
Hydroxygenkwanin Improves the Efficacy of Cytotoxic Drugs in ABCG2-Overexpressing Multidrug-Resistant Cancer Cells
 , and
, and
Abstract
:1. Introduction
2. Results
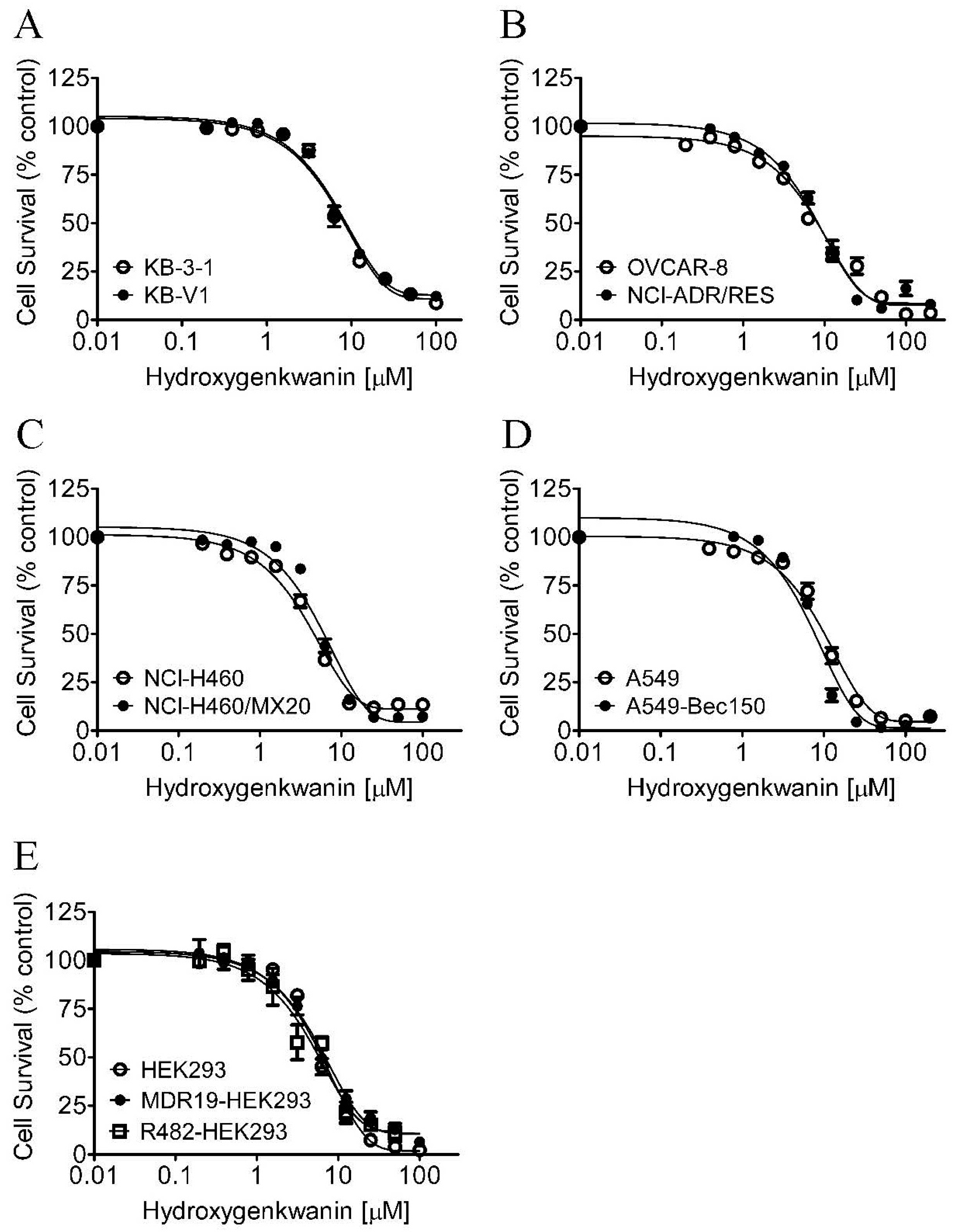
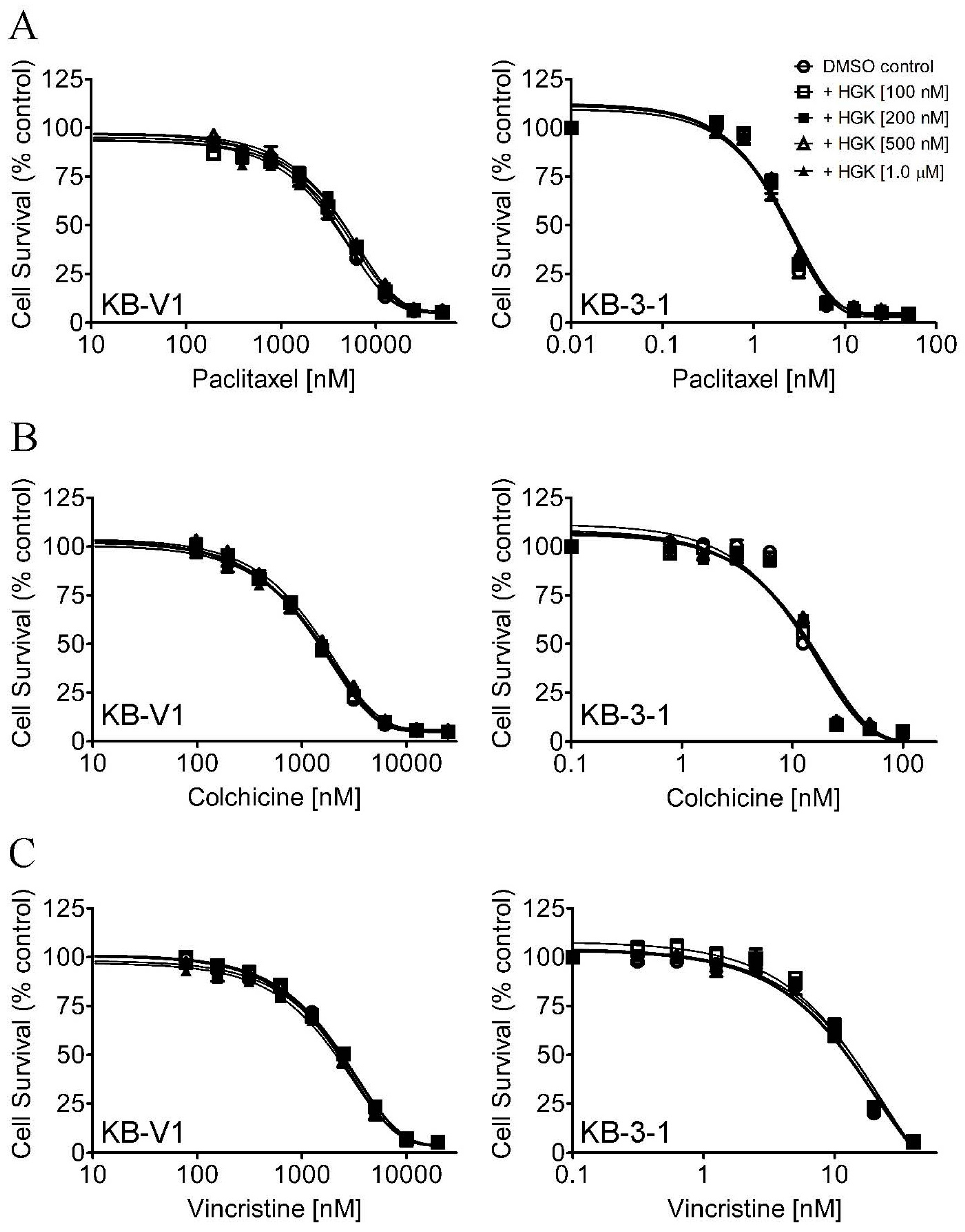
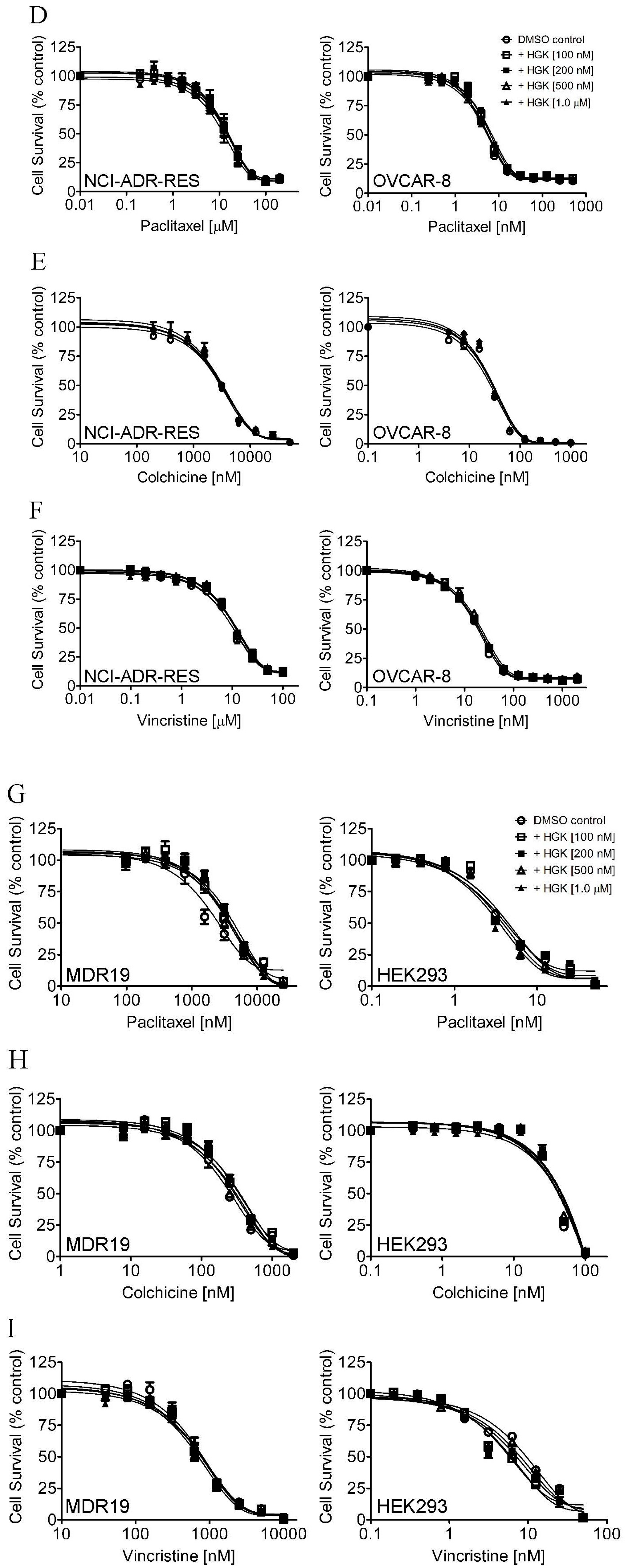
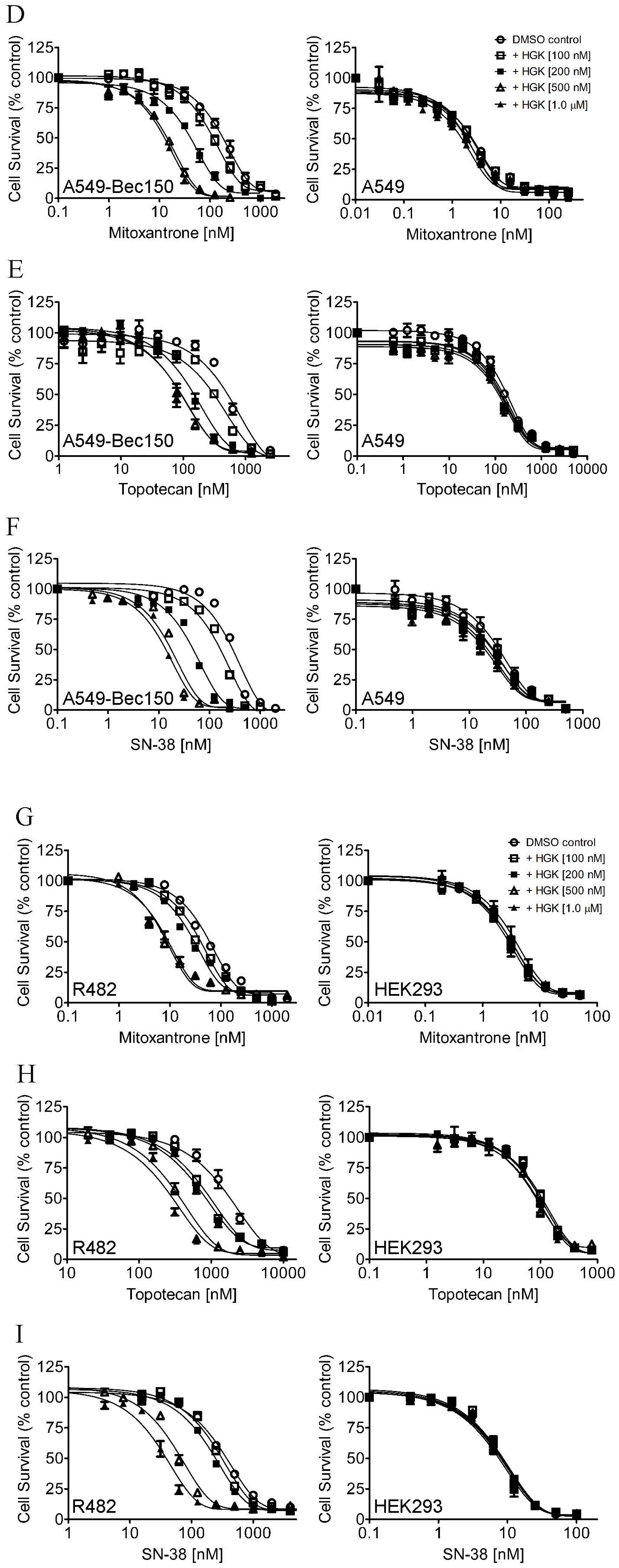
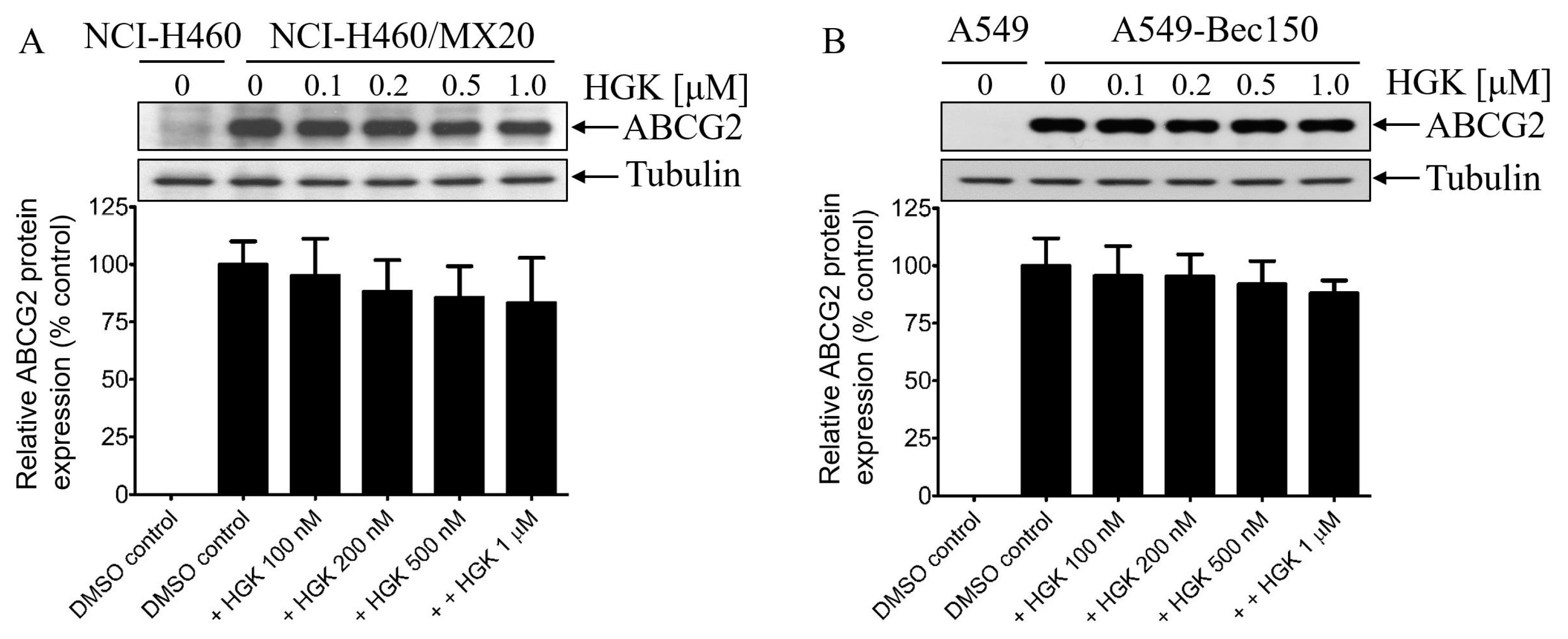
2.1. Hydroxygenkwanin Sensitizes ABCG2-Overexpressing Multidrug-Resistant Cells to Cytotoxic Anticancer Drugs
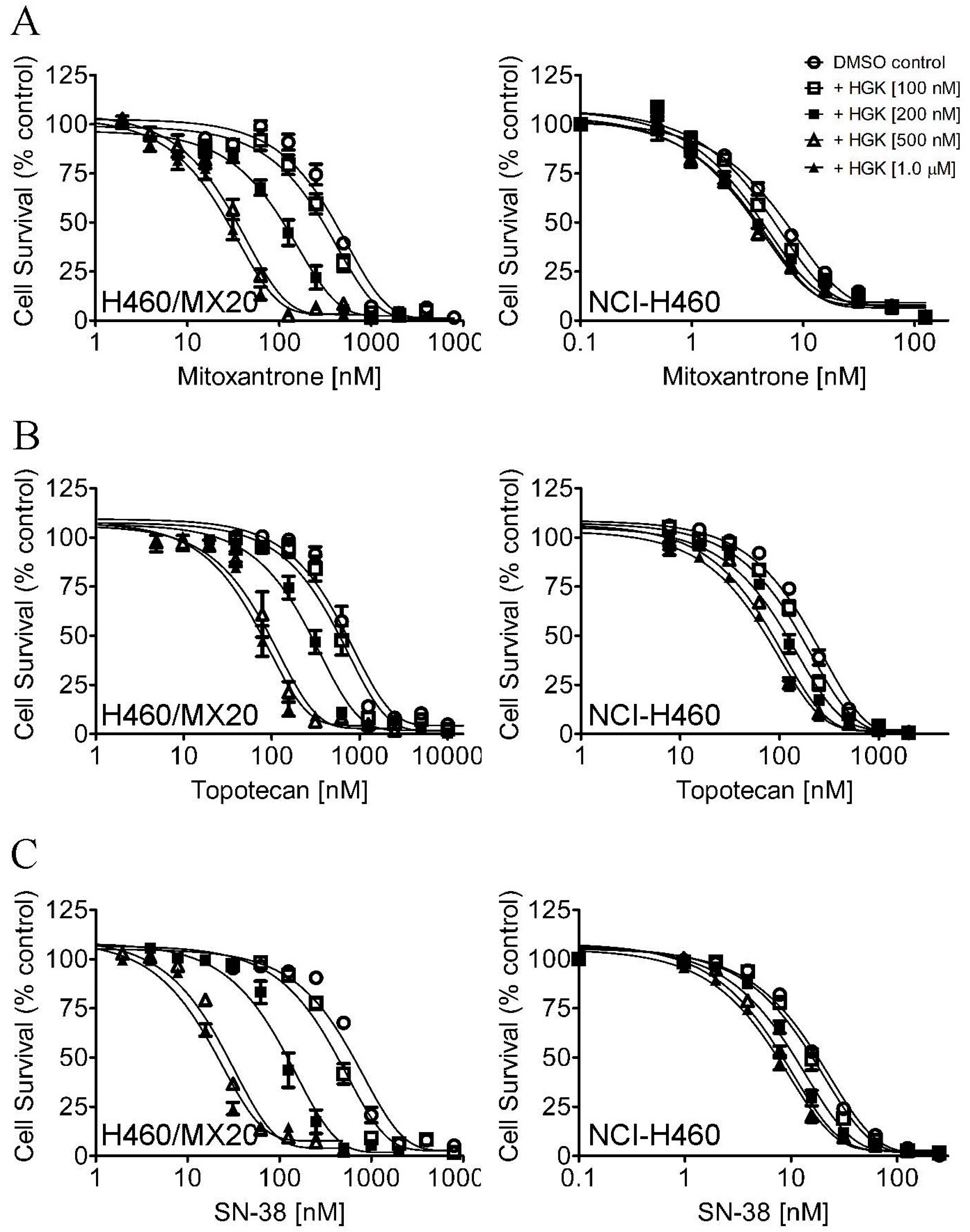
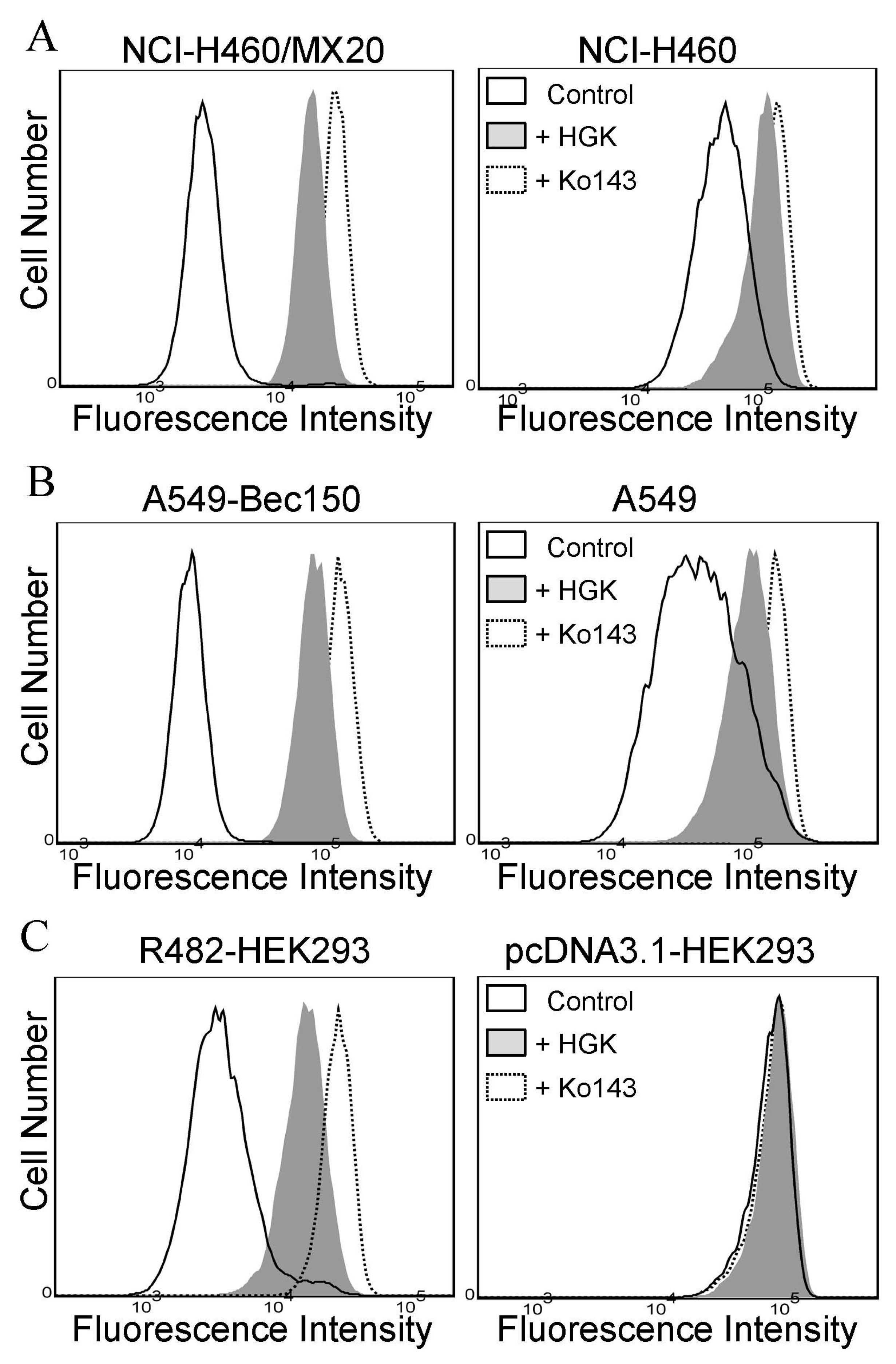
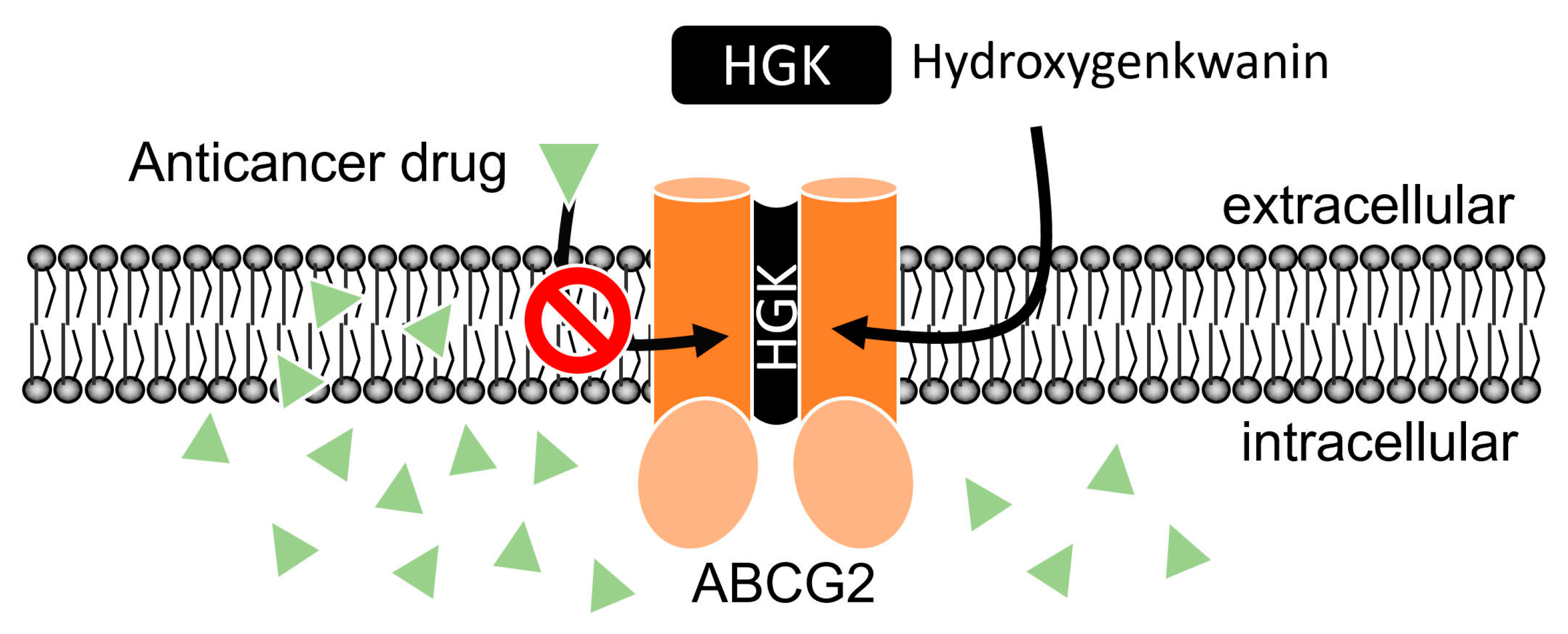
2.2. Hydroxygenkwanin Attenuates the Drug Transport Function of ABCG2
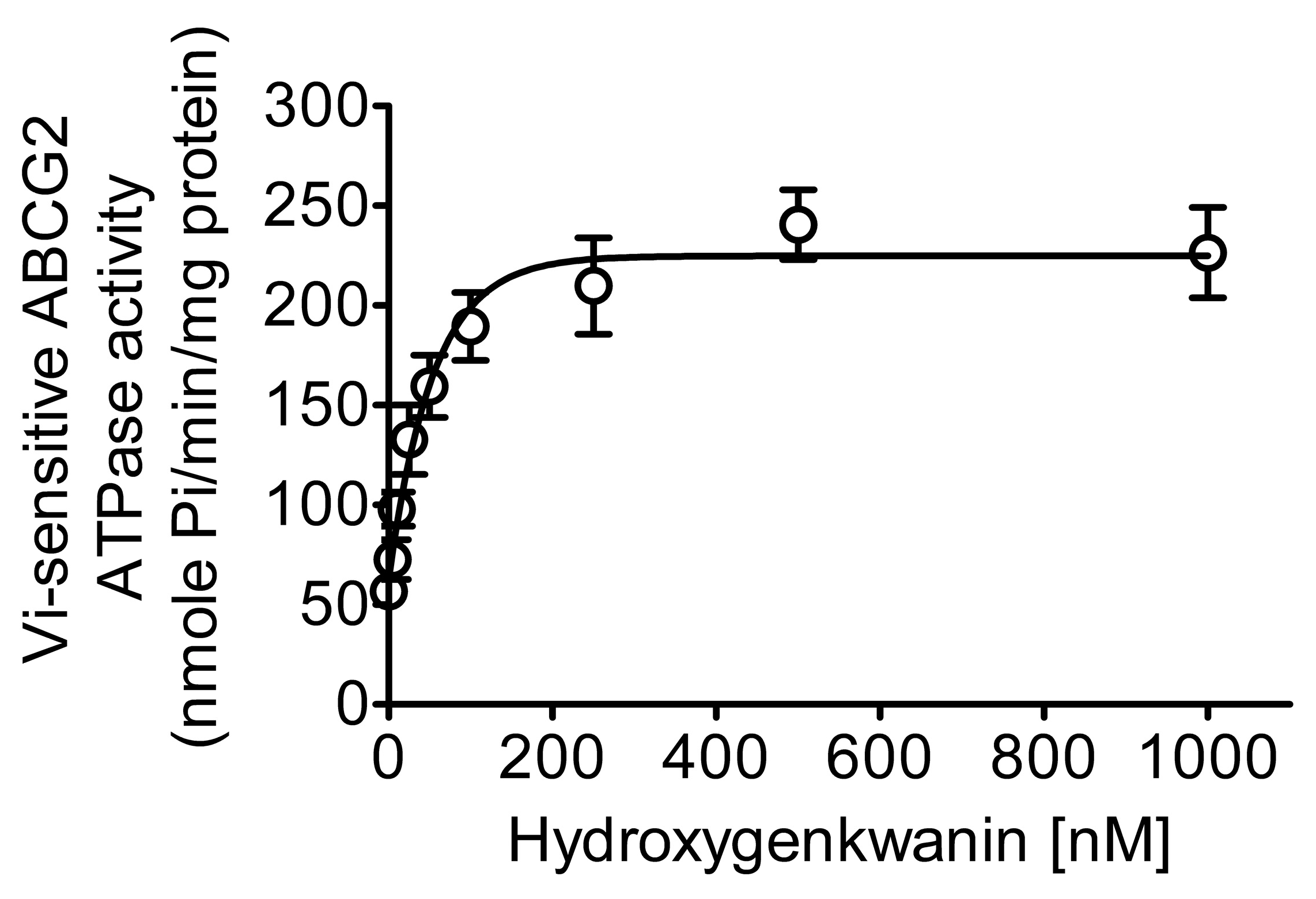
2.3. Hydroxygenkwanin Stimulates the ATPase Activity of ABCG2
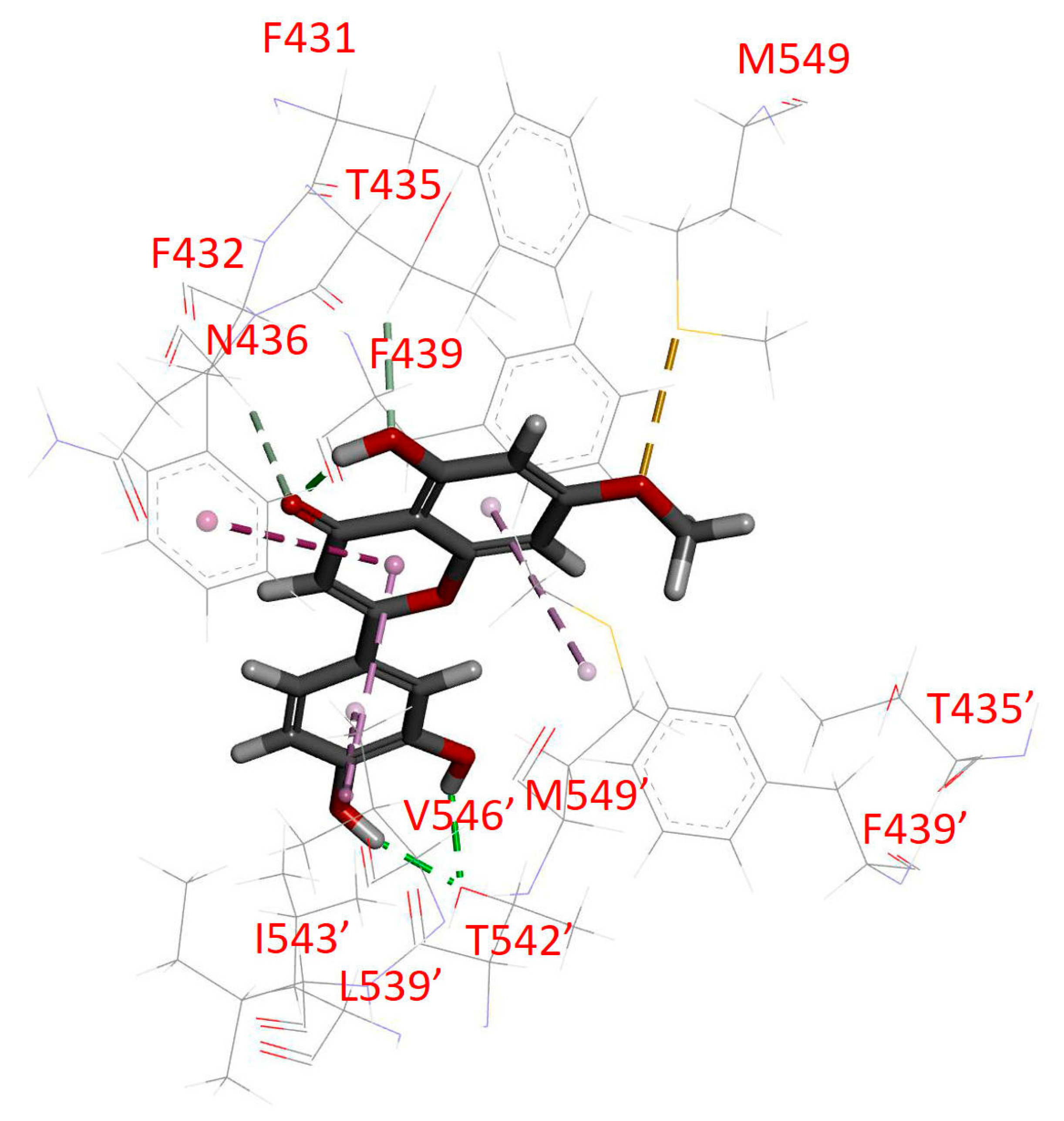
2.4. Docking of Hydroxygenkwanin in the Drug-Binding Pocket of ABCG2
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Cell Lines
4.3. Cell Viability Assay
4.4. Flow Cytometry
4.5. Immunoblotting
4.6. ATPase Assay
4.7. Docking Analysis
4.8. Data Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Szakacs, G.; Paterson, J.K.; Ludwig, J.A.; Booth-Genthe, C.; Gottesman, M.M. Targeting multidrug resistance in cancer. Nat. Rev. Drug Discov. 2006, 5, 219–234. [Google Scholar] [CrossRef]
- Robey, R.W.; Pluchino, K.M.; Hall, M.D.; Fojo, A.T.; Bates, S.E.; Gottesman, M.M. Revisiting the role of ABC transporters in multidrug-resistant cancer. Nat. Rev. Cancer 2018, 18, 452–464. [Google Scholar] [CrossRef] [PubMed]
- Breedveld, P.; Pluim, D.; Cipriani, G.; Wielinga, P.; van Tellingen, O.; Schinkel, A.H.; Schellens, J.H. The effect of Bcrp1 (Abcg2) on the in vivo pharmacokinetics and brain penetration of imatinib mesylate (Gleevec): Implications for the use of breast cancer resistance protein and P-glycoprotein inhibitors to enable the brain penetration of imatinib in patients. Cancer Res. 2005, 65, 2577–2582. [Google Scholar] [PubMed] [Green Version]
- Sarkadi, B.; Homolya, L.; Szakacs, G.; Váradi, A. Human multidrug resistance ABCB and ABCG transporters: Participation in a chemoimmunity defense system. Physiol. Rev. 2006, 86, 1179–1236. [Google Scholar] [CrossRef] [PubMed]
- Giacomini, K.M.; Huang, S.M.; Tweedie, D.J.; Benet, L.Z.; Brouwer, K.L.; Chu, X.; Dahlin, A.; Evers, R.; Fischer, V.; Hillgren, K.M.; et al. Membrane transporters in drug development. Nat. Rev. Drug Discov. 2010, 9, 215–236. [Google Scholar] [CrossRef] [PubMed]
- Tsunoda, S.; Okumura, T.; Ito, T.; Kondo, K.; Ortiz, C.; Tanaka, E.; Watanabe, G.; Itami, A.; Sakai, Y.; Shimada, Y. ABCG2 expression is an independent unfavorable prognostic factor in esophageal squamous cell carcinoma. Oncology 2006, 71, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Zeng, H.; Ying, K. The combination of stem cell markers CD133 and ABCG2 predicts relapse in stage I non-small cell lung carcinomas. Med. Oncol. 2011, 28, 1458–1462. [Google Scholar] [CrossRef]
- Hang, D.; Dong, H.-C.; Ning, T.; Dong, B.; Hou, D.-L.; Xu, W.-G. Prognostic value of the stem cell markers CD133 and ABCG2 expression in esophageal squamous cell carcinoma. Dis. Esophagus 2012, 25, 638–644. [Google Scholar] [CrossRef]
- Kovalev, A.A.; Tsvetaeva, D.A.; Grudinskaja, T.V. Role of ABC-cassette transporters (MDR1, MRP1, BCRP) in the development of primary and acquired multiple drug resistance in patients with early and metastatic breast cancer. Exp. Oncol. 2013, 35, 287–290. [Google Scholar]
- Ross, D.D.; Karp, J.E.; Chen, T.T.; Doyle, L.A. Expression of breast cancer resistance protein in blast cells from patients with acute leukemia. Blood 2000, 96, 365–368. [Google Scholar] [CrossRef] [PubMed]
