
Identification of the Karyopherin Superfamily in Maize and Its Functional Cues in Plant Development

Abstract
:1. Introduction
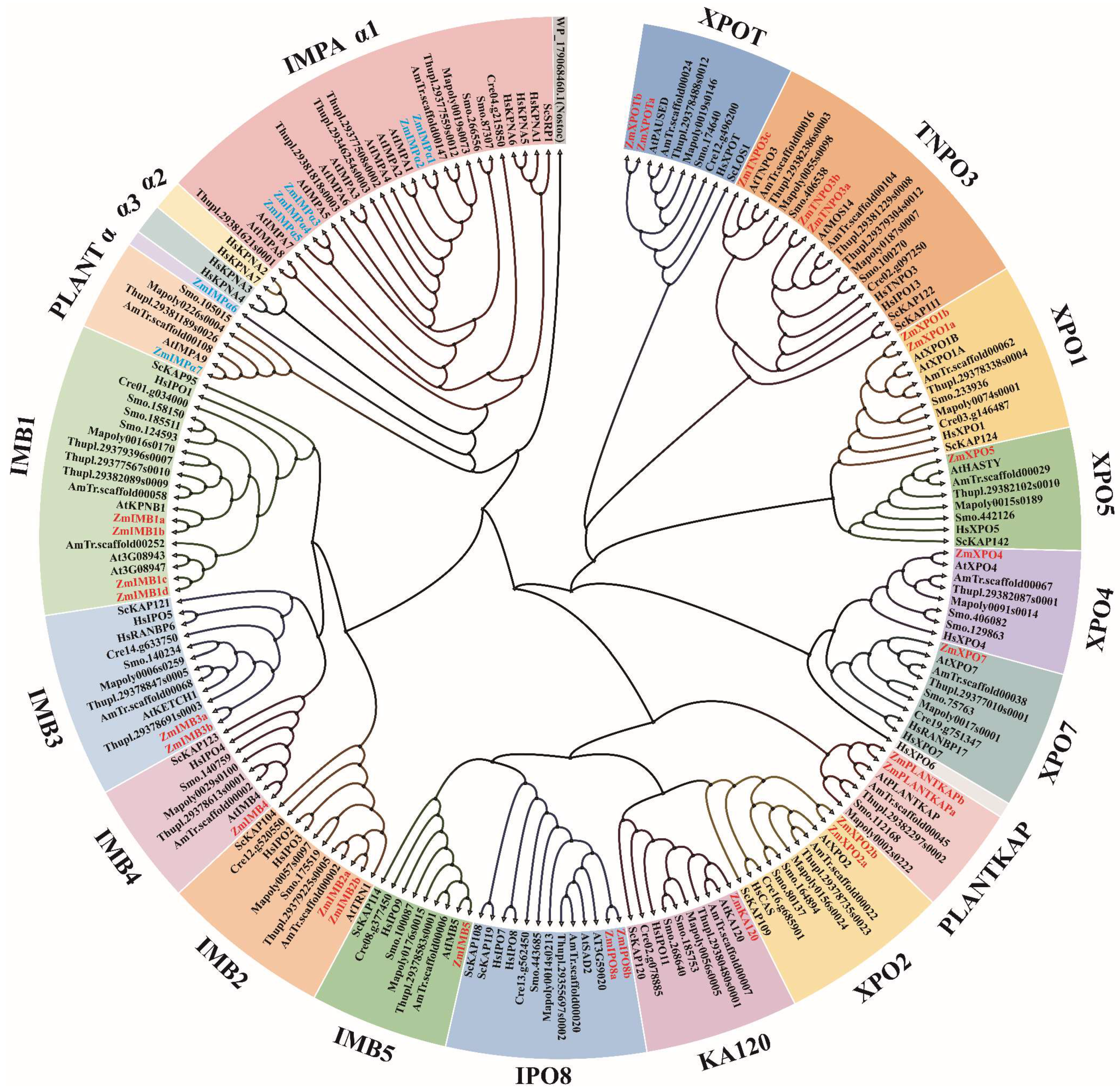
2. A General View of the Karyopherin Superfamily
2.1. Evolution of the Karyopherins
2.1.1. Importin α
2.1.2. Importin β
2.2. The Karyopherin-Mediated NCT Pathway
2.2.1. The Classical Nuclear Protein Import Cycle in Yeast and Mammals
2.2.2. The IMPα- and IMPβ-Mediated Nuclear Transport Pathway in Plants
2.2.3. The IMPβ-Dependent Nuclear Translocating Pathway
3. Importin α Family in Maize and Arabidopsis
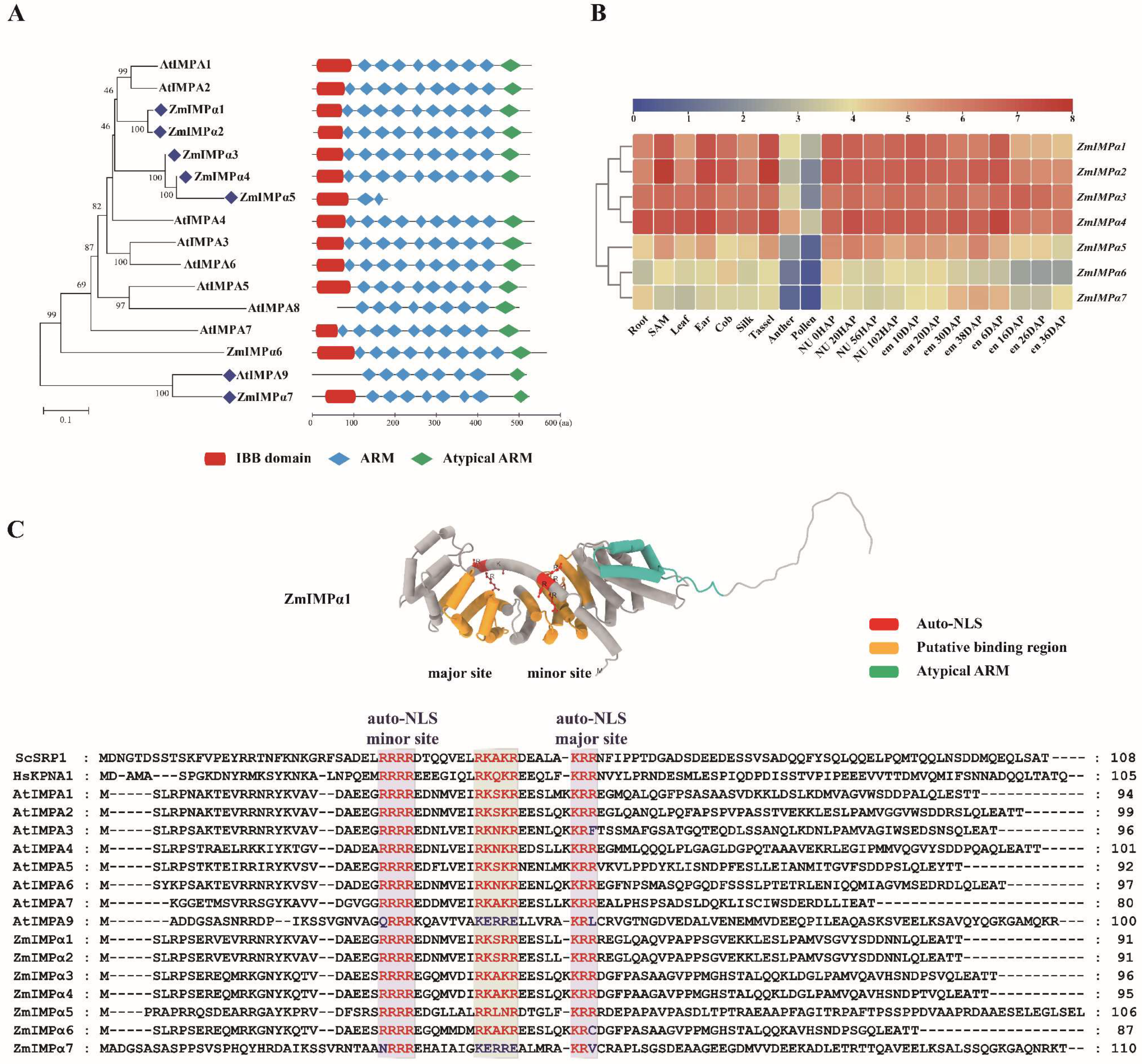
3.1. Protein Domain Distribution and Gene Expression Profiles of the ZmIMPαs
3.2. Multifunctionality of the IBB Domain
3.3. The ARM Repeat and Classical NLS Recognition
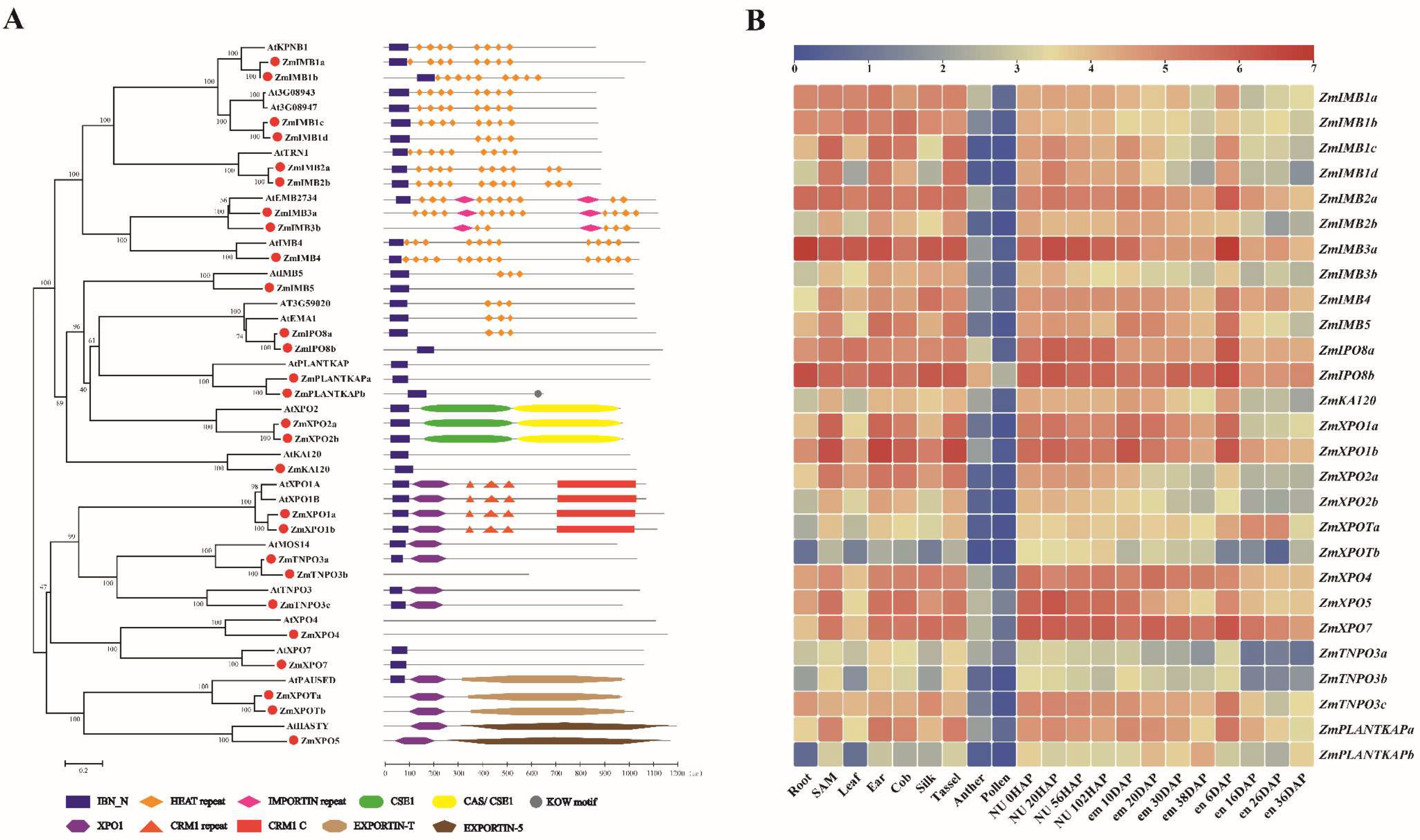
4. Importin β Family in Maize and Arabidopsis
4.1. The Characteristic Domains of IMPβ Proteins
4.1.1. Importin
4.1.2. Exportin
4.2. The Function of the IBN_N Domain and Ran System
4.3. The Non-Classical NLS and NES Recognized by Importin β
4.3.1. The Non-Classical NLS
4.3.2. NES
5. Functional Cues of Karyopherins in Hormone Signaling and Plant Development
5.1. The Roles of Arabidopsis KAPs in Hormone Signaling
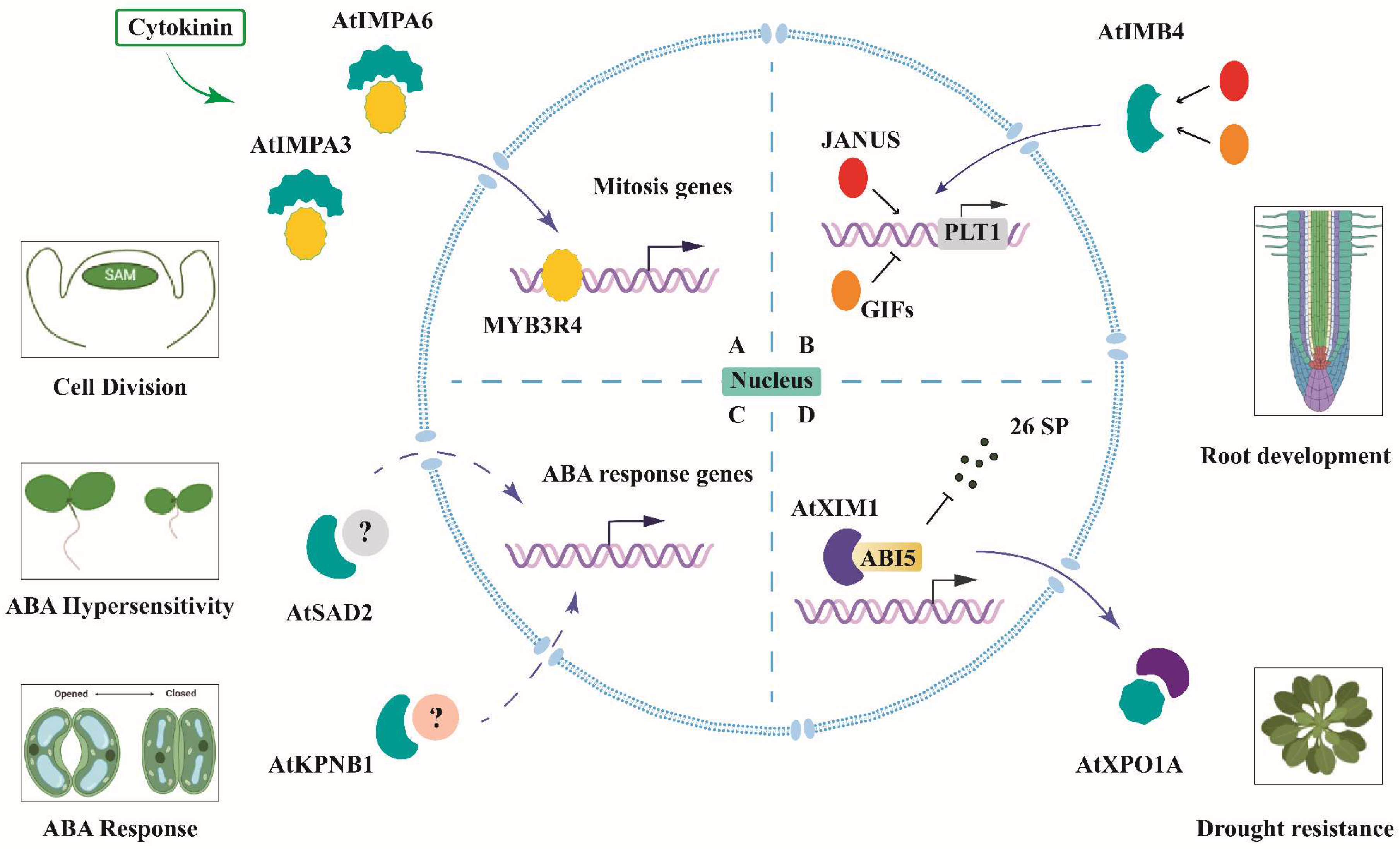
5.1.1. AtIMB4 and PLT1-Mediated Root Development
5.1.2. AtIMPA3/6 and Cytokinin-Activated Cell Division in Shoot Apical Meristem
5.1.3. AtIMPβs and ABA Signaling in Response to Abiotic Stress
5.2. The Predicted Interacting Protein of the ZmKAPs Involved in the Auxin Pathway
5.2.1. Auxin Biosynthesis
5.2.2. Auxin Transport
5.2.3. Auxin Signaling
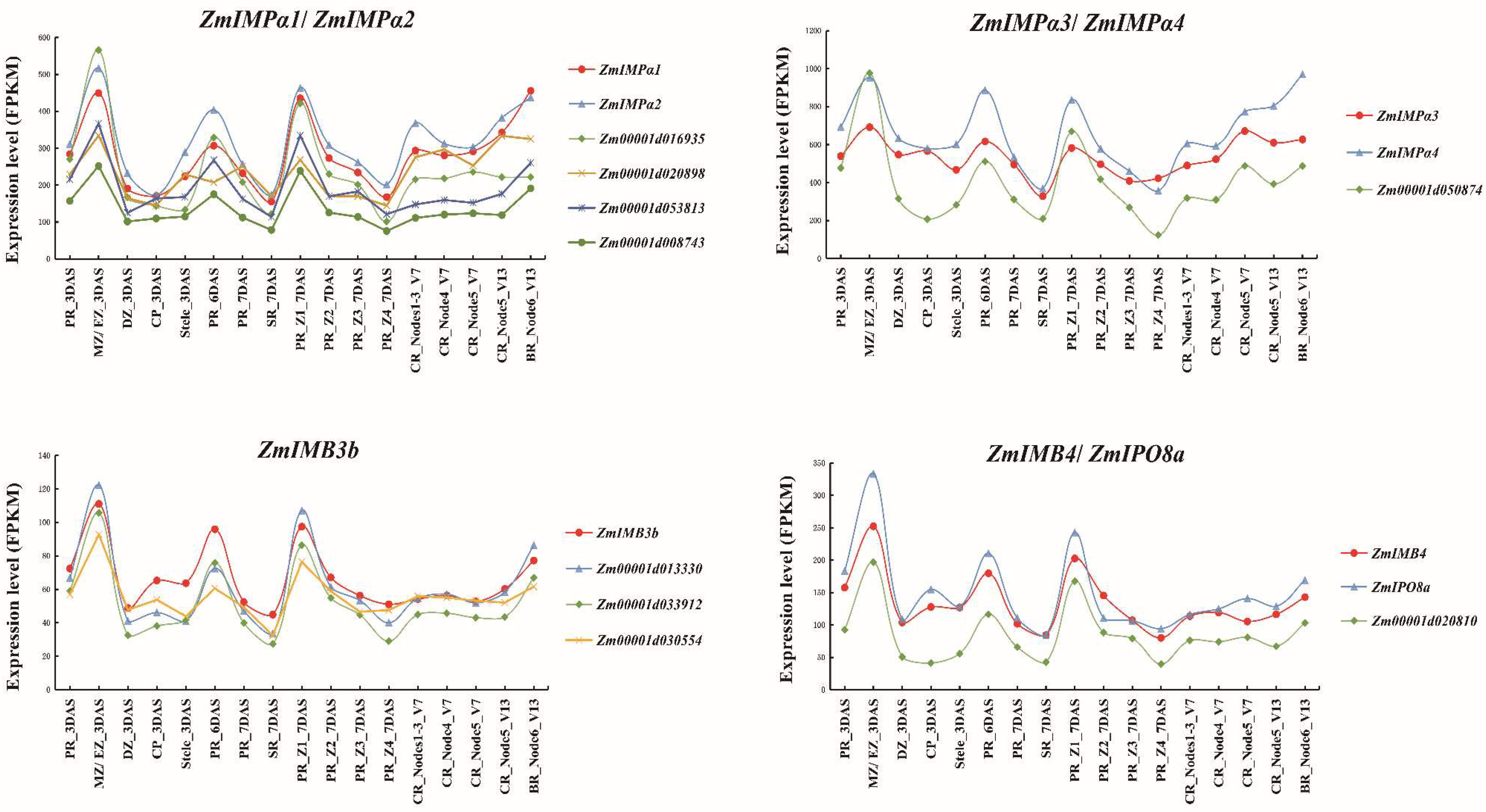
5.3. Expression Profiles of ZmKAPs and Corresponding Interaction Partners in Root Development
6. Conclusions and Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Beck, M.; Hurt, E. The nuclear pore complex: Understanding its function through structural insight. Nat. Rev. Mol. Cell Biol. 2017, 18, 73–89. [Google Scholar] [CrossRef] [PubMed]
- Ribbeck, K.; Görlich, D. Kinetic analysis of translocation through nuclear pore complexes. EMBO J. 2001, 20, 1320–1330. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.H.; Hoelz, A. The Structure of the Nuclear Pore Complex (An Update). Annu. Rev. Biochem. 2019, 88, 725–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aramburu, I.V.; Lemke, E.A. Floppy but not sloppy: Interaction mechanism of FG-nucleoporins and nuclear transport receptors. Semin. Cell Dev. Biol. 2017, 68, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Paci, G.; Caria, J.; Lemke, E.A. Cargo transport through the nuclear pore complex at a glance. J. Cell Sci. 2021, 134, jcs247874. [Google Scholar] [CrossRef]
- Knockenhauer, K.E.; Schwartz, T.U. The Nuclear Pore Complex as a Flexible and Dynamic Gate. Cell 2016, 164, 1162–1171. [Google Scholar] [CrossRef] [Green Version]
- Chook, Y.M.; Blobel, G. Karyopherins and nuclear import. Curr. Opin. Struct. Biol. 2001, 11, 703–715. [Google Scholar] [CrossRef]
- Cook, A.; Bono, F.; Jinek, M.; Conti, E. Structural Biology of Nucleocytoplasmic Transport. Annu. Rev. Biochem. 2007, 76, 647–671. [Google Scholar] [CrossRef]
- Chang, C.C.; Hsia, K.C. More than a zip code: Global modulation of cellular function by nuclear localization signals. FEBS J. 2021, 288, 5569–5585. [Google Scholar] [CrossRef]
- Gerace, L. Nuclear export signals and the fast track to the cytoplasm. Cell 1995, 82, 341–344. [Google Scholar] [CrossRef]
- Allen, J.R.; Strader, L.C. Nucleocytoplasmic partitioning as a mechanism to regulate Arabidopsis signaling events. Curr. Opin. Cell Biol. 2021, 69, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Merkle, T. Nucleo-cytoplasmic partitioning of proteins in plants: Implications for the regulation of environmental and developmental signalling. Curr. Genet. 2003, 44, 231–260. [Google Scholar] [CrossRef] [PubMed]
- Xiong, F.; Groot, E.P.; Zhang, Y.; Li, S. Functions of plant importin β proteins beyond nucleocytoplasmic transport. J. Exp. Bot. 2021, 72, 6140–6149. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Wang, Y.E.; Palazzo, A.F. Crosstalk between nucleocytoplasmic trafficking and the innate immune response to viral infection. J. Biol. Chem. 2021, 297, 100856. [Google Scholar] [CrossRef] [PubMed]
- Kai, Z.S.; Pasquinelli, A.E. MicroRNA assassins: Factors that regulate the disappearance of miRNAs. Nat. Struct. Mol. Biol. 2010, 17, 5–10. [Google Scholar] [CrossRef]
- Li, C.; Qi, W.; Liang, Z.; Yang, X.; Ma, Z.; Song, R. A SnRK1-ZmRFWD3-Opaque2 Signaling Axis Regulates Diurnal Nitrogen Accumulation in Maize Seeds. Plant Cell 2020, 32, 2823–2841. [Google Scholar] [CrossRef]
- Zhang, N.; Qiao, Z.; Liang, Z.; Mei, B.; Xu, Z.; Song, R. Zea mays Taxilin Protein Negatively Regulates Opaque-2 Transcriptional Activity by Causing a Change in Its Sub-Cellular Distribution. PLoS ONE 2012, 7, e43822. [Google Scholar] [CrossRef] [Green Version]
- Tsuji, L.; Takumi, T.; Imamoto, N.; Yoneda, Y. Identification of novel homologues of mouse importin alpha, the alpha subunit of the nuclear pore-targeting complex, and their tissue-specific expression. FEBS Lett. 1997, 416, 30–34. [Google Scholar] [CrossRef] [Green Version]
- Quan, Y.; Ji, Z.-L.; Wang, X.; Tartakoff, A.M.; Tao, T. Evolutionary and transcriptional analysis of karyopherin beta superfamily proteins. Mol. Cell. Proteom. 2008, 7, 1254–1269. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Hara-Nishimura, I. Functional insights of nucleocytoplasmic transport in plants. Front. Plant Sci. 2014, 5, 118. [Google Scholar] [CrossRef]
- Xu, Y.; Liu, L.; Zhao, P.; Tong, J.; Zhong, N.; Zhang, H.; Liu, N. Genome-Wide Identification, Expression Profile and Evolution Analysis of Karyopherin β Gene Family in Solanum tuberosum Group Phureja DM1-3 Reveals Its Roles in Abiotic Stresses. Int. J. Mol. Sci. 2020, 21, 931. [Google Scholar] [CrossRef] [Green Version]
- Goldfarb, D.S.; Corbett, A.H.; Mason, D.A.; Harreman, M.T.; Adam, S.A. Importin alpha: A multipurpose nuclear-transport receptor. Trends Cell Biol. 2004, 14, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Görlich, D.; Kutay, U. Transport between the Cell Nucleus and the Cytoplasm. Annu. Rev. Cell Dev. Biol. 1999, 15, 607–660. [Google Scholar] [CrossRef]
