
Neuroprotection and Non-Invasive Brain Stimulation: Facts or Fiction?
 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. NIBS and Neuroprotection in Parkinson’s Disease
2.1. tDCS
2.2. rTMS
3. NIBS and Neuroprotection in Alzheimer’s Disease
4. Beyond Neurodegenerative Disorders: Neuroprotection and Stroke
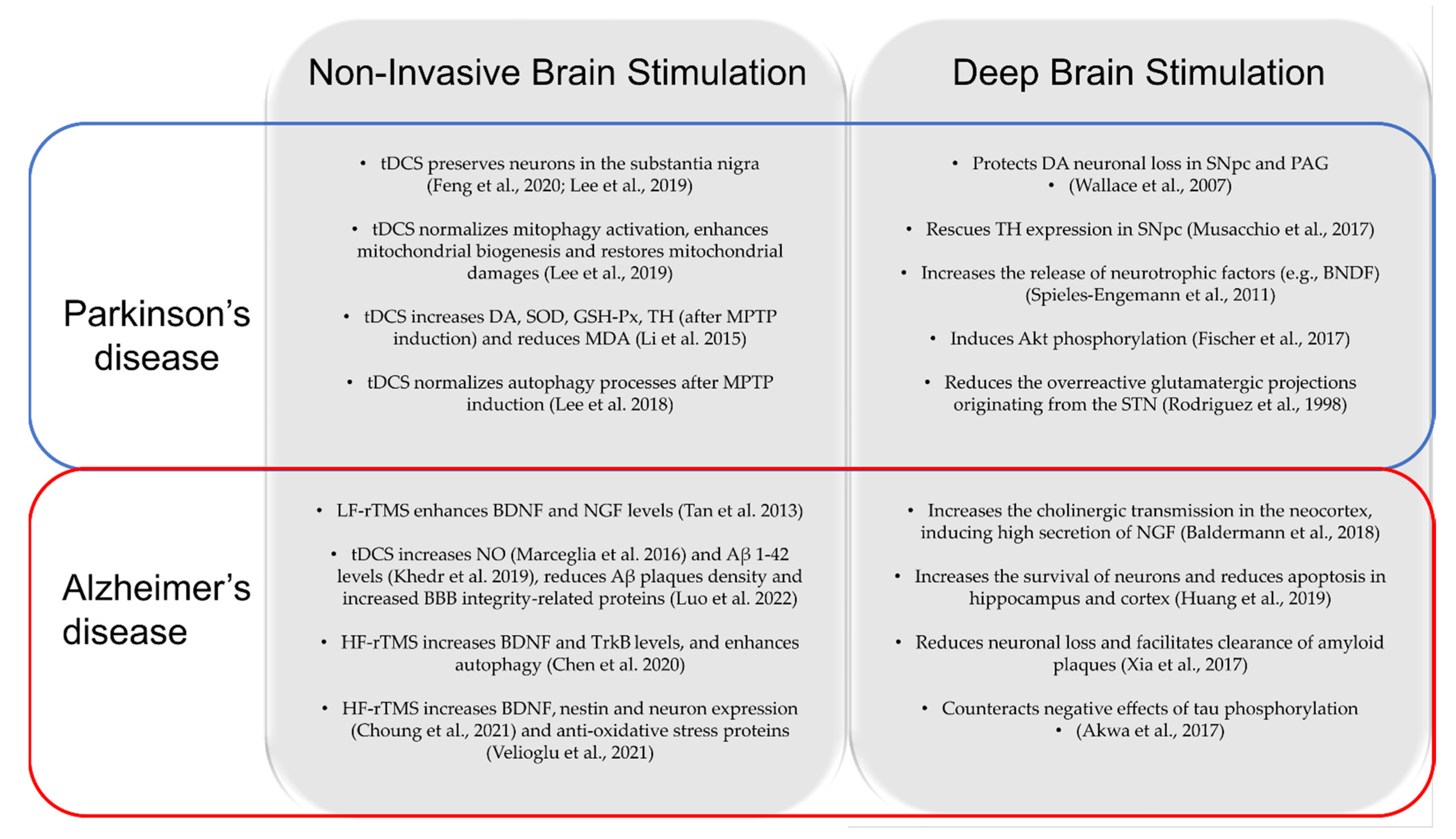
5. Differences and Similarities in Neuroprotective Mechanisms between NIBS and DBS
5.1. DBS and NIBS: Neuroprotection in PD
5.2. DBS and NIBS: Neuroprotection in AD
6. Limitations and Future Directions
7. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ferrucci, R.; Mameli, F.; Guidi, I.; Mrakic-Sposta, S.; Vergari, M.; Marceglia, S.; Cogiamanian, F.; Barbieri, S.; Scarpini, E.; Priori, A. Transcranial Direct Current Stimulation Improves Recognition Memory in Alzheimer Disease. Neurology 2008, 71, 493–498. [Google Scholar] [CrossRef] [Green Version]
- Lefaucheur, J.P.; Antal, A.; Ayache, S.S.; Benninger, D.H.; Brunelin, J.; Cogiamanian, F.; Cotelli, M.; De Ridder, D.; Ferrucci, R.; Langguth, B.; et al. Evidence-Based Guidelines on the Therapeutic Use of Transcranial Direct Current Stimulation (TDCS); Elsevier Ireland Ltd.: Amsterdam, The Netherlands, 2017; Volume 128, pp. 56–92. [Google Scholar]
- Cespón, J.; Rodella, C.; Miniussi, C.; Pellicciari, M.C. Behavioural and Electrophysiological Modulations Induced by Transcranial Direct Current Stimulation in Healthy Elderly and Alzheimer’s Disease Patients: A Pilot Study. Clin. Neurophysiol. 2019, 130, 2038–2052. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.H.S.; Chen, R. Invasive and Noninvasive Brain Stimulation in Parkinson’s Disease: Clinical Effects and Future Perspectives. Clin. Pharmacol. Ther. 2019, 106, 763–775. [Google Scholar] [CrossRef] [PubMed]
- Ferrucci, R.; Bocci, T.; Cortese, F.; Ruggiero, F.; Priori, A. Cerebellar Transcranial Direct Current Stimulation in Neurological Disease; BioMed Central Ltd.: London, UK, 2016; Volume 3. [Google Scholar]
- Guidetti, M.; Arlotti, M.; Bocci, T.; Bianchi, A.M.; Parazzini, M.; Ferrucci, R.; Priori, A. Electric Fields Induced in the Brain by Transcranial Electric Stimulation: A Review of In Vivo Recordings. Biomedicines 2022, 10, 2333. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Liu, H.; Liu, Z.; Rao, J.; Wang, J.; Wang, P.; Gong, X.; Wen, Y. Transcranial Direct Current Stimulation for Parkinson’s Disease: A Systematic Review and Meta-Analysis. Front. Aging Neurosci. 2021, 13, 691. [Google Scholar] [CrossRef]
- Dagan, M.; Herman, T.; Harrison, R.; Zhou, J.; Giladi, N.; Ruffini, G.; Manor, B.; Hausdorff, J.M. Multitarget Transcranial Direct Current Stimulation for Freezing of Gait in Parkinson’s Disease. Mov. Disord. 2018, 33, 642–646. [Google Scholar] [CrossRef] [PubMed]
- Manor, B.; Dagan, M.; Herman, T.; Gouskova, N.A.; Vanderhorst, V.G.; Giladi, N.; Travison, T.G.; Pascual-Leone, A.; Lipsitz, L.A.; Hausdorff, J.M. Multitarget Transcranial Electrical Stimulation for Freezing of Gait: A Randomized Controlled Trial. Mov. Disord. 2021, 36, 2693–2698. [Google Scholar] [CrossRef]
- Putzolu, M.; Ogliastro, C.; Lagravinese, G.; Bonassi, G.; Trompetto, C.; Marchese, R.; Avanzino, L.; Pelosin, E. Investigating the Effects of Transcranial Direct Current Stimulation on Obstacle Negotiation Performance in Parkinson Disease with Freezing of Gait: A Pilot Study. Brain Stimul. 2019, 12, 1583–1585. [Google Scholar] [CrossRef]
- Valentino, F.; Cosentino, G.; Brighina, F.; Pozzi, N.G.; Sandrini, G.; Fierro, B.; Savettieri, G.; D’Amelio, M.; Pacchetti, C. Transcranial Direct Current Stimulation for Treatment of Freezing of Gait: A Cross-over Study. Mov. Disord. 2014, 29, 1064–1069. [Google Scholar] [CrossRef]
- Rascol, O.; Payoux, P.; Ory, F.; Ferreira, J.J.; Brefel-Courbon, C.; Montastruc, J.-L. Limitations of Current Parkinson’s Disease Therapy. Ann. Neurol. 2003, 53 (Suppl. S3), S3–S15. [Google Scholar] [CrossRef]
- Hadar, R.; Winter, R.; Edemann-Callesen, H.; Wieske, F.; Habelt, B.; Khadka, N.; Felgel-Farnholz, V.; Barroeta-Hlusicka, E.; Reis, J.; Tatarau, C.A.; et al. Prevention of Schizophrenia Deficits via Non-Invasive Adolescent Frontal Cortex Stimulation in Rats. Mol. Psychiatry 2019, 25, 896–905. [Google Scholar] [CrossRef] [PubMed]
- McKinnon, C.; Gros, P.; Lee, D.J.; Hamani, C.; Lozano, A.M.; Kalia, L.V.; Kalia, S.K. Deep Brain Stimulation: Potential for Neuroprotection. Ann. Clin. Transl. Neurol. 2018, 6, 174–185. [Google Scholar] [CrossRef] [Green Version]
- de Lau, L.M.; Breteler, M.M. Epidemiology of Parkinson’s Disease. Lancet Neurol. 2006, 5, 525–535. [Google Scholar] [CrossRef]
- Balestrino, R.; Schapira, A.H.V. Parkinson Disease. Eur. J. Neurol. 2020, 27, 27–42. [Google Scholar] [CrossRef]
- Lee, S.B.; Youn, J.; Jang, W.; Yang, H.O. Neuroprotective Effect of Anodal Transcranial Direct Current Stimulation on 1-Methyl-4-Phenyl-1,2,3,6-Tetrahydropyridine (MPTP)-Induced Neurotoxicity in Mice through Modulating Mitochondrial Dynamics. Neurochem. Int. 2019, 129, 104491. [Google Scholar] [CrossRef]