- Steinbach, D.; Sell, W.; Voigt, A.; Hermann, J.; Zintl, F.; Sauerbrey, A. BCRP gene expression is associated with a poor response to remission induction therapy in childhood acute myeloid leukemia. Leukemia 2002, 16, 1443–1447. [Google Scholar] [CrossRef] [Green Version]
- Schwarzenbach, H. Expression of MDR1/P-glycoprotein, the multidrug resistance protein MRP, and the lung-resistance protein LRP in multiple myeloma. Med. Oncol. 2002, 19, 87–104. [Google Scholar] [CrossRef]
- Uggla, B.; Ståhl, E.; Wågsäter, D.; Paul, C.; Karlsson, M.G.; Sirsjö, A.; Tidefelt, U. BCRP mRNA expression v. clinical outcome in 40 adult AML patients. Leuk. Res. 2005, 29, 141–146. [Google Scholar] [CrossRef]
- Matthews, C.; Catherwood, M.A.; Larkin, A.-M.; Clynes, M.; Morris, T.C.; Alexander, H.D. MDR-1, but not MDR-3 gene expression, is associated with unmutated IgVH genes and poor prognosis chromosomal aberrations in chronic lymphocytic leukemia. Leuk. Lymphoma 2006, 47, 2308–2313. [Google Scholar] [CrossRef]
- Turner, J.G.; Gump, J.L.; Zhang, C.; Cook, J.M.; Marchion, D.; Hazlehurst, L.; Munster, P.; Schell, M.J.; Dalton, W.S.; Sullivan, D.M. ABCG2 expression, function, and promoter methylation in human multiple myeloma. Blood 2006, 108, 3881–3889. [Google Scholar] [CrossRef] [PubMed]
- Tsubaki, M.; Satou, T.; Itoh, T.; Imano, M.; Komai, M.; Nishinobo, M.; Yamashita, M.; Yanae, M.; Yamazoe, Y.; Nishida, S. Overexpression of MDR1 and survivin, and decreased Bim expression mediate multidrug-resistance in multiple myeloma cells. Leuk. Res. 2012, 36, 1315–1322. [Google Scholar] [CrossRef]
- Gillet, J.-P.; Gottesman, M.M. Mechanisms of multidrug resistance in cancer. In Multi-Drug Resistance in Cancer; Methods in Molecular Biology; Springer: Berlin/Heidelberg, Germany, 2010; Volume 596, pp. 47–76. [Google Scholar] [CrossRef]
- Wu, C.-P.; Calcagno, A.M.; Ambudkar, S.V. Reversal of ABC drug transporter-mediated multidrug resistance in cancer cells: Evaluation of current strategies. Curr. Mol. Pharmacol. 2008, 1, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Nobili, S.; Landini, I.; Giglioni, B.; Mini, E. Pharmacological strategies for overcoming multidrug resistance. Curr. Drug Targets 2006, 7, 861–879. [Google Scholar] [CrossRef]
- Adamska, A.; Falasca, M. ATP-binding cassette transporters in progression and clinical outcome of pancreatic cancer: What is the way forward? World J. Gastroenterol. 2018, 24, 3222–3238. [Google Scholar] [CrossRef]
- Xiao, H.; Zheng, Y.; Ma, L.; Tian, L.; Sun, Q. Clinically-Relevant ABC Transporter for Anti-Cancer Drug Resistance. Front. Pharmacol. 2021, 12, 648407. [Google Scholar] [CrossRef]
- Engle, K.; Kumar, G. Cancer multidrug-resistance reversal by ABCB1 inhibition: A recent update. Eur. J. Med. Chem. 2022, 239, 114542. [Google Scholar] [CrossRef] [PubMed]
- Zattoni, I.F.; Delabio, L.C.; Dutra, J.D.P.; Kita, D.H.; Scheiffer, G.; Hembecker, M.; Pereira, G.D.S.; Moure, V.R.; Valdameri, G. Targeting breast cancer resistance protein (BCRP/ABCG2): Functional inhibitors and expression modulators. Eur. J. Med. Chem. 2022, 237, 114346. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Wu, C.-P.; Ambudkar, S.V. Development of inhibitors of ATP-binding cassette drug transporters—Present status and challenges. Expert Opin. Drug Metab. Toxicol. 2008, 4, 205–223. [Google Scholar] [CrossRef]