- Yoshida, K.; Blobel, G. The Karyopherin Kap142p/Msn5p Mediates Nuclear Import and Nuclear Export of Different Cargo Proteins. J. Cell Biol. 2001, 152, 729–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gontan, C.; Güttler, T.; Engelen, E.; Demmers, J.; Fornerod, M.; Grosveld, F.G.; Tibboel, D.; Görlich, D.; Poot, R.A.; Rottier, R.J. Exportin 4 mediates a novel nuclear import pathway for Sox family transcription factors. J. Cell Biol. 2009, 185, 27–34. [Google Scholar] [CrossRef] [Green Version]
- Grünwald, M.; Lazzaretti, D.; Bono, F. Structural basis for the nuclear export activity of Importin13. EMBO J. 2013, 32, 899–913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aksu, M.; Pleiner, T.; Karaca, S.; Kappert, C.; Dehne, H.J.; Seibel, K.; Urlaub, H.; Bohnsack, M.T.; Görlich, D. Xpo7 is a broad-spectrum exportin and a nuclear import receptor. J. Cell Biol. 2018, 217, 2329–2340. [Google Scholar] [CrossRef]
- Aksu, M.; Trakhanov, S.; Rodriguez, A.V.; Görlich, D. Structural basis for the nuclear import and export functions of the biportin Pdr6/Kap122. J. Cell Biol. 2019, 218, 1839–1852. [Google Scholar] [CrossRef] [Green Version]
- Schnable, J.C.; Springer, N.M.; Freeling, M. Differentiation of the maize subgenomes by genome dominance and both ancient and ongoing gene loss. Proc. Natl. Acad. Sci. USA 2011, 108, 4069–4074. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Ngu, D.W.; Carvalho, D.; Liang, Z.; Qiu, Y.; Roston, R.L.; Schnable, J.C. Differentially Regulated Orthologs in Sorghum and the Subgenomes of Maize. Plant Cell 2017, 29, 1938–1951. [Google Scholar] [CrossRef]
- Umeda, M.; Izaddoost, S.; Cushman, I.; Moore, M.S.; Sazer, S. The Fission Yeast Schizosaccharomyces pombe Has Two Importin-alpha Proteins, Imp1p and Cut15p, Which Have Common and Unique Functions in Nucleocytoplasmic Transport and Cell Cycle Progression. Genetics 2005, 171, 7–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mason, D.A.; Stage, D.E.; Goldfarb, D.S. Evolution of the Metazoan-Specific Importin alpha Gene Family. J. Mol. Evol. 2009, 68, 351–365. [Google Scholar] [CrossRef] [PubMed]
- O’Reilly, A.J.; Dacks, J.B.; Field, M.C. Evolution of the Karyopherin-β Family of Nucleocytoplasmic Transport Factors; Ancient Origins and Continued Specialization. PLoS ONE 2011, 6, e19308. [Google Scholar] [CrossRef] [Green Version]
- Koumandou, V.L.; Wickstead, B.; Ginger, M.L.; van der Giezen, M.; Dacks, J.B.; Field, M.C. Molecular paleontology and complexity in the last eukaryotic common ancestor. Crit. Rev. Biochem. Mol. Biol. 2013, 48, 373–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, M. Molecular mechanism of the nuclear protein import cycle. Nat. Rev. Mol. Cell Biol. 2007, 8, 195–208. [Google Scholar] [CrossRef]
- Görlich, D.; Kostka, S.; Kraft, R.; Dingwall, C.; Laskey, R.A.; Hartmann, E.; Prehn, S. Two different subunits of importin cooperate to recognize nuclear localization signals and bind them to the nuclear envelope. Curr. Biol. 1995, 5, 383–392. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.M.; Stewart, M. Structural Basis for the High-affinity Binding of Nucleoporin Nup1p to the Saccharomyces cerevisiae Importin-beta Homologue, Kap95p. J. Mol. Biol. 2005, 349, 515–525. [Google Scholar] [CrossRef]
- Lee, S.J.; Matsuura, Y.; Liu, S.M.; Stewart, M. Structural basis for nuclear import complex dissociation by RanGTP. Nature 2005, 435, 693–696. [Google Scholar] [CrossRef]
- Kobe, B. Autoinhibition by an internal nuclear localization signal revealed by the crystal structure of mammalian importin alpha. Nat. Struct. Biol. 1999, 6, 388–397. [Google Scholar] [CrossRef]
- Matsuura, Y.; Stewart, M. Structural basis for the assembly of a nuclear export complex. Nature 2004, 432, 872–877. [Google Scholar] [CrossRef]
- Luo, Y.; Wang, Z.; Ji, H.; Fang, H.; Wang, S.; Tian, L.; Li, X. An Arabidopsis homolog of importin β1 is required for ABA response and drought tolerance. Plant J. 2013, 75, 377–389. [Google Scholar] [CrossRef] [PubMed]
- Haasen, D.; Merkle, T. Characterization of an Arabidopsis thaliana Homologue of the Nuclear Export Receptor CAS by its Interaction with Importin α. Plant Biol. 2008, 14, 432–439. [Google Scholar] [CrossRef]
- Cambiagno, D.A.; Giudicatti, A.J.; Arce, A.L.; Gagliardi, D.; Li, L.; Yuan, W.; Lundberg, D.S.; Weigel, D.; Manavella, P.A. HASTY modulates miRNA biogenesis by linking pri-miRNA transcription and processing. Mol. Plant 2021, 14, 426–439. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.-J.; Imamoto, N.; Matsuki, R.; Yoneda, Y.; Yamamoto, N. Functional characterization of a plant importin alpha homologue. Nuclear localization signal (NLS)-selective binding and mediation of nuclear import of NLS proteins in vitro. J. Biol. Chem. 1998, 273, 24083–24087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hübner, S.; Smith, H.; Hu, W.; Chan, C.K.; Rihs, H.P.; Paschal, B.M.; Raikhel, N.V.; Jans, D.A. Plant importin alpha binds nuclear localization sequences with high affinity and can mediate nuclear import independent of importin beta. J. Biol. Chem. 1999, 274, 22610–22617. [Google Scholar] [CrossRef] [Green Version]
- Pemberton, L.F.; Paschal, B.M. Mechanisms of Receptor-Mediated Nuclear Import and Nuclear Export. Traffic 2005, 6, 187–198. [Google Scholar] [CrossRef]
- Cingolani, G.; Bednenko, J.; Gillespie, M.; Gerace, L. Molecular Basis for the Recognition of a Nonclassical Nuclear Localization Signal by Importin beta. Mol. Cell 2002, 10, 1345–1353. [Google Scholar] [CrossRef]
- Tran, E.J.; Bolger, T.A.; Wente, S.R. SnapShot: Nuclear Transport. Cell 2007, 131, 420. [Google Scholar] [CrossRef] [Green Version]
- Kapinos, L.E.; Schoch, R.L.; Wagner, R.S.; Schleicher, K.D.; Lim, R.Y. Karyopherin-Centric Control of Nuclear Pores Based on Molecular Occupancy and Kinetic Analysis of Multivalent Binding with FG Nucleoporins. Biophys. J. 2014, 106, 1751–1762. [Google Scholar] [CrossRef] [Green Version]
- Nachury, M.V.; Weis, K. The direction of transport through the nuclear pore can be inverted. Proc. Natl. Acad. Sci. USA 1999, 96, 9622–9627. [Google Scholar] [CrossRef]
- Izaurralde, E.; Kutay, U.; von Kobbe, C.; Mattaj, I.W.; Görlich, D. The asymmetric distribution of the constituents of the Ran system is essential for transport into and out of the nucleus. EMBO J. 1997, 16, 6535–6547. [Google Scholar] [CrossRef] [PubMed]