- Hattori, N.; Mizuno, Y. Mitochondrial Dysfunction in Parkinson’s Disease. Exp. Neurobiol. 2015, 24, 103. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Lu, C.; Wei, Y.; Hu, R.; Wang, Y.; Li, K. Transcranial Direct Current Stimulation Ameliorates Behavioral Deficits and Reduces Oxidative Stress in 1-Methyl-4-Phenyl-1,2,3,6-Tetrahydropyridine-Induced Mouse Model of Parkinson’s Disease. Neuromodul. Technol. Neural Interface 2015, 18, 442–447. [Google Scholar] [CrossRef]
- Laste, G.; Caumo, W.; Adachi, L.N.S.; Rozisky, J.R.; De MacEdo, I.C.; Filho, P.R.M.; Partata, W.A.; Fregni, F.; Torres, I.L.S. After-Effects of Consecutive Sessions of Transcranial Direct Current Stimulation (TDCS) in a Rat Model of Chronic Inflammation. Exp. Brain Res. 2012, 221, 75–83. [Google Scholar] [CrossRef]
- Lynch-Day, M.A.; Mao, K.; Wang, K.; Zhao, M.; Klionsky, D.J. The Role of Autophagy in Parkinson’s Disease. Cold Spring Harb. Perspect. Med. 2012, 2, a009357. [Google Scholar] [CrossRef] [Green Version]
- Guidetti, M.; Marceglia, S.; Loh, A.; Harmsen, I.E.; Meoni, S.; Foffani, G.; Lozano, A.M.; Moro, E.; Volkmann, J.; Priori, A. Clinical Perspectives of Adaptive Deep Brain Stimulation. Brain Stimul. 2021, 14, 1238–1247. [Google Scholar] [CrossRef]
- Hariz, M.; Blomstedt, P. Deep Brain Stimulation for Parkinson’s Disease. J. Intern. Med. 2022, 292, 764–778. [Google Scholar] [CrossRef] [PubMed]
- Barker, R.A.; Drouin-Ouellet, J.; Parmar, M. Cell-Based Therapies for Parkinson Disease—Past Insights and Future Potential. Nat. Rev. Neurol. 2015, 11, 492–503. [Google Scholar] [CrossRef] [PubMed]
- Madrid, J.; Benninger, D.H. Non-Invasive Brain Stimulation for Parkinson’s Disease: Clinical Evidence, Latest Concepts and Future Goals: A Systematic Review. J. Neurosci. Methods 2021, 347, 108957. [Google Scholar] [CrossRef]
- Ganguly, J.; Murgai, A.; Sharma, S.; Aur, D.; Jog, M. Non-Invasive Transcranial Electrical Stimulation in Movement Disorders. Front. Neurosci. 2020, 14, 522. [Google Scholar] [CrossRef] [PubMed]
- Benninger, D.H.; Lomarev, M.; Lopez, G.; Wassermann, E.M.; Li, X.; Considine, E.; Hallett, M. Transcranial Direct Current Stimulation for the Treatment of Parkinson’s Disease. J. Neurol. Neurosurg. Psychiatry 2010, 81, 1105–1111. [Google Scholar] [CrossRef] [Green Version]
- Simpson, M.W.; Mak, M. The Effect of Transcranial Direct Current Stimulation on Upper Limb Motor Performance in Parkinson’s Disease: A Systematic Review. J. Neurol. 2020, 267, 3479–3488. [Google Scholar] [CrossRef]
- Broeder, S.; Nackaerts, E.; Heremans, E.; Vervoort, G.; Meesen, R.; Verheyden, G.; Nieuwboer, A. Transcranial Direct Current Stimulation in Parkinson’s Disease: Neurophysiological Mechanisms and Behavioral Effects. Neurosci. Biobehav. Rev. 2015, 57, 105–117. [Google Scholar] [CrossRef]
- Fregni, F.; Boggio, P.S.; Santos, M.C.; Lima, M.; Vieira, A.L.; Rigonatti, S.P.; Silva, M.T.A.; Barbosa, E.R.; Nitsche, M.A.; Pascual-Leone, A. Noninvasive Cortical Stimulation with Transcranial Direct Current Stimulation in Parkinson’s Disease. Mov. Disord. 2006, 21, 1693–1702. [Google Scholar] [CrossRef]
- Elsner, B.; Kugler, J.; Pohl, M.; Mehrholz, J. Transcranial Direct Current Stimulation (TDCS) for Idiopathic Parkinson’s Disease. Cochrane Database Syst. Rev. 2016, 2016, CD010916. [Google Scholar] [CrossRef]
- Doruk, D.; Gray, Z.; Bravo, G.L.; Pascual-Leone, A.; Fregni, F. Effects of TDCS on Executive Function in Parkinson’s Disease. Neurosci. Lett. 2014, 582, 27–31. [Google Scholar] [CrossRef]
- Manto, M.; Argyropoulos, G.P.D.; Bocci, T.; Celnik, P.A.; Corben, L.A.; Guidetti, M.; Koch, G.; Priori, A.; Rothwell, J.C.; Sadnicka, A.; et al. Consensus Paper: Novel Directions and Next Steps of Non-Invasive Brain Stimulation of the Cerebellum in Health and Disease. Cerebellum 2021, 21, 1092–1122. [Google Scholar] [CrossRef] [PubMed]
- Sala, G.; Bocci, T.; Borzì, V.; Parazzini, M.; Priori, A.; Ferrarese, C. Direct Current Stimulation Enhances Neuronal Alpha-Synuclein Degradation in Vitro. Sci. Rep. 2021, 11, 2197. [Google Scholar] [CrossRef]
- Fukai, M.; Bunai, T.; Hirosawa, T.; Kikuchi, M.; Ito, S.; Minabe, Y.; Ouchi, Y. Endogenous Dopamine Release under Transcranial Direct-Current Stimulation Governs Enhanced Attention: A Study with Positron Emission Tomography. Transl. Psychiatry 2019, 9, 115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonteneau, C.; Redoute, J.; Haesebaert, F.; Le Bars, D.; Costes, N.; Suaud-Chagny, M.F.; Brunelin, J. Frontal Transcranial Direct Current Stimulation Induces Dopamine Release in the Ventral Striatum in Human. Cereb. Cortex 2018, 28, 2636–2646. [Google Scholar] [CrossRef]
- Tanaka, T.; Takano, Y.; Tanaka, S.; Hironaka, N.; Kobayashi, K.; Hanakawa, T.; Watanabe, K.; Honda, M. Transcranial Direct-Current Stimulation Increases Extracellular Dopamine Levels in the Rat Striatum. Front. Syst. Neurosci. 2013, 7, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fritsch, B.; Reis, J.; Martinowich, K.; Schambra, H.M.; Ji, Y.; Cohen, L.G.; Lu, B. Direct Current Stimulation Promotes BDNF-Dependent Synaptic Plasticity: Potential Implications for Motor Learning. Neuron 2010, 66, 198–204. [Google Scholar] [CrossRef] [Green Version]