- Wu, C.P.; Ohnuma, S.; Ambudkar, S.V. Discovering natural product modulators to overcome multidrug resistance in cancer chemotherapy. Curr. Pharm. Biotechnol. 2011, 12, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Jaitak, V. Natural products as multidrug resistance modulators in cancer. Eur. J. Med. Chem. 2019, 176, 268–291. [Google Scholar] [CrossRef]
- Ye, Q.; Liu, K.; Shen, Q.; Li, Q.; Hao, J.; Han, F.; Jiang, R.-W. Reversal of Multidrug Resistance in Cancer by Multi-Functional Flavonoids. Front. Oncol. 2019, 9, 487. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.-P.; Lusvarghi, S.; Hsiao, S.-H.; Liu, T.-C.; Li, Y.-Q.; Huang, Y.-H.; Hung, T.-H.; Ambudkar, S.V. Licochalcone A Selectively Resensitizes ABCG2-Overexpressing Multidrug-Resistant Cancer Cells to Chemotherapeutic Drugs. J. Nat. Prod. 2020, 83, 1461–1472. [Google Scholar] [CrossRef]
- Wu, C.-P.; Li, Y.-Q.; Hung, T.-H.; Chang, Y.-T.; Huang, Y.-H.; Wu, Y.-S. Sophoraflavanone G Resensitizes ABCG2-Overexpressing Multidrug-Resistant Non-Small-Cell Lung Cancer Cells to Chemotherapeutic Drugs. J. Nat. Prod. 2021, 84, 2544–2553. [Google Scholar] [CrossRef]
- Sanders, K.; Moran, Z.; Shi, Z.; Paul, R.; Greenlee, H. Natural Products for Cancer Prevention: Clinical Update 2016. Semin. Oncol. Nurs. 2016, 32, 215–240. [Google Scholar] [CrossRef]
- Li, T.; Wang, N.; Zhang, T.; Zhang, B.; Sajeevan, T.P.; Joseph, V.; Armstrong, L.; He, S.; Yan, X.; Naman, C.B. A Systematic Review of Recently Reported Marine Derived Natural Product Kinase Inhibitors. Mar. Drugs 2019, 17, 493. [Google Scholar] [CrossRef]
- Khalifa, S.A.M.; Elias, N.; Farag, M.A.; Chen, L.; Saeed, A.; Hegazy, M.-E.F.; Moustafa, M.S.; Abd El-Wahed, A.; Al-Mousawi, S.M.; Musharraf, S.G.; et al. Marine Natural Products: A Source of Novel Anticancer Drugs. Mar. Drugs 2019, 17, 491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, H.; Liang, Y.; Ito, Y.; Wang, X.; Chen, R.; He, J.; Li, H.; Zhang, T. Preparative Isolation and Purification of Four Flavonoids from Daphne Genkwa Sieb. Et Zucc. By High-Speed Countercurrent Chromatography. J. Liq. Chromatogr. Relat. Technol. 2011, 34, 2360–2372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.-W.; Bao, Y.; Yu, H.; Chen, Q.J.; Lu, F.; Zhai, S.; Zhang, C.-F.; Li, F.; Wang, C.-Z.; Yuan, C.-S. Anti-rheumatoid arthritis effects of flavonoids from Daphne genkwa. Int. Immunopharmacol. 2020, 83, 106384. [Google Scholar] [CrossRef]
- Zhou, D.-C.; Zheng, G.; Jia, L.-Y.; He, X.; Zhang, C.-F.; Wang, C.-Z.; Yuan, C.-S. Comprehensive evaluation on anti-inflammatory and anti-angiogenic activities in vitro of fourteen flavonoids from Daphne Genkwa based on the combination of efficacy coefficient method and principal component analysis. J. Ethnopharmacol. 2021, 268, 113683. [Google Scholar] [CrossRef]
- Mohamed, T.A.; Hegazy, M.-E.F.; Abd El Aty, A.A.; Ghabbour, H.A.; Alsaid, M.S.; Shahat, A.A.; Paré, P.W. Antimicrobial sesquiterpene lactones from Artemisia sieberi. J. Asian Nat. Prod. Res. 2017, 19, 1093–1101. [Google Scholar] [CrossRef]
- Taguchi, N.; Yuriguchi, M.; Ando, T.; Kitai, R.; Aoki, H.; Kunisada, T. Flavonoids with Two OH Groups in the B-Ring Promote Pigmented Hair Regeneration. Biol. Pharm. Bull. 2019, 42, 1446–1449. [Google Scholar] [CrossRef] [Green Version]
- Shen, H.-Y.; Li, X.-Q.; Fan, W.-Q.; Wang, Y.-W.; Huang, F.; Wu, J.-Q.; Zhang, W.; Feng, X.-S.; Chao, X. A systematic approach to decode the mechanism of Cornus in the treatment of hepatocellular carcinoma (HCC). Eur. J. Pharmacol. 2021, 909, 174405. [Google Scholar] [CrossRef]
- Chen, C.-C.; Chen, C.-Y.; Cheng, S.-F.; Shieh, T.-M.; Leu, Y.-L.; Chuang, W.-Y.; Liu, K.-T.; Ueng, S.-H.; Shih, Y.-H.; Chou, L.-F.; et al. Hydroxygenkwanin Increases the Sensitivity of Liver Cancer Cells to Chemotherapy by Inhibiting DNA Damage Response in Mouse Xenograft Models. Int. J. Mol. Sci. 2021, 22, 9766. [Google Scholar] [CrossRef]
