
The Role of Transmembrane Proteins in Plant Growth, Development, and Stress Responses

Abstract
:1. Introduction
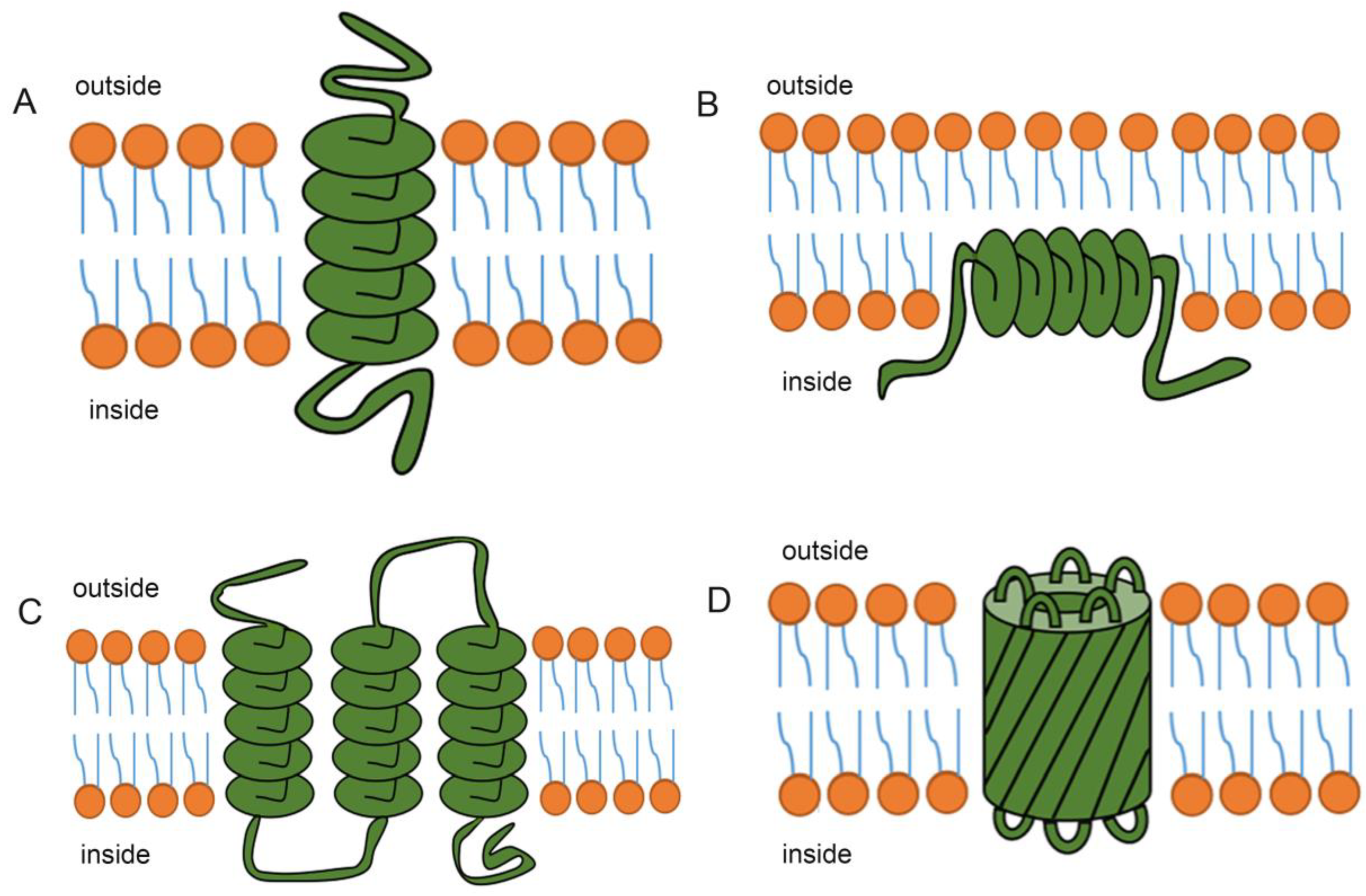
2. The Structure of Transmembrane Proteins
3. Methods Used to Study Transmembrane Proteins
3.1. Prediction by the Classical Kyte and Doolittle Method
3.2. Prediction Using the “Positive Charge Residence Rule”
3.3. Prediction Based on Hidden Markov Models
3.4. Statistical Analysis and Model Parameter Prediction
3.5. Prediction by Wavelet Analysis
4. The Role of Transmembrane Proteins in Plant Growth and Development
5. Transmembrane Proteins Participate in Various Stress Responses
6. Response to Salt Stress
7. Response to Drought Stress
8. Response to Cold and Heat Stress
9. Responsive to Biotic Stresses
10. Future Perspectives
11. Application of Transmembrane Protein in Crop Breeding
12. Receptor Proteins for Various External Stimuli
13. Interactions between Transmembrane Proteins and Intracellular Proteins
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Marinko, J.T.; Huang, H.; Penn, W.D.; Capra, J.A.; Schlebach, J.P.; Sanders, C.R. Folding and misfolding of human membrane proteins in health and disease: From single molecules to cellular proteostasis. Chem. Rev. 2019, 119, 5537–5606. [Google Scholar] [CrossRef] [PubMed]
- Goossens, K.; De Winter, H. Molecular dynamics simulations of membrane proteins: An overview. J. Chem. Inf. Model. 2018, 58, 2193–2202. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Li, J.; Sun, J.; Wang, L.; Fan, C.; Shen, J. Recent Advances of DNA Nanostructure-Based Cell Membrane Engineering. Adv. Healthc. Mater. 2021, 10, e2001718. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, E.P.; Beis, K.; Cameron, A.D.; Iwata, S. Overcoming the challenges of membrane protein crystallography. Curr. Opin. Struct. Biol. 2008, 18, 581–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esseghir, S.; Reis-Filho, J.S.; Kennedy, A.; James, M.; O′Hare, M.J.; Jeffery, R.; Poulsom, R.; Isacke, C.M. Identification of transmembrane proteins as potential prognostic markers and therapeutic targets in breast cancer by a screen for signal sequence encoding transcripts. J. Pathol. 2006, 210, 420–430. [Google Scholar] [CrossRef] [PubMed]
- Breton, G.; Danyluk, J.; Charron, J.B.; Sarhan, F. Expression profiling and bioinformatic analyses of a novel stress-regulated multispanning transmembrane protein family from cereals and Arabidopsis. Plant Physiol. 2003, 132, 64–74. [Google Scholar] [CrossRef] [Green Version]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [Green Version]
- Fleishman, S.J.; Unger, V.M.; Ben-Tal, N. Transmembrane protein structures without X-rays. Trends Biochem. Sci. 2006, 31, 106–113. [Google Scholar] [CrossRef]
- Muller, M.P.; Jiang, T.; Sun, C.; Lihan, M.; Pant, S.; Mahinthichaichan, P.; Trifan, A.; Tajkhorshid, E. Characterization of lipid–protein interactions and lipid-mediated modulation of membrane protein function through molecular simulation. Chem. Rev. 2019, 119, 6086–6161. [Google Scholar] [CrossRef]
- Nakao, H.; Sugimoto, Y.; Ikeda, K.; Saito, H.; Nakano, M. Structural feature of lipid scrambling model transmembrane peptides: Same-side positioning of hydrophilic residues and their deeper position. J. Phys. Chem. Lett. 2020, 11, 1662–1667. [Google Scholar] [CrossRef]
- Nawrocki, G.; Im, W.; Sugita, Y.; Feig, M. Clustering and dynamics of crowded proteins near membranes and their influence on membrane bending. Proc. Natl. Acad. Sci. USA 2019, 116, 24562–24567. [Google Scholar] [CrossRef] [PubMed]
- Corradi, V.; Mendez-Villuendas, E.; Ingólfsson, H.I.; Gu, R.X.; Siuda, I.; Melo, M.N.; Moussatova, A.; DeGagné, L.J.; Sejdiu, B.I.; Singh, G.; et al. Lipid–protein interactions are unique fingerprints for membrane proteins. ACS Cent. Sci. 2018, 4, 709–717. [Google Scholar] [CrossRef] [PubMed]
- Nakao, H.; Hayashi, C.; Ikeda, K.; Saito, H.; Nagao, H.; Nakano, M. Effects of hydrophilic residues and hydrophobic length on flip-flop promotion by transmembrane peptides. J. Phys. Chem. B 2018, 122, 4318–4324. [Google Scholar] [CrossRef] [PubMed]
- Rich, P.R.; Maréchal, A. Functions of the hydrophilic channels in protonmotive cytochrome c oxidase. J. R. Soc. Interface 2013, 10, e20130183. [Google Scholar] [CrossRef] [Green Version]
- Becker, O.M.; Marantz, Y.; Shacham, S.; Inbal, B.; Heifetz, A.; Kalid, O.; Bar-Haim, S.; Warshaviak, D.; Fichman, M.; Noiman, S. G protein-coupled receptors: In silico drug discovery in 3D. Proc. Natl. Acad. Sci. USA 2004, 101, 11304–11309. [Google Scholar] [CrossRef] [Green Version]
- Heng, B.C.; Aubel, D.; Fussenegger, M. An overview of the diverse roles of G-protein coupled receptors (GPCRs) in the pathophysiology of various human diseases. Biotechnol. Adv. 2013, 31, 1676–1694. [Google Scholar] [CrossRef]