- Spezia Adachi, L.N.; Caumo, W.; Laste, G.; Fernandes Medeiros, L.; Ripoll Rozisky, J.; De Souza, A.; Fregni, F.; Torres, I.L.S. Reversal of Chronic Stress-Induced Pain by Transcranial Direct Current Stimulation (TDCS) in an Animal Model. Brain Res. 2012, 1489, 17–26. [Google Scholar] [CrossRef] [Green Version]
- Ranieri, F.; Podda, M.V.; Riccardi, E.; Frisullo, G.; Dileone, M.; Profice, P.; Pilato, F.; di Lazzaro, V.; Grassi, C. Modulation of LTP at Rat Hippocampal CA3-CA1 Synapses by Direct Current Stimulation. J. Neurophysiol. 2012, 107, 1868–1880. [Google Scholar] [CrossRef] [Green Version]
- Zigova, T.; Pencea, V.; Wiegand, S.J.; Luskin, M.B. Intraventricular Administration of BDNF Increases the Number of Newly Generated Neurons in the Adult Olfactory Bulb. Mol. Cell. Neurosci. 1998, 11, 234–245. [Google Scholar] [CrossRef]
- Benraiss, A.; Chmielnicki, E.; Lerner, K.; Roh, D.; Goldman, S.A. Adenoviral Brain-Derived Neurotrophic Factor Induces Both Neostriatal and Olfactory Neuronal Recruitment from Endogenous Progenitor Cells in the Adult Forebrain. J. Neurosci. 2001, 21, 6718–6731. [Google Scholar] [CrossRef]
- Huang, E.J.; Reichardt, L.F. Neurotrophins: Roles in Neuronal Development and Function. Annu. Rev. Neurosci. 2001, 24, 677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, X.J.; Huang, Y.T.; Huang, Y.Z.; Kuo, C.W.; Peng, C.W.; Rotenberg, A.; Juan, C.H.; Pei, Y.C.; Chen, Y.H.; Chen, K.Y.; et al. Early Transcranial Direct Current Stimulation Treatment Exerts Neuroprotective Effects on 6-OHDA-Induced Parkinsonism in Rats. Brain Stimul. 2020, 13, 655–663. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.B.; Kim, H.T.; Yang, H.O.; Jang, W. Anodal Transcranial Direct Current Stimulation Prevents Methyl-4-Phenyl-1,2,3,6-Tetrahydropyridine (MPTP)-Induced Neurotoxicity by Modulating Autophagy in an in Vivo Mouse Model of Parkinson’s Disease. Sci. Rep. 2018, 8, 15165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alam, G.; Richardson, J.R. Regulation of Tyrosine Hydroxylase: Relevance to Parkinson’s Disease. Genet. Neurol. Behav. Diet Park. Dis. 2020, 2, 51–66. [Google Scholar] [CrossRef]
- Ischiropoulos, H.; Beckman, J.S. Oxidative Stress and Nitration in Neurodegeneration: Cause, Effect, or Association? J. Clin. Investig. 2003, 111, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deas, E.; Wood, N.W.; Plun-Favreau, H. Mitophagy and Parkinson’s Disease: The PINK1–Parkin Link. Biochim. Biophys. Acta-Mol. Cell Res. 2011, 1813, 623–633. [Google Scholar] [CrossRef] [Green Version]
- Rae, C.D.; Lee, V.H.C.; Ordidge, R.J.; Alonzo, A.; Loo, C. Anodal Transcranial Direct Current Stimulation Increases Brain Intracellular PH and Modulates Bioenergetics. Int. J. Neuropsychopharmacol. 2013, 16, 1695–1706. [Google Scholar] [CrossRef] [Green Version]
- Filichia, E.; Hoffer, B.; Qi, X.; Luo, Y. Inhibition of Drp1 Mitochondrial Translocation Provides Neural Protection in Dopaminergic System in a Parkinson’s Disease Model Induced by MPTP. Sci. Rep. 2016, 6, 32656. [Google Scholar] [CrossRef] [Green Version]
- Majeski, A.E.; Fred Dice, J. Mechanisms of Chaperone-Mediated Autophagy. Int. J. Biochem. Cell Biol. 2004, 36, 2435–2444. [Google Scholar] [CrossRef]
- Cuervo, A.M.; Stafanis, L.; Fredenburg, R.; Lansbury, P.T.; Sulzer, D. Impaired Degradation of Mutant Alpha-Synuclein by Chaperone-Mediated Autophagy. Science 2004, 305, 1292–1295. [Google Scholar] [CrossRef]
- Lu, J.; Wu, M.; Yue, Z. Autophagy and Parkinson’s Disease. Adv. Exp. Med. Biol. 2020, 1207, 21–51. [Google Scholar] [CrossRef] [PubMed]
- Vekrellis, K.; Stefanis, L. Targeting Intracellular and Extracellular Alpha-Synuclein as a Therapeutic Strategy in Parkinson’s Disease and Other Synucleinopathies. Expert Opin. Ther. Targets 2012, 16, 421–432. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Xiong, L.J.; Tong, Y.; Mao, M. Neuroprotective Effect of Brain-Derived Neurotrophic Factor Mediated by Autophagy through the PI3K/Akt/MTOR Pathway. Mol. Med. Rep. 2013, 8, 1011–1016. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, T.H.; He, X.K.; Liu, H.H.; Chen, J.J.J.; Peng, C.W.; Liu, H.L.; Rotenberg, A.; Chen, K.T.; Chang, M.Y.; Chiang, Y.H.; et al. Early Repetitive Transcranial Magnetic Stimulation Exerts Neuroprotective Effects and Improves Motor Functions in Hemiparkinsonian Rats. Neural Plast. 2021, 2021, 1763533. [Google Scholar] [CrossRef] [PubMed]
- Ba, M.; Ma, G.; Ren, C.; Sun, X.; Kong, M. Repetitive Transcranial Magnetic Stimulation for Treatment of Lactacystin-Induced Parkinsonian Rat Model. Oncotarget 2017, 8, 50921–50929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knopman, D.S.; Amieva, H.; Petersen, R.C.; Chételat, G.; Holtzman, D.M.; Hyman, B.T.; Nixon, R.A.; Jones, D.T. Alzheimer Disease. Nat. Rev. Dis. Prim. 2021, 7, 33. [Google Scholar] [CrossRef]
- Chou, Y.H.; Ton That, V.; Sundman, M. A Systematic Review and Meta-Analysis of RTMS Effects on Cognitive Enhancement in Mild Cognitive Impairment and Alzheimer’s Disease. Neurobiol. Aging 2020, 86, 1–10. [Google Scholar] [CrossRef]
