
Three Heat Shock Protein Genes and Antioxidant Enzymes Protect Pardosa pseudoannulata (Araneae: Lycosidae) from High Temperature Stress

Abstract
:1. Introduction
2. Results
2.1. Sequence Analysis and Phylogenetic Tree
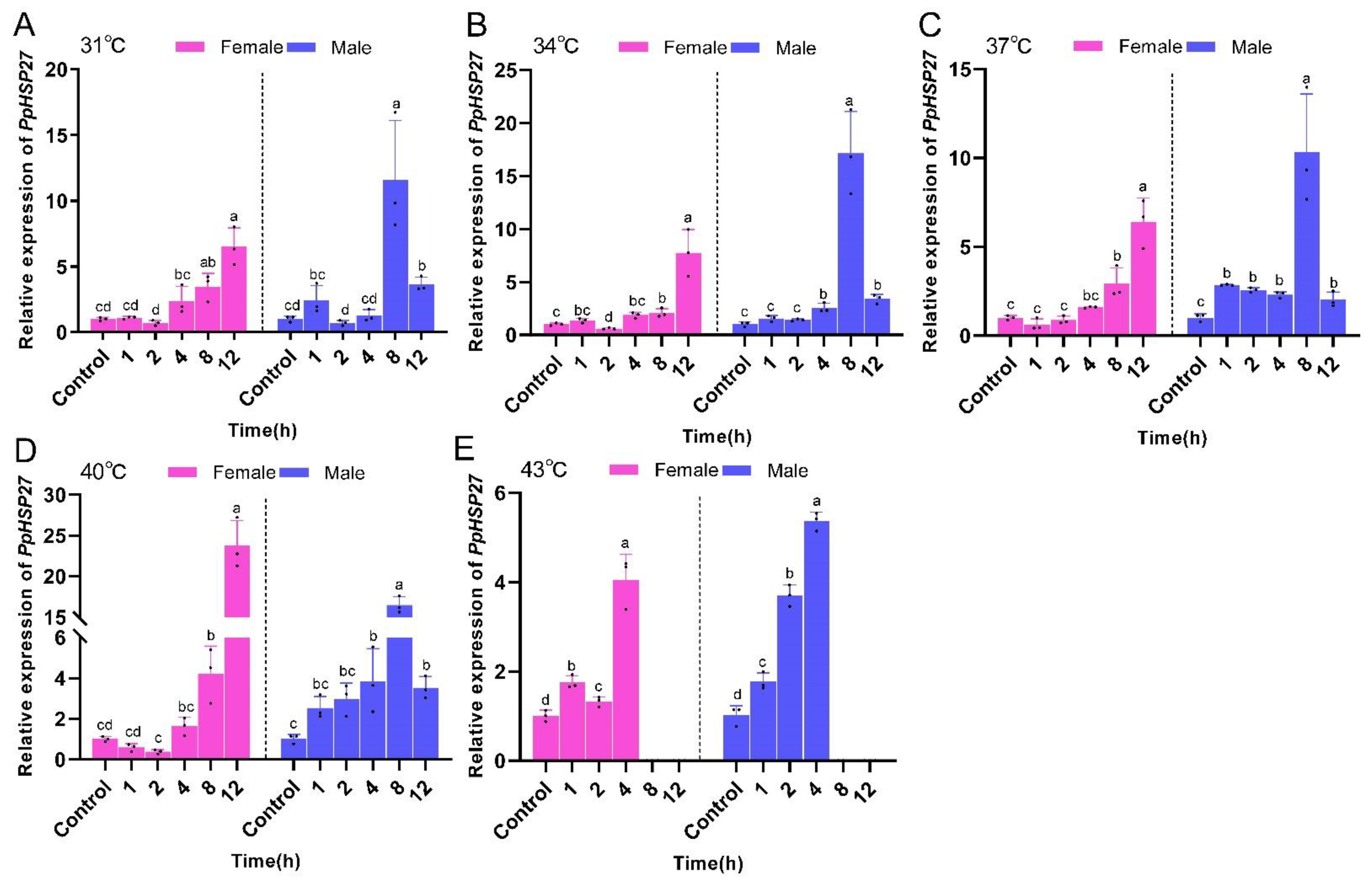
2.2. Effects of High Temperature Stress on PpHSP Genes Expression
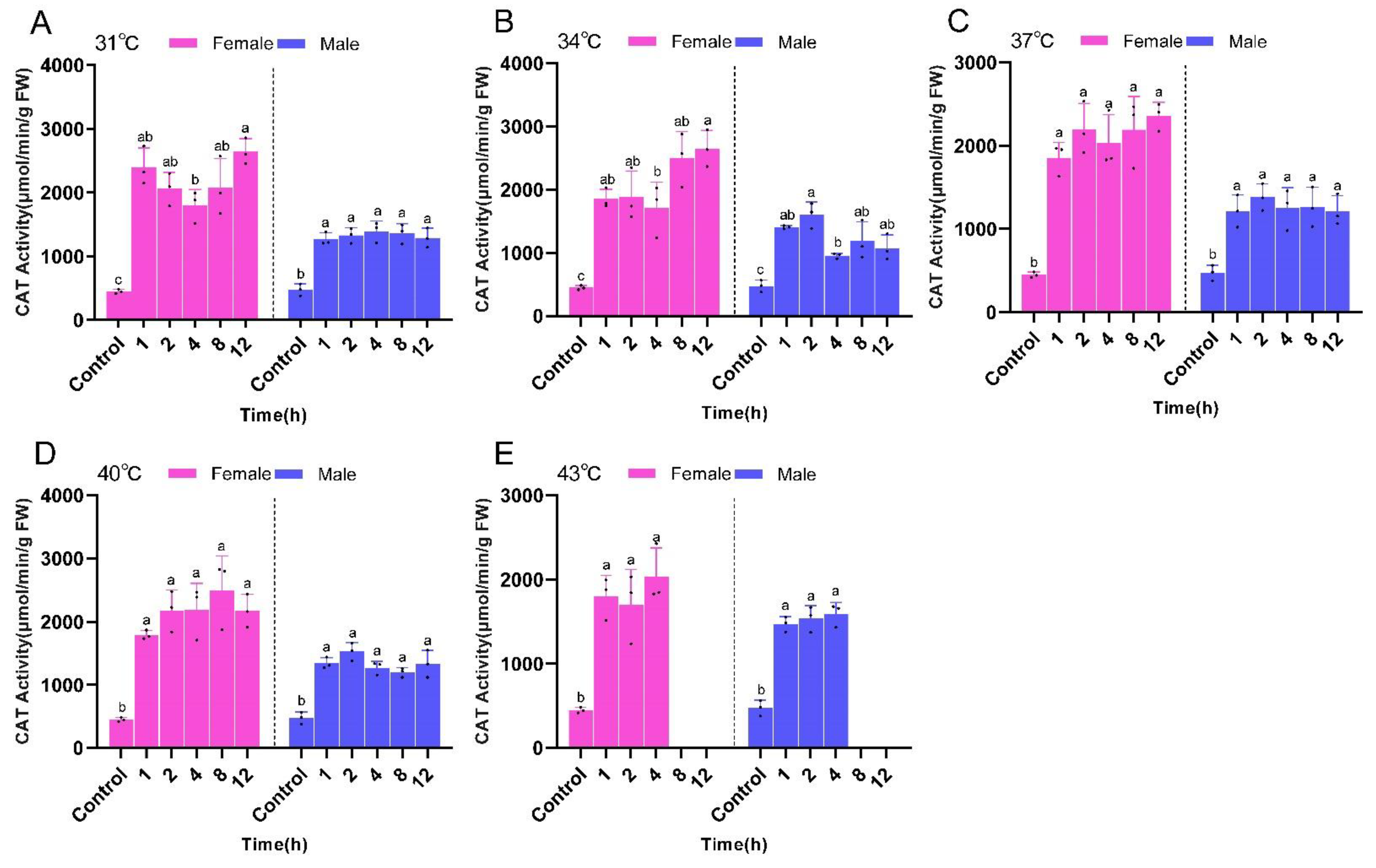
2.3. Changes in Antioxidant Enzyme Activity
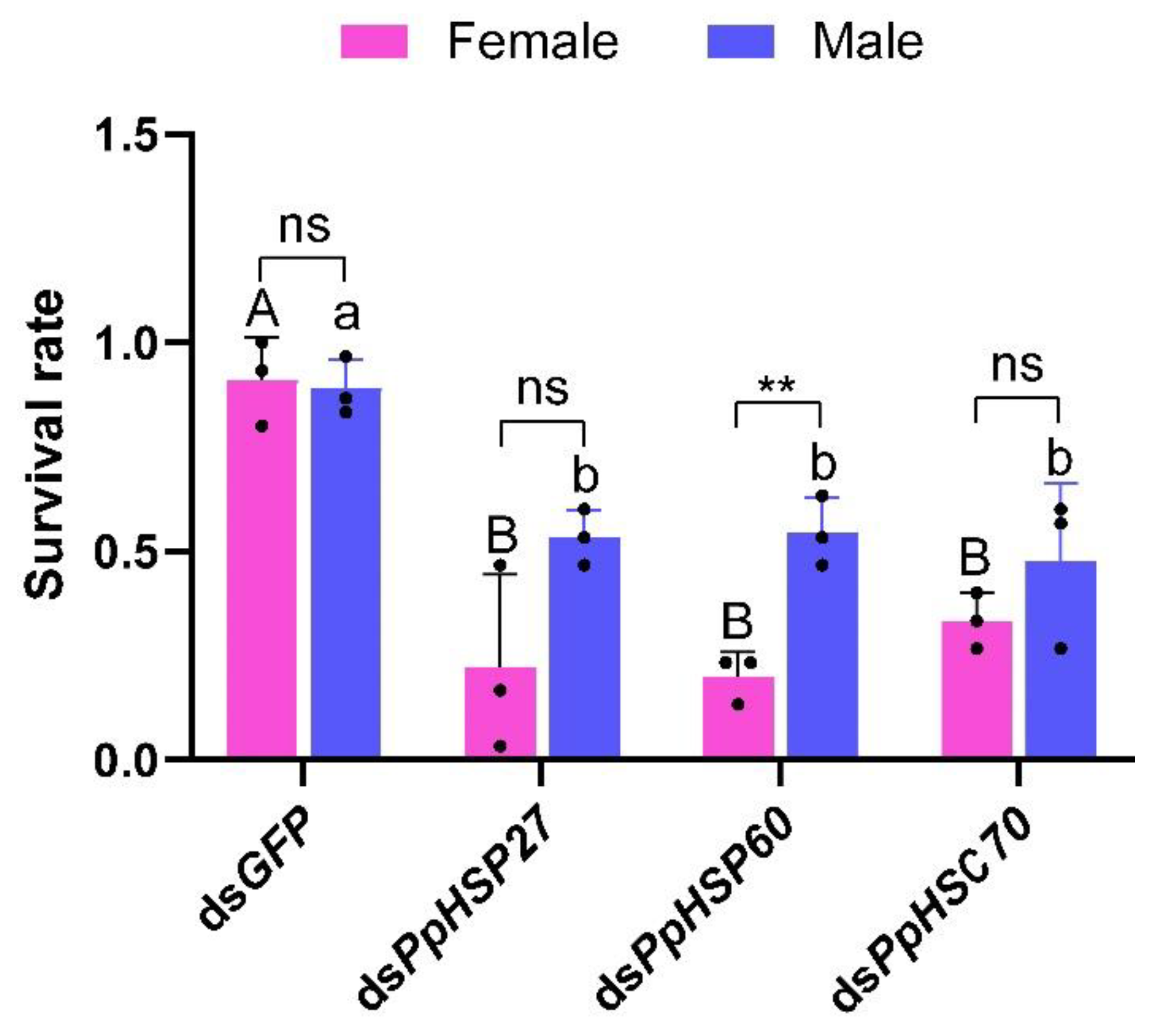
2.4. Effect of Silencing Three HSP Genes on High Temperature Tolerance in P. pseudoannulata
3. Discussion
4. Materials and Methods
4.1. Spider Feeding and Experimental Handling
4.2. RNA Extraction and cDNA Synthesis
4.3. Cloning the Full-Length cDNA of HSP Genes
4.4. Sequence Analysis
4.5. Quantitative Real-Time PCR
4.6. RNA Interference
4.7. Antioxidant Enzyme Activity Detection
4.8. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bale, J.S.; Masters, G.J.; Hodkinson, I.D.; Awmack, C.; Bezemer, T.M.; Brown, V.K.; Butterfield, J.; Buse, A.; Coulson, J.C.; Farrar, J.; et al. Herbivory in global climate change research: Direct effects of rising temperature on insect herbivores. Global Change Biol. 2002, 8, 1–16. [Google Scholar] [CrossRef]
- Horn, D.J. Temperature synergism in integrated pest management. In Temperature Sensitivity in Insects and Application in Integrated Pest Management; CRC Press: Boca Raton, FL, USA, 2019; pp. 125–140. [Google Scholar]
- Perkins-Kirkpatrick, S.E.; Lewis, S.C. Increasing trends in regional heatwaves. Nat. Commun. 2020, 11, 3357. [Google Scholar] [CrossRef]
- Wang, X.X.; Jiang, D.B.; Lang, X.M. Future extreme climate changes linked to global warming intensity. Sci. Bull. 2017, 62, 1673–1680. [Google Scholar] [CrossRef] [Green Version]
- Feder, M.E.; Hofmann, G.E. Heat-shock proteins, molecular chaperones, and the stress response: Evolutionary and ecological physiology. Annu. Rev. Physiol. 1999, 61, 243–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srensen, J.G.; Kristensen, T.N.; Loeschcke, V. The evolutionary and ecological role of heat shock proteins. Ecol. Lett. 2010, 6, 1025–1037. [Google Scholar] [CrossRef]
- Felton, G.W.; Summers, C.B. Antioxidant systems in insects. Arch. Insect Biochem. Physiol. 1995, 29, 187–197. [Google Scholar] [CrossRef]
- Perez, R.; Aron, S. Adaptations to thermal stress in social insects: Recent advances and future directions. Biol. Rev. 2020, 95, 1535–1553. [Google Scholar] [CrossRef]
