

Analysis of the Site-Specific Myoglobin Modifications in the Melibiose-Derived Novel Advanced Glycation End-Product
 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Identification of AGEs Generated on the Model Protein MB
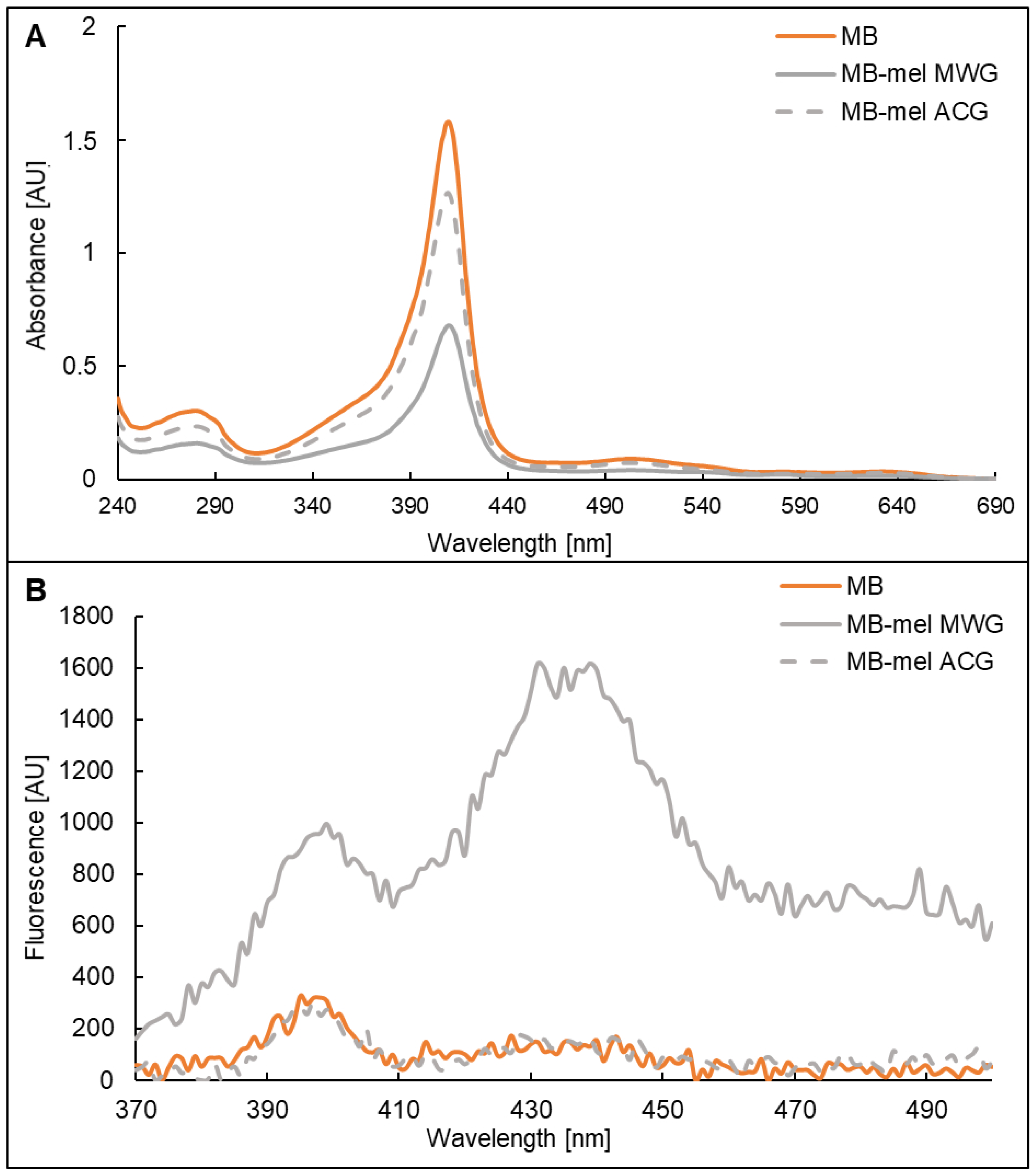
2.2. Absorption and Fluorescence Properties of MB-mel Products
2.3. Fourier-Transform Infrared Spectroscopy (FTIR) of the Glycated Myoglobin
2.4. Analysis of the Site-Specific Modifications Present on MB Glycated with Mel
2.5. MAGE Adducts Consisting of an Intact Melibiose
3. Discussion
4. Materials and Methods
4.1. Synthesis of Protein AGEs
4.2. SDS-PAGE and Western Blotting
4.3. Absorption Spectra
4.4. Fluorescence Spectra
4.5. Fourier-Transform Infrared Spectroscopy (FTIR)
4.6. Mass Spectrometry Analysis
4.7. Proteomic Data Analysis
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Cho, S.-J.; Roman, G.; Yeboah, F.; Konishi, Y. The Road to Advanced Glycation End Products: A Mechanistic Perspective. Curr. Med. Chem. 2007, 14, 1653–1671. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.-A.; Wu, C.-H.; Yen, G.-C. Perspective of Advanced Glycation End Products on Human Health. J. Agric. Food Chem. 2018, 66, 2065–2070. [Google Scholar] [CrossRef] [PubMed]
- Fournet, M.; Bonté, F.; Desmoulière, A. Glycation damage: A possible hub for major pathophysiological disorders and aging. Aging Dis. 2018, 9, 880–900. [Google Scholar] [CrossRef] [Green Version]
- Krhač, M.; Lovrenčić, M.V. Update on biomarkers of glycemic control. World J. Diabetes 2019, 10, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Vetter, S.W. Glycated Serum Albumin and AGE Receptors, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2015; Volume 72, ISBN 9780128033142. [Google Scholar]
- Khan, M.S.; Rabbani, N.; Tabrez, S.; Ul Islam, B.; Malik, A.; Ahmed, A.; Alsenaidy, M.A.; Alsenaidy, A.M. Glycation Induced Generation of Amyloid Fibril Structures by Glucose Metabolites. Protein Pept. Lett. 2016, 23, 892–897. [Google Scholar] [CrossRef]
- Chen, Y.; Yu, L.; Wang, Y.; Wei, Y.; Xu, Y.; He, T.; He, R. D-Ribose contributes to the glycation of serum protein. Biochim. Biophys. Acta-Mol. Basis Dis. 2019, 1865, 2285–2292. [Google Scholar] [CrossRef]
- Amani, S.; Fatima, S. Glycation with Fructose: The Bitter Side of Nature’s Own Sweetener. Curr. Diabetes Rev. 2020, 16, 962–970. [Google Scholar] [CrossRef]
- Lin, J.-A.A.; Wu, C.-H.H.; Lu, C.-C.C.; Hsia, S.-M.M.; Yen, G.-C.C. Glycative stress from advanced glycation end products (AGEs) and dicarbonyls: An emerging biological factor in cancer onset and progression. Mol. Nutr. Food Res. 2016, 60, 1850–1864. [Google Scholar] [CrossRef]
- Ahmad, S.; Khan, M.S.; Akhter, F.; Khan, M.S.; Khan, A.; Ashraf, J.M.; Pandey, R.P.; Shahab, U. Glycoxidation of biological macromolecules: A critical approach to halt the menace of glycation. Glycobiology 2014, 24, 979–990. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, S.; Maity, S.; Chakraborti, A.S. Methylglyoxal-induced modification causes aggregation of myoglobin. Spectrochim. Acta-Mol. Biomol. Spectrosc. 2016, 155, 1–10. [Google Scholar] [CrossRef]