- Cammisuli, D.M.; Cignoni, F.; Ceravolo, R.; Bonuccelli, U.; Castelnuovo, G. Transcranial Direct Current Stimulation (TDCS) as a Useful Rehabilitation Strategy to Improve Cognition in Patients With Alzheimer’s Disease and Parkinson’s Disease: An Updated Systematic Review of Randomized Controlled Trials. Front. Neurol. 2022, 12, 2648. [Google Scholar] [CrossRef]
- Rajji, T.K. Transcranial Magnetic and Electrical Stimulation in Alzheimer’s Disease and Mild Cognitive Impairment: A Review of Randomized Controlled Trials. Clin. Pharmacol. Ther. 2019, 106, 776–780. [Google Scholar] [CrossRef]
- Teselink, J.; Bawa, K.K.; Koo, G.K.; Sankhe, K.; Liu, C.S.; Rapoport, M.; Oh, P.; Marzolini, S.; Gallagher, D.; Swardfager, W.; et al. Efficacy of Non-Invasive Brain Stimulation on Global Cognition and Neuropsychiatric Symptoms in Alzheimer’s Disease and Mild Cognitive Impairment: A Meta-Analysis and Systematic Review. Ageing Res. Rev. 2021, 72, 101499. [Google Scholar] [CrossRef]
- Lefaucheur, J.P. A Comprehensive Database of Published TDCS Clinical Trials (2005–2016). Neurophysiol. Clin. 2016, 46, 319–398. [Google Scholar] [CrossRef] [PubMed]
- Xiao, N.; Le, Q.T. Neurotrophic Factors and Their Potential Applications in Tissue Regeneration. Arch. Immunol. Ther. Exp. (Warsz) 2016, 64, 89–99. [Google Scholar] [CrossRef]
- Mitra, S.; Behbahani, H.; Eriksdotter, M. Innovative Therapy for Alzheimer’s Disease-With Focus on Biodelivery of NGF. Front. Neurosci. 2019, 13, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, T.K.S.; Ho, C.S.H.; Tam, W.W.S.; Kua, E.H.; Ho, R.C.M. Decreased Serum Brain-Derived Neurotrophic Factor (BDNF) Levels in Patients with Alzheimer’s Disease (AD): A Systematic Review and Meta-Analysis. Int. J. Mol. Sci. 2019, 20, 257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Straten, G.; Eschweiler, G.W.; Maetzler, W.; Laske, C.; Leyhe, T. Glial Cell-Line Derived Neurotrophic Factor (GDNF) Concentrations in Cerebrospinal Fluid and Serum of Patients with Early Alzheimer’s Disease and Normal Controls. J. Alzheimer’s Dis. 2009, 18, 331–337. [Google Scholar] [CrossRef]
- Rockenstein, E.; Ubhi, K.; Doppler, E.; Novak, P.; Moessler, H.; Li, B.; Blanchard, J.; Grundke-Iqbal, I.; Iqbal, K.; Mante, M.; et al. Regional Comparison of the Neurogenic Effects of CNTF-Derived Peptides and Cerebrolysin in AβPP Transgenic Mice. J. Alzheimer’s Dis. 2011, 27, 743–752. [Google Scholar] [CrossRef] [Green Version]
- Knipper, M.; da Penha Berzaghi, M.; Blöchl, A.; Breer, H.; Thoenen, H.; Lindholm, D. Positive Feedback between Acetylcholine and the Neurotrophins Nerve Growth Factor and Brain-Derived Neurotrophic Factor in the Rat Hippocampus. Eur. J. Neurosci. 1994, 6, 668–671. [Google Scholar] [CrossRef]
- Choung, J.S.; Kim, J.M.; Ko, M.H.; Cho, D.S.; Kim, M.Y. Therapeutic Efficacy of Repetitive Transcranial Magnetic Stimulation in an Animal Model of Alzheimer’s Disease. Sci. Rep. 2021, 11, 437. [Google Scholar] [CrossRef]
- Velioglu, H.A.; Hanoglu, L.; Bayraktaroglu, Z.; Toprak, G.; Guler, E.M.; Bektay, M.Y.; Mutlu-Burnaz, O.; Yulug, B. Left Lateral Parietal RTMS Improves Cognition and Modulates Resting Brain Connectivity in Patients with Alzheimer’s Disease: Possible Role of BDNF and Oxidative Stress. Neurobiol. Learn. Mem. 2021, 180, 107410. [Google Scholar] [CrossRef]
- Chen, X.; Chen, S.; Liang, W.; Ba, F. Administration of Repetitive Transcranial Magnetic Stimulation Attenuates A β 1-42-Induced Alzheimer’s Disease in Mice by Activating β-Catenin Signaling. Biomed Res. Int. 2019, 2019, 1431760. [Google Scholar] [CrossRef]
- Tan, T.; Xie, J.; Liu, T.; Chen, X.; Zheng, X.; Tong, Z.; Tian, X. Low-Frequency (1Hz) Repetitive Transcranial Magnetic Stimulation (RTMS) Reverses Aβ1–42-Mediated Memory Deficits in Rats. Exp. Gerontol. 2013, 48, 786–794. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Dong, G.Y.; Wang, L.X. High-Frequency Transcranial Magnetic Stimulation Protects APP/PS1 Mice against Alzheimer’s Disease Progress by Reducing APOE and Enhancing Autophagy. Brain Behav. 2020, 10, e01740. [Google Scholar] [CrossRef] [PubMed]
- Ohira, K.; Hayashi, M. A New Aspect of the TrkB Signaling Pathway in Neural Plasticity. Curr. Neuropharmacol. 2009, 7, 276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obulesu, M.; Lakshmi, M.J. Apoptosis in Alzheimer’s Disease: An Understanding of the Physiology, Pathology and Therapeutic Avenues. Neurochem. Res. 2014, 39, 2301–2312. [Google Scholar] [CrossRef]
- Paradis, E.; Douillard, H.; Koutroumanis, M.; Goodyer, C.; LeBlanc, A. Amyloid Beta Peptide of Alzheimer’s Disease Downregulates Bcl-2 and Upregulates Bax Expression in Human Neurons. J. Neurosci. 1996, 16, 7533–7539. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Wang, X. Antioxidant Therapies for Alzheimer’s Disease. Oxid. Med. Cell. Longev. 2012, 2012, 472932. [Google Scholar] [CrossRef] [Green Version]
- Selley, M.L. Increased Concentrations of Homocysteine and Asymmetric Dimethylarginine and Decreased Concentrations of Nitric Oxide in the Plasma of Patients with Alzheimer’s Disease. Neurobiol. Aging 2003, 24, 903–907. [Google Scholar] [CrossRef]
- Guix, F.X.; Uribesalgo, I.; Coma, M.; Muñoz, F.J. The Physiology and Pathophysiology of Nitric Oxide in the Brain. Prog. Neurobiol. 2005, 76, 126–152. [Google Scholar] [CrossRef]