- Gonzalez-Tokman, D.; Cordoba-Aguilar, A.; Dattilo, W.; Lira-Noriega, A.; Sanchez-Guillen, R.A.; Villalobos, F. Insect responses to heat: Physiological mechanisms, evolution and ecological implications in a warming world. Biol. Rev. 2020, 95, 802–821. [Google Scholar] [CrossRef]
- Kregel Kevin, C. Invited Review: Heat shock proteins: Modifying factors in physiological stress responses and acquired thermotolerance. J. Appl. Physiol. 2002, 192, 2177–2186. [Google Scholar] [CrossRef] [Green Version]
- King, A.M.; MacRae, T.H. Insect heat shock proteins during stress and diapause. In Annual Review of Entomology; Berenbaum, M.R., Ed.; Palo Alto: Santa Clara, CA, USA, 2015; Volume 60, pp. 59–75. [Google Scholar]
- Ganea, E. Chaperone-like activity of alpha-crystallin and other small heat shock proteins. Curr. Protein Pept. Sci. 2001, 2, 205–225. [Google Scholar] [CrossRef]
- Basha, E.; O’Neill, H.; Vierling, E. Small heat shock proteins and alpha-crystallins: Dynamic proteins with flexible functions. Trends Biochem. Sci. 2012, 37, 106–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fink, A.L. Chaperone-Mediated Protein Folding. Physiol. Rev. 1999, 79, 425–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morimoto, R.I. Cells in stress: Transcriptional activation of heat shock genes. Science 1993, 259, 1409–1410. [Google Scholar] [CrossRef] [PubMed]
- Bukau, B.; Horwich, A.L. The Hsp70 and Hsp60 chaperone machines. Cell 1998, 92, 351–366. [Google Scholar] [CrossRef] [Green Version]
- Mayer, M.P.; Bukau, B. Hsp70 chaperones: Cellular functions and molecular mechanism. Cell. Mol. Life Sci. 2005, 62, 670–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karunanithi, S.; Barclay, J.W.; Robertson, R.M.; Brown, I.R.; Atwood, H.L. Neuroprotection at Drosophila synapses conferred by prior heat shock. J. Neurosci. 1999, 19, 4360–4369. [Google Scholar] [CrossRef] [Green Version]
- Benoit, J.B.; Lopez-Martinez, G.; Patrick, K.R.; Phillips, Z.P.; Krause, T.B.; Denlinger, D.L. Drinking a hot blood meal elicits a protective heat shock response in mosquitoes. Proc. Natl. Acad. Sci. USA 2011, 108, 8026–8029. [Google Scholar] [CrossRef] [Green Version]
- Paim, R.M.; Araujo, R.N.; Leis, M.; Sant’Anna, M.R.; Gontijo, N.F.; Lazzari, C.R.; Pereira, M.H. Functional evaluation of Heat Shock Proteins 70 (HSP70/HSC70) on Rhodnius prolixus (Hemiptera, Reduviidae) physiological responses associated with feeding and starvation. Insect Biochem. Mol. Biol. 2016, 77, 10–20. [Google Scholar] [CrossRef]
- Lu, K.; Chen, X.; Liu, W.; Zhou, Q. Characterization of heat shock cognate protein 70 gene and its differential expression in response to thermal stress between two wing morphs of Nilaparvata lugens (Stål). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2016, 199, 47–53. [Google Scholar] [CrossRef]