- Hsu, Y.H.; Chen, Y.W.; Wu, M.H.; Tu, L.H. Protein Glycation by Glyoxal Promotes Amyloid Formation by Islet Amyloid Polypeptide. Biophys. J. 2019, 116, 2304–2313. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.U.; Thorpe, S.R.; Baynes, J.W. Identification of N(ε)-carboxymethyllysine as a degradation product of fructoselysine in glycated protein. J. Biol. Chem. 1986, 261, 4889–4894. [Google Scholar] [CrossRef]
- Ikeda, K.; Higashi, T.; Sano, H.; Jinnouchi, Y.; Yoshida, M.; Araki, T.; Ueda, S.; Horiuchi, S. Ne-(carboxymethyl)lysine protein adduct is a major immunological epitope in proteins modified with advanced glycation end products of the maillard reaction. Biochemistry 1996, 35, 8075–8083. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.U.; Brinkmann Frye, E.; Degenhardt, T.P.; Thorpe, S.R.; Baynes, J.W. Nε-(Carboxyethyl)lysine, a product of the chemical modification of proteins by methylglyoxal, increases with age in human lens proteins. Biochem. J. 1997, 324, 565–570. [Google Scholar] [CrossRef]
- Portero-Otin, M.; Nagaraj, R.H.; Monnier, V.M. Chromatographic evidence for pyrraline formation during protein glycation in vitro and in vivo. Biochim. Biophys. Acta (BBA) Protein Struct. Mol. Enzymol. 1995, 1247, 74–80. [Google Scholar] [CrossRef]
- Wa, C.; Cerny, R.L.; Clarke, W.A.; Hage, D.S. Characterization of glycation adducts on human serum albumin by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. Clin. Chim. Acta 2007, 385, 48–60. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.-S.; Park, S.; Kim, J. The role of glycation in the pathogenesis of aging and its prevention through herbal products and physical exercise. J. Exerc. Nutr. Biochem. 2017, 21, 55–61. [Google Scholar] [CrossRef]
- Henning, C.; Glomb, M.A. Pathways of the Maillard reaction under physiological conditions. Glycoconj. J. 2016, 33, 499–512. [Google Scholar] [CrossRef]
- Cotham, W.E.; Metz, T.O.; Ferguson, P.L.; Brock, J.W.C.C.; Hinton, D.J.S.S.; Thorpe, S.R.; Baynes, J.W.; Ames, J.M. Proteomic analysis of arginine adducts on glyoxal-modified ribonuclease. Mol. Cell. Proteom. 2004, 3, 1145–1153. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, N.; Thornalley, P.J.; Dawczynski, J.; Franke, S.; Strobel, J.; Stein, G.; Haik, G.M. Methylglyoxal-Derived Hydroimidazolone Advanced Glycation End-Products of Human Lens Proteins. Investig. Ophthalmol. Vis. Sci. 2003, 44, 5287–5292. [Google Scholar] [CrossRef]
- Grandhee, S.K.; Monnier, V.M. Mechanism of formation of the maillard protein cross-link pentosidine: Glucose, fructose, and ascorbate as pentosidine precursors. J. Biol. Chem. 1991, 266, 11649–11653. [Google Scholar] [CrossRef]
- Staniszewska, M.; Bronowicka-Szydełko, A.; Gostomska-Pampuch, K.; Szkudlarek, J.; Bartyś, A.; Bieg, T.; Gamian, E.; Kochman, A.; Picur, B.; Pietkiewicz, J.; et al. The melibiose-derived glycation product mimics a unique epitope present in human and animal tissues. Sci. Rep. 2021, 11, 2940. [Google Scholar] [CrossRef]