- Katusic, Z.S.; Austin, S.A. Endothelial Nitric Oxide: Protector of a Healthy Mind. Eur. Heart J. 2014, 35, 888–894. [Google Scholar] [CrossRef] [Green Version]
- Trivedi, D.P.; Hallock, K.J.; Bergethon, P.R. Electric Fields Caused by Blood Flow Modulate Vascular Endothelial Electrophysiology and Nitric Oxide Production. Bioelectromagnetics 2013, 34, 22–30. [Google Scholar] [CrossRef]
- Marceglia, S.; Mrakic-Sposta, S.; Rosa, M.; Ferrucci, R.; Mameli, F.; Vergari, M.; Arlotti, M.; Ruggiero, F.; Scarpini, E.; Galimberti, D.; et al. Transcranial Direct Current Stimulation Modulates Cortical Neuronal Activity in Alzheimer’s Disease. Front. Neurosci. 2016, 10, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirabali, T.; Rust, R.; Rigotti, S.; Siccoli, A.; Nitsch, R.M.; Kulic, L. Distinct Changes in All Major Components of the Neurovascular Unit across Different Neuropathological Stages of Alzheimer’s Disease. Brain Pathol. 2020, 30, 1056–1070. [Google Scholar] [CrossRef] [PubMed]
- Bai, H.; Forrester, J.V.; Zhao, M. DC Electric Stimulation Upregulates Angiogenic Factors in Endothelial Cells through Activation of VEGF Receptors. Cytokine 2011, 55, 110–115. [Google Scholar] [CrossRef] [Green Version]
- Rueger, M.A.; Keuters, M.H.; Walberer, M.; Braun, R.; Klein, R.; Sparing, R.; Fink, G.R.; Graf, R.; Schroeter, M. Multi-Session Transcranial Direct Current Stimulation (TDCS) Elicits Inflammatory and Regenerative Processes in the Rat Brain. PLoS ONE 2012, 7, e43776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cancel, L.M.; Arias, K.; Bikson, M.; Tarbell, J.M. Direct Current Stimulation of Endothelial Monolayers Induces a Transient and Reversible Increase in Transport Due to the Electroosmotic Effect. Sci. Rep. 2018, 8, 9265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Y.; Yang, H.; Yan, X.; Wu, Y.; Wei, G.; Wu, X.; Tian, X.; Xiong, Y.; Wu, G.; Wen, H. Transcranial Direct Current Stimulation Alleviates Neurovascular Unit Dysfunction in Mice With Preclinical Alzheimer’s Disease. Front. Aging Neurosci. 2022, 14, 857415. [Google Scholar] [CrossRef]
- Teter, B. Rodent Aging. In Encyclopedia of Neuroscience; Academic Press: Oxford, UK, 2009; pp. 397–406. [Google Scholar] [CrossRef]
- Khedr, E.M.; Salama, R.H.; Abdel Hameed, M.; Abo Elfetoh, N.; Seif, P. Therapeutic Role of Transcranial Direct Current Stimulation in Alzheimer Disease Patients: Double-Blind, Placebo-Controlled Clinical Trial. Neurorehabil. Neural Repair 2019, 33, 384–394. [Google Scholar] [CrossRef]
- Dhaynaut, M.; Sprugnoli, G.; Cappon, D.; Macone, J.; Sanchez, J.S.; Normandin, M.D.; Guehl, N.J.; Koch, G.; Paciorek, R.; Connor, A.; et al. Impact of 40 Hz Transcranial Alternating Current Stimulation on Cerebral Tau Burden in Patients with Alzheimer’s Disease: A Case Series. J. Alzheimer’s Dis. 2022, 85, 1667–1676. [Google Scholar] [CrossRef]
- Luo, Y.; Yang, W.; Li, N.; Yang, X.; Zhu, B.; Wang, C.; Hou, W.; Wang, X.; Wen, H.; Tian, X. Anodal Transcranial Direct Current Stimulation Can Improve Spatial Learning and Memory and Attenuate Aβ 42 Burden at the Early Stage of Alzheimer’s Disease in APP/PS1 Transgenic Mice. Front. Aging Neurosci. 2020, 12, 134. [Google Scholar] [CrossRef]
- Storch, S.; Samantzis, M.; Balbi, M. Driving Oscillatory Dynamics: Neuromodulation for Recovery After Stroke. Front. Syst. Neurosci. 2021, 15, 65. [Google Scholar] [CrossRef]
- Yang, Y.; Yang, L.Y.; Orban, L.; Cuylear, D.; Thompson, J.; Simon, B.; Yang, Y. Non-Invasive Vagus Nerve Stimulation Reduces Blood-Brain Barrier Disruption in a Rat Model of Ischemic Stroke. Brain Stimul. 2018, 11, 689–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.P.; Zhao, Y.; Qin, X.Y.; Wan, L.Y.; Fan, X.X. Non-Invasive Vagus Nerve Stimulation Protects Against Cerebral Ischemia/Reperfusion Injury and Promotes Microglial M2 Polarization Via Interleukin-17A Inhibition. J. Mol. Neurosci. 2019, 67, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Notturno, F.; Pace, M.; Zappasodi, F.; Cam, E.; Bassetti, C.L.; Uncini, A. Neuroprotective Effect of Cathodal Transcranial Direct Current Stimulation in a Rat Stroke Model. J. Neurol. Sci. 2014, 342, 146–151. [Google Scholar] [CrossRef]
- Priori, A.; Maiorana, N.; Dini, M.; Guidetti, M.; Marceglia, S.; Ferrucci, R. Adaptive Deep Brain Stimulation (ADBS). Int. Rev. Neurobiol. 2021, 159, 111–127. [Google Scholar] [CrossRef] [PubMed]
- Lozano, A.M.; Lipsman, N.; Bergman, H.; Brown, P.; Chabardes, S.; Chang, J.W.; Matthews, K.; McIntyre, C.C.; Schlaepfer, T.E.; Schulder, M.; et al. Deep Brain Stimulation: Current Challenges and Future Directions. Nat. Rev. Neurol. 2019, 15, 148–160. [Google Scholar] [CrossRef] [PubMed]
- Herrington, T.M.; Cheng, J.J.; Eskandar, E.N. Mechanisms of Deep Brain Stimulation. J. Neurophysiol. 2016, 115, 19–38. [Google Scholar] [CrossRef] [Green Version]
- Wallace, B.A.; Ashkan, K.; Heise, C.E.; Foote, K.D.; Torres, N.; Mitrofanis, J.; Benabid, A.L. Survival of Midbrain Dopaminergic Cells after Lesion or Deep Brain Stimulation of the Subthalamic Nucleus in MPTP-Treated Monkeys. Brain 2007, 130, 2129–2145. [Google Scholar] [CrossRef] [Green Version]