- Qi, X.; Gao, J.; Li, Z.; Zhang, G.; Li, J.; Fu, Y.; Cai, M.; Wang, H.; Tong, T. CDCP1: A promising diagnostic biomarker and therapeutic target for human cancer. Life Sci. 2022, 301, 120600. [Google Scholar] [CrossRef]
- Hanton, S.L.; Renna, L.; Bortolotti, L.E.; Chatre, L.; Stefano, G.; Brandizzi, F. Diacidic motifs influence the export of transmembrane proteins from the endoplasmic reticulum in plant cells. Plant Cell 2005, 17, 3081–3093. [Google Scholar] [CrossRef] [Green Version]
- Jin, L.; Li, R.; Cao, H.; Yan, Y. Identification and Expression Analysis of Fatty Acid Export Gene FAX1 in Oil Palm (Elaeis guineensis). Chin. J. Trop. Crops 2022, 43, 27–33. [Google Scholar]
- Kleczkowski, L.A.; Igamberdiev, A.U. Magnesium signaling in plants. Int. J. Mol. Sci. 2021, 22, 1159. [Google Scholar] [CrossRef]
- Corradi, V.; Sejdiu, B.I.; Mesa-Galloso, H.; Abdizadeh, H.; Noskov, S.Y.; Marrink, S.J.; Tieleman, D.P. Emerging diversity in lipid–protein interactions. Chem. Rev. 2019, 119, 5775–5848. [Google Scholar] [CrossRef] [PubMed]
- Barrera, E.E.; Machado, M.R.; Pantano, S. Fat SIRAH: Coarse-grained phospholipids to explore membrane–protein dynamics. J. Chem. Theory Comput. 2019, 15, 5674–5688. [Google Scholar] [CrossRef] [PubMed]
- Renne, M.F.; de Kroon, A.I.P.M. The role of phospholipid molecular species in determining the physical properties of yeast membranes. FEBS Lett. 2018, 592, 1330–1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thoma, J.; Manioglu, S.; Kalbermatter, D.; Bosshart, P.D.; Fotiadis, D.; Müller, D.J. Protein-enriched outer membrane vesicles as a native platform for outer membrane protein studies. Commun. Biol. 2018, 1, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulmschneider, M.B.; Sansom, M.S.P. Amino acid distributions in integral membrane protein structures. Biochim. Biophys. Acta 2001, 1512, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Bordag, N.; Keller, S. α-Helical transmembrane peptides: A “divide and conquer” approach to membrane proteins. Chem. Phys. Lipids 2010, 163, 1–26. [Google Scholar] [CrossRef]
- Harrison, R.S.; Shepherd, N.E.; Hoang, H.N.; Ruiz-Gómez, G.; Hill, T.A.; Driver, R.W.; Desai, V.S.; Young, P.R.; Abbenante, G.; Fairlie, D.P. Downsizing human, bacterial, and viral proteins to short water-stable alpha helices that maintain biological potency. Proc. Natl. Acad. Sci. USA 2010, 107, 11686–11691. [Google Scholar] [CrossRef] [Green Version]
- Lundbaek, J.A.; Collingwood, S.A.; Ingólfsson, H.I.; Kapoor, R.; Andersen, O.S. Lipid bilayer regulation of membrane protein function: Gramicidin channels as molecular force probes. J. R. Soc. Interface 2010, 7, 373–395. [Google Scholar] [CrossRef] [Green Version]
- Langosch, D.; Brosig, B.; Kolmar, H.; Fritz, H.J. Dimerisation of the glycophorin A transmembrane segment in membranes probed with the ToxR transcription activator. J. Mol. Biol. 1996, 263, 525–530. [Google Scholar] [CrossRef]
- Baeza-Delgado, C.; Marti-Renom, M.A.; Mingarro, I. Structure-based statistical analysis of transmembrane helices. Eur. Biophys. J. 2013, 42, 199–207. [Google Scholar] [CrossRef] [Green Version]
- Shinoda, K.; Shinoda, W.; Mikami, M. Molecular dynamics simulation of an archaeal lipid bilayer with sodium chloride. Phys. Chem. Chem. Phys. 2007, 9, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Almén, M.S.; Nordström, K.J.; Fredriksson, R.; Schiöth, H.B. Mapping the human membrane proteome: A majority of the human membrane proteins can be classified according to function and evolutionary origin. BMC Biol. 2009, 7, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bethani, I.; Skånland, S.S.; Dikic, I.; Acker-Palmer, A. Spatial organization of transmembrane receptor signalling. EMBO J. 2010, 29, 2677–2688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Remorino, A.; Hochstrasser, R.M. Three-dimensional structures by two-dimensional vibrational spectroscopy. Acc. Chem. Res. 2012, 45, 1896–1905. [Google Scholar] [CrossRef] [Green Version]
- Fairman, J.W.; Noinaj, N.; Buchanan, S.K. The structural biology of β-barrel membrane proteins: A summary of recent reports. Curr. Opin. Struct. Biol. 2011, 21, 523–531. [Google Scholar] [CrossRef] [Green Version]
- Fujiyoshi, Y.; Mitsuoka, K.; de Groot, B.L.; Philippsen, A.; Grubmüller, H.; Agre, P.; Engel, A. Structure and function of water channels. Curr. Opin. Struct. Biol. 2002, 12, 509–515. [Google Scholar] [CrossRef] [Green Version]
- Del Piccolo, N.; Sarabipour, S.; Hristova, K. A new method to study heterodimerization of membrane proteins and its application to fibroblast growth factor receptors. J. Biol. Chem. 2017, 292, 1288–1301. [Google Scholar] [CrossRef] [Green Version]