- Gostomska-Pampuch, K.; Gamian, A.; Rawicz-Pruszyński, K.; Gęca, K.; Tkaczuk-Włach, J.; Jonik, I.; Ożga, K.; Staniszewska, M. Proteins in human body fluids contain in vivo antigen analog of the melibiose-derived glycation product: MAGE. Sci. Rep. 2022, 12, 7520. [Google Scholar] [CrossRef] [PubMed]
- Czech, M.; Konopacka, M.; Rogoliński, J.; Maniakowski, Z.; Staniszewska, M.; Łaczmański, Ł.; Witkowska, D.; Gamian, A. The Genotoxic and Pro-Apoptotic Activities of Advanced Glycation End-Products (MAGE) Measured with Micronuclei Assay Are Inhibited by Their Low Molecular Mass Counterparts. Genes 2021, 12, 729. [Google Scholar] [CrossRef] [PubMed]
- Bronowicka-szydełko, A.; Krzystek-korpacka, M.; Gacka, M. Association of Novel Advanced Glycation End-Product (AGE10) with Complications of Diabetes as Measured by Enzyme-Linked Immunosorbent Assay. J. Clin. Med. 2021, 10, 4499. [Google Scholar] [CrossRef] [PubMed]
- Indyk, D.; Bronowicka-Szydełko, A.; Gamian, A.; Kuzan, A. Advanced glycation end products and their receptors in serum of patients with type 2 diabetes. Sci. Rep. 2021, 11, 13264. [Google Scholar] [CrossRef]
- O’Connell, K.J.; Motherway, M.O.C.; O’Callaghan, J.; Fitzgerald, G.F.; Paul Ross, R.; Ventura, M.; Stanton, C.; van Sinderen, D. Metabolism of four α-glycosidic linkage-containing oligosaccharides by Bifidobacterium breve UCC2003. Appl. Environ. Microbiol. 2013, 79, 6280–6292. [Google Scholar] [CrossRef] [Green Version]
- O’Callaghan, A.; van Sinderen, D. Bifidobacteria and their role as members of the human gut microbiota. Front. Microbiol. 2016, 7, 925. [Google Scholar] [CrossRef] [Green Version]
- Yoon, M.Y.; Hwang, H.J. Reduction of soybean oligosaccharides and properties of α-d-galactosidase from Lactobacillus curvatus R08 and Leuconostoc mesenteriodes JK55. Food Microbiol. 2008, 25, 815–823. [Google Scholar] [CrossRef]
- Baú, T.R.; Garcia, S.; Ida, E.I. Changes in soymilk during fermentation with kefir culture: Oligosaccharides hydrolysis and isoflavone aglycone production. Int. J. Food Sci. Nutr. 2015, 66, 845–850. [Google Scholar] [CrossRef]
- Schievano, E.; Tonoli, M.; Rastrelli, F. NMR Quantification of Carbohydrates in Complex Mixtures. A Challenge on Honey. Anal. Chem. 2017, 89, 13405–13414. [Google Scholar] [CrossRef] [PubMed]
- Sandek, A.; Bjarnason, I.; Volk, H.D.; Crane, R.; Meddings, J.B.; Niebauer, J.; Kalra, P.R.; Buhner, S.; Herrmann, R.; Springer, J.; et al. Studies on bacterial endotoxin and intestinal absorption function in patients with chronic heart failure. Int. J. Cardiol. 2012, 157, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Staniszewska, M.; Jarosz, S.; Jon, M.; Gamian, A. Advanced glycation end-products prepared in solution under high pressure contain epitopes distinct from those formed in the dry reaction at high temperature. Arch. Immunol. Ther. Exp. 2005, 53, 71–78. [Google Scholar]