- Andrade, M.A.; Petosa, C.; O’Donoghue, S.I.; Müller, C.W.; Bork, P. Comparison of ARM and HEAT protein repeats. J. Mol. Biol. 2001, 309, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Malik, H.S.; Eickbush, T.H.; Goldfarb, D.S. Evolutionary specialization of the nuclear targeting apparatus. Proc. Natl. Acad. Sci. USA 1997, 94, 13738–13742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cingolani, G.; Petosa, C.; Weis, K.; Müller, C.W. Structure of importin-β bound to the IBB domain of importin-alpha. Nature 1999, 399, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Lange, A.; Mills, R.E.; Lange, C.J.; Stewart, M.; Devine, S.E.; Corbett, A.H. Classical Nuclear Localization Signals: Definition, Function, and Interaction with Importin alpha. J. Biol. Chem. 2007, 282, 5101–5105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herold, A.; Truant, R.; Wiegand, H.; Cullen, B.R. Determination of the Functional Domain Organization of the Importin α Nuclear Import Factor. J. Cell Biol. 1998, 143, 309–318. [Google Scholar] [CrossRef] [Green Version]
- Yi, F.; Gu, W.; Chen, J.; Song, N.; Gao, X.; Zhang, X.; Zhou, Y.; Ma, X.; Song, W.; Zhao, H.; et al. High Temporal-Resolution Transcriptome Landscape of Early Maize Seed Development. Plant Cell 2019, 31, 974–992. [Google Scholar] [CrossRef] [Green Version]
- Wirthmueller, L.; Roth, C.; Fabro, G.; Caillaud, M.C.; Rallapalli, G.; Asai, S.; Sklenar, J.; Jones, A.M.E.; Wiermer, M.; Jones, J.D.G.; et al. Probing formation of cargo/importin-α transport complexes in plant cells using a pathogen effector. Plant J. 2015, 81, 40–52. [Google Scholar] [CrossRef] [Green Version]
- Fanara, P.; Hodel, M.R.; Corbett, A.H.; Hodel, A.E. Quantitative analysis of nuclear localization signal (NLS)-importin alpha interaction through fluorescence depolarization. Evidence for auto-inhibitory regulation of NLS binding. J. Biol. Chem. 2000, 275, 21218–21223. [Google Scholar] [CrossRef] [Green Version]
- Harreman, M.T.; Hodel, M.R.; Fanara, P.; Hodel, A.E.; Corbett, A.H. The Auto-inhibitory Function of Importin α Is Essentialin Vivo. J. Biol. Chem. 2003, 278, 5854–5863. [Google Scholar] [CrossRef]
- Harreman, M.T.; Cohen, P.E.; Hodel, M.R.; Truscott, G.J.; Corbett, A.H.; Hodel, A.E. Characterization of the Auto-inhibitory Sequence within the N-terminal Domain of Importin alpha. J. Biol. Chem. 2003, 278, 21361–21369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jibiki, K.; Liu, M.; Lei, C.; Kodama, T.S.; Kojima, C.; Fujiwara, T.; Yasuhara, N. Biochemical propensity mapping for structural and functional anatomy of importin α IBB domain. Genes Cells 2022, 27, 173–191. [Google Scholar] [CrossRef] [PubMed]
- Oostdyk, L.T.; McConnell, M.J.; Paschal, B.M. Characterization of the Importin-β binding domain in nuclear import receptor KPNA7. Biochem. J. 2019, 476, 3413–3434. [Google Scholar] [CrossRef] [PubMed]
- Jibiki, K.; Kodama, T.S.; Suenaga, A.; Kawase, Y.; Shibazaki, N.; Nomoto, S.; Nagasawa, S.; Nagashima, M.; Shimodan, S.; Kikuchi, R.; et al. Importin α2 association with chromatin: Direct DNA binding via a novel DNA-binding domain. Genes Cells 2021, 26, 945–966. [Google Scholar] [CrossRef]
- Miyamoto, Y.; Yamada, K.; Yoneda, Y. Importin α: A key molecule in nuclear transport and non-transport functions. J. Biochem. 2016, 160, 69–75. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.W.; Couñago, R.L.; Williams, S.J.; Bodén, M.; Kobe, B. Crystal Structure of Rice Importin-α and Structural Basis of Its Interaction with Plant-Specific Nuclear Localization Signals. Plant Cell 2012, 24, 5074–5088. [Google Scholar] [CrossRef] [Green Version]
- Marfori, M.; Mynott, A.; Ellis, J.J.; Mehdi, A.M.; Saunders, N.F.; Curmi, P.M.; Forwood, J.K.; Bodén, M.; Kobe, B. Molecular basis for specificity of nuclear import and prediction of nuclear localization. Biochim. Biophys. Acta 2011, 1813, 1562–1577. [Google Scholar] [CrossRef]
- Lu, J.; Wu, T.; Zhang, B.; Liu, S.; Song, W.; Qiao, J.; Ruan, H. Types of nuclear localization signals and mechanisms of protein import into the nucleus. Cell Commun. Signal. 2021, 19, 60. [Google Scholar] [CrossRef]
- Kosugi, S.; Hasebe, M.; Matsumura, N.; Takashima, H.; Miyamoto-Sato, E.; Tomita, M.; Yanagawa, H. Six Classes of Nuclear Localization Signals Specific to Different Binding Grooves of Importin alpha. J. Biol. Chem. 2009, 284, 478–485. [Google Scholar] [CrossRef] [Green Version]
- He, S.; Huang, K.; Zhang, X.; Yu, X.; Huang, P.; An, C. The LSD1-Type Zinc Finger Motifs of Pisum sativa LSD1 Are a Novel Nuclear Localization Signal and Interact with Importin Alpha. PLoS ONE 2011, 6, e22131. [Google Scholar] [CrossRef]
- Adam, S.A.; Lobl, T.J.; Mitchell, M.A.; Gerace, L. Identification of specific binding proteins for a nuclear location sequence. Nature 1989, 337, 276–279. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.W.; Couñago, R.M.; Williams, S.J.; Bodèn, M.; Kobe, B. The distribution of different classes of nuclear localization signals (NLSs) in diverse organisms and the utilization of the minor NLS-binding site inplantnuclear import factor importin-α. Plant Signal. Behav. 2013, 8, e25976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fontes, M.R.; Teh, T.; Jans, D.; Brinkworth, R.I.; Kobe, B. Structural Basis for the Specificity of Bipartite Nuclear Localization Sequence Binding by Importin-alpha. J. Biol. Chem. 2003, 278, 27981–27987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helizon, H.; Rösler-Dalton, J.; Gasch, P.; Von Horsten, S.; Essen, L.-O.; Zeidler, M. Arabidopsis phytochrome A nuclear translocation is mediated by a far-red elongated hypocotyl 1-importin complex. Plant J. 2018, 96, 1255–1268. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Masi, R.; Lintermann, R.; Wirthmueller, L. Nuclear Import of Arabidopsis Poly(ADP-Ribose) Polymerase 2 Is Mediated by Importin-α and a Nuclear Localization Sequence Located Between the Predicted SAP Domains. Front. Plant Sci. 2018, 9, 1581. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Kim, D.; Yun, H.R.; Lee, Y.M.; Yogendra, B.; Bo, Z.; Kim, H.E.; Min, J.H.; Lee, Y.; Gil Rim, Y.; et al. Nuclear import of LIKE HETEROCHROMATIN PROTEIN1 is redundantly mediated by importins α-1, α-2 and α-3. Plant J. 2020, 103, 1205–1214. [Google Scholar] [CrossRef]
- Contreras, R.; Kallemi, P.; González-García, M.P.; Lazarova, A.; Sánchez-Serrano, J.J.; Sanmartín, M.; Rojo, E. Identification of Domains and Factors Involved in MINIYO Nuclear Import. Front. Plant Sci. 2019, 10, 1044. [Google Scholar] [CrossRef]
- Gerth, K.; Lin, F.; Daamen, F.; Menzel, W.; Heinrich, F.; Heilmann, M. Arabidopsis phosphatidylinositol 4-phosphate 5-kinase 2 contains a functional nuclear localization sequence and interacts with alpha-importins. Plant J. 2017, 92, 862–878. [Google Scholar] [CrossRef]