- Musacchio, T.; Rebenstorff, M.; Fluri, F.; Brotchie, J.M.; Volkmann, J.; Koprich, J.B.; Ip, C.W. Subthalamic Nucleus Deep Brain Stimulation Is Neuroprotective in the A53T α-Synuclein Parkinson’s Disease Rat Model. Ann. Neurol. 2017, 81, 825–836. [Google Scholar] [CrossRef] [Green Version]
- Spieles-Engemann, A.L.; Steece-Collier, K.; Behbehani, M.M.; Collier, T.J.; Wohlgenant, S.L.; Kemp, C.J.; Cole-Strauss, A.; Levine, N.D.; Gombash, S.E.; Thompson, V.B.; et al. Subthalamic Nucleus Stimulation Increases Brain Derived Neurotrophic Factor in the Nigrostriatal System and Primary Motor Cortex. J. Park. Dis. 2011, 1, 123. [Google Scholar] [CrossRef]
- Fischer, D.L.; Kemp, C.J.; Cole-Strauss, A.; Polinski, N.K.; Paumier, K.L.; Lipton, J.W.; Steece-Collier, K.; Collier, T.J.; Buhlinger, D.J.; Sortwell, C.E. Subthalamic Nucleus Deep Brain Stimulation Employs TrkB Signaling for Neuroprotection and Functional Restoration. J. Neurosci. 2017, 37, 6786–6796. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, M.C.; Obeso, J.A.; Olanow, C.W. Subthalamic Nucleus-Mediated Excitotoxicity in Parkinson’s Disease: A Target for Neuroprotection. Ann. Neurol. 1998, 44, S175–S188. [Google Scholar] [CrossRef] [PubMed]
- Baldermann, J.C.; Hardenacke, K.; Hu, X.; Köster, P.; Horn, A.; Freund, H.J.; Zilles, K.; Sturm, V.; Visser-Vandewalle, V.; Jessen, F.; et al. Neuroanatomical Characteristics Associated with Response to Deep Brain Stimulation of the Nucleus Basalis of Meynert for Alzheimer’s Disease. Neuromodulation 2018, 21, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Chu, H.; Ma, Y.; Zhou, Z.; Dai, C.; Huang, X.; Fang, L.; Ao, Q.; Huang, D. The Neuroprotective Effect of Deep Brain Stimulation at Nucleus Basalis of Meynert in Transgenic Mice with Alzheimer’s Disease. Brain Stimul. 2019, 12, 161–174. [Google Scholar] [CrossRef] [PubMed]
- Xia, F.; Yiu, A.; Stone, S.S.D.; Oh, S.; Lozano, A.M.; Josselyn, S.A.; Frankland, P.W. Entorhinal Cortical Deep Brain Stimulation Rescues Memory Deficits in Both Young and Old Mice Genetically Engineered to Model Alzheimer’s Disease. Neuropsychopharmacology 2017, 42, 2493–2503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akwa, Y.; Gondard, E.; Mann, A.; Capetillo-Zarate, E.; Alberdi, E.; Matute, C.; Marty, S.; Vaccari, T.; Lozano, A.M.; Baulieu, E.E.; et al. Synaptic Activity Protects against AD and FTD-like Pathology via Autophagic-Lysosomal Degradation. Mol. Psychiatry 2017, 23, 1530–1540. [Google Scholar] [CrossRef] [Green Version]
- Shaw, V.E.; Keay, K.A.; Ashkan, K.; Benabid, A.L.; Mitrofanis, J. Dopaminergic Cells in the Periaqueductal Grey Matter of MPTP-Treated Monkeys and Mice; Patterns of Survival and Effect of Deep Brain Stimulation and Lesion of the Subthalamic Nucleus. Park. Relat. Disord. 2010, 16, 338–344. [Google Scholar] [CrossRef]
- Harnack, D.; Meissner, W.; Jira, J.A.; Winter, C.; Morgenstern, R.; Kupsch, A. Placebo-Controlled Chronic High-Frequency Stimulation of the Subthalamic Nucleus Preserves Dopaminergic Nigral Neurons in a Rat Model of Progressive Parkinsonism. Exp. Neurol. 2008, 210, 257–260. [Google Scholar] [CrossRef]
- Spieles-Engemann, A.L.; Behbehani, M.M.; Collier, T.J.; Wohlgenant, S.L.; Steece-Collier, K.; Paumier, K.; Daley, B.F.; Gombash, S.; Madhavan, L.; Mandybur, G.T.; et al. Stimulation of the Rat Subthalamic Nucleus Is Neuroprotective Following Significant Nigral Dopamine Neuron Loss. Neurobiol. Dis. 2010, 39, 105–115. [Google Scholar] [CrossRef] [Green Version]
- Hilker, R.; Portman, A.T.; Voges, J.; Staal, M.J.; Burghaus, L.; Van Laar, T.; Koulousakis, A.; Maguire, R.P.; Pruim, J.; De Jong, B.M.; et al. Disease Progression Continues in Patients with Advanced Parkinson’s Disease and Effective Subthalamic Nucleus Stimulation. J. Neurol. Neurosurg. Psychiatry 2005, 76, 1217–1221. [Google Scholar] [CrossRef]
- Pal, G.D.; Ouyang, B.; Serrano, G.; Shill, H.A.; Goetz, C.; Stebbins, G.; Metman, L.V.; Driver-Dunckley, E.; Mehta, S.H.; Caviness, J.N.; et al. Comparison of Neuropathology in Parkinson’s Disease Subjects with and without Deep Brain Stimulation. Mov. Disord. 2017, 32, 274–277. [Google Scholar] [CrossRef]
- Fischer, D.L.; Manfredsson, F.P.; Kemp, C.J.; Cole-Strauss, A.; Lipton, J.W.; Duffy, M.F.; Polinski, N.K.; Steece-Collier, K.; Collier, T.J.; Gombash, S.E.; et al. Subthalamic Nucleus Deep Brain Stimulation Does Not Modify the Functional Deficits or Axonopathy Induced by Nigrostriatal α-Synuclein Overexpression. Sci. Rep. 2017, 7, 16356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obeso, J.A.; Rodriguez-Oroz, M.C.; Rodriguez, M.; Macias, R.; Alvarez, L.; Guridi, J.; Vitek, J.; DeLong, M.R. Pathophysiologic Basis of Surgery for Parkinson’s Disease. Neurology 2000, 55, S7–S12. [Google Scholar] [PubMed]
- Maesawa, S.; Kaneoke, Y.; Kajita, Y.; Usui, N.; Misawa, N.; Nakayama, A.; Yoshida, J. Long-Term Stimulation of the Subthalamic Nucleus in Hemiparkinsonian Rats: Neuroprotection of Dopaminergic Neurons. J. Neurosurg. 2004, 100, 679–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gratwicke, J.; Kahan, J.; Zrinzo, L.; Hariz, M.; Limousin, P.; Foltynie, T.; Jahanshahi, M. The Nucleus Basalis of Meynert: A New Target for Deep Brain Stimulation in Dementia? Neurosci. Biobehav. Rev. 2013, 37, 2676–2688. [Google Scholar] [CrossRef] [PubMed]