- Anderson, A.B.; Robertson, C.R. Absorption spectra indicate conformational alteration of myoglobin adsorbed on polydimethylsiloxane. Biophys. J. 1995, 68, 2091–2097. [Google Scholar] [CrossRef] [Green Version]
- Karumanchi, D.K.; Karunaratne, N.; Lurio, L.; Dillon, J.P.; Gaillard, E.R. Non-enzymatic glycation of α-crystallin as an in vitro model for aging, diabetes and degenerative diseases. Amino Acids 2015, 47, 2601–2608. [Google Scholar] [CrossRef]
- Jahouh, F.; Hou, S.J.; Kováč, P.; Banoub, J.H. Determination of glycation sites by tandem mass spectrometry in a synthetic lactose-bovine serum albumin conjugate, a vaccine model prepared by dialkyl squarate chemistry. Rapid Commun. Mass Spectrom. 2012, 26, 749–758. [Google Scholar] [CrossRef] [Green Version]
- Pietkiewicz, J.; Gamian, A.; Staniszewska, M.; Danielewicz, R. Inhibition of human muscle-specific enolase by methylglyoxal and irreversible formation of advanced glycation end products. J. Enzyme Inhib. Med. Chem. 2009, 24, 356–364. [Google Scholar] [CrossRef]
- Banerjee, S.; Chakraborti, A.S. In vitro study on structural alteration of myoglobin by methylglyoxal. Protein J. 2013, 32, 216–222. [Google Scholar] [CrossRef]
- Liu, J.J.; You, Y.; Gao, S.Q.; Tang, S.; Chen, L.; Wen, G.B.; Lin, Y.W. Identification of the Protein Glycation Sites in Human Myoglobin as Rapidly Induced by d-Ribose. Molecules 2021, 26, 5829. [Google Scholar] [CrossRef]
- Castellani, R.; Smith, M.A.; Richey, P.L.; Perry, G. Glycoxidation and oxidative stress in Parkinson disease and diffuse Lewy body disease. Brain Res. 1996, 737, 195–200. [Google Scholar] [CrossRef]
- Oya, T.; Hattori, N.; Mizuno, Y.; Miyata, S.; Maeda, S.; Osawa, T.; Uchida, K. Methylglyoxal modification of protein. Chemical and immunochemical characterization of methylglyoxal-arginine adducts. J. Biol. Chem. 1999, 274, 18492–18502. [Google Scholar] [CrossRef] [PubMed]
- Nagai, R.; Hayashi, C.M.; Xia, L.; Takeya, M.; Horiuchi, S. Identification in human atherosclerotic lesions of GA-pyridine, a novel structure derived from glycolaldehyde-modified proteins. J. Biol. Chem. 2002, 277, 48905–48912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alizadeh-Rahrovi, J.; Shayesteh, A.; Ebrahim-Habibi, A. Structural stability of myoglobin and glycomyoglobin: A comparative molecular dynamics simulation study. J. Biol. Phys. 2015, 41, 349–366. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, S. Formation of Pentosidine Cross-Linking in Myoglobin by Glyoxal: Detection of Fluorescent Advanced Glycation End Product. J. Fluoresc. 2017, 27, 1213–1219. [Google Scholar] [CrossRef] [PubMed]
- Ghelani, H.; Razmovski-Naumovski, V.; Pragada, R.R.; Nammi, S. Attenuation of glucose-induced myoglobin glycation and the formation of advanced glycation end products (AGEs) by (R)-α-lipoic acid in vitro. Biomolecules 2018, 8, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makita, Z.; Vlassara, H.; Cerami, A.; Bucala, R. Immunochemical detection of advanced glycosylation end products in vivo. J. Biol. Chem. 1992, 267, 5133–5138. [Google Scholar] [CrossRef]