- Krebs, R.A.; Feder, M.E.; Lee, J. Heritability of expression of the 70KD heat-shock protein in Drosophila melanogaster and its relevance to the evolution of thermotolerance. Evolution 1998, 52, 841–847. [Google Scholar] [CrossRef]
- Lü, Z.-C.; Wan, F.-H. Using double-stranded RNA to explore the role of heat shock protein genes in heat tolerance in Bemisia tabaci (Gennadius). J. Exp. Biol. 2011, 214, 764–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, D.; Shao, L.; Spitz, D.R. Reactive oxygen species in normal and tumor stem cells. Adv. Cancer Res. 2014, 122, 1–67. [Google Scholar] [PubMed] [Green Version]
- Jena, K.; Kar, P.K.; Kausar, Z.; Babu, C.S. Effects of temperature on modulation of oxidative stress and antioxidant defenses in testes of tropical tasar silkworm Antheraea mylitta. J. Therm. Biol. 2013, 38, 199–204. [Google Scholar] [CrossRef]
- Parsell, D.A.; Lindquist, S. The Function of Heat-Shock Proteins in Stress Tolerance: Degradation and Reactivation of Damaged Proteins. Annu. Rev. Genet. 1993, 27, 437–497. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ma, W.; Shen, J.; Long, D.; Jiang, Y. Tolerance and response of two honeybee species Apis cerana and Apis mellifera to high temperature and relative humidity. PLoS ONE 2019, 14, e0217921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, Z.Q.; Tu, Y.Q.; Guo, P.Y.; He, W.; Wei, D.D. Antioxidant Enzymes and Heat Shock Protein Genes from Liposcelis bostrychophila Are Involved in Stress Defense upon Heat Shock. Insects 2020, 11, 839. [Google Scholar] [CrossRef]
- Khurshid, A.; Inayat, R.; Tamkeen, A.; Haq, I.U.; Li, C.; Boamah, S.; Zhou, J.-J.; Liu, C. Antioxidant Enzymes and Heat-Shock Protein Genes of Green Peach Aphid (Myzus persicae) Under Short-Time Heat Stress. Front. Physiol. 2021, 12, 805509. [Google Scholar] [CrossRef]
- Yang, L.H.; Huang, H.; Wang, J.J. Antioxidant responses of citrus red mite, Panonychus citri (McGregor) (Acari: Tetranychidae), exposed to thermal stress. J. Insect Physiol. 2010, 56, 1871–1876. [Google Scholar] [CrossRef]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [Green Version]
- Dong, L.; He, Y.; Wang, Y.; Dong, Z. Research progress on application of superoxide dismutase (SOD). J. Agric. Sci. Technol. 2013, 15, 53–58. [Google Scholar]
- Mhamdi, A.; Queval, G.; Chaouch, S.; Vanderauwera, S.; Van Breusegem, F.; Noctor, G. Catalase function in plants: A focus on Arabidopsis mutants as stress-mimic models. J. Exp. Bot. 2010, 61, 4197–4220. [Google Scholar] [CrossRef] [PubMed]