- Hotta, H.; Kagitani, F.; Kondo, M.; Uchida, S. Basal Forebrain Stimulation Induces NGF Secretion in Ipsilateral Parietal Cortex via Nicotinic Receptor Activation in Adult, but Not Aged Rats. Neurosci. Res. 2009, 63, 122–128. [Google Scholar] [CrossRef]
- Kilgard, M.P.; Merzenich, M.M. Cortical Map Reorganization Enabled by Nucleus Basalis Activity. Science 1998, 279, 1714–1718. [Google Scholar] [CrossRef]
- Leplus, A.; Lauritzen, I.; Melon, C.; Kerkerian-Le Goff, L.; Fontaine, D.; Checler, F. Chronic Fornix Deep Brain Stimulation in a Transgenic Alzheimer’s Rat Model Reduces Amyloid Burden, Inflammation, and Neuronal Loss. Brain Struct. Funct. 2019, 224, 363–372. [Google Scholar] [CrossRef]
- Li, Y.; Liu, L.; Barger, S.W.; Griffin, W.S.T. Interleukin-1 Mediates Pathological Effects of Microglia on Tau Phosphorylation and on Synaptophysin Synthesis in Cortical Neurons through a P38-MAPK Pathway. J. Neurosci. 2003, 23, 1605–1611. [Google Scholar] [CrossRef] [Green Version]
- Laxton, A.W.; Tang-Wai, D.F.; McAndrews, M.P.; Zumsteg, D.; Wennberg, R.; Keren, R.; Wherrett, J.; Naglie, G.; Hamani, C.; Smith, G.S.; et al. A phase I trial of deep brain stimulation of memory circuits in Alzheimer's disease. Ann. Neurol. 2010, 68, 521–534. [Google Scholar] [CrossRef]
- Lozano, A.M.; Fosdick, L.; Chakravarty, M.M.; Leoutsakos, J.M.; Munro, C.; Oh, E.; Drake, K.E.; Lyman, C.H.; Rosenberg, P.B.; Anderson, W.S.; et al. A Phase II Study of Fornix Deep Brain Stimulation in Mild Alzheimer’s Disease. J. Alzheimer’s Dis. 2016, 54, 777–787. [Google Scholar] [CrossRef] [Green Version]
- Hadar, R.; Bikovski, L.; Soto-Montenegro, M.L.; Schimke, J.; Maier, P.; Ewing, S.; Voget, M.; Wieske, F.; Götz, T.; Desco, M.; et al. Early Neuromodulation Prevents the Development of Brain and Behavioral Abnormalities in a Rodent Model of Schizophrenia. Mol. Psychiatry 2017, 23, 943–951. [Google Scholar] [CrossRef] [PubMed]
- Zeiss, C.J.; Shin, D.; Wyk, B.V.; Beck, A.P.; Zatz, N.; Sneiderman, C.A.; Kilicoglu, H. Menagerie: A Text-Mining Tool to Support Animal-Human Translation in Neurodegeneration Research. PLoS ONE 2019, 14, e0226176. [Google Scholar] [CrossRef] [PubMed]
- Konnova, E.A.; Swanberg, M. Animal Models of Parkinson’s Disease. In Parkinson’s Disease: Pathogenesis and Clinical Aspects; Codon Publications: Brisbane, Australia, 2018; pp. 83–106. [Google Scholar] [CrossRef]
- Anselmi, C.; Kowarsky, M.; Gasparini, F.; Caicci, F.; Ishizuka, K.J.; Palmeri, K.J.; Raveh, T.; Sinha, R.; Neff, N.; Quake, S.R.; et al. Two Distinct Evolutionary Conserved Neural Degeneration Pathways Characterized in a Colonial Chordate. Proc. Natl. Acad. Sci. USA 2022, 119, e2203032119. [Google Scholar] [CrossRef] [PubMed]
- Sekiya, M.; Wang, M.; Fujisaki, N.; Sakakibara, Y.; Quan, X.; Ehrlich, M.E.; De Jager, P.L.; Bennett, D.A.; Schadt, E.E.; Gandy, S.; et al. Integrated Biology Approach Reveals Molecular and Pathological Interactions among Alzheimer’s Aβ42, Tau, TREM2, and TYROBP in Drosophila Models. Genome Med. 2018, 10, 26. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Tames, J.; Asai, A.; Hirata, A. Significant Group-Level Hotspots Found in Deep Brain Regions during Transcranial Direct Current Stimulation (TDCS): A Computational Analysis of Electric Fields. Clin. Neurophysiol. 2020, 131, 755–765. [Google Scholar] [CrossRef] [PubMed]
- Karimi, F.; Attarpour, A.; Amirfattahi, R.; Nezhad, A.Z. Computational Analysis of Non-Invasive Deep Brain Stimulation Based on Interfering Electric Fields. Phys. Med. Biol. 2019, 64, 235010. [Google Scholar] [CrossRef]
- Missey, F.; Rusina, E.; Acerbo, E.; Botzanowski, B.; Trébuchon, A.; Bartolomei, F.; Jirsa, V.; Carron, R.; Williamson, A. Orientation of Temporal Interference for Non-Invasive Deep Brain Stimulation in Epilepsy. Front. Neurosci. 2021, 15, 656. [Google Scholar] [CrossRef]
- Bocci, T.; Caleo, M.; Tognazzi, S.; Francini, N.; Briscese, L.; Maffei, L.; Rossi, S.; Priori, A.; Sartucci, F. Evidence for Metaplasticity in the Human Visual Cortex. J. Neural Transm. 2013, 121, 221–231. [Google Scholar] [CrossRef]
- Stagg, C.J.; Best, J.G.; Stephenson, M.C.; O’Shea, J.; Wylezinska, M.; Kineses, Z.T.; Morris, P.G.; Matthews, P.M.; Johansen-Berg, H. Polarity-Sensitive Modulation of Cortical Neurotransmitters by Transcranial Stimulation. J. Neurosci. 2009, 29, 5202–5206. [Google Scholar] [CrossRef] [Green Version]
- Bikson, M.; Dmochowski, J. What It Means to Go Deep with Non-Invasive Brain Stimulation. Clin. Neurophysiol. 2020, 131, 752–754. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Study | Sample/Animals | Polarity | Configuration | Parameters | Biological Outcomes | Biological Results |
|---|---|---|---|---|---|---|
| Li et al., 2015 [19] | 36 C57Bl mice (n = 9 in control group; n = 9 in sham tDCS group; n = 9 in tDCS groups; n = 9 in drug group) | Anodal/Sham | AE: left frontal cortex; R: between the shoulders | 0.2 mA, 10 min/day, 21 consecutive days AEA: 3.5 mm2 CD: 5.7 mA/cm2 | DA, TH, SOD and GSH-PX activities, nonenzymatic MDA activity | tDCS increased DA, SOD and GSH-Px; after MPTP induction, anodal tDCS increased TH and reduced MDA |