- Chen, C.-Y.; Chen, C.-C.; Chuang, W.-Y.; Leu, Y.-L.; Ueng, S.-H.; Hsueh, C.; Yeh, C.-T.; Wang, T.-H. Hydroxygenkwanin Inhibits Class I HDAC Expression and Synergistically Enhances the Antitumor Activity of Sorafenib in Liver Cancer Cells. Front. Oncol. 2020, 10, 216. [Google Scholar] [CrossRef]
- Leu, Y.-L.; Wang, T.-H.; Wu, C.-C.; Huang, K.-Y.; Jiang, Y.-W.; Hsu, Y.-C.; Chen, C.-Y. Hydroxygenkwanin Suppresses Non-Small Cell Lung Cancer Progression by Enhancing EGFR Degradation. Molecules 2020, 25, 941. [Google Scholar] [CrossRef]
- Chou, L.-F.; Chen, C.-Y.; Yang, W.-H.; Chen, C.-C.; Chang, J.-L.; Leu, Y.-L.; Liou, M.-J.; Wang, T.-H. Suppression of Hepatocellular Carcinoma Progression through FOXM1 and EMT Inhibition via Hydroxygenkwanin-Induced miR-320a Expression. Biomolecules 2019, 10, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.-C.; Lee, P.-C.; Wang, J.J.; Hsu, Y.-C. Anticancer Effect and Mechanism of Hydroxygenkwanin in Oral Squamous Cell Carcinoma. Front. Oncol. 2019, 9, 911. [Google Scholar] [CrossRef]
- Li, N.; Liu, J.-H.; Zhang, J.; Yu, B.-Y. Comparative evaluation of cytotoxicity and antioxidative activity of 20 flavonoids. J. Agric. Food Chem. 2008, 56, 3876–3883. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, Y.S.; Yin, L.H.; Xu, L.N.; Peng, J.Y.; Zhou, H.; Kang, W. Synergistic anti-glioma effect of Hydroxygenkwanin and Apigenin in vitro. Chem. Biol. Interact. 2013, 206, 346–355. [Google Scholar] [CrossRef] [PubMed]
- Kartner, N.; Riordan, J.R.; Ling, V. Cell surface P-glycoprotein associated with multidrug resistance in mammalian cell lines. Science 1983, 221, 1285–1288. [Google Scholar] [CrossRef]
- Maliepaard, M.; van Gastelen, M.A.; de Jong, L.A.; Pluim, D.; van Waardenburg, R.C.; Ruevekamp-Helmers, M.C.; Floot, B.G.; Schellens, J.H. Overexpression of the BCRP/MXR/ABCP gene in a topotecan-selected ovarian tumor cell line. Cancer Res. 1999, 59, 4559–4563. [Google Scholar]
- Robey, R.W.; Medina-Pérez, W.Y.; Nishiyama, K.; Lahusen, T.; Miyake, K.; Litman, T.; Senderowicz, A.M.; Ross, D.D.; Bates, S.E. Overexpression of the ATP-binding Cassette Half-Transporter, ABCG2 (MXR/BCRP/ABCP1), in Flavopiridol-resistant Human Breast Cancer Cells. Clin. Cancer Res. 2001, 7, 145–152. [Google Scholar]
- Bates, S.E.; Medina-Pérez, W.Y.; Kohlhagen, G.; Antony, S.; Nadjem, T.; Robey, R.W.; Pommier, Y. ABCG2 mediates differential resistance to SN-38 (7-ethyl-10-hydroxycamptothecin) and homocamptothecins. J. Pharmacol. Exp. Ther. 2004, 310, 836–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galetti, M.; Petronini, P.G.; Fumarola, C.; Cretella, D.; La Monica, S.; Bonelli, M.; Cavazzoni, A.; Saccani, F.; Caffarra, C.; Andreoli, R.; et al. Effect of ABCG2/BCRP Expression on Efflux and Uptake of Gefitinib in NSCLC Cell Lines. PLoS ONE 2015, 10, e0141795. [Google Scholar] [CrossRef]
- Robey, R.W.; Honjo, Y.; van de Laar, A.; Miyake, K.; Regis, J.T.; Litman, T.; Bates, S.E. A functional assay for detection of the mitoxantrone resistance protein, MXR (ABCG2). Biochim. Biophys. Acta (BBA)—Biomembr. 2001, 1512, 171–182. [Google Scholar] [CrossRef] [Green Version]