- Vaish, S.; Gupta, D.; Mehrotra, R.; Mehrotra, S.; Basantani, M.K. Glutathione S-transferase: A versatile protein family. 3 Biotech 2020, 10, 321. [Google Scholar] [CrossRef] [PubMed]
- Sigsgaard, L. Early season natural control of the brown planthopper, Nilaparvata lugens: The contribution and interaction of two spider species and a predatory bug. Bull. Entomol. Res. 2007, 97, 533–544. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Peng, Y.; Tian, J.; Wang, J.; Wei, B.; Xie, C.; Wang, Z. Rice Field Spiders in China: A Review of the Literature. J. Econ. Entomol. 2018, 111, 53–64. [Google Scholar] [CrossRef] [PubMed]
- World Spider Catalog. In World Spider Catalog, Version 21.0; Natural History Museum Bern: London, UK, 2020.
- Wang, Z. Bionomics and behavior of the wolf spider, Pardosa pseudoannulata (Araneae: Lycosidae). Acta Entomol. Sin. 2007, 50, 927–932. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, L.; Shi, Y.; Tang, M.; Wang, Z. Effect of temperature on growth and development of Pardosa pseudoannulata. Chin. J. Biol. Control 2017, 33, 597–603. [Google Scholar] [CrossRef]
- Zhao, J.; Yuan, A.; Yu, K. Effect of temperature on the development and fertility of Pardosa pseudoannulata. J. Hubei Univ. 1989, 1, 1–9. [Google Scholar]
- Song, Y.; Wang, C.; Linderholm, H.W.; Fu, Y.; Cai, W.; Xu, J.; Zhuang, L.; Wu, M.; Shi, Y.; Wang, G. The negative impact of increasing temperatures on rice yields in southern China. Sci. Total Environ. 2022, 820, 153262. [Google Scholar] [CrossRef]
- Xiao, R.; Wang, L.; Cao, Y.; Zhang, G. Transcriptome response to temperature stress in the wolf spider Pardosa pseudoannulata (Araneae: Lycosidae). Ecol. Evol. 2016, 6, 3540–3554. [Google Scholar] [CrossRef] [Green Version]
- Zeng, L.; Cheng, Y.; Yan, Z.; Ma, J.; Ren, S.; Wei, L.; Xue, Z. Predation of Pardosa pseudoannulata and Pardosa procurva Against Plutella xylostella. Agric. Sci. Technol. 2016, 17, 2866. [Google Scholar] [CrossRef]
- Li, C.; Wang, Y.; Yao, G.; Li, G. Cloning and analysis of Pardosa pseudoannulata heat shock protein 20 gene. J. South. Agric. 2018, 49, 1525–1530. [Google Scholar]
- Sun, L.-Y.; Liu, J.; Li, Q.; Fu, D.; Zhu, J.-Y.; Guo, J.-J.; Xiao, R.; Jin, D.-C. Cloning and differential expression of three heat shock protein genes associated with thermal stress from the wolf spider Pardosa pseudoannulata (Araneae: Lycosidae). J. Asia Pac. Entomol. 2021, 24, 158–166. [Google Scholar] [CrossRef]