- Tofani, L.; Feis, A.; Snoke, R.E.; Berti, D.; Baglioni, P.; Smulevich, G. Spectroscopic and interfacial properties of myoglobin/surfactant complexes. Biophys. J. 2004, 87, 1186–1195. [Google Scholar] [CrossRef] [Green Version]
- Raupbach, J.; Ott, C.; Koenig, J.; Grune, T. Proteasomal degradation of glycated proteins depends on substrate unfolding: Preferred degradation of moderately modified myoglobin. Free Radic. Biol. Med. 2020, 152, 516–524. [Google Scholar] [CrossRef]
- Banerjee, S.; Chakraborti, A.S. Structural alterations of hemoglobin and myoglobin by glyoxal: A comparative study. Int. J. Biol. Macromol. 2014, 66, 311–318. [Google Scholar] [CrossRef]
- Mallya, M.; Shenoy, R.; Kodyalamoole, G.; Biswas, M.; Karumathil, J.; Kamath, S. Absorption spectroscopy for the estimation of glycated hemoglobin (HbA1c) for the diagnosis and management of diabetes mellitus: A pilot study. Photomed. Laser Surg. 2013, 31, 219–224. [Google Scholar] [CrossRef]
- Chhabra, A.; Bhatia, A.; Ram, A.K.; Goel, S. Increased advanced glycation end product specific fluorescence in repeatedly heated used cooking oil. J. Food Sci. Technol. 2017, 54, 2602–2606. [Google Scholar] [CrossRef] [PubMed]
- GhoshMoulick, R.; Bhattacharya, J.; Roy, S.; Basak, S.; Dasgupta, A.K. Compensatory secondary structure alterations in protein glycation. Biochim. Biophys. Acta-Proteins Proteom. 2007, 1774, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Ball, P. Water as an Active Constituent in Cell Biology. Chem. Rev. 2008, 108, 74–108. [Google Scholar] [CrossRef] [PubMed]
- Merzel, F.; Avbelj, F. Why do water molecules around small hydrophobic solutes form stronger hydrogen bonds than in the bulk? Biochim. Biophys. Acta (BBA)-Gen. Subj. 2020, 1864, 129537. [Google Scholar] [CrossRef] [PubMed]
- Carulli, S.; Calvano, C.D.; Palmisano, F.; Pischetsrieder, M. MALDI-TOF MS characterization of glycation products of whey proteins in a glucose/galactose model system and lactose-free milk. J. Agric. Food Chem. 2011, 59, 1793–1803. [Google Scholar] [CrossRef]
- Montgomery, H.; Tanaka, K.; Belgacem, O. Glycation pattern of peptides condensed with maltose, lactose and glucose determined by ultraviolet matrix-assisted laser desorption/ionization tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2010, 24, 841–848. [Google Scholar] [CrossRef]
- Shinohara, Y.; Ozawa, Y.; Okita, A.; Kato, K.; Chiba, T.; Miura, N.; Furukawa, J.I. Analysis of the susceptibility of reducing disaccharides composed of D-glucose to glycation using the Maillard reaction and a novel sensitive method that measures the percentage of the open-ring form. Carbohydr. Res. 2020, 493, 108019. [Google Scholar] [CrossRef]
- Hull, G.L.J.; Woodside, J.V.; Ames, J.M.; Cuskelly, G.J. N*-(carboxymethyl)lysine content of foods commonly consumed in a Western style diet. Food Chem. 2012, 131, 170–174. [Google Scholar] [CrossRef]
