossRef]
- Uvelius, B.; Persson, L.; Mattiasson, A. Smooth muscle cell hypertrophy and hyperplasia in the rat detrusor after short-time in-fravesical outflow obstruction. J. Urol. 1984, 131, 173–176. [Google Scholar] [CrossRef]
- Saito, M.; Longhurst, P.A.; Murphy, M.; Monson, F.C.; Wein, A.J.; Levin, R.M. 3H-thymidine uptake by the rat urinary bladder after partial outflow obstruction. Neurourol. Urodynamics 1994, 13, 63–69. [Google Scholar] [CrossRef]
- Malmqvist, U.; Arner, A.; Uvelius, B. Contractile and cytoskeletal proteins in smooth muscle during hypertrophy and its reversal. Am. J. Physiol. Physiol. 1991, 260, C1085–C1093. [Google Scholar] [CrossRef]
- Malmqvist, U.; Arner, A.; Uvelius, B. Cytoskeletal and Contractile Proteins in Detrusor Smooth Muscle from Bladders with Outlet Obstruction—A Comparative Study in RAT and MAN. Scand. J. Urol. Nephrol. 1991, 25, 261–267. [Google Scholar] [CrossRef]
- Uvelius, B.; Mattiasson, A. Collagen Content in the Rat Urinary Bladder Subjected to Infra Vesical Outflow Obstruction. J. Urol. 1984, 132, 587–590. [Google Scholar] [CrossRef]
- Damaser, M.S.; Arner, A.; Uvelius, B. Partial outlet obstruction induces chronic distension and increased stiffness of rat urinary bladder. Neurourol Urodyn. 1996, 15, 650–665. [Google Scholar] [CrossRef]
- Sadegh, M.K.; Ekman, M.; Krawczyk, K.; Svensson, D.; Göransson, O.; Dahan, D.; Nilsson, B.-O.; Albinsson, S.; Uvelius, B.; Swärd, K. Detrusor Induction of miR-132/212 following Bladder Outlet Obstruction: Association with MeCP2 Repression and Cell Viability. PLoS ONE 2015, 10, e0116784. [Google Scholar] [CrossRef]
- Turner, W.H.; Brading, A.F. Smooth muscle of the bladder in the normal and the diseased state: Pathophysiology, diagnosis and treatment. Pharmacol. Ther. 1997, 75, 77–110. [Google Scholar] [CrossRef]
- Lindner, P.; Mattiasson, A.; Persson, L.; Uvelius, B. Reversibility of detrusor hypertrophy and hyperplasia after removal of in-fravesical outflow obstruction in the rat. J. Urol. 1988, 140, 642–646. [Google Scholar] [CrossRef] [PubMed]
- Gabella, G.; Uvelius, B. Reversal of muscle hypertrophy in the rat urinary bladder after removal of urethral obstruction. Cell Tissue Res. 1994, 277, 333–339. [Google Scholar] [CrossRef]
- Malmgren, A.; Uvelius, B.; Andersson, K.-E.; Andersson, P. On the Reversibility of Functional Bladder Changes Induced by Infra Vesical Outflow Obstruction in the Rat. J. Urol. 1990, 143, 1026–1031. [Google Scholar] [CrossRef]
- Chai, T.C.; Gemalmaz, H.; Andersson, K.E.; Tuttle, J.B.; Steers, W.D. Persistently increased voiding frequency despite relief of bladder outlet obstruction. J. Urol. 1999, 161, 1689–1693. [Google Scholar] [CrossRef]
- Jin, L.-H.; Andersson, K.-E.; Han, J.-U.; Kwon, Y.-H.; Park, C.-S.; Shin, H.-Y.; Yoon, S.-M.; Lee, T. Persistent detrusor overactivity in rats after relief of partial urethral obstruction. Am. J. Physiol. Integr. Comp. Physiol. 2011, 301, R896–R904. [Google Scholar] [CrossRef]
- Ekman, M.; Bhattachariya, A.; Dahan, D.; Uvelius, B.; Albinsson, S.; Swärd, K. Mir-29 Repression in Bladder Outlet Obstruction Contributes to Matrix Remodeling and Altered Stiffness. PLoS ONE 2013, 8, e82308. [Google Scholar] [CrossRef]
- Gheinani, A.H.; Köck, I.; Vasquez, E.; Baumgartner, U.; Bigger-Allen, A.; Sack, B.S.; Burkhard, F.C.; Adam, R.M.; Monastyrskaya, K. Concordant miRNA and mRNA expression profiles in humans and mice with bladder outlet obstruction. Am. J. Clin. Exp. Urol. 2018, 6, 219–233. [Google Scholar]