- Jin, J.; Zhao, M.; Wang, Y.; Zhou, Z.; Guo, J. Induced Thermotolerance and Expression of Three Key Hsp Genes (Hsp70, Hsp21, and sHsp21) and Their Roles in the High Temperature Tolerance of Agasicles hygrophila. Front. Physiol. 2020, 10, 1593. [Google Scholar] [CrossRef] [PubMed]
- Tian, C.; Li, Y.; Wu, Y.; Chu, W.; Liu, H. Sustaining induced heat shock protein 70 confers biological thermotolerance in a high-temperature adapted predatory mite Neoseiulus barkeri (Hughes). Pest Manag. Sci. 2021, 77, 939–948. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.-M.; Zhu, X.-L.; Lu, J.; Cai, W.-J.; Ye, Y.-P.; Lv, Y.-P. Effect of high temperature stress on heat shock protein expression and antioxidant enzyme activity of two morphs of the mud crab Scylla paramamosain. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2018, 223, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Flaherty, K.M.; DeLuca-Flaherty, C.; McKay, D.B. Three-dimensional structure of the ATPase fragment of a 70K heat-shock cognate protein. Nature 1990, 346, 623–628. [Google Scholar] [CrossRef] [PubMed]
- Snutch, T.P.; Heschl, M.F.; Baillie, D.L. The Caenorhabditis elegans hsp70 gene family: A molecular genetic characterization. Gene 1988, 64, 241–255. [Google Scholar] [CrossRef]
- Shim, J.-K.; Jung, D.-O.; Park, J.-W.; Kim, D.-W.; Ha, D.-M.; Lee, K.-Y. Molecular cloning of the heat-shock cognate 70 (Hsc70) gene from the two-spotted spider mite, Tetranychus urticae, and its expression in response to heat shock and starvation. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2006, 145, 288–295. [Google Scholar] [CrossRef]
- Cheng, W.; Li, D.; Wang, Y.; Liu, Y.; Zhu-Salzman, K. Cloning of heat shock protein genes (hsp70, hsc70 and hsp90) and their expression in response to larval diapause and thermal stress in the wheat blossom midge, Sitodiplosis mosellana. J. Insect Physiol. 2016, 95, 66–77. [Google Scholar] [CrossRef]
- Wang, H.; Dong, S.Z.; Kai, L.; Cui, H.; Ye, G.Y. A heat shock cognate 70 gene in the endoparasitoid, Pteromalus puparum, and its expression in relation to thermal stress. BMB Rep. 2008, 41, 388–393. [Google Scholar] [CrossRef] [Green Version]
- Sonoda, S.; Ashfaq, M.; Tsumuki, H. Cloning and nucleotide sequencing of three heat shock protein genes (hsp90, hsc70, and hsp19. 5) from the diamondback moth, Plutella xylostella (L.) and their expression in relation to developmental stage and temperature. Arch. Insect Biochem. Physiol. 2006, 62, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Koehn, R.K.; Bayne, B.L. Towards a physiological and genetical understanding of the energetics of the stress response. Biol. J. Linn. Soc. 1989, 37, 157–171. [Google Scholar] [CrossRef]