- Perrone, A.; Giovino, A.; Benny, J.; Martinelli, F. Advanced Glycation End Products (AGEs): Biochemistry, Signaling, Analytical Methods, and Epigenetic Effects. Oxid. Med. Cell. Longev. 2020, 2020, 3818196. [Google Scholar] [CrossRef] [Green Version]
- Schalkwijk, C.G.; Stehouwer, C.D.A. Methylglyoxal, a Highly Reactive Dicarbonyl Compound, in Diabetes, Its Vascular Complications, and Other Age-Related Diseases. Physiol. Rev. 2020, 100, 407–461. [Google Scholar] [CrossRef]
- Chen, C.; Zhang, J.-Q.; Li, L.; Guo, M.; He, Y.; Dong, Y.; Meng, H.; Yi, F. Advanced Glycation End Products in the Skin: Molecular Mechanisms, Methods of Measurement, and Inhibitory Pathways. Front. Med. 2022, 9, 837222. [Google Scholar] [CrossRef] [PubMed]
- Allaman, I.; Bélanger, M.; Magistretti, P.J. Methylglyoxal, the dark side of glycolysis. Front. Neurosci. 2015, 9, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klaus, A.; Rau, R.; Glomb, M.A. Modification and Cross-Linking of Proteins by Glycolaldehyde and Glyoxal: A Model System. J. Agric. Food Chem. 2018, 66, 10835–10843. [Google Scholar] [CrossRef] [PubMed]
- Wiśniewski, J.R. Filter Aided Sample Preparation—A tutorial. Anal. Chim. Acta 2019, 1090, 23–30. [Google Scholar] [CrossRef]
- Wiśniewski, J.R.; Gaugaz, F.Z. Fast and Sensitive Total Protein and Peptide Assays for Proteomic Analysis. Anal. Chem. 2015, 87, 4110–4116. [Google Scholar] [CrossRef]
- Rappsilber, J.; Ishihama, Y.; Mann, M. Stop and Go Extraction tips for matrix-assisted laser desorption/ionization, nanoelectrospray, and LC/MS sample pretreatment in proteomics. Anal. Chem. 2003, 75, 663–670. [Google Scholar] [CrossRef]
- Wiśniewski, J.R. Quantitative Evaluation of Filter Aided Sample Preparation (FASP) and Multienzyme Digestion FASP Protocols. Anal. Chem. 2016, 88, 5438–5443. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Modification | Modified Residue | Detected m/z | ∆m [Da] | Score | Peptide Sequence |
|---|---|---|---|---|---|
| CML | K78 | 412.225 | +58.005 | 6.89 × 1014 | _K(CML)KGHHEAELK_ |
| CML | K87 | 412.224 | +58.005 | 3.76 × 1014 | _KKGHHEAELK(CML)_ |
| CML | K56 | 470.553 | +58.005 | 5.69 × 1014 | _TEAEMK(CML)ASEDLK_ |
| CEL | K145 | 716.894 | +72.021 | 6.9 × 1014 | _ALELFRNDIAAK(CEL)_ |
| CML | K63 | 522.318 | +58.005 | 1.97 × 1014 | _K(CML)HGTVVLTALGGILK_ |
| Hex, CML 2× CEL | H64, K63, K62, K77 | 1257.686 | +364.100 | 5.35 × 1014 | _ASEDLK(CEL)K(CML)H(Hex)GTVVLTALGGILK(CEL)_ |
| MGO | K56 | 469.222 | +54.010 | 4.69 × 1014 | _TEAEMK(MGO)ASEDLK_ |
| Type of Modification | Modified Residue | Detected m/z | ∆m [Da] | Score | Peptide Sequence |
|---|---|---|---|---|---|
| 2× Hex | K63, K77 | 611.018 | +324.106 | 7.45 × 1014 | _K(Hex)HGTVVLTALGGILK(Hex)_ |
| 3× MOLD | K63, K77, K78 | 594.690 | +147.024 | 3.97 × 1014 | _K(MOLD)HGTVVLTALGGILK(MOLD)K(MOLD)_ |
| CML | K87 | 412.224 | +58.005 | 1.75 × 1014 | _KKGHHEAELK(CML)_ |
| CML | K96 | 425.898 | +58.005 | 3.81 × 1014 | _PLAQSHATK(CML)HK_ |
| CML | K56 | 470.553 | +58.005 | 8.26 × 1014 | _TEAEMK(CML)ASEDLK_ |