- Fink, A.; Sal-Man, N.; Gerber, D.; Shai, Y. Transmembrane domains interactions within the membrane milieu: Principles, advances and challenges. Biochim. Biophys. Acta 2012, 1818, 974–983. [Google Scholar] [CrossRef] [Green Version]
- Zagnoni, M. Miniaturised technologies for the development of artificial lipid bilayer systems. Lab Chip 2012, 12, 1026–1039. [Google Scholar] [CrossRef]
- Tamm, L.K.; Hong, H.; Liang, B. Folding and assembly of β-barrel membrane proteins. Biochim. Biophys. Acta 2004, 1666, 250–263. [Google Scholar] [CrossRef] [Green Version]
- Schulz, G.E. β-barrel membrane proteins. Curr. Opin. Struct. Biol. 2000, 10, 443–447. [Google Scholar] [CrossRef]
- McMorran, L.M.; Brockwell, D.J.; Radford, S.E. Mechanistic studies of the biogenesis and folding of outer membrane proteins in vitro and in vivo: What have we learned to date? Arch. Biochem. Biophys. 2014, 564, 265–280. [Google Scholar] [CrossRef] [Green Version]
- Yarov-Yarovoy, V.; Schonbrun, J.; Baker, D. Multipass membrane protein structure prediction using Rosetta. Proteins Struct. Funct. Bioinform. 2006, 62, 1010–1025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tusnády, G.E.; Dosztányi, Z.; Simon, I. Transmembrane proteins in the Protein Data Bank: Identification and classification. Bioinformatics 2004, 20, 2964–2972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, J.; Stevens, T.J.; Samsó, M. Membrane proteins: The ‘Wild West’of structural biology. Trends Biochem. Sci. 2003, 28, 137–144. [Google Scholar] [CrossRef]
- Chou, K.C.; Zhang, C.T. Prediction of protein structural classes. Crit. Rev. Biochem. Mol. Biol. 1995, 30, 275–349. [Google Scholar] [CrossRef]
- Kyte, J.; Doolittle, R.F. A simple method for displaying the hydropathic character of a protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef] [Green Version]
- Von Heijne, G. Membrane protein structure prediction: Hydrophobicity analysis and the positive-inside rule. J. Mol. Biol. 1992, 225, 487–494. [Google Scholar] [CrossRef]
- Sonnhammer, E.L.; Von Heijne, G.; Krogh, A. A hidden Markov model for predicting transmembrane helices in protein sequences. Proc. Int. Conf. Intell. Syst. Mol. Biol. 1998, 6, 175–182. [Google Scholar]
- Nugent, T.; Ward, S.; Jones, D.T. The MEMPACK alpha-helical transmembrane protein structure prediction server. Bioinformatics 2011, 27, 1438–1439. [Google Scholar] [CrossRef] [Green Version]
- Torrence, C.; Compo, G.P. A practical guide to wavelet analysis. Bull. Am. Meteorol. Soc. 1998, 79, 61–78. [Google Scholar] [CrossRef]
- Rost, B.; Sander, C.; Casadio, R.; Fariselli, P. Transmembrane helices predicted at 95% accuracy. Protein Sci. 1995, 4, 521–533. [Google Scholar] [CrossRef] [PubMed]
- Möller, S.; Croning MD, R.; Apweiler, R. Evaluation of methods for the prediction of membrane spanning regions. Bioinformatics 2001, 17, 646–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. A model recognition approach to the prediction of all-helical membrane protein structure and topology. Biochemistry 1994, 33, 3038–3049. [Google Scholar] [CrossRef] [PubMed]
- Argos, P.; Persson, B. Topology prediction of membrane proteins. Protein Sci. 1996, 5, 363–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Persson, B.; Argos, P. Prediction of transmembrane segments in proteins utilising multiple sequence alignments. J. Mol. Biol. 1994, 237, 182–192. [Google Scholar] [CrossRef]
- Banach, M.; Fabian, P.; Stapor, K.; Konieczny, L.; Roterman, A.I. Structure of the hydrophobic core determines the 3D protein structure—Verification by single mutation proteins. Biomolecules 2020, 10, 767. [Google Scholar] [CrossRef]
- Schwacke, R.; Schneider, A.; van der Graaff, E.; Fischer, K.; Catoni, E.; Desimone, M.; Frommer, W.B.; Flügge, U.I.; Kunze, R. ARAMEMNON, a novel database for Arabidopsis integral membrane proteins. Plant Physiol. 2003, 131, 16–26. [Google Scholar] [CrossRef] [Green Version]
- Käll, L.; Sonnhammer, E.L.L. Reliability of transmembrane predictions in whole-genome data. FEBS Lett. 2002, 532, 415–418. [Google Scholar] [CrossRef] [Green Version]
- Viklund, H.; Elofsson, A. Best α-helical transmembrane protein topology predictions are achieved using hidden Markov models and evolutionary information. Protein Sci. 2004, 13, 1908–1917. [Google Scholar] [CrossRef]
- Cserzö, M.; Wallin, E.; Simon, I.; von Heijne, G.; Elofsson, A. Prediction of transmembrane alpha-helices in prokaryotic membrane proteins: The dense alignment surface method. Protein Eng. 1997, 10, 673–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirakawa, H.; Muta, S.; Kuhara, S. The hydrophobic cores of proteins predicted by wavelet analysis. Bioinformatics 1999, 15, 141–148. [Google Scholar] [CrossRef]