- Ming, L.I.; Wen-Cai, L.U.; Feng, H.Z.; Yuan, L.; Wang, J.J.; Lin, H.E. Cloning and expression analysis of a heat shock protein 70 gene TCHSP70–4 from Tetranychus cinnabarinus (Acari:Tetranychidae). Acta Entomol. Sin. 2008, 14, 3036–3051. [Google Scholar]
- Andersen, J.P.; Schwartz, A.; Gramsbergen, J.B.; Loeschcke, V. Dopamine levels in the mosquito Aedes aegypti during adult development, following blood feeding and in response to heat stress. J. Insect Physiol. 2006, 52, 1163–1170. [Google Scholar] [CrossRef] [PubMed]
- Sarup, P.; Sørensen, J.; Dimitrov, K.; Barker, J.; Loeschcke, V. Climatic adaptation of Drosophila buzzatii populations in southeast Australia. Heredity 2006, 96, 479–486. [Google Scholar] [CrossRef]
- Wei, J.; Wang, R.; Wan, F. Comparative analysis of the copy numbers of hsp70 and hsp90 in Trialeurodes vaporariorum and three cryptic species of Bemisia tabaci complex (Hemiptera: Aleyrodidae). Acta Entomol. Sin. 2014, 57, 647–655. [Google Scholar] [CrossRef]
- Wen, L.-L.; Yang, F.; Chen, Y.-Y.; Guo, Y.-Y.; Wang, S.-Y.; Jiao, X.-G. Comparison of stress tolerance between the two wolf spiders Pardosa astrigera and Pardosa pseudoannulata. J. Hubei Univ. (Nat. Sci.) 2015, 37, 231–234. [Google Scholar]
- Krishnan, N.; Kodrík, D. Endocrine control of oxidative stress in insects. In Oxidative Stress in Vertebrates and Invertebrates: Molecular Aspects of Cell Signaling; John Wiley & Sons: Hoboken, NJ, USA, 2011; pp. 259–270. [Google Scholar]
- Chen, H.; Sarwar, S.G.; Guo, J.; Wan, F.; Zhou, Z. Antioxidant Responses of Ragweed Leaf Beetle Ophraella communa (Coleoptera: Chrysomelidae) Exposed to Thermal Stress. Front. Physiol. 2018, 9, e808. [Google Scholar] [CrossRef] [Green Version]
- Ju, R.-T.; Wei, H.-P.; Wang, F.; Zhou, X.-H.; Li, B. Anaerobic respiration and antioxidant responses of Corythucha ciliata (Say) adults to heat-induced oxidative stress under laboratory and field conditions. Cell Stress Chaperones 2014, 19, 255–262. [Google Scholar] [CrossRef] [Green Version]
- Lu, F.; Chen, Q.; Chen, Z.; Lu, H.; Xu, X.; Jing, F. Effects of heat stress on development, reproduction and activities of protective enzymes in Mononychellus mcgregori. Exp. Appl. Acarol. 2014, 63, 267–284. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, Y.; He, M.; Yun, Y.; Peng, Y. Antioxidant responses of the pest natural enemy Hylyphantes graminicola (Araneae: Linyphiidae) exposed to short-term heat stress. J. Therm. Biol. 2020, 87, 102477. [Google Scholar] [CrossRef]