| CML | K77 | 782.974 | +58.005 | 9.56 × 1014 | _HGTVVLTALGGILK(CML)K_ |
| CML | K63 | 522.319 | +58.005 | 7.59 × 1014 | _K(CML)HGTVVLTALGGILK_ |
| GAP | K78 | 539.325 | +109.029 | 7.48 × 1014 | _HGTVVLTALGGILKK(GAP)_ |
| GOLD, DOGDIC | K78, K79 | 668.834 | +160.016 | 5.56 × 1014 | _K(GOLD)K(DOGDIC)GHHEAELK_ |
| GOLD, DOGDIC | K87, K78 | 668.834 | +160.016 | 4.61 × 1014 | _K(DOGDIC)KGHHEAELK(GOLD)_ |
| Hex | K62 | 476.747 | +162.053 | 5.48 × 1014 | _ASEDLK(Hex)K_ |
| Hex | H93 | 611.018 | +162.053 | 4.7 × 1014 | _PLAQSH(Hex)ATKHKIPIK_ |
| Hex | K98 | 611.017 | +162.053 | 4.31 × 1014 | _PLAQSHATKHK(Hex)IPIK_ |
| Hex | K96 | 611.018 | +162.053 | 2.39 × 1014 | _PLAQSHATK(Hex)HKIPIK_ |
| mel | R139 | 659.345 | +324.106 | 4.39 × 1014 | _ALELFR(mel)NDIAAKYK_ |
| mel | K62 | 557.771 | +324.106 | 3.57 × 1013 | _ASEDLK(mel)K_ |
| mel | K63 | 611.018 | +324.106 | 4.88 × 1014 | _K(mel)HGTVVLTALGGILK_ |
| mel | H64 | 611.018 | +324.106 | 6.83 × 1014 | _H(mel)GTVVLTALGGILKK_ |
| mel | K77 | 611.018 | +324.106 | 4.32 × 1014 | _KHGTVVLTALGGILK(mel)_ |
| mel | K77 | 611.019 | +324.106 | 7.53 × 1014 | _HGTVVLTALGGILK(mel)K_ |
| mel, 2× CML | H97, K96, K98 | 553.268 | +440.116 | 7.05 × 1014 | _PLAQSHATK(CML)H(mel)K(CML)_ |
| MG-H1 | R139 | 569.313 | +54.010 | 4.32 × 1014 | _ALELFR(MG-H1)NDIAAKYK_ |
| MG-H1 | R139 | 472.261 | +54.010 | 8.01 × 1014 | _ALELFR(MG-H1)NDIAAK_ |
| Pyrraline | K96 | 593.008 | +108.021 | 1.18 × 1014 | _PLAQSHATK(Pyrraline)HKIPIK_ |
| 4× GA | H97, K98, H113, H116 | 1003.870 | +408.128 | 2.72 × 1014 | _H(GA)K(GA)IPIKYLEFISDAIIH(GA)VLH(GA)SK_ |
| GLX | K96 | 496.762 | +39.995 | 1.18 × 1014 | _PLAQSHATK(GLX)_ |
| GLX | H119 | 771.837 | +39.995 | 3.32 × 1014 | _H(GLX)PGDFGADAQGAMTK_ |
| 2× GLX | H93, K96 | 516.761 | +79.99 | 3.93 × 1014 | _PLAQSH(GLX)ATK(GLX)_ |
| MGO | K56 | 469.224 | +54.010 | 4.34 × 1014 | _TEAEMK(MGO)ASEDLK_ |
| MGO | K147 | 569.313 | +54.010 | 3.15 × 1014 | _ALELFRNDIAAKYK(MGO)_ |
| 2× MGO | K96, K102 | 593.008 | +108.020 | 1.18 × 1014 | _PLAQSHATK(MGO)HKIPIK(MGO)_ |
| 5× MGO | H93, K96, H97, K98, K102 | 647.021 | +270.050 | 1.15 × 1014 | _PLAQSH(MGO)ATK(MGO)H(MGO)K(MGO)IPIK(MGO)_ |
| 3× MGO | H93, K98, K102 | 611.010 | +162.030 | 3.62 × 1014 | _PLAQSH(MGO)ATKHK(MGO)IPIK(MGO)_ |
| MGO | K145 | 472.261 | +54.010 | 5.68 × 1014 | _ALELFRNDIAAK(MGO)_ |
| Type of Modification | Modified Residue | Detected m/z | ∆m [Da] | Score | Peptide Sequence |
|---|---|---|---|---|---|
| 2× Hex | K56, K62 | 559.253 | +324.106 | 8.10428 × 1014 | _TEAEMK(Hex)ASEDLK(Hex)_ |
| 2× Hex, 2× CML | H64, K77, K78, K79 | 692.390 | +324.106 | 7.97853 × 1014 | _H(Hex)GTVVLTALGGILK(Hex)K(CML)K(CML)_ |
| 3× mel, CML, CEL | H81, H82, K87, K78, K79 | 760.337 | +1102.344 | 6.10181 × 1014 | _K(CML)K(CEL)GH(mel)H(mel)EAELK(mel)_ |
| 3× MOLD | K62, K63, K77 | 766.430 | +147.024 | 7.30365 × 1014 | _ASEDLK(MOLD)K(MOLD)HGTVVLTALGGILK(MOLD)_ |
| CEL | K145 | 478.264 | +72.021 | 8.29719 × 1014 | _ALELFRNDIAAK(CEL)_ |
| CEL | K77 | 725.934 | +72.021 | 3.53221 × 1014 | _HGTVVLTALGGILK(CEL)_ |