- Pumroy, R.A.; Cingolani, G. Diversification of importin-α isoforms in cellular trafficking and disease states. Biochem. J. 2015, 466, 13–28. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Wang, H.; Liu, J.; Han, S.; Lin, M.; Guo, Z.; Chen, X. OsWRKY62 and OsWRKY76 Interact with Importin α1s for Negative Regulation of Defensive Responses in Rice Nucleus. Rice 2022, 15, 12. [Google Scholar] [CrossRef]
- Jiang, C.J.; Shoji, K.; Matsuki, R.; Baba, A.; Inagaki, N.; Ban, H.; Iwasaki, T.; Imamoto, N.; Yoneda, Y.; Deng, X.W.; et al. Molecular Cloning of a Novel Importin α Homologue from Rice, by Which Constitutive Photomorphogenic 1 (COP1) Nuclear Localization Signal (NLS)-Protein Is Preferentially Nuclear Imported. J. Biol. Chem. 2001, 276, 9322–9329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganguly, A.; DeMott, L.; Zhu, C.; McClosky, D.D.; Anderson, C.T.; Dixit, R. Importin-β Directly Regulates the Motor Activity and Turnover of a Kinesin-4. Dev. Cell 2018, 44, 642–651.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziemienowicz, A.; Haasen, D.; Staiger, D.; Merkle, T. Arabidopsis transportin1 is the nuclear import receptor for the circadian clock-regulated RNA-binding protein AtGRP7. Plant Mol. Biol. 2003, 53, 201–212. [Google Scholar] [CrossRef]
- Ma, X.; Huang, X.; Jiao, Z.; He, L.; Li, Y.; Ow, D.W. Overproduction of plant nuclear export signals enhances diamide tolerance in Schizosaccharomyces pombe. Biochem. Biophys. Res. Commun. 2020, 531, 335–340. [Google Scholar] [CrossRef]
- Blanvillain, R.; Wei, S.; Wei, P.; Kim, J.H.; Ow, D.W. Stress tolerance to stress escape in plants: Role of the OXS2 zinc-finger transcription factor family. EMBO J. 2011, 30, 3812–3822. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Shimura, H.; Sueda, K.; Masuta, C. Importin/exportin-mediated nucleocytoplasmic shuttling of cucumber mosaic virus 2b protein is required for 2b’s efficient suppression of RNA silencing. PLoS Pathog. 2022, 18, e1010267. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhan, Q.; Shi, T.; Liu, J.; Zhao, K.; Gao, Y. The nuclear transporter SAD2 plays a role in calcium- and H2 O2 -mediated cell death in Arabidopsis. Plant J. 2020, 101, 324–333. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Guo, X.; Ge, C.; Ma, Z.; Jiang, M.; Li, T.; Koiwa, H.; Yang, S.W.; Zhang, X. KETCH1 imports HYL1 to nucleus for miRNA biogenesis in Arabidopsis. Proc. Natl. Acad. Sci. USA 2017, 114, 4011–4016. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.; Fang, X.; Qi, Y. TRANSPORTIN1 Promotes the Association of MicroRNA with ARGONAUTE1 in Arabidopsis. Plant Cell 2016, 28, 2576–2585. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Ye, R.; Xin, Y.; Fang, X.; Li, C.; Shi, H.; Zhou, X.; Qi, Y. An importin β protein negatively regulates MicroRNA activity in Arabidopsis. Plant Cell 2011, 23, 3565–3576. [Google Scholar] [CrossRef]
- Xu, S.; Zhang, Z.; Jing, B.; Gannon, P.; Ding, J.; Xu, F.; Li, X.; Zhang, Y. Transportin-SR is required for proper splicing of resistance genes and plant immunity. PLoS Genet. 2011, 7, e1002159. [Google Scholar] [CrossRef] [Green Version]
- Jia, M.; Shen, X.; Tang, Y.; Shi, X.; Gu, Y. A karyopherin constrains nuclear activity of the NLR protein SNC1 and is essential to prevent autoimmunity in Arabidopsis. Mol. Plant. 2021, 14, 1733–1744. [Google Scholar] [CrossRef] [PubMed]
- Petosa, C.; Schoehn, G.; Askjaer, P.; Bauer, U.; Moulin, M.; Steuerwald, U.; Soler-López, M.; Baudin, F.; Mattaj, I.W.; Müller, C.W. Architecture of CRM1/Exportin1 Suggests How Cooperativity Is Achieved during Formation of a Nuclear Export Complex. Mol. Cell 2004, 16, 761–775. [Google Scholar] [CrossRef] [PubMed]
- Monecke, T.; Haselbach, D.; Voß, B.; Russek, A.; Neumann, P.; Thomson, E.; Hurt, E.; Zachariae, U.; Stark, H.; Grubmüller, H.; et al. Structural basis for cooperativity of CRM1 export complex formation. Proc. Natl. Acad. Sci. USA 2013, 110, 960–965. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Gong, P.; Ge, L.; Chang, Z.; Cheng, X.; Zhou, X.; Wang, A.; Li, F. Nuclear exportin 1 facilitates turnip mosaic virus infection by exporting the sumoylated viral replicase and by repressing plant immunity. New Phytol. 2021, 232, 1382–1398. [Google Scholar] [CrossRef] [PubMed]
- Cook, A.; Fernandez, E.; Lindner, D.; Ebert, J.; Schlenstedt, G.; Conti, E. The Structure of the Nuclear Export Receptor Cse1 in Its Cytosolic State Reveals a Closed Conformation Incompatible with Cargo Binding. Mol. Cell 2005, 18, 355–367. [Google Scholar] [CrossRef] [Green Version]
- Leisegang, M.S.; Martin, R.; Ramírez, A.S.; Bohnsack, M.T. Exportin T and Exportin 5: tRNA and miRNA biogenesis—And beyond. Biol. Chem. 2012, 393, 599–604. [Google Scholar] [CrossRef]
- Cook, A.G.; Fukuhara, N.; Jinek, M.; Conti, E. Structures of the tRNA export factor in the nuclear and cytosolic states. Nature 2009, 461, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Okada, C.; Yamashita, E.; Lee, S.J.; Shibata, S.; Katahira, J.; Nakagawa, A.; Yoneda, Y.; Tsukihara, T. A High-Resolution Structure of the Pre-microRNA Nuclear Export Machinery. Science 2009, 326, 1275–1279. [Google Scholar] [CrossRef] [PubMed]
- Chook, Y.M.; Blobel, G. Structure of the nuclear transport complex karyopherin-beta2-Ran x GppNHp. Nature 1999, 399, 230–237. [Google Scholar] [CrossRef]
- Vetter, I.R.; Arndt, A.; Kutay, U.; Görlich, D.; Wittinghofer, A. Structural View of the Ran–Importin β Interaction at 2.3 Å Resolution. Cell 1999, 97, 635–646. [Google Scholar] [CrossRef] [Green Version]
- Aksu, M.; Trakhanov, S.; Görlich, D. Structure of the exportin Xpo4 in complex with RanGTP and the hypusine-containing translation factor eIF5A. Nat. Commun. 2016, 7, 11952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bollman, K.M.; Aukerman, M.J.; Park, M.Y.; Hunter, C.; Berardini, T.Z.; Poethig, R.S. HASTY, the Arabidopsis ortholog of exportin 5/MSN5, regulates phase change and morphogenesis. Development 2003, 130, 1493–1504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilsson, J.; Askjaer, P.; Kjems, J. A Role for the Basic Patch and the C Terminus of RanGTP in Regulating the Dynamic Interactions with Importin β, CRM1 and RanBP1. J. Mol. Biol. 2001, 305, 231–243. [Google Scholar] [CrossRef] [PubMed]
- Vetter, I.R.; Nowak, C.; Nishimoto, T.; Kuhlmann, J.; Wittinghofer, A. Structure of a Ran-binding domain complexed with Ran bound to a GTP analogue: Implications for nuclear transport. Nature 1999, 398, 39–46. [Google Scholar] [CrossRef]