- Ali, A.; Rashid, M.; Huang, Q.; Wong, C.; Lei, C.-L. Response of antioxidant enzymes in Mythimna separata (Lepidoptera: Noctuidae) exposed to thermal stress. Bull. Entomol. Res. 2017, 107, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Yu, N.; Christiaens, O.; Liu, J.; Niu, J.; Cappelle, K.; Caccia, S.; Huvenne, H.; Smagghe, G. Delivery of dsRNA for RNAi in insects: An overview and future directions. Insect Sci. 2013, 20, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Li, C.; Bao, H.; Fang, J.; Liu, Z.; Zhang, Y. Validating the importance of two acetylcholinesterases in insecticide sensitivities by RNAi in Pardosa pseudoannulata, an important predatory enemy against several insect pests. Pestic. Biochem. Physiol. 2015, 125, 26–30. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.M.; Lu, T.Y.; Wu, Y.; Yu, N.; Xu, G.M.; Han, Q.Q.; Liu, Z.W. The importance of vitellogenin receptors in the oviposition of the pond wolf spider, Pardosa pseudoannulata. Insect Sci. 2022, 29, 443–452. [Google Scholar] [CrossRef]
- Niu, D.; Zhao, Y.; Gong, X.; Yang, R.; Hu, L.; Zhang, W. Stress response and silencing verification of heat shock proteins in Dermatophagoides farinae under temperature stress. Int. J. Biol. Macromol. 2020, 144, 351–361. [Google Scholar] [CrossRef]
- Kim, M.; Lee, S.; Chun, Y.S.; Na, J.; Kwon, H.; Kim, W.; Kim, Y. Heat tolerance induction of the Indian meal moth (Lepidoptera: Pyralidae) is accompanied by upregulation of heat shock proteins and polyols. Environ. Entomol. 2017, 46, 1005–1011. [Google Scholar] [CrossRef]
- Yang, X.-Q.; Zhang, Y.-L.; Wang, X.-Q.; Dong, H.; Gao, P.; Jia, L.-Y. Characterization of multiple heat-shock protein transcripts from Cydia pomonella: Their response to extreme temperature and insecticide exposure. J. Agric. Food Chem. 2016, 64, 4288–4298. [Google Scholar] [CrossRef]
- Elisabeth, G.; Alexandre, G.; Christine, H.; Ivan, I.; Appel, R.D.; Amos, B. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [Green Version]
- Edouard, D.C.; Sigrist, C.J.A.; Alexandre, G.; Virginie, B.; Langendijk-Genevaux, P.S.; Elisabeth, G.; Amos, B.; Nicolas, H. ScanProsite: Detection of PROSITE signature matches and ProRule-associated functional and structural residues in proteins. Nucleic Acids Res. 2006, 34, W362–W365. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.M.; Huang, H.L.; Yuan, Y.; Wan, T.Y.; Jiang, C.F.; Dai, Z.H.; Xiong, S.L.; Cao, M.H.; Tu, S.X. Silicone stressed response of crayfish (Procambarus clarkii) in antioxidant enzyme activity and related gene expression. Environ. Pollut. 2021, 274, 115836. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Names | Primer Sequences | Use of Primers |