- Zeidan, A.; Nordström, I.; Dreja, K.; Malmqvist, U.; Hellstrand, P. Stretch-Dependent Modulation of Contractility and Growth in Smooth Muscle of Rat Portal Vein. Circ. Res. 2000, 87, 228–234. [Google Scholar] [CrossRef]
- Saito, M.; Longhurst, P.A.; Murphy, M.; Monson, F.C.; Wein, A.J.; Levin, R.M. Effect of slow and rapid cystometry on in vitro rat urinary bladder DNA synthesis. Gen. Pharmacol. Vasc. Syst. 1994, 25, 1021–1025. [Google Scholar] [CrossRef]
- Levin, R.M.; O’Connor, L.J.; Leggett, R.E.; Whitbeck, C.; Chichester, P. Focal hypoxia of the obstructed rabbit bladder wall correlates with intermediate decompensation. Neurourol. Urodynamics 2003, 22, 156–163. [Google Scholar] [CrossRef]
- Uvelius, B.; Lindner, P.; Mattiasson, A. Collagen content in the rat urinary bladder following removal of an experimental in-fravesical outlet obstruction. Urol Int. 1991, 47, 245–249. [Google Scholar] [CrossRef] [PubMed]
- Ekman, M.; Uvelius, B.; Albinsson, S.; Swärd, K. HIF-mediated metabolic switching in bladder outlet obstruction mitigates the relaxing effect of mitochondrial inhibition. Lab. Investig. 2014, 94, 557–568. [Google Scholar] [CrossRef] [Green Version]
- Fry, C.H.; McCloskey, K.D. Spontaneous Activity and the Urinary Bladder. Adv. Exp. Med. Biol. 2019, 1124, 121–147. [Google Scholar] [CrossRef]
- Drake, M.; Harvey, I.J.; Gillespie, J.I. Autonomous Activity in the Isolated Guinea Pig Bladder. Exp. Physiol. 2003, 88, 19–30. [Google Scholar] [CrossRef]
- Ekström, J.; Uvelius, B. Length-tension relations of smooth muscle from normal and denervated rat urinary bladders. Acta Physiol. Scand. 1981, 112, 443–447. [Google Scholar] [CrossRef]
- Arner, A.; Malmqvist, U.; Uvelius, B. Metabolism and force in hypertrophic smooth muscle from rat urinary bladder. Am. J. Physiol. Physiol. 1990, 258, C923–C932. [Google Scholar] [CrossRef]
- Polyanska, M.; Arner, A.; Malmquist, U.; Uvelius, B. Lactate Dehydrogenase Activity and Isoform Distribution in the Rat Urinary Bladder: Effects of Outlet Obstruction and its Removal. J. Urol. 1993, 150, 543–545. [Google Scholar] [CrossRef]
- Uvelius, B.; Arner, A. Changed metabolism of detrusor muscle cells from obstructed rat urinary bladder. Scand. J. Urol. Nephrol. Suppl. 1997, 184, 59–65. [Google Scholar]
- Gabella, G. Structure of the intramural nerves of the rat bladder. J. Neurocytol. 1999, 28, 615–637. [Google Scholar] [CrossRef]
- Gabella, G. The structural relations between nerve fibres and muscle cells in the urinary bladder of the rat. J. Neurocytol. 1995, 24, 159–187. [Google Scholar] [CrossRef]
- Bibbiani, C.; Tongiani, R.; Viola-Magni, M.P.I. QUANTITATIVE DETERMINATION OF THE AMOUNT OF DNA PER NUCLEUS BY INTERFERENCE MICROSCOPY. J. Cell Biol. 1969, 42, 444–451. [Google Scholar] [CrossRef]
- Goodenough, D.A.; Paul, D.L. Gap Junctions. Cold Spring Harb. Perspect. Biol. 2009, 1, a002576. [Google Scholar] [CrossRef] [PubMed]
- Christ, G.J.; Day, N.S.; Day, M.; Zhao, W.; Persson, K.; Pandita, R.K.; Andersson, K.-E. Increased connexin43-mediated intercellular communication in a rat model of bladder overactivity in vivo. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2003, 284, R1241–R1248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haefliger, J.-A.; Tissières, P.; Tawadros, T.; Formenton, A.; Bény, J.L.; Nicod, P.; Frey, P.; Meda, P. Connexins 43 and 26 Are Differentially Increased after Rat Bladder Outlet Obstruction. Exp. Cell Res. 2002, 274, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Nitti, V.W.; Rosenberg, S.; Mitcheson, D.H.; He, W.; Fakhoury, A.; Martin, N.E. Urodynamics and Safety of the β3-Adrenoceptor Agonist Mirabegron in Males with Lower Urinary Tract Symptoms and Bladder Outlet Obstruction. J. Urol. 2013, 190, 1320–1327. [Google Scholar] [CrossRef]