- Mandell, A.J.; Selz, K.A.; Shlesinger, M.F. Wavelet transformation of protein hydrophobicity sequences suggests their memberships in structural families. Phys. A Stat. Mech. Its Appl. 1997, 244, 254–262. [Google Scholar] [CrossRef]
- Sasidharan, K.; Tomita, M.; Aon, M.; Lloyd, D.; Murray, D.B. Time-structure of the yeast metabolism in vivo. In Advances in Systems Biology; Springer: New York, NY, USA, 2012; pp. 359–379. [Google Scholar]
- Qiu, J.; Liang, R.; Zou, X.; Mo, J. Prediction of transmembrane proteins based on the continuous wavelet transform. J. Chem. Inf. Comput. Sci. 2004, 44, 741–747. [Google Scholar] [CrossRef]
- Qiu, J.; Liang, R.; Zou, X.; Mo, J. Prediction of protein secondary structure based on continuous wavelet transform. Talanta 2003, 61, 285–293. [Google Scholar] [CrossRef]
- Cai, F.; Sun, F.; Dai, H.; Zhu, X.; Zhang, L. Application of wavelet and Fourier transform in time series analysis. GNSS World China 2019, 44, 40–46. [Google Scholar]
- Kiefer, D.; Kuhn, A. Hydrophobic forces drive spontaneous membrane insertion of the bacteriophage Pf3 coat protein without topological control. EMBO J. 1999, 18, 6299–6306. [Google Scholar] [CrossRef]
- Harley, C.A.; Holt, J.A.; Turner, R.; Tipper, D.J. Transmembrane protein insertion orientation in yeast depends on the charge difference across transmembrane segments, their total hydrophobicity, and its distribution. J. Biol. Chem. 1998, 273, 24963–24971. [Google Scholar] [CrossRef] [Green Version]
- Sharpe, H.J.; Stevens, T.J.; Munro, S. A comprehensive comparison of transmembrane domains reveals organelle-specific properties. Cell 2010, 142, 158–169. [Google Scholar] [CrossRef] [Green Version]
- Kraffe, E.; Marty, Y.; Guderley, H. Changes in mitochondrial oxidative capacities during thermal acclimation of rainbow trout Oncorhynchus mykiss: Roles of membrane proteins, phospholipids and their fatty acid compositions. J. Exp. Biol. 2007, 210, 149–165. [Google Scholar] [CrossRef] [Green Version]
- Funakoshi, K.; Suzuki, H.; Takeuchi, S. Lipid bilayer formation by contacting monolayers in a microfluidic device for membrane protein analysis. Anal. Chem. 2006, 78, 8169–8174. [Google Scholar] [CrossRef] [PubMed]
- Kelkar, D.A.; Chattopadhyay, A. The gramicidin ion channel: A model membrane protein. Biochim. Biophys. Acta 2007, 1768, 2011–2025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saier, M.H., Jr. A functional-phylogenetic classification system for transmembrane solute transporters. Microbiol. Mol. Biol. Rev. 2000, 64, 354–411. [Google Scholar] [CrossRef] [PubMed]
- Clark, H.F.; Gurney, A.L.; Abaya, E.; Baker, K.; Baldwin, D.; Brush, J.; Chen, J.; Chow, B.; Chui, C.; Crowley, C.; et al. The secreted protein discovery initiative (SPDI), a large-scale effort to identify novel human secreted and transmembrane proteins: A bioinformatics assessment. Genome Res. 2003, 13, 2265–2270. [Google Scholar] [CrossRef] [Green Version]
- Ehlers, M.R.W.; Riordan, J.F. Membrane proteins with soluble counterparts: Role of proteolysis in the release of transmembrane proteins. Biochemistry 1991, 30, 10065–10074. [Google Scholar] [CrossRef] [PubMed]
- Nühse, T.S.; Bottrill, A.R.; Jones, A.M.; Peck, S.C. Quantitative phosphoproteomic analysis of plasma membrane proteins reveals regulatory mechanisms of plant innate immune responses. Plant J. 2007, 51, 931–940. [Google Scholar] [CrossRef] [Green Version]
- Xue, S.X.; Jia, M.; Yu, F. MATE transporter gene of Zea mays regulates plant development and hypocotyl elongation. J. Northwest A F Univ. Nat. Sci. Ed. 2018, 46, 29–38. [Google Scholar]
- Wang, L.; Bei, X.; Gao, J.; Li, Y.; Yan, Y.; Hu, Y. The similar and different evolutionary trends of MATE family occurred between rice and Arabidopsis thaliana. BMC Plant Biol. 2016, 16, 207. [Google Scholar] [CrossRef] [Green Version]
- Du, Z.; Su, Q.; Wu, Z.; Huang, Z.; Bao, J.; Li, J.; Tu, H.; Zeng, C.; Fu, J.; He, H. Genome-wide characterization of MATE gene family and expression profiles in response to abiotic stresses in rice (Oryza sativa). BMC Ecol. Evol. 2021, 21, 141. [Google Scholar] [CrossRef]
- Li, G.W.; Peng, Y.H.; Yu, X.; Zhang, M.H.; Cai, W.M.; Sun, W.N.; Su, W.A. Transport functions and expression analysis of vacuolar membrane aquaporins in response to various stresses in rice. J. Plant Physiol. 2008, 165, 1879–1888. [Google Scholar] [CrossRef]
- Johansson, I.; Karlsson, M.; Johanson, U.; Larsson, C.; Kjellbom, P. The role of aquaporins in cellular and whole plant water balance. Biochim. Biophys. Acta 2000, 1465, 324–342. [Google Scholar] [CrossRef] [Green Version]
- An, J.; Cheng, C.; Hu, Z.; Chen, H.; Cai, W.; Yu, B. The Panax ginseng PgTIP1 gene confers enhanced salt and drought tolerance to transgenic soybean plants by maintaining homeostasis of water, salt ions and ROS. Environ. Exp. Bot. 2018, 155, 45–55. [Google Scholar] [CrossRef]