- Dai, C.-L.; Tiwari, A.K.; Wu, C.-P.; Su, X.-D.; Wang, S.-R.; Liu, D.-G.; Ashby, C.R., Jr.; Huang, Y.; Robey, R.W.; Liang, Y.-J.; et al. Lapatinib (Tykerb, GW572016) reverses multidrug resistance in cancer cells by inhibiting the activity of ATP-binding cassette subfamily B member 1 and G member 2. Cancer Res. 2008, 68, 7905–7914. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, A.K.; Sodani, K.; Dai, C.-L.; Abuznait, A.H.; Singh, S.; Xiao, Z.-J.; Patel, A.; Talele, T.T.; Fu, L.; Kaddoumi, A.; et al. Nilotinib potentiates anticancer drug sensitivity in murine ABCB1-, ABCG2-, and ABCC10-multidrug resistance xenograft models. Cancer Lett. 2013, 328, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.-P.; Hsiao, S.-H.; Murakami, M.; Lu, M.-J.; Li, Y.-Q.; Hsieh, C.-H.; Ambudkar, S.V.; Wu, Y.-S. Tyrphostin RG14620 selectively reverses ABCG2-mediated multidrug resistance in cancer cell lines. Cancer Lett. 2017, 409, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.-P.; Murakami, M.; Hsiao, S.-H.; Liu, T.-C.; Yeh, N.; Li, Y.-Q.; Hung, T.-H.; Wu, Y.-S.; Ambudkar, S.V. SIS3, a specific inhibitor of Smad3 reverses ABCB1- and ABCG2-mediated multidrug resistance in cancer cell lines. Cancer Lett. 2018, 433, 259–272. [Google Scholar] [CrossRef]
- Zhang, G.-N.; Zhang, Y.-K.; Wang, Y.-J.; Gupta, P.; Ashby, C.R., Jr.; Alqahtani, S.; Deng, T.; Bates, S.E.; Kaddoumi, A.; Wurpel, J.N.D.; et al. Epidermal growth factor receptor (EGFR) inhibitor PD153035 reverses ABCG2-mediated multidrug resistance in non-small cell lung cancer: In vitro and in vivo. Cancer Lett. 2018, 424, 19–29. [Google Scholar] [CrossRef]
- Toyoda, Y.; Takada, T.; Suzuki, H. Inhibitors of Human ABCG2: From Technical Background to Recent Updates with Clinical Implications. Front. Pharmacol. 2019, 10, 208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuestas, M.L.; Castillo, A.I.; Sosnik, A.; Mathet, V.L. Downregulation of mdr1 and abcg2 genes is a mechanism of inhibition of efflux pumps mediated by polymeric amphiphiles. Bioorgan. Med. Chem. Lett. 2012, 22, 6577–6579. [Google Scholar] [CrossRef]
- Bhullar, J.; Natarajan, K.; Shukla, S.; Mathias, T.J.; Sadowska, M.; Ambudkar, S.V.; Baer, M.R. The FLT3 inhibitor quizartinib inhibits ABCG2 at pharmacologically relevant concentrations, with implications for both chemosensitization and adverse drug interactions. PLoS ONE 2013, 8, e71266. [Google Scholar] [CrossRef] [Green Version]
- Natarajan, K.; Bhullar, J.; Shukla, S.; Burcu, M.; Chen, Z.-S.; Ambudkar, S.V.; Baer, M.R. The Pim kinase inhibitor SGI-1776 decreases cell surface expression of P-glycoprotein (ABCB1) and breast cancer resistance protein (ABCG2) and drug transport by Pim-1-dependent and -independent mechanisms. Biochem. Pharmacol. 2013, 85, 514–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robey, R.W.; Steadman, K.; Polgar, O.; Morisaki, K.; Blayney, M.; Mistry, P.; Bates, S.E. Pheophorbide a is a specific probe for ABCG2 function and inhibition. Cancer Res. 2004, 64, 1242–1246. [Google Scholar] [CrossRef] [Green Version]
- Özvegy, C.; Litman, T.; Szakacs, G.; Nagy, Z.; Bates, S.; Váradi, A.; Sarkadi, B. Functional characterization of the human multidrug transporter, ABCG2, expressed in insect cells. Biochem. Biophys. Res. Commun. 2001, 285, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Ozvegy-Laczka, C.; Hegedűs, T.; Várady, G.; Ujhelly, O.; Schuetz, J.D.; Váradi, A.; Kéri, G.; Örfi, L.; Német, K.; Sarkadi, B. High-affinity interaction of tyrosine kinase inhibitors with the ABCG2 multidrug transporter. Mol. Pharmacol. 2004, 65, 1485–1495. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.-P.; Hsiao, S.-H.; Sim, H.-M.; Luo, S.-Y.; Tuo, W.-C.; Cheng, H.-W.; Li, Y.-Q.; Huang, Y.-H.; Ambudkar, S.V. Human ABCB1 (P-glycoprotein) and ABCG2 mediate resistance to BI 2536, a potent and selective inhibitor of Polo-like kinase 1. Biochem. Pharmacol. 2013, 86, 904–913. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.-P.; Lusvarghi, S.; Wang, J.-C.; Hsiao, S.-H.; Huang, Y.-H.; Hung, T.-H.; Ambudkar, S.V. Avapritinib: A Selective Inhibitor of KIT and PDGFRα that Reverses ABCB1 and ABCG2-Mediated Multidrug Resistance in Cancer Cell Lines. Mol. Pharm. 2019, 16, 3040–3052. [Google Scholar] [CrossRef]