- Kalita, J.; Kapinos, L.E.; Lim, R.Y.H. On the asymmetric partitioning of nucleocytoplasmic transport—Recent insights and open questions. J. Cell Sci. 2021, 134, jcs240382. [Google Scholar] [CrossRef]
- Soniat, M.; Chook, Y.M. Nuclear localization signals for four distinct karyopherin-β nuclear import systems. Biochem. J. 2015, 468, 353–362. [Google Scholar] [CrossRef]
- Süel, K.E.; Chook, Y.M. Kap104p Imports the PY-NLS-containing Transcription Factor Tfg2p into the Nucleus. J. Biol. Chem. 2009, 284, 15416–15424. [Google Scholar] [CrossRef] [Green Version]
- Mboukou, A.; Rajendra, V.; Kleinova, R.; Tisné, C.; Jantsch, M.F.; Barraud, P. Transportin-1: A Nuclear Import Receptor with Moonlighting Functions. Front. Mol. Biosci. 2021, 8, 638149. [Google Scholar] [CrossRef]
- Lee, B.J.; Cansizoglu, A.E.; Süel, K.E.; Louis, T.H.; Zhang, Z.; Chook, Y.M. Rules for Nuclear Localization Sequence Recognition by Karyopherin beta 2. Cell 2006, 126, 543–558. [Google Scholar] [CrossRef]
- Wing, C.E.; Fung, H.Y.J.; Chook, Y.M. Karyopherin-mediated nucleocytoplasmic transport. Nat. Rev. Mol. Cell Biol. 2022, 23, 307–328. [Google Scholar] [CrossRef] [PubMed]
- Lai, M.C.; Lin, R.I.; Huang, S.Y.; Tsai, C.W.; Tarn, W.Y. A Human Importin-β Family Protein, Transportin-SR2, Interacts with the Phosphorylated RS Domain of SR Proteins. J. Biol. Chem. 2000, 275, 7950–7957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haasen, D.; Köhler, C.; Neuhaus, G.; Merkle, T. Nuclear export of proteins in plants: AtXPO1 is the export receptor for leucine-rich nuclear export signals in Arabidopsis thaliana. Plant J. 1999, 20, 695–705. [Google Scholar] [CrossRef] [PubMed]
- Fung, H.Y.; Fu, S.C.; Chook, Y.M. Nuclear export receptor CRM1 recognizes diverse conformations in nuclear export signals. eLife 2017, 6, e23961. [Google Scholar] [CrossRef] [PubMed]
- Fung, H.Y.; Fu, S.C.; Brautigam, C.A.; Chook, Y.M. Structural determinants of nuclear export signal orientation in binding to exportin CRM1. eLife 2015, 4, e10034. [Google Scholar] [CrossRef]
- Xu, D.; Farmer, A.; Chook, Y.M. Recognition of nuclear targeting signals by Karyopherin-β proteins. Curr. Opin. Struct. Biol. 2010, 20, 782–790. [Google Scholar] [CrossRef] [Green Version]
- Verma, V.; Ravindran, P.; Kumar, P.P. Plant hormone-mediated regulation of stress responses. BMC Plant Biol. 2016, 16, 86. [Google Scholar] [CrossRef] [Green Version]
- Aida, M.; Beis, D.; Heidstra, R.; Willemsen, V.; Blilou, I.; Galinha, C.; Nussaume, L.; Noh, Y.S.; Amasino, R.; Scheres, B. The PLETHORA Genes Mediate Patterning of the Arabidopsis Root Stem Cell Niche. Cell 2004, 119, 109–120. [Google Scholar] [CrossRef] [Green Version]
- Mähönen, A.P.; Ten Tusscher, K.; Siligato, R.; Smetana, O.; Díaz-Triviño, S.; Salojärvi, J.; Wachsman, G.; Prasad, K.; Heidstra, R.; Scheres, B. PLETHORA gradient formation mechanism separates auxin responses. Nature 2014, 515, 125–129. [Google Scholar] [CrossRef] [Green Version]
- Xiong, F.; Zhang, B.K.; Liu, H.H.; Wei, G.; Wu, J.H.; Wu, Y.N.; Zhang, Y.; Li, S. Transcriptional Regulation of PLETHORA1 in the Root Meristem Through an Importin and Its Two Antagonistic Cargos. Plant Cell 2020, 32, 3812–3824. [Google Scholar] [CrossRef]
- Yang, W.; Cortijo, S.; Korsbo, N.; Roszak, P.; Schiessl, K.; Gurzadyan, A.; Wightman, R.; Jönsson, H.; Meyerowitz, E. Molecular mechanism of cytokinin-activated cell division in Arabidopsis. Science 2021, 371, 1350–1355. [Google Scholar] [CrossRef] [PubMed]
- Verslues, P.E.; Guo, Y.; Dong, C.H.; Ma, W.; Zhu, J.K. Mutation of SAD2, an importin β-domain protein in Arabidopsis, alters abscisic acid sensitivity. Plant J. 2006, 47, 776–787. [Google Scholar] [CrossRef] [PubMed]
- Oh, T.R.; Yu, S.G.; Yang, H.W.; Kim, J.H.; Kim, W.T. AtKPNB1, an Arabidopsis importin-β protein, is downstream of the RING E3 ubiquitin ligase AtAIRP1 in the ABA-mediated drought stress response. Planta 2020, 252, 93. [Google Scholar] [CrossRef]
- Xu, X.; Wan, W.; Jiang, G.; Xi, Y.; Huang, H.; Cai, J.; Chang, Y.; Duan, C.G.; Mangrauthia, S.K.; Peng, X.; et al. Nucleocytoplasmic Trafficking of the Arabidopsis WD40 Repeat Protein XIW1 Regulates ABI5 Stability and Abscisic Acid Responses. Mol. Plant 2019, 12, 1598–1611. [Google Scholar] [CrossRef]
- McWhite, C.D.; Papoulas, O.; Drew, K.; Cox, R.M.; June, V.; Dong, O.X.; Kwon, T.; Wan, C.; Salmi, M.L.; Roux, S.J.; et al. A Pan-plant Protein Complex Map Reveals Deep Conservation and Novel Assemblies. Cell 2020, 181, 460–474.e14. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING database in 2021: Customizable protein–protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Yang, H.; Shang, C.; Ma, S.; Liu, L.; Cheng, J. The Roles of Auxin Biosynthesis YUCCA Gene Family in Plants. Int. J. Mol. Sci. 2019, 20, 6343. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Prunet, N.; Gan, E.S.; Wang, Y.; Stewart, D.; Wellmer, F.; Huang, J.; Yamaguchi, N.; Tatsumi, Y.; Kojima, M.; et al. SUPERMAN regulates floral whorl boundaries through control of auxin biosynthesis. EMBO J. 2018, 37, e97499. [Google Scholar] [CrossRef]
- Rizzardi, K.; Landberg, K.; Nilsson, L.; Ljung, K.; Sundås-Larsson, A. TFL2/LHP1 is involved in auxin biosynthesis through positive regulation of YUCCA genes. Plant J. 2011, 65, 897–906. [Google Scholar] [CrossRef]
- Yamaguchi, N.; Huang, J.; Tatsumi, Y.; Abe, M.; Sugano, S.S.; Kojima, M.; Takebayashi, Y.; Kiba, T.; Yokoyama, R.; Nishitani, K.; et al. Chromatin-mediated feed-forward auxin biosynthesis in floral meristem determinacy. Nat. Commun. 2018, 9, 5290. [Google Scholar] [CrossRef]
- Kriechbaumer, V.; Wang, P.; Hawes, C.; Abell, B.M. Alternative splicing of the auxin biosynthesis gene YUCCA4 determines its subcellular compartmentation. Plant J. 2012, 70, 292–302. [Google Scholar] [CrossRef]