- Sudhakaran, S.; Thakral, V.; Padalkar, G.; Rajora, N.; Dhiman, P.; Raturi, G.; Sharma, Y.; Tripathi, D.K.; Deshmukh, R.; Sharma, T.R. Significance of solute specificity, expression, and gating mechanism of tonoplast intrinsic protein during development and stress response in plants. Physiol. Plant. 2021, 172, 258–274. [Google Scholar] [CrossRef]
- Diener, A.C.; Gaxiola, R.A.; Fink, G.R. Arabidopsis ALF5, a multidrug efflux transporter gene family member, confers resistance to toxins. Plant Cell 2001, 13, 1625–1638. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Peng, Y.; Li, G.; Arora, R.; Tang, Z.; Su, W.; Cai, W. Isolation and functional characterization of PgTIP1, a hormone-autotrophic cells-specific tonoplast aquaporin in ginseng. J. Exp. Bot. 2007, 58, 947–956. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Cai, W. A ginseng PgTIP1 gene whose protein biological activity related to Ser128 residue confers faster growth and enhanced salt stress tolerance in Arabidopsis. Plant Sci. 2015, 234, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Boutrot, F.; Chantret, N.; Gautier, M.F. Genome-wide analysis of the rice and Arabidopsis non-specific lipid transfer protein (nsLtp) gene families and identification of wheat nsLtp genes by EST data mining. BMC Genom. 2008, 9, 86. [Google Scholar] [CrossRef] [Green Version]
- Ramesh, S.A.; Tyerman, S.D.; Xu, B.; Bose, J.; Kaur, S.; Conn, V.; Domingos, P.; Ullah, S.; Wege, S.; Shabala, S. GABA signalling modulates plant growth by directly regulating the activity of plant-specific anion transporters. Nat. Commun. 2015, 6, 7879. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Gügel, I.L.; Giavalisco, P.; Zeisler, V.; Schreiber, L.; Soll, J.; Philippar, K. FAX1, a novel membrane protein mediating plastid fatty acid export. PLoS Biol. 2015, 13, e1002053. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, M.; Sato, Y.; Wu, S.; Kang, B.H.; McCarty, D.R. Conserved functions of the MATE transporter BIG EMBRYO1 in regulation of lateral organ size and initiation rate. Plant Cell 2015, 27, 2288–2300. [Google Scholar] [CrossRef] [Green Version]
- Suhail, M. Na+, K+-ATPase: Ubiquitous multifunctional transmembrane protein and its relevance to various pathophysiological conditions. J. Clin. Med. Res. 2010, 2, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romanov, G.A.; Lomin, S.N.; Schmülling, T. Cytokinin signaling: From the ER or from the PM? That is the question! New Phytol. 2018, 218, 41–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, B.; Xue, X.Y.; Hu, W.L.; Wang, L.J.; Chen, X.Y. An ABC transporter gene of Arabidopsis thaliana, AtWBC11, is involved in cuticle development and prevention of organ fusion. Plant Cell Physiol. 2007, 48, 1790–1802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ung, K.L.; Winkler, M.; Schulz, L.; Kolb, M.; Janacek, D.P.; Dedic, E.; Stokes, D.L.; Hammes, U.Z.; Pedersen, B.P. Structures and mechanism of the plant PIN-FORMED auxin transporter. Nature 2022, 609, 605–610. [Google Scholar] [CrossRef]
- Tena, G. PIN finally up. Nat. Plants 2022, 8, 725. [Google Scholar] [CrossRef]
- Gomes, G.L.B.; Scortecci, K.C. Auxin and its role in plant development: Structure, signalling, regulation and response mechanisms. Plant Biol. 2021, 23, 894–904. [Google Scholar] [CrossRef]
- Courville, P.; Chaloupka, R.; Cellier, M.F.M. Recent progress in structure–function analyses of Nramp proton-dependent metal-ion transporters. Biochem. Cell Biol. 2006, 84, 960–978. [Google Scholar] [CrossRef]
- Mani, A.; Sankaranarayanan, K. In silico analysis of natural resistance-associated macrophage protein (NRAMP) family of transporters in rice. Protein J. 2018, 37, 237–247. [Google Scholar] [CrossRef]
- Qin, L.; Han, P.; Chen, L.; Walk, T.C.; Li, Y.; Hu, X.; Xie, L.; Liao, H.; Liao, X. Genome-wide identification and expression analysis of NRAMP family genes in soybean (Glycine max L.). Front. Plant Sci. 2017, 8, 1436. [Google Scholar] [CrossRef] [Green Version]
- Segond, D.; Dellagi, A.; Lanquar, V.; Rigault, M.; Patrit, O.; Thomine, S.; Expert, D. NRAMP genes function in Arabidopsis thaliana resistance to Erwinia chrysanthemi infection. Plant J. 2009, 58, 195–207. [Google Scholar] [CrossRef]
- Li, L.; Zhu, Z.; Liao, Y.; Yang, C.; Fan, N.; Zhang, J.; Yamaji, N.; Dirick, L.; Ma, J.F.; Curie, C.; et al. NRAMP6 and NRAMP1 cooperatively regulate root growth and manganese translocation under manganese deficiency in Arabidopsis. Plant J. 2022, 110, 1564–1577. [Google Scholar] [CrossRef] [PubMed]