| Lee et al., 2018 [45] | 60 Male C57BL/6 mice (n = 15 in control group; n = 15 in anodal tDCS group; n = 15 in MPTP group; n = 15 in MPTP + tDCS group) | Anodal/Sham | AE: left motor cortex; R: between the shoulders | 0.1 mA, 30 min/day, 5 consecutive days AEA: 3.1 mm2 CD: 3.2 mA/cm2 | TH-positive cells; TH; α-synuclein protein; loss of dopaminergic neuron cells; ratio of LC3-II/LC3-I; p62; PI3K; mTOR; AMPK; ULK | tDCS attenuated decrease of TH, p62, mTOR, PI3K, BDNF; attenuated increase of α-synuclein, LC3-II/LC3-I, AMPK and ULK |
| Lee et al., 2019 [17] | Male C57BL/6 mice (number n.r.) | Anodal/Sham | AE: on motor cortex; R: between the shoulders | 0.1 mA, 30 min/day, 5 days/week, 1 week; AEA: 3.1 mm2 CD: 3.2 mA/cm2 | Expression of: TH, mitophagy-related proteins; marker of degradation phase of autophagy; mitochondrial biogenesis-related proteins; mitochondrial fission and fusion -related proteins; ATP concentration. Mitochondrial GDH activity | tDCS preserved neurons and fibers in substantia nigra and striatum; attenuated mitochondrial GDH activity, ATP concentration; increased mitophagy-related and mitochondrial biogenesis proteins |
| Feng et al., 2020 [44] | 16 male Wistar (n = 8 in anodal group; n = 8 in sham group) | Anodal/Sham | AE: skull bregma; R: anterior chest | 300 μA, 20 min/day, 5 days/week, 4 weeks; AEA: 37.9 mm2; CD: 0.16 mA/cm2 | Loss of dopaminergic nigrostriatal neurons and fibers | tDCS preserved neurons in the substantia nigra, but not fibers in the striatum |
| Study | NIBS Method | Sample/Animals | Configuration | Parameters | Biological Outcomes | Biological Results |
|---|---|---|---|---|---|---|
| Tan et al., 2013 [73] | rTMS (LF) | 84 mice (n = 21 in control group; n = 21 rTMS group; n = 21 in Aβ injection; n = 21 Aβ injection + rTMS) | Whole brain stimulation | 400 pulses per session, 7 days/week, 2 weeks LF-rTMS: 20 trains (20 pulses at 1 Hz, 10 s inter-interval) | Neuroplasticity-related proteins (BDNF, NGF and NMDA receptor) levels | LF-rTMS reversed NMDA receptor suppression, enhanced, BDNF and NGF levels |
| Marceglia et al., 2016 [83] | tDCS (anodal/sham) | 7 AD patients (n = 7 tDCS; n = 7 sham) | AE: bilateral temporo-parietal area; R: right arm | 1.5 mA, 15 min/day, 1 day AEA: 25 cm2 CD: 0.06 mA/cm2 | total NO levels | tDCS increased NO levels |
| Khedr et al., 2019 [90] | tDCS (anodal) | 46 AD patients (n = 23 tDCS; n = 23 sham) | AE: bilateral temporo-parietal area; R: left arm | 2 mA, 20 min each side (5 min in between), 5 days/week, 2 weeks AEA: 35 cm2 CD = 0.057 mA/cm2 | AD brain damage biomarkers levels (TAU and Aβ 1-42) | tDCS increased Aβ 1-42 |
| Chen et al., 2020 [74] | rTMS (HF) | 30 mice (n = 15 rTMS; n = 15 sham) | Whole brain stimulation, | 600 pulses per session, 7 days/week, 2 weeks HF-rTMS 20 trains (30 pulses at 5 Hz, 2 s inter-interval) | Synaptic plasticity-related proteins (PSD95), neurotrophic factors (BDNF, TrkB and AKT), autophagy marker proteins (p62 and LC3-II/LC3-I) | HF-rTMS increased BDNF and TrkB levels, and enhanced hippocampal cellular autophagy |
| Choung et al., 2021 [70] | rTMS (HF/LF/sham) | 24 mice (n = 8 HF-rTMS; n = 8 LF-rTMS; n = 8 sham) | Whole brain stimulation | 1600 pulses per session, 5 days/week, 2 weeks HF-rTMS: 40 trains (2 s duration at 20 Hz, 28 s inter-interval) LF-rTMS: continuous stimulation (1 Hz). | BDNF, nestin and neuron protein levels | HF-rTMS increased BDNF, nestin and neuron expression levels in hippocampus and cortex, compared to sham |
| Velioglu et al., 2021 [71] | rTMS (HF) | 15 subjects | Left parietal cortex stimulation | 1640 pulses per session, 5 days/week, 2 weeks HF-rTMS: 42 trains (2 s duration at 20 Hz, 28 s inter-interval) | BDNF and anti-oxidative stress proteins levels | HF-rTMS increased BDNF and anti-oxidative stress proteins levels |
| Luo et al., 2022 [88] | tDCS (anodal/sham) | 33 AD model mice (n = 11 tDCS; n = 11 not treated; n = 11 sham) | AE: frontal cortex; R: thorax | 150 µA, 30 min/day, 5 days/week, 2 weeks AEA:nr CD: nr | Aβ plaques density in the hippocampus and frontal cortex, NVU integrity | tDCS reduced Aβ plaques density and increased BBB integrity-related proteins |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guidetti, M.; Bertini, A.; Pirone, F.; Sala, G.; Signorelli, P.; Ferrarese, C.; Priori, A.; Bocci, T. Neuroprotection and Non-Invasive Brain Stimulation: Facts or Fiction? Int. J. Mol. Sci. 2022, 23, 13775. https://doi.org/10.3390/ijms232213775
Guidetti M, Bertini A, Pirone F, Sala G, Signorelli P, Ferrarese C, Priori A, Bocci T. Neuroprotection and Non-Invasive Brain Stimulation: Facts or Fiction? International Journal of Molecular Sciences. 2022; 23(22):13775. https://doi.org/10.3390/ijms232213775
Chicago/Turabian StyleGuidetti, Matteo, Alessandro Bertini, Francesco Pirone, Gessica Sala, Paola Signorelli, Carlo Ferrarese, Alberto Priori, and Tommaso Bocci. 2022. "Neuroprotection and Non-Invasive Brain Stimulation: Facts or Fiction?" International Journal of Molecular Sciences 23, no. 22: 13775. https://doi.org/10.3390/ijms232213775