- Tsugeki, R.; Tanaka-Sato, N.; Maruyama, N.; Terada, S.; Kojima, M.; Sakakibara, H.; Okada, K. CLUMSY VEIN, the Arabidopsis DEAH-box Prp16 ortholog, is required for auxin-mediated development. Plant J. 2015, 81, 183–197. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.J.; Luo, J. The PIN-FORMED Auxin Efflux Carriers in Plants. Int. J. Mol. Sci. 2018, 19, 2759. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.; Ma, J.; Wu, D.; Shen, W.H.; Zhu, Y. Functional Coordination of the Chromatin-Remodeling Factor AtINO80 and the Histone Chaperones NRP1/2 in Inflorescence Meristem and Root Apical Meristem. Front. Plant Sci. 2019, 10, 115. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Zheng, B.; Yu, Y.; Won, S.Y.; Mo, B.; Chen, X. The role of Mediator in small and long noncoding RNA production inArabidopsis thaliana. EMBO J. 2011, 30, 814–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raya-González, J.; Ávalos-Rangel, A.; Ruiz-Herrera, L.F.; Valdez-Alarcón, J.J.; López-Bucio, J. The RNA polymerase II subunit NRPB2 is required for indeterminate root development, cell viability, stem cell niche maintenance, and de novo root tip regeneration in Arabidopsis. Protoplasma 2022, 259, 1175–1188. [Google Scholar] [CrossRef]
- Terrile, M.C.; Tebez, N.M.; Colman, S.L.; Mateos, J.L.; Morato-López, E.; Sánchez-López, N.; Izquierdo-Álvarez, A.; Marina, A.; Calderón Villalobos, L.I.A.; Estelle, M.; et al. S-Nitrosation of E3 Ubiquitin Ligase Complex Components Regulates Hormonal Signalings in Arabidopsis. Front. Plant Sci. 2022, 12, 794582. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Zhang, F.; Friml, J.; Ding, Z. Auxin signaling: Research advances over the past 30 years. J. Integr. Plant Biol. 2022, 64, 371–392. [Google Scholar] [CrossRef]
- Zhu, G.; Chang, Y.; Xu, X.; Tang, K.; Chen, C.; Lei, M.; Zhu, J.K.; Duan, C.G. EXPORTIN 1A prevents transgene silencing in Arabidopsis by modulating nucleo-cytoplasmic partitioning of HDA6. J. Integr. Plant Biol. 2019, 61, 1243–1254. [Google Scholar] [CrossRef]
- Yuan, L.; Chen, X.; Chen, H.; Wu, K.; Huang, S. Histone deacetylases HDA6 and HDA9 coordinately regulate valve cell elongation through affecting auxin signaling in Arabidopsis. Biochem. Biophys. Res. Commun. 2019, 508, 695–700. [Google Scholar] [CrossRef]
- Forestan, C.; Farinati, S.; Rouster, J.; Lassagne, H.; Lauria, M.; Dal Ferro, N.; Varotto, S. Control of Maize Vegetative and Reproductive Development, Fertility, and rRNAs Silencing by HISTONE DEACETYLASE 108. Genetics 2018, 208, 1443–1466. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, E.; Mano, S.; Nomoto, M.; Tada, Y.; Hara-Nishimura, I.; Nishimura, M.; Yamada, K. HSP90 Stabilizes Auxin-Responsive Phenotypes by Masking a Mutation in the Auxin Receptor TIR1. Plant Cell Physiol. 2016, 57, 2245–2254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, E.; Mano, S.; Hara-Nishimura, I.; Nishimura, M.; Yamada, K. HSP90 stabilizes auxin receptor TIR1 and ensures plasticity of auxin responses. Plant Signal. Behav. 2017, 12, e1311439. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Zhang, Y.; Kieffer, M.; Yu, H.; Kepinski, S.; Estelle, M. HSP90 regulates temperature-dependent seedling growth in Arabidopsis by stabilizing the auxin co-receptor F-box protein TIR1. Nat. Commun. 2016, 7, 10269. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Ito, H.; Quint, M.; Huang, H.; Noël, L.D.; Gray, W.M. Genetic analysis of CAND1–CUL1 interactions in Arabidopsis supports a role for CAND1-mediated cycling of the SCFTIR1 complex. Proc. Natl. Acad. Sci. USA 2008, 105, 8470–8475. [Google Scholar] [CrossRef] [Green Version]
- Qin, N.; Xu, D.; Li, J.; Deng, X.W. COP9 signalosome: Discovery, conservation, activity, and function. J. Integr. Plant Biol. 2020, 62, 90–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dohmann, E.M.; Levesque, M.P.; Isono, E.; Schmid, M.; Schwechheimer, C. Auxin Responses in Mutants of the Arabidopsis CONSTITUTIVE PHOTOMORPHOGENIC9 Signalosome. Plant Physiol. 2008, 147, 1369–1379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pacurar, D.I.; Pacurar, M.L.; Lakehal, A.; Pacurar, A.M.; Ranjan, A.; Bellini, C. The Arabidopsis Cop9 signalosome subunit 4 (CNS4) is involved in adventitious root formation. Sci. Rep. 2017, 7, 628. [Google Scholar] [CrossRef] [Green Version]
- Majumdar, P.; Karidas, P.; Siddiqi, I.; Nath, U. The Ubiquitin-Specific Protease TNI/UBP14 Functions in Ubiquitin Recycling and Affects Auxin Response. Plant Physiol. 2020, 184, 1499–1513. [Google Scholar] [CrossRef]
- Majumdar, P.; Karidas, P.; Nath, U. The TARANI/UBIQUITIN PROTEASE 14 protein is required for lateral root development in Arabidopsis. Plant Signal. Behav. 2021, 16, 1860386. [Google Scholar] [CrossRef]
- David, R.; Burgess, A.; Parker, B.; Li, J.; Pulsford, K.; Sibbritt, T.; Preiss, T.; Searle, I.R. Transcriptome-Wide Mapping of RNA 5-Methylcytosine in Arabidopsis mRNAs and Noncoding RNAs. Plant Cell 2017, 29, 445–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, X.; Liang, Z.; Shen, L.; Zhang, Q.; Bao, S.; Geng, Y.; Zhang, B.; Leo, V.; Vardy, L.A.; Lu, T.; et al. 5-Methylcytosine RNA Methylation in Arabidopsis Thaliana. Mol. Plant 2017, 10, 1387–1399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goh, T.; Joi, S.; Mimura, T.; Fukaki, H. The establishment of asymmetry in Arabidopsis lateral root founder cells is regulated by LBD16/ASL18 and related LBD/ASL proteins. Development 2012, 139, 883–893. [Google Scholar] [CrossRef] [Green Version]
- Ötvös, K.; Miskolczi, P.; Marhavý, P.; Cruz-Ramírez, A.; Benková, E.; Robert, S.; Bakó, L. Pickle Recruits Retinoblastoma Related 1 to Control Lateral Root Formation in Arabidopsis. Int. J. Mol. Sci. 2021, 22, 3862. [Google Scholar] [CrossRef]
- Flores-Pérez, U.; Pérez-Gil, J.; Closa, M.; Wright, L.P.; Botella-Pavía, P.; Phillips, M.A.; Ferrer, A.; Gershenzon, J.; Rodríguez-Concepción, M. PLEIOTROPIC REGULATORY LOCUS 1 (PRL1) Integrates the Regulation of Sugar Responses with Isoprenoid Metabolism in Arabidopsis. Mol. Plant 2010, 3, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.; Wang, S.; Li, K.; Szakonyi, D.; Koncz, C.; Li, X. PRL1 modulates root stem cell niche activity and meristem size through WOX5 and PLTs in Arabidopsis. Plant J. 2015, 81, 399–412. [Google Scholar] [CrossRef]
- Xiang, D.; Yang, H.; Venglat, P.; Cao, Y.; Wen, R.; Ren, M.; Stone, S.; Wang, E.; Wang, H.; Xiao, W.; et al. POPCORN Functions in the Auxin Pathway to Regulate Embryonic Body Plan and Meristem Organization in Arabidopsis. Plant Cell 2011, 23, 4348–4367. [Google Scholar] [CrossRef] [Green Version]
- Gallois, J.L.; Guyon-Debast, A.; Lécureuil, A.; Vezon, D.; Carpentier, V.; Bonhomme, S.; Guerche, P. The Arabidopsis Proteasome RPT5 Subunits Are Essential for Gametophyte Development and Show Accession-Dependent Redundancy. Plant Cell 2009, 21, 442–459. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, T.; Tsujimoto-Inui, Y.; Sotta, N.; Hirakawa, T.; Matsunaga, T.M.; Fukao, Y.; Matsunaga, S.; Fujiwara, T. Proteasomal degradation of BRAHMA promotes Boron tolerance in Arabidopsis. Nat. Commun. 2018, 9, 5285. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, T.; Sotta, N.; Suzuki, T.; Fujiwara, T.; Matsunaga, S. The 26S Proteasome Is Required for the Maintenance of Root Apical Meristem by Modulating Auxin and Cytokinin Responses Under High-Boron Stress. Front. Plant Sci. 2019, 10, 590. [Google Scholar] [CrossRef]