- Osakabe, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Tran, L.S. Sensing the environment: Key roles of membrane-localized kinases in plant perception and response to abiotic stress. J. Exp. Bot. 2013, 64, 445–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmid, C.; Bauer, S.; Müller, B.; Bartelheimer, M. Belowground neighbor perception in Arabidopsis thaliana studied by transcriptome analysis: Roots of Hieracium pilosella cause biotic stress. Front. Plant. Sci. 2013, 13, 296. [Google Scholar] [CrossRef] [Green Version]
- Rawat, N.; Singla-Pareek, S.L.; Pareek, A. Membrane dynamics during individual and combined abiotic stresses in plants and tools to study the same. Physiol. Plant. 2021, 171, 653–676. [Google Scholar] [CrossRef] [PubMed]
- Rathor, P.; Borza, T.; Liu, Y.; Qin, Y.; Stone, S.; Zhang, J.; Hui, J.P.M.; Berrue, F.; Groisillier, A.; Tonon, T.; et al. Low mannitol concentrations in Arabidopsis thaliana expressing Ectocarpus genes improve salt tolerance. Plants 2020, 9, 1508. [Google Scholar] [CrossRef]
- Al-Saharin, R.; Hellmann, H.; Mooney, S. Plant E3 Ligases and Their Role in Abiotic Stress Response. Cells 2022, 11, 890. [Google Scholar] [CrossRef]
- Tian, M.; Lou, L.; Liu, L.; Yu, F.; Zhao, Q.; Zhang, H.; Wu, Y.; Tang, S.; Xia, R.; Zhu, B.; et al. The RING finger E3 ligase STRF1 is involved in membrane trafficking and modulates salt-stress response in Arabidopsis thaliana. Plant J. 2015, 82, 81–92. [Google Scholar] [CrossRef]
- Isono, E.; Kalinowska, K. ESCRT-dependent degradation of ubiquitylated plasma membrane proteins in plants. Curr. Opin. Plant Biol. 2017, 40, 49–55. [Google Scholar] [CrossRef]
- Liu, H.; Yang, W.; Zhao, X.; Kang, G.; Li, N.; Xu, H. Genome-wide analysis and functional characterization of CHYR gene family associated with abiotic stress tolerance in bread wheat (Triticum aestivum L.). BMC Plant Biol. 2022, 22, 204. [Google Scholar] [CrossRef]
- He, F.; Wang, H.L.; Li, H.G.; Su, Y.; Li, S.; Yang, Y.; Feng, C.H.; Yin, W.; Xia, X. Pe CHYR 1, a ubiquitin E3 ligase from Populus euphratica, enhances drought tolerance via ABA-induced stomatal closure by ROS production in Populus. Plant Biotechnol. J. 2018, 16, 1514–1528. [Google Scholar] [CrossRef] [Green Version]
- Doroodian, P.; Hua, Z. The ubiquitin switch in plant stress response. Plants 2021, 10, 246. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.H.; Kim, S.H.; Berberich, T.; Kusano, T. Identification and properties of a small protein that interacts with a tobacco bZIP-type transcription factor TBZF. Plant Biotechnol. 2012, 29, 395–399. [Google Scholar] [CrossRef]
- Maurel, C.; Chrispeels, M.J. Aquaporins. A molecular entry into plant water relations. Plant Physiol. 2001, 125, 135–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, C.; Chen, K.; Ding, Q.; Mou, Y.; Yang, R.; Zhao, M.; Ma, B.; Xu, Z.; Ma, Y.; Pan, Y. Proteomic analysis of the function of a novel cold-regulated multispanning transmembrane protein COR413-PM1 in Arabidopsis. Int. J. Mol. Sci. 2018, 19, 2572. [Google Scholar] [CrossRef] [PubMed]
- Eisenbarth, D.A.; Weig, A.R. Dynamics of aquaporins and water relations during hypocotyl elongation in Ricinus communis L. seedlings. J. Exp. Bot. 2005, 56, 1831–1842. [Google Scholar] [CrossRef] [Green Version]
- Zou, Z.; Yang, L.; Gong, J.; Mo, Y.; Wang, J.; Cao, J.; An, F.; Xie, G. Genome-wide identification of Jatropha curcas aquaporin genes and the comparative analysis provides insights into the gene family expansion and evolution in Hevea brasiliensis. Front. Plant Sci. 2016, 7, 395. [Google Scholar] [CrossRef] [Green Version]
- Zha, Z.; Wang, D.; Hong, W.; Liu, L.; Zhou, S.; Feng, X.; Qin, B.; Wang, J.; Yang, Y.; Du, L.; et al. Influence of europium speciation on its accumulation in Brassica napus and over-expressing BnTR1 lines. J. Radioanal. Nucl. Chem. 2014, 301, 257–262. [Google Scholar] [CrossRef]
- Bo, S.; Yuanyou, Y.; Jiali, L.; Jijun, Y.; Jun, T.; Ning, L. The Influences of BnTR1 on Ca2+ Absorption of Rapes: Preliminary Investigation by 45 Ca. J. Nucl. Agric. Sci. 2015, 29, 1366. [Google Scholar]
- Liu, Z.B.; Wang, J.M.; Yang, F.X.; Yang, L.; Yue, Y.F.; Xiang, J.B.; Gao, M.; Xiong, F.J.; Lv, D.; Wu, X.J.; et al. A novel membrane-bound E3 ubiquitin ligase enhances the thermal resistance in plants. Plant Biotechnol. J. 2014, 12, 93–104. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Zhang, Y.; Ding, P.; Johnson, K.; Li, X.; Zhang, Y. The ankyrin-repeat transmembrane protein BDA1 functions downstream of the receptor-like protein SNC2 to regulate plant immunity. Plant Physiol. 2012, 159, 1857–1865. [Google Scholar] [CrossRef] [Green Version]
- Sanderfoot, A. Increases in the number of SNARE genes parallels the rise of multicellularity among the green plants. Plant Physiol. 2007, 144, 6–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Deng, M.; Yang, C.; Yu, Q.; Zhang, L.; Zhu, Q.; Guo, X. A Qa-SNARE complex contributes to soybean cyst nematode resistance via regulation of mitochondria-mediated cell death. J. Exp. Bot. 2021, 72, 7145–7162. [Google Scholar] [CrossRef] [PubMed]