- Wu, C.-P.; Shukla, S.; Calcagno, A.M.; Hall, M.D.; Gottesman, M.M.; Ambudkar, S.V. Evidence for dual mode of action of a thiosemicarbazone, NSC73306: A potent substrate of the multidrug resistance linked ABCG2 transporter. Mol. Cancer Ther. 2007, 6, 3287–3296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orlando, B.J.; Liao, M. ABCG2 transports anticancer drugs via a closed-to-open switch. Nat. Commun. 2020, 11, 2264. [Google Scholar] [CrossRef]
- Kowal, J.; Ni, D.; Jackson, S.M.; Manolaridis, I.; Stahlberg, H.; Locher, K.P. Structural Basis of Drug Recognition by the Multidrug Transporter ABCG2. J. Mol. Biol. 2021, 433, 166980. [Google Scholar] [CrossRef]
- Jackson, S.M.; Manolaridis, I.; Kowal, J.; Zechner, M.; Taylor, N.M.I.; Bause, M.; Bauer, S.; Bartholomaeus, R.; Bernhardt, G.; Koenig, B.; et al. Structural basis of small-molecule inhibition of human multidrug transporter ABCG2. Nat. Struct. Mol. Biol. 2018, 25, 333–340. [Google Scholar] [CrossRef]
- Manolaridis, I.; Jackson, S.M.; Taylor, N.M.I.; Kowal, J.; Stahlberg, H.; Locher, K.P. Cryo-EM structures of a human ABCG2 mutant trapped in ATP-bound and substrate-bound states. Nature 2018, 563, 426–430. [Google Scholar] [CrossRef] [Green Version]
- Haider, A.J.; Cox, M.H.; Jones, N.; Goode, A.J.; Bridge, K.S.; Wong, K.; Briggs, D.; Kerr, I.D. Identification of residues in ABCG2 affecting protein trafficking and drug transport, using co-evolutionary analysis of ABCG sequences. Biosci. Rep. 2015, 35, e00241. [Google Scholar] [CrossRef] [Green Version]
- Weidner, L.D.; Zoghbi, S.S.; Lu, S.; Shukla, S.; Ambudkar, S.V.; Pike, V.W.; Mulder, J.; Gottesman, M.M.; Innis, R.B.; Hall, M.D. The Inhibitor Ko143 Is Not Specific for ABCG2. J. Pharmacol. Exp. Ther. 2015, 354, 384–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishiyama, M.; Kuga, T. Central effects of the neurotropic mycotoxin fumitremorgin A in the rabbit (I). Effects on the spinal cord. Jpn. J. Pharmacol. 1989, 50, 167–173. [Google Scholar] [CrossRef]
- Liu, K.; Zhu, J.; Huang, Y.; Li, C.; Lu, J.; Sachar, M.; Li, S.; Ma, X. Metabolism of KO143, an ABCG2 inhibitor. Drug Metab. Pharmacokinet. 2017, 32, 193–200. [Google Scholar] [CrossRef]
- Ahmed-Belkacem, A.; Pozza, A.; Munoz-Martinez, F.; Bates, S.E.; Castanys, S.; Gamarro, F.; Di Pietro, A.; Perez-Victoria, J.M. Flavonoid structure-activity studies identify 6-prenylchrysin and tectochrysin as potent and specific inhibitors of breast cancer resistance protein ABCG2. Cancer Res. 2005, 65, 4852–4860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, R.J.; Baptista, R.; Moreno, A.; Madeira, P.G.; Khonkarn, R.; Baubichon-Cortay, H.; Dos Santos, D.J.; Falson, P.; Ferreira, M.-J.U. Optimizing the flavanone core toward new selective nitrogen-containing modulators of ABC transporters. Future Med. Chem. 2018, 10, 725–741. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhai, Z.; Li, H.; Wang, X.; Huang, Y.; Su, X. Guajadial reverses multidrug resistance by inhibiting ABC transporter expression and suppressing the PI3K/Akt pathway in drug-resistant breast cancer cells. Chem. Biol. Interact. 2019, 305, 98–104. [Google Scholar] [CrossRef]