- Stelpflug, S.C.; Sekhon, R.S.; Vaillancourt, B.; Hirsch, C.N.; Buell, C.R.; De Leon, N.; Kaeppler, S.M. An Expanded Maize Gene Expression Atlas based on RNA Sequencing and its Use to Explore Root Development. Plant Genome 2016, 9, 1. [Google Scholar] [CrossRef]
- Qian, Y.; Ren, Q.; Zhang, J.; Chen, L. Transcriptomic analysis of the maize (Zea mays L.) inbred line B73 response to heat stress at the seedling stage. Gene 2019, 692, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.F.; He, F.F.; Ma, X.X.; Mao, C.Z.; Hodgman, C.; Lu, C.G.; Wu, P. OsCAND1 Is Required for Crown Root Emergence in Rice. Mol. Plant 2011, 4, 289–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbasi, N.; Kim, H.B.; Park, N.I.; Kim, H.S.; Kim, Y.K.; Park, Y.I.; Choi, S.B. APUM23, a nucleolar Puf domain protein, is involved in pre-ribosomal RNA processing and normal growth patterning in Arabidopsis. Plant J. 2010, 64, 960–976. [Google Scholar] [CrossRef] [PubMed]
- Oka, M.; Yoneda, Y. Importin α: Functions as a nuclear transport factor and beyond. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2018, 94, 259–274. [Google Scholar] [CrossRef] [Green Version]
- Brioudes, F.; Jay, F.; Sarazin, A.; Grentzinger, T.; A Devers, E.; Voinnet, O. HASTY, the Arabidopsis EXPORTIN5 ortholog, regulates cell-to-cell and vascular microRNA movement. EMBO J. 2021, 40, e107455. [Google Scholar] [CrossRef]
- Parween, D.; Sahu, B.B. An Arabidopsis nonhost resistance gene, IMPORTIN ALPHA 2 provides immunity against rice sheath blight pathogen, Rhizoctonia solani. Curr. Res. Microb. Sci. 2022, 3, 100109. [Google Scholar] [CrossRef]
- Lüdke, D.; Roth, C.; Kamrad, S.A.; Messerschmidt, J.; Hartken, D.; Appel, J.; Hörnich, B.F.; Yan, Q.; Kusch, S.; Klenke, M.; et al. Functional requirement of the Arabidopsis importin-α nuclear transport receptor family in autoimmunity mediated by the NLR protein SNC1. Plant J. 2021, 105, 994–1009. [Google Scholar] [CrossRef]
- Bhattacharjee, S.; Lee, L.Y.; Oltmanns, H.; Cao, H.; Veena; Cuperus, J.; Gelvin, S.B. IMPa-4, an Arabidopsis Importin α Isoform, Is Preferentially Involved in Agrobacterium-Mediated Plant Transformation. Plant Cell 2008, 20, 2661–2680. [Google Scholar] [CrossRef] [Green Version]
- Xiong, F.; Duan, C.Y.; Liu, H.H.; Wu, J.H.; Zhang, Z.H.; Li, S.; Zhang, Y. Arabidopsis KETCH1 Is Critical for the Nuclear Accumulation of Ribosomal Proteins and Gametogenesis. Plant Cell 2020, 32, 1270–1284. [Google Scholar] [CrossRef]
- Liu, H.H.; Xiong, F.; Duan, C.Y.; Wu, Y.N.; Zhang, Y.; Li, S. Importin β4 Mediates Nuclear Import of GRF-Interacting Factors to Control Ovule Development in Arabidopsis. Plant Physiol. 2019, 179, 1080–1092. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Zhang, W.; Zhao, Y.; Gong, X.; Guo, L.; Zhu, G.; Wang, X.; Gong, Z.; Schumaker, K.S.; Guo, Y. SAD2, an Importin β-Like Protein, Is Required for UV-B Response in Arabidopsis by Mediating MYB4 Nuclear Trafficking. Plant Cell 2007, 19, 3805–3818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Gong, X.; Cao, W.; Zhao, J.; Fu, L.; Wang, X.; Schumaker, K.S.; Guo, Y. SAD2 in Arabidopsis Functions in Trichome Initiation through Mediating GL3 Function and Regulating GL1, TTG1 and GL2 Expression. J. Integr. Plant Biol. 2008, 50, 906–917. [Google Scholar] [CrossRef] [PubMed]
- Panda, C.; Li, X.; Wager, A.; Chen, H.; Li, X. An importin-beta-like protein mediates lignin-modification-induced dwarfism in Arabidopsis. Plant J. 2020, 102, 1281–1293. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.J.; Wang, L.C.; Yeh, C.H.; Lu, C.A.; Wu, S.J. Isolation and characterization of the Arabidopsis heat-intolerant 2 (hit2) mutant reveal the essential role of the nuclear export receptor EXPORTIN1A (XPO1A) in plant heat tolerance. New Phytol. 2010, 186, 833–842. [Google Scholar] [CrossRef]
- Blanvillain, R.; Boavida, L.C.; McCormick, S.; Ow, D.W. EXPORTIN1 Genes Are Essential for Development and Function of the Gametophytes in Arabidopsis thaliana. Genetics 2008, 180, 1493–1500. [Google Scholar] [CrossRef] [Green Version]
- Hunter, C.A.; Aukerman, M.J.; Sun, H.; Fokina, M.; Poethig, R.S. PAUSED Encodes the Arabidopsis Exportin-t Ortholog. Plant Physiol. 2003, 132, 2135–2143. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Chen, X. PAUSED, a Putative Exportin-t, Acts Pleiotropically in Arabidopsis Development But Is Dispensable for Viability. Plant Physiol. 2003, 132, 1913–1924. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.; Jia, M.; Li, X.; Tang, Y.; Jiang, K.; Bao, J.; Gu, Y. Exportin-4 coordinates nuclear shuttling of TOPLESS family transcription corepressors to regulate plant immunity. Plant Cell 2021, 33, 697–713. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name a | Locus ID b | Chromosomal Location c | Transcript ID | Putative Proteins d | |||||
|---|---|---|---|---|---|---|---|---|---|
| Chr | Chr_start | Chr_end | Length (aa) | MW (kDa) | Subcellular Location | ||||
| IMPα | ZmIMPα1 | Zm00001d008345 | 8 | 5938159 | 5944491 (−) | T001 | 527 | 57.85 | Nucleus/Cytoplasm |
| ZmIMPα2 | Zm00001d040274 | 3 | 35350411 | 35356233 (+) | T001 | 529 | 57.95 | Nucleus/Cytoplasm | |
| ZmIMPα3 | Zm00001d037606 | 6 | 131468248 | 85071126 (−) | T001 | 528 | 58.13 | Nucleus/Cytoplasm | |
| ZmIMPα4 | Zm00001d009850 | 8 | 85065908 | 131476305 (−) | T005 | 529 | 58.20 | Nucleus/Cytoplasm | |
| ZmIMPα5 | Zm00001d040153 | 3 | 29316628 | 29318539 (+) | T004 | 183 | 20.42 | Nucleus/Cytoplasm | |
| ZmIMPα6 | Zm00001d022536 | 7 | 179671127 | 179674969 (+) | T008 | 568 | 61.71 | Nucleus | |
| ZmIMPα7 | Zm00001d008640 | 8 | 15537598 | 15544131 (+) | T002 | 526 | 56.54 | Nucleus/Cytoplasm | |
| IMPβ | ZmIMB1a | Zm00001d030694 | 1 | 153742904 | 153749377 (+) | T002 | 1074 | 116.51 | Nucleus/Cytoplasm |
| ZmIMB1b | Zm00001d041556 | 3 | 127112005 | 127118515 (−) | T002 | 987 | 107.94 | Nucleus/Cytoplasm | |
| ZmIMB1c | Zm00001d038021 | 6 | 145393970 | 145398983 (−) | T001 | 879 | 96.77 | Nucleus/Cytoplasm | |
| ZmIMB1d | Zm00001d010512 | 8 | 118588081 | 118591573 (−) | T001 | 876 | 96.15 | Nucleus/Cytoplasm | |