- Bai, Z.Y.; Wang, T.; Wu, Y.H.; Wang, K.; Liang, Q.Y.; Pan, Y.Z.; Jiang, B.B.; Zhang, L.; Liu, G.L.; Jia, Y.; et al. Whole-transcriptome sequence analysis of differentially expressed genes in Phormium tenax under drought stress. Sci. Rep. 2017, 7, 41700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miura, K.; Ashikari, M.; Matsuoka, M. The role of QTLs in the breeding of high-yielding rice. Trends Plant Sci. 2011, 16, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Shi, G.; Schwartz, O.; Compton, A.A. More than meets the I: The diverse antiviral and cellular functions of interferon-induced transmembrane proteins. Retrovirology 2017, 14, 53. [Google Scholar] [CrossRef]
- Ye, Y.; Ding, Y.; Jiang, Q.; Wang, F.; Sun, J.; Zhu, C. The role of receptor-like protein kinases (RLKs) in abiotic stress response in plants. Plant Cell Rep. 2017, 36, 235–242. [Google Scholar] [CrossRef]
- Jose, J.; Ghantasala, S.; Roy Choudhury, S. Arabidopsis transmembrane receptor-like kinases (RLKs): A bridge between extracellular signal and intracellular regulatory machinery. Int. J. Mol. Sci. 2020, 21, 4000. [Google Scholar] [CrossRef]
- Shpak, E.D. Diverse roles of ERECTA family genes in plant development. J. Integr. Plant Biol. 2013, 55, 1238–1250. [Google Scholar] [CrossRef]
- Song, W.Y.; Wang, G.L.; Chen, L.L.; Kim, H.S.; Pi, L.Y.; Holsten, T.; Gardner, J.; Wang, B.; Zhai, W.X.; Zhu, L.H.; et al. A receptor kinase-like protein encoded by the rice disease resistance gene, Xa21. Science 1995, 270, 1804–1806. [Google Scholar] [CrossRef] [Green Version]
- She, J.; Han, Z.; Kim, T.W.; Wang, J.; Cheng, W.; Chang, J.; Shi, S.; Wang, J.; Yang, M.; Wang, Z.Y.; et al. Structural insight into brassinosteroid perception by BRI1. Nature 2011, 474, 472–476. [Google Scholar] [CrossRef] [Green Version]
- Tang, W.; Kim, T.W.; Oses-Prieto, J.A.; Sun, Y.; Deng, Z.; Zhu, S.; Wang, R.; Burlingame, A.L.; Wang, Z.Y. BSKs mediate signal transduction from the receptor kinase BRI1 in Arabidopsis. Science 2008, 321, 557–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.B.; Liu, C.; Tang, D.Y.; Yan, L.; Wang, D.; Yang, Y.Z.; Gui, J.S.; Zhao, X.Y.; Li, L.G.; Tang, X.D.; et al. The receptor-like cytoplasmic kinase STRK1 phosphorylates and activates CatC, thereby regulating H2O2 homeostasis and improving salt tolerance in rice. Plant Cell 2018, 30, 1100–1118. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.L.; Xue, H.W. Global analysis of expression profiles of rice receptor-like kinase genes. Mol. Plant 2012, 5, 143–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Yan, L.; Tang, D.Y. Molecular mechanism of receptor-like kinases involved in the response to salt stress in rice. In Proceedings of the 2018 Proceedings of the National Congress of Plant Biology, Taian, China, 18 October 2018. [Google Scholar]
- Lu, K.; Chen, T.; Yao, S.; Liang, W.; Wei, X.; Zhang, Y.; Wang, C. Functional analysis on four receptor-like protein kinases under salt stress in Rice. Chin. J. Rice Sci. 2021, 35, 103–111. [Google Scholar]
- Pandey, S. Heterotrimeric G-protein signaling in plants: Conserved and novel mechanisms. Annu. Rev. Plant Biol. 2019, 70, 213–238. [Google Scholar] [CrossRef] [PubMed]
- Patel, J.S.; Selvaraj, V.; Gunupuru, L.R.; Kharwar, R.N.; Sarma, B.K. Plant G-protein signaling cascade and host defense. 3 Biotech 2020, 10, 219. [Google Scholar] [CrossRef]
- DeFalco, T.A.; Bender, K.W.; Snedden, W.A. Breaking the code: Ca2+ sensors in plant signalling. Biochem. J. 2010, 425, 27–40. [Google Scholar] [CrossRef] [Green Version]
- Meisrimler, C.N.; Planchon, S.; Renaut, J.; Sergeant, K.; Lüthje, S. Alteration of plasma membrane-bound redox systems of iron deficient pea roots by chitosan. J. Proteom. 2011, 74, 1437–1449. [Google Scholar] [CrossRef]
- Perfus-Barbeoch, L.; Jones, A.M.; Assmann, S.M. Plant heterotrimeric G protein function: Insights from Arabidopsis and rice mutants. Curr. Opin. Plant Biol. 2004, 7, 719–731. [Google Scholar] [CrossRef]
- Jones, A.M.; Assmann, S.M. Plants: The latest model system for G-protein research. EMBO Rep. 2004, 5, 572–578. [Google Scholar] [CrossRef] [Green Version]
- Testerink, C.; Munnik, T. Phosphatidic acid: A multifunctional stress signaling lipid in plants. Trends Plant Sci. 2005, 10, 368–375. [Google Scholar] [CrossRef]