- Liao, C.-H.; Kuo, H.-C. Mirabegron 25 mg Monotherapy Is Safe but Less Effective in Male Patients With Overactive Bladder and Bladder Outlet Obstruction. Urology 2018, 117, 115–119. [Google Scholar] [CrossRef]
- Zeng, J.; Ekman, M.; Jiang, C.; Uvelius, B.; Swärd, K. Non-uniform changes in membrane receptors in the rat urinary bladder following outlet obstruction. Eur. J. Pharmacol. 2015, 762, 82–88. [Google Scholar] [CrossRef]
- Lee, J.Y.; Park, J.M.; Gil Na, Y.G.; Song, K.H.; Lim, J.S.; Yang, S.W.; Kim, G.H.; Shin, J.H. Expression of bladder α1-adrenoceptor subtype after relief of partial bladder outlet obstruction in a rat model. Investig. Clin. Urol. 2020, 61, 297–303. [Google Scholar] [CrossRef]
- Zhu, B.; Ekman, M.; Svensson, D.; Lindvall, J.M.; Nilsson, B.-O.; Uvelius, B.; Swärd, K. Array profiling reveals contribution of Cthrc1 to growth of the denervated rat urinary bladder. Am. J. Physiol. Renal. Physiol. 2018, 314, F893–F905. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Gene Description | Fold Change |
|---|---|---|
| Grem1 | gremlin 1, cysteine knot superfamily, homolog (Xenopus laevis) | 17,13979 |
| Trim59 | tripartite motif-containing 59 | 2,722014 |
| Cdkn1c | cyclin-dependent kinase inhibitor 1C | 2,672856 |
| Rspo3 | R-spondin 3 homolog (Xenopus laevis) | 2,502077 |
| RGD1564327 | similar to integrin alpha 8 | 2,322538 |
| Cthrc1 | collagen triple helix repeat containing 1 | 2,212791 |
| Eda2r | ectodysplasin A2 receptor | 2,182682 |
| Fxyd6 | FXYD domain-containing ion transport regulator 6 | 2,136691 |
| Adora2b | adenosine A2B receptor | 2,105719 |
| Edn1 | endothelin 1 | 2,099241 |
| Acadsb | acyl-Coenzyme A dehydrogenase, short/branched chain | 2,090642 |
| Angptl1 | angiopoietin-like 1 | 2,064316 |
| Abcb1b | ATP-binding cassette, sub-family B (MDR/TAP), member 1B | 2,057768 |
| Phlda1 | pleckstrin homology-like domain, family A, member 1 | 0,494031 |
| Cadps | Ca++-dependent secretion activator | 0,49359 |
| Col1a1 | collagen type I alpha 1 | 0,492934 |
| Nrk | Nik related kinase | 0,491153 |
| Mfap2 | microfibrillar-associated protein 2 | 0,469284 |
| Nrg1 | neuregulin 1 | 0,466933 |
| Itm2a | integral membrane protein 2A | 0,450881 |
| Slc30a10 | solute carrier family 30, member 10 | 0,445451 |
| Hhip | Hedgehog-interacting protein | 0,437063 |
| Ahrr | aryl-hydrocarbon receptor repressor | 0,43424 |
| Krt20 | keratin 20 | 0,415004 |
| Fam111a | family with sequence similarity 111, member A | 0,403881 |
| Igfbp3 | insulin-like growth factor binding protein 3 | 0,322226 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uvelius, B.; Andersson, K.-E. Molecular and Morphological Characteristics of the De-Obstructed Rat Urinary Bladder—An Update. Int. J. Mol. Sci. 2022, 23, 11330. https://doi.org/10.3390/ijms231911330
Uvelius B, Andersson K-E. Molecular and Morphological Characteristics of the De-Obstructed Rat Urinary Bladder—An Update. International Journal of Molecular Sciences. 2022; 23(19):11330. https://doi.org/10.3390/ijms231911330
Chicago/Turabian StyleUvelius, Bengt, and Karl-Erik Andersson. 2022. "Molecular and Morphological Characteristics of the De-Obstructed Rat Urinary Bladder—An Update" International Journal of Molecular Sciences 23, no. 19: 11330. https://doi.org/10.3390/ijms231911330