| ZmIMB2a | Zm00001d002936 | 2 | 27303853 | 27322287 (+) | T010 | 891 | 98.80 | Nucleus/Cytoplasm | |
| ZmIMB2b | Zm00001d026696 | 10 | 150180000 | 150204009 (+) | T005 | 890 | 98.86 | Nucleus/Cytoplasm | |
| ZmIMB3a | Zm00001d021893 | 7 | 165532355 | 165542662 (−) | T002 | 1126 | 123.26 | Nucleus/Cytoplasm | |
| ZmIMB3b | Zm00001d033632 | 1 | 269308497 | 269321829 (+) | T008 | 1132 | 123.78 | Nucleus/Cytoplasm | |
| ZmIMB4 | Zm00001d028511 | 1 | 37580598 | 37594480 (−) | T008 | 1047 | 114.94 | Nucleus/Cytoplasm | |
| ZmIMB5 | Zm00001d045725 | 9 | 35215354 | 35239296 (−) | T001 | 1028 | 113.35 | Nucleus envelope/Cytosol | |
| ZmIPO8a | Zm00001d050526 | 4 | 96801958 | 96828810 (+) | T008 | 1145 | 128.01 | Nucleus envelope/Cytosol | |
| ZmIPO8b | Zm00001d016479 | 5 | 164246013 | 164266562 (+) | T001 | 1036 | 131.22 | Nucleus envelope/Cytosol | |
| ZmKA120 | Zm00001d007225 | 2 | 225128676 | 225140701 (+) | T019 | 1115 | 116.49 | Nucleus/Cytoplasm | |
| ZmXPO1a | Zm00001d012815 | 5 | 776419 | 787466 (+) | T022 | 1151 | 132.30 | Nucleus envelope/Cytosol | |
| ZmXPO1b | Zm00001d034914 | 1 | 305341236 | 305352529 (−) | T037 | 1122 | 128.54 | Nucleus envelope/Cytosol | |
| ZmXPO2a | Zm00001d033764 | 1 | 272997605 | 273005246 (+) | T002 | 981 | 108.32 | Nucleus/Cytoplasm | |
| ZmXPO2b | Zm00001d013417 | 5 | 10817793 | 10829003 (+) | T005 | 982 | 108.52 | Nucleus/Cytoplasm | |
| ZmXPOTa | Zm00001d022125 | 7 | 170837895 | 170846340 (+) | T002 | 978 | 107.96 | Nucleus/Cytoplasm | |
| ZmXPOTb | Zm00001d006845 | 2 | 217770387 | 217778785 (+) | T005 | 1024 | 113.22 | Nucleus/Cytoplasm | |
| ZmXPO4 | Zm00001d032704 | 1 | 235324863 | 235346931 (−) | T036 | 1165 | 129.84 | Nucleus/Cytoplasm | |
| ZmXPO5 | Zm00001d009270 | 8 | 49685540 | 49721525 (+) | T001 | 1175 | 130.20 | Nucleus/Cytoplasm | |
| ZmXPO7 | Zm00001d037100 | 6 | 112267718 | 112290870 (+) | T051 | 1067 | 121.03 | Nucleus/Cytoplasm | |
| ZmTNPO3a | Zm00001d052632 | 4 | 195421971 | 195454136 (+) | T005 | 1038 | 114.09 | Cytoplasm | |
| ZmTNPO3b | Zm00001d014033 | 5 | 29806869 | 29825510 (−) | T001 | 564 | 62.27 | Cytoplasm | |
| ZmTNPO3c | Zm00001d032699 | 1 | 235073263 | 235106928 (−) | T030 | 981 | 109.64 | Cytoplasm | |
| ZmPLANTKAPa | Zm00001d048628 | 4 | 1742938 | 1750365 (+) | T001 | 1092 | 120.63 | Nucleus envelope/Cytosol | |
| ZmPLANTKAPb | Zm00001d019335 | 7 | 28400881 | 28407780 (−) | T004 | 655 | 73.54 | Nucleus envelope/Cytosol | |
| Type | Consensus Motifs | Cargo | Sequence | NTR | Source |
|---|---|---|---|---|---|
| MP-cNLS | Class I— KR (K/R) R or K (K/R) RK | AtFHY1/AtFHL | 40KKRK | AtIMPA1 | Arabidopsis [74] |
| AtPARP2 | 48KRKR | AtIMPA2 | Arabidopsis [75] | ||
| AtLHP1 | 173RKRKRK | AtIMPA1/2/3 | Arabidopsis [76] | ||
| AtMINIYO | 253KLKKRRK | AtIMPA4 | Arabidopsis [77] | ||
| Class II— (P/R) XXKR (^DE) (K/R) | AtVRN1 | 173PTPTPKIPKKRGRKKKNADPE | AtIMPA1/2/3 | Arabidopsis [76] | |
| Class III— KRX (W/F/Y) XXAF | AtPIP5K2 | 239ATRKRSSVDSGAGSLTGEKIFPRIC | AtIMPA6/9 | Arabidopsis [78] | |
| Class IV— (R/P) XXKR (K/R) (^DE) | – | – | – | – | |
| Class V— LGKR (K/R) (W/F/Y) | VQ-protein | 92LGLGKRKRGPGVSGGKQTKRRSR | AtIMPA1/2/3 | Arabidopsis [76] | |
| BP-cNLS | Class VI— KRX10–12K(KR) (KR) or KRX10–12K(KR) X (K/R) | AtMINIYO | 1401RKR–1414RYKK, | AtIMPA4 | Arabidopsis [77] |
| OsWRKY62/OsWRKY76 | 8RK–36KKK | OsIMPα1 | Oryza Sativa [80] | ||
| OsCOP1 | 294RKKR–312KRR | OsIMPα1b | Oryza sativa [81] | ||
| ZmOpaque2 | 230RKRK–241RRSRYRK | OsIMPα1b ZmIMPα4 | Oryza sativa [81], Zea mays [16] | ||
| PY-NLS | (basic/hydrophobic) Xn— (R/H/K) (X)2–5 PY | AtFRA1 | 311KKRK–320PY | AtIMB4 | Arabidopsis [82] |
| M9-like domain | AtGRP7 | 112SGGGGSYGGGGGRREGGGGYSG | AtTRN1 | Arabidopsis [83] | |
| Other NLS | Zinc finger motifs | PsLSD1 | 7CNGCRNMLLYPRGATNVCCALC– 46CGGCRTLLMYTRGATSVRCSCC– 84CANCRTTLMYPYGAPSVKCAVC | AtIMPA1 | Pisum sativa [70] |
| NES | Φ-X2–3-Φ-X2–3-Φ-X-Φ | OXS2 | 699LEAWIEQMQL/LGALLEQMQL | - | Arabidopsis [84,85] |
| AtFHY1 | 54LLPL | AtXPO1 | Arabidopsis [74] | ||
| OsWRKY62 | 308VDQIPHIPV | AtXPO1 | Oryza Sativa [80] | ||
| CMV 2b | 79L-85L-87L | AtXPO1 | Mosaic Virus [86] |
| NTR | Putative Interactor in Maize | Interactive Score | Ortholog of the Putative Interactor in Arabidopsis | |
|---|---|---|---|---|
| Name | Gene ID | |||
| ZmIMPα1/2/3/4 (P, S) | Zm00001d009312 | P-0.208, S-0.582 | CHR11 | AT3G06400 |
| Zm00001d040831 | CHR17 | AT5G18620 | ||
| ZmIMPα1/2/3/4 (S) | Zm00001d014449 | S-0.781 | LHP1 | AT5G17690 |
| ZmIMPα1/2/3/4 (P) | Zm00001d050874 | P-0.242 | NRP1 | AT5G17690 |
| Zm00001d016935 | NRP2 | AT1G74560 | ||
| ZmIMB1c/d (P) | Zm00001d033218 | P-0.333 | NRPB2 | AT4G21710 |
| Zm00001d013683 | ||||
| ZmIMPα1/2 (P) | Zm00001d020898 | P-0.631 | HSP90.2 | AT5G56030 |
| Zm00001d031332 | ||||
| ZmIMPα1/2 (P), ZmXPO1 (S) | Zm00001d053813 | P-0.208, S-0.582 | CAND1 | AT2G02560 |
| ZmIMPα1/2/3/4 (P), ZmIMB1 (P) | Zm00001d028143 | P-0.243, P-0.363 | CSN4 | AT5G42970 |
| ZmIMPα1/2 (P), ZmIMB3 (P) | Zm00001d008743 | P-0.299, P-0.255 | UBP14 | AT3G20630 |
| ZmIMPα1/2/3/4 (P), ZmIMB1c/d (P) | Zm00001d045109 | P-0.299, P-0.303 | PKL | AT2G25170 |
| ZmIMPα4 (S) | Zm00001d033309 | S-0.421 | PRL1 | AT4G15900 |
| ZmIMB3, ZmIMB4 (S), ZmIPO8 (S), ZmXPOT (S) | Zm00001d020810 | S-0.716, S-0.640, S-0.505 | TRM4B | AT2G22400 |
| ZmIMPα1/2/3/4 (P), ZmXPO1/5 (S) | Zm00001d050139 | P-0.270, S-0.805 | HDA6 | AT5G63110 |
| ZmIMB3, ZmIMB4 (S) | Zm00001d013330 | S-0.639 | PCN | AT4G07410 |
| Zm00001d033912 | ||||
| ZmIPO8 (S) | Zm00001d006459 | S-0.655 | PRP16 | AT5G13010 |
| ZmIMB3, ZmIMB4 (S) | Zm00001d030554 | S-0.489 | APUM23 | AT1G72320 |
| ZmIMPα1/2/3/4 (P) | Zm00001d037481 | P-0.231 | RPT5A | AT3G05530 |
| Zm00001d018409 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, L.; Zhang, G.; Yang, G.; Dong, J. Identification of the Karyopherin Superfamily in Maize and Its Functional Cues in Plant Development. Int. J. Mol. Sci. 2022, 23, 14103. https://doi.org/10.3390/ijms232214103
Jin L, Zhang G, Yang G, Dong J. Identification of the Karyopherin Superfamily in Maize and Its Functional Cues in Plant Development. International Journal of Molecular Sciences. 2022; 23(22):14103. https://doi.org/10.3390/ijms232214103
Chicago/Turabian StyleJin, Lu, Guobin Zhang, Guixiao Yang, and Jiaqiang Dong. 2022. "Identification of the Karyopherin Superfamily in Maize and Its Functional Cues in Plant Development" International Journal of Molecular Sciences 23, no. 22: 14103. https://doi.org/10.3390/ijms232214103