|---|---|---|
| PpHSP27-1F | GGTTCCAGAAGCGTCTCT | Amplification of internal HSP genes fragment |
| PpHSP27-1R | CGTCCGAGTCCTGTATGAG | |
| PpHSP27-2F | CATTCAGAGCCACAGCACAG | |
| PpHSP27-2R | TTCAGAAGCGGTTACAGAGAAC | |
| PpHSP60-F | GGTGTAGATGTCTTGGCTGAT | |
| PpHSP60-R | CAGTTGGAATGGCATGAGTTC | |
| PpHSC70-F | ATGTGTTGCTGTTATGGATGG | |
| PpHSC70-R | ATGCTTGCTGTAGTTCTTGTG | |
| 3′-PpHSP27-F | CAGATGGCGTCCTCACAGT | 3′RACE |
| 3′-PpHSP60-F | GATGCTGCTGTAGTCACTCA | |
| 3′-PpHSC70-F | AGAGTCACCAGAAACATTGCGTAAAG | |
| 5′-PpHSP27-R | TATGAGGGGCGAGCTGAGTCCGTCGAGC | 5′RACE |
| 5′-PpHSP60-R | GCCTCCAACCTTGAGAAGTGCAACACCA | |
| 5′-PpHSC70-R | CCCTTGAGCTTCAACCCAAGCATCACC | |
| qPpHSP27-F | GGTTCAAGCCAAACACGAAGA | qPCR |
| qPpHSP27-R | GTGAGGACGCCATCTGTGA | |
| qPpHSP60-F | TGATGCTGCTGTAGTCACTCA | |
| qPpHSP60-R | ACCCATACCGCCCATACCT | |
| qPpHSC70-F | GGCTATATCTGATGCTGATGTGA | |
| qPpHSC70-R | AAGAACACCACCCTGAATTGC | |
| β-actin-F | GACCCAATACTTCTAACG | |
| β-actin-R | ACAGCAGGAAACACTTA | |
| dsPpHSP27-F | taatacgactcactatagggTGAGGAGTGACTTCAGCGTG | RNAi |
| dsPpHSP27-R | taatacgactcactatagggCAGGCTGATATTGGCTGACA | |
| dsPpHSP60-F | taatacgactcactatagggCTCCAGAAGAAATAGCCCAGGTT | |
| dsPpHSP60-R | taatacgactcactatagggAACGAGATCAGCATCATCACCAA | |
| dsPpHSC70-F | taatacgactcactatagggGCTGGTGATGTCACAGATGTT | |
| dsPpHSC70-R | taatacgactcactatagggCTTCTTCAGGTAACTGGCTCTT | |
| dsGFP-F | taatacgactcactatagggGCCAACACTTGTCACTACTT | |
| dsGFP-R | taatacgactcactatagggGGAGTATTTTGTTGATAATGGTC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, D.; Liu, J.; Pan, Y.-N.; Zhu, J.-Y.; Xiao, F.; Liu, M.; Xiao, R. Three Heat Shock Protein Genes and Antioxidant Enzymes Protect Pardosa pseudoannulata (Araneae: Lycosidae) from High Temperature Stress. Int. J. Mol. Sci. 2022, 23, 12821. https://doi.org/10.3390/ijms232112821
Fu D, Liu J, Pan Y-N, Zhu J-Y, Xiao F, Liu M, Xiao R. Three Heat Shock Protein Genes and Antioxidant Enzymes Protect Pardosa pseudoannulata (Araneae: Lycosidae) from High Temperature Stress. International Journal of Molecular Sciences. 2022; 23(21):12821. https://doi.org/10.3390/ijms232112821
Chicago/Turabian StyleFu, Di, Jing Liu, Ying-Na Pan, Jia-Yun Zhu, Feng Xiao, Min Liu, and Rong Xiao. 2022. "Three Heat Shock Protein Genes and Antioxidant Enzymes Protect Pardosa pseudoannulata (Araneae: Lycosidae) from High Temperature Stress" International Journal of Molecular Sciences 23, no. 21: 12821. https://doi.org/10.3390/ijms232112821