- Hunt, L.; Otterhag, L.; Lee, J.C.; Lasheen, T.; Hunt, J.; Seki, M.; Shinozaki, K.; Sommarin, M.; Gilmour, D.J.; Pical, C.; et al. Gene-specific expression and calcium activation of Arabidopsis thaliana phospholipase C isoforms. New Phytol. 2004, 162, 643–654. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.; Zhu, J.K. Abiotic stress signal transduction in plants: Molecular and genetic perspectives. Physiol. Plant. 2001, 112, 152–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Tan, B.; Zhou, X.; Xue, J.; Zhang, X.; Wang, P.; Shao, C.; Li, Y.; Li, C.; Xia, H.; et al. Interferon-inducible transmembrane protein 3 genetic variant rs12252 and influenza susceptibility and severity: A meta-analysis. PLoS ONE 2015, 10, e0124985. [Google Scholar] [CrossRef] [PubMed]
- Østergaard, L.; Yanofsky, M.F. Establishing gene function by mutagenesis in Arabidopsis thaliana. Plant J. 2004, 39, 682–696. [Google Scholar] [CrossRef]
- Li, Z.; Zou, Y.; Li, Y.; Sun, C.; Liu, Y.; Cui, L.; Liu, J.; Yang, Z.; Chen, Y. Exploring the salt-and drought-tolerant genes of alfalfa through expression library screening strategy. Grass Res. 2021, 1, 11. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Classifications | Transmembrane Protein | Roles | Species |
|---|---|---|---|
| Leaf development | ZmMATE884 | Participating in the growth and development regulation of the branches and leaves. | Maize [80] |
| PgTIP1 | Promoting the leaf size of panax ginseng and accelerating the growth of rape seed and Arabidopsis thaliana. | Ginseng [83], Arabidopsis, and rape [84] | |
| Root development | ZmMATE884 | Participating in the growth and development regulation of the root system of maize. | maize [78] |
| ALF5 | Participating in lateral root development. | Arabidopsis [85] | |
| PgTIP1 | Promoting root growth. | Ginseng [83], Arabidopsis, and rape [87] | |
| Reproductive growth | ALMT | Regulating the growth of pollen tubes. | Arabidopsis [89] |
| FAX1 | Enhacing the male fertility, lipid synthesis, and horn synthesis of leaves and flowers. | Arabidopsis [86] | |
| ZmMATE884 | Participating in the growth and development regulation of the embryo after pollination. | Maize [80] | |
| BIGEl | Regulating organogenesis and organ size. | Maize [91] | |
| PgTIP1 | Promoting photosynthesis and growth of plants. | Ginseng [87] | |
| Phytohormone regulation | PIN and AUX family | Responsible for polar transport of auxin. | Arabidopsis thaliana [93,80] |
| ZmMATE884 | Regulating the development of plant hypocotyls by affecting synthesis, transportation, or response of plant hormones. | Maize [87] | |
| AtWBC11 | Abscisic acid was involved in its expression to regulate hormones and light signals. | Arabidopsis [92] | |
| NRAMP | Participating in plant hormone regulation. | Arabidopsis, paddy, soybean, and tobacco [99] | |
| Response to salt stress | MaTET | Participating in SOS signal transduction pathway under salt stress. | Arabidopsis and banana [103] |
| GhSARP1 | Negatively regulating ubiquitous-mediated salt stress response. | Arabidopsis and cotton [104] | |
| AtSTRF1 | Promoting the tolerance to salt ion and osmotic stress. | Arabidopsis [105] | |
| PgTIP1 | Improving the salt tolerance. | Ginseng and Arabidopsis [106] | |
| Response to drought stress | MaTET | Participating in molecular responses. | Banana and Arabidopsis [103] |
| AtCHYR1 | Promoting ABA-induced stomatal closure and ROS production. | Arabidopsis [108] | |
| PeCHYR1 | Inducing stomatal closure to improve drought resistance. | Poplar and Arabidopsis [109] | |
| NtTIP1 | Enhancing channel activity to promote water absorption and maintain osmotic balance. | Tobacco [111] | |
| Response to temperature stress | COR413-PM1 | Regulating tolerance of freezing stress. | Arabidopsis [112] |
| RcPIP1;3, RcPIP2;1, RcPIP2;2 | A functional antagonistic relationship exists among the three genes to regulate cold adaptation. | Castor-oil plant [114] | |
| BnTR1 | Regulating Ca2+ dynamics to induce heat shock protein expressions and promote the tolerance of heat stress. | Rape, rice [117] | |
| Responsive to biotic stresses | BDA1 | Activating SA synthesis and WRKY70-dependent defense gene expression to enhance plant defense responses. | Arabidopsis [118] |
| AtSYP121, AtSYP122 | Resistant to powdery mildew. | Arabidopsis [119] | |
| OsPENl | Disease resistance to a variety of pathogens. | Rice [120] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Wang, B.; Yuan, F. The Role of Transmembrane Proteins in Plant Growth, Development, and Stress Responses. Int. J. Mol. Sci. 2022, 23, 13627. https://doi.org/10.3390/ijms232113627
Zhou Y, Wang B, Yuan F. The Role of Transmembrane Proteins in Plant Growth, Development, and Stress Responses. International Journal of Molecular Sciences. 2022; 23(21):13627. https://doi.org/10.3390/ijms232113627
Chicago/Turabian StyleZhou, Yingli, Baoshan Wang, and Fang Yuan. 2022. "The Role of Transmembrane Proteins in Plant Growth, Development, and Stress Responses" International Journal of Molecular Sciences 23, no. 21: 13627. https://doi.org/10.3390/ijms232113627