- Silbermann, K.; Shah, C.P.; Sahu, N.U.; Juvale, K.; Stefan, S.M.; Kharkar, P.S.; Wiese, M. Novel chalcone and flavone derivatives as selective and dual inhibitors of the transport proteins ABCB1 and ABCG2. Eur. J. Med. Chem. 2019, 164, 193–213. [Google Scholar] [CrossRef]
- Huang, W.; Wan, C.; Luo, Q.; Huang, Z.; Luo, Q. Genistein-inhibited cancer stem cell-like properties and reduced chemoresistance of gastric cancer. Int. J. Mol. Sci. 2014, 15, 3432–3443. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Wei, Y.-C.; Li, W.-C.; Chen, H.-Y.; Lin, H.-Y.; Chiang, C.-P.; Chen, H.-M. Natural Compounds Modulate Drug Transporter Mediated Oral Cancer Treatment. Biomolecules 2020, 10, 1335. [Google Scholar] [CrossRef] [PubMed]
- Shen, D.-W.; Fojo, A.; Chin, J.E.; Roninson, I.B.; Richert, N.; Pastan, I.; Gottesman, M.M. Human multidrug-resistant cell lines: Increased mdr1 expression can precede gene amplification. Science 1986, 232, 643–645. [Google Scholar] [CrossRef]
- Roschke, A.V.; Tonon, G.; Gehlhaus, K.S.; McTyre, N.; Bussey, K.J.; Lababidi, S.; Scudiero, D.A.; Weinstein, J.N.; Kirsch, I.R. Karyotypic complexity of the NCI-60 drug-screening panel. Cancer Res. 2003, 63, 8634–8647. [Google Scholar] [PubMed]
- Henrich, C.J.; Bokesch, H.R.; Dean, M.; Bates, S.E.; Robey, R.W.; Goncharova, E.I.; Wilson, J.A.; McMahon, J.B. A high-throughput cell-based assay for inhibitors of ABCG2 activity. J. Biomol. Screen. 2006, 11, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Robey, R.W.; Obrzut, T.; Shukla, S.; Polgar, O.; Macalou, S.; Bahr, J.C.; Di Pietro, A.; Ambudkar, S.V.; Bates, S.E. Becatecarin (rebeccamycin analog, NSC 655649) is a transport substrate and induces expression of the ATP-binding cassette transporter, ABCG2, in lung carcinoma cells. Cancer Chemother. Pharmacol. 2009, 64, 575–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robey, R.W.; Shukla, S.; Finley, E.M.; Oldham, R.K.; Barnett, D.; Ambudkar, S.V.; Fojo, T.; Bates, S.E. Inhibition of P-glycoprotein (ABCB1)- and multidrug resistance-associated protein 1 (ABCC1)-mediated transport by the orally administered inhibitor, CBT-1®. Biochem. Pharmacol. 2008, 75, 1302–1312. [Google Scholar] [CrossRef] [Green Version]
- Robey, R.W.; Honjo, Y.; Morisaki, K.; Nadjem, T.A.; Runge, S.; Risbood, M.; Poruchynsky, M.S.; Bates, S.E. Mutations at amino-acid 482 in the ABCG2 gene affect substrate and antagonist specificity. Br. J. Cancer 2003, 89, 1971–1978. [Google Scholar] [CrossRef]
- Ishiyama, M.; Tominaga, H.; Shiga, M.; Sasamoto, K.; Ohkura, Y.; Ueno, K. A combined assay of cell viability and in vitro cytotoxicity with a highly water-soluble tetrazolium salt, neutral red and crystal violet. Biol. Pharm. Bull. 1996, 19, 1518–1520. [Google Scholar] [CrossRef] [Green Version]
- Hsiao, S.-H.; Lu, Y.-J.; Li, Y.-Q.; Huang, Y.-H.; Hsieh, C.-H.; Wu, C.-P. Osimertinib (AZD9291) Attenuates the Function of Multidrug Resistance-Linked ATP-Binding Cassette Transporter ABCB1 in Vitro. Mol. Pharm. 2016, 13, 2117–2125. [Google Scholar] [CrossRef]
- Wu, C.-P.; Hsiao, S.-H.; Su, C.-Y.; Luo, S.-Y.; Li, Y.-Q.; Huang, Y.-H.; Hsieh, C.-H.; Huang, C.-W. Human ATP-Binding Cassette transporters ABCB1 and ABCG2 confer resistance to CUDC-101, a multi-acting inhibitor of histone deacetylase, epidermal growth factor receptor and human epidermal growth factor receptor 2. Biochem. Pharmacol. 2014, 92, 567–576. [Google Scholar] [CrossRef]
- Ambudkar, S.V. Drug-stimulatable ATPase activity in crude membranes of human MDR1-transfected mammalian cells. Methods Enzym. 1998, 292, 504–514. [Google Scholar]