| CML | K77 | 782.973 | +58.005 | 3.3215 × 1014 | _HGTVVLTALGGILK(CML)K_ |
| CML | K145 | 570.644 | +58.005 | 6.11103 × 1014 | _ALELFRNDIAAK(CML)YK_ |
| GOLD | K133 | 769.332 | +34.992 | 4.66304 × 1014 | _HPGDFGADAQGAMTK(GOLD)_ |
| GOLD, DOGDIC | K87, K78 | 668.834 | +160.016 | 8.9105 × 1014 | _K(DOGDIC)KGHHEAELK(GOLD)_ |
| Hex | K62 | 476.746 | +162.053 | 1.08165 × 1014 | _ASEDLK(Hex)K_ |
| mel, 2× CML | H64, K63, K77 | 692.390 | +440.116 | 6.8908 × 1014 | _K(CML)H(mel)GTVVLTALGGILK(CML)K_ |
| mel, CEL | K56, K62 | 583.261 | +396.127 | 5.48394 × 1014 | _TEAEMK(mel)ASEDLK(CEL)_ |
| MOLD | K47 | 556.313 | +49.008 | 4.13698 × 1014 | _FDKFK(MOLD)HLK_ |
| Pentosidine | K62 | 737.084 | +58.992 | 5.06918 × 1014 | _ASEDLK(Pentosidine)KHGTVVLTALGGILK_ |
| 2× GA | K62, K63 | 332.171 | +204.064 | 2.58266 × 1014 | _ASEDLK(GA)K(GA)_ |
| 4× GA | K79, H81, H82, K87 | 486.233 | +408.128 | 9.06642 × 1014 | _K(GA)GH(GA)H(GA)EAELK(GA)_ |
| GLX | H64 | 709.921 | +39.995 | 3.29039 × 1014 | _H(GLX)GTVVLTALGGILK_ |
| GLX | K77 | 473.617 | +39.995 | 8.98953 × 1014 | _HGTVVLTALGGILK(GLX)_ |
| GLX | H113 | 642.341 | +39.995 | 8.29994 × 1014 | _YLEFISDAIIH(GLX)VLHSK_ |
| MGO | K56 | 469.223 | +54.010 | 3.19781 × 1014 | _TEAEMK(MGO)ASEDLK_ |
| Glycation Type | Residue (Amount within MB) | Residues with Intact Mel | % of Residue Type | % of Nucleophilic Residues (32 aa) | % of Total Residues (153 aa) |
|---|---|---|---|---|---|
| MWG | K (19) | 4 | 21.1 | 21.9 | 4.6 |
| H (11) | 2 | 18.2 | |||
| R (2) | 1 | 50.0 | |||
| ACG | K (19) | 2 | 10.5 | 15.6 | 3.3 |
| H (11) | 3 | 27.3 | |||
| R (2) | 0 | 0.0 |
| AGE Modification Type | Monoisotopic Mass [Da] [Da] [Da] |
|---|---|
| CML | 58.005 |
| CEL | 72.021 |
| Hex/FL | 162.053 |
| Pyrraline | 108.021 |
| Pentosidine | 58.992 |
| GAP | 109.029 |
| MG-H1 (MGO on R) | 54.010 |
| GOLD | 34.992 |
| MOLD | 49.008 |
| DOGDIC | 125.024 |
| mel | 324.106 |
| GA | 102.032 |
| GLX | 39.995 |
| MGO (on K, H) | 54.010 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gostomska-Pampuch, K.; Wiśniewski, J.R.; Sowiński, K.; Gruszecki, W.I.; Gamian, A.; Staniszewska, M. Analysis of the Site-Specific Myoglobin Modifications in the Melibiose-Derived Novel Advanced Glycation End-Product. Int. J. Mol. Sci. 2022, 23, 13036. https://doi.org/10.3390/ijms232113036
Gostomska-Pampuch K, Wiśniewski JR, Sowiński K, Gruszecki WI, Gamian A, Staniszewska M. Analysis of the Site-Specific Myoglobin Modifications in the Melibiose-Derived Novel Advanced Glycation End-Product. International Journal of Molecular Sciences. 2022; 23(21):13036. https://doi.org/10.3390/ijms232113036
Chicago/Turabian StyleGostomska-Pampuch, Kinga, Jacek R. Wiśniewski, Karol Sowiński, Wieslaw I. Gruszecki, Andrzej Gamian, and Magdalena Staniszewska. 2022. "Analysis of the Site-Specific Myoglobin Modifications in the Melibiose-Derived Novel Advanced Glycation End-Product" International Journal of Molecular Sciences 23, no. 21: 13036. https://doi.org/10.3390/ijms232113036