- Wu, C.-P.; Hsiao, S.-H.; Huang, Y.-H.; Hung, L.-C.; Yu, Y.-J.; Chang, Y.-T.; Hung, T.-H.; Wu, Y.-S. Sitravatinib Sensitizes ABCB1- and ABCG2-Overexpressing Multidrug-Resistant Cancer Cells to Chemotherapeutic Drugs. Cancers 2020, 12, 195. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Transporter Expressed | IC50 (μM) 1 | RF 2 |
|---|---|---|---|
| KB-3-1 | - | 7.54 ± 1.25 | 1.0 |
| KB-V1 | ABCB1 | 7.94 ± 1.15 | 1.1 |
| OVCAR-8 | - | 7.73 ± 0.64 | 1.0 |
| NCI-ADR-RES | ABCB1 | 7.31 ± 1.34 | 0.9 |
| NCI-H460 | - | 4.36 ± 0.83 | 1.0 |
| NCI-H460/MX20 | ABCG2 | 5.20 ± 1.32 | 1.2 |
| A549 | - | 8.96 ± 1.80 | 1.0 |
| A549-Bec150 | ABCG2 | 5.93 ± 1.95 | 0.7 |
| pcDNA3.1-HEK293 | - | 5.15 ± 1.20 | 1.0 |
| MDR19-HEK293 | ABCB1 | 6.10 ± 0.67 | 1.2 |
| R482-HEK293 | ABCG2 | 5.07 ± 0.86 | 1.0 |
| Mean IC50 1 ± SD and (FR 2) | |||
|---|---|---|---|
| Treatment | Concentration (μM) | OVCAR-8 (Parental) [nM] | NCI-ADR-RES (Resistant) [μM] |
| Colchicine | - | 22.31 ± 6.77 (1.0) | 2.50 ± 0.51 (1.0) |
| +HGK | 0.1 | 23.11 ± 6.57 (1.0) | 2.70 ± 0.59 (0.9) |
| +HGK | 0.2 | 22.74 ± 7.17 (1.0) | 2.75 ± 0.58 (0.9) |
| +HGK | 0.5 | 23.25 ± 7.58 (1.0) | 2.71 ± 0.54 (0.9) |
| +HGK | 1.0 | 20.92 ± 6.17 (1.1) | 2.67 ± 0.51 (0.9) |
| +Tariquidar | 1.0 | 22.67 ± 8.45 (1.0) | 28.69 ± 8.59 [nM] ** (87) |
| [nM] | [μM] | ||
| Vincristine | - | 16.47 ± 2.36 (1.0) | 10.25 ± 0.95 (1.0) |
| +HGK | 0.1 | 17.97 ± 2.08 (0.9) | 11.09 ± 1.18 (0.9) |
| +HGK | 0.2 | 18.28 ± 2.18 (0.9) | 11.75 ± 1.15 (0.9) |
| +HGK | 0.5 | 20.25 ± 2.91 (0.8) | 11.41 ± 1.33 (0.9) |
| +HGK | 1.0 | 18.44 ± 2.21 (0.9) | 10.83 ± 1.37 (0.9) |
| +Tariquidar | 1.0 | 12.11 ± 1.93 (1.4) | 95.48 ± 13.84 [nM] *** (107) |
| [nM] | [μM] | ||
| Paclitaxel | - | 5.11 ± 1.01 (1.0) | 13.84 ± 1.29 (1.0) |
| +HGK | 0.1 | 5.98 ± 1.21 (0.9) | 13.03 ± 1.46 (1.1) |
| +HGK | 0.2 | 6.69 ± 1.33 (0.8) | 13.71 ± 1.58 (1.0) |
| +HGK | 0.5 | 5.45 ± 1.06 (0.9) | 13.42 ± 1.77 (1.0) |
| +HGK | 1.0 | 5.31 ± 0.96 (1.0) | 12.05 ± 1.22 (1.1) |
| +Tariquidar | 1.0 | 4.09 ± 0.86 (1.2) | 10.61 ± 2.21 [nM] *** (1304) |
| Treatment | Concentration (μM) | KB-3-1 (Parental) [nM] | KB-V1 (Resistant) [μM] |
| Colchicine | - | 10.69 ± 4.21 (1.0) | 1.19 ± 0.17 (1.0) |
| +HGK | 0.1 | 11.03 ± 4.36 (1.0) | 1.20 ± 0.16 (1.0) |
| +HGK | 0.2 | 11.58 ± 4.53 (0.9) | 1.28 ± 0.16 (0.9) |
| +HGK | 0.5 | 11.89 ± 4.43 (0.9) | 1.32 ± 0.17 (0.9) |
| +HGK | 1.0 | 12.28 ± 4.92 (0.9) | 1.23 ± 0.12 (1.0) |
| +Tariquidar | 1.0 | 10.89 ± 4.32 (1.0) | 10.29 ± 3.65 [nM] *** (115) |
| [nM] | [μM] | ||
| Vincristine | - | 10.03 ± 2.92 (1.0) | 2.02 ± 0.33 (1.0) |
| +HGK | 0.1 | 10.53 ± 3.09 (1.0) | 2.04 ± 0.31 (1.0) |
| +HGK | 0.2 | 10.95 ± 3.29 (0.9) | 2.00 ± 0.31 (1.0) |
| +HGK | 0.5 | 10.20 ± 2.87 (1.0) | 1.89 ± 0.31 (1.1) |
| +HGK | 1.0 | 10.18 ± 2.86 (1.0) | 1.86 ± 0.30 (1.1) |
| +Tariquidar | 1.0 | 10.33 ± 3.09 (1.0) | 14.61 ± 3.87 [nM] *** (138) |
| [nM] | [μM] | ||
| Paclitaxel | - | 1.60 ± 0.55 (1.0) | 3.41 ± 0.46 (1.0) |
| +HGK | 0.1 | 1.67 ± 0.55 (1.0) | 3.97 ± 0.58 (0.9) |
| +HGK | 0.2 | 1.72 ± 0.50 (0.9) | 4.17 ± 0.59 (0.8) |
| +HGK | 0.5 | 1.81 ± 0.54 (0.9) | 4.16 ± 0.51 (0.8) |
| +HGK | 1.0 | 1.71 ± 0.46 (0.9) | 3.43 ± 0.43 (1.0) |
| +Tariquidar | 1.0 | 1.52 ± 0.46 (1.1) | 1.50 ± 0.40 [nM] *** (2273) |
| Treatment | Concentration (μM) | pcDNA3.1-HEK293 (Parental) [nM] | MDR19-HEK293 (Resistant) [nM] |
| Colchicine | - | 33.96 ± 13.18 (1.0) | 206.78 ± 40.87 (1.0) |
| +HGK | 0.1 | 34.85 ± 12.59 (1.0) | 269.33 ± 57.01 (0.8) |
| +HGK | 0.2 | 36.92 ± 13.78 (0.9) | 279.90 ± 67.09 (0.7) |
| +HGK | 0.5 | 38.28 ± 13.96 (0.9) | 236.33 ± 57.01 (0.9) |
| +HGK | 1.0 | 35.99 ± 13.07 (0.9) | 252.20 ± 60.70 (0.8) |
| +Tariquidar | 1.0 | 30.91 ± 12.38 (1.1) | 16.81 ± 5.14 ** (12.3) |
| [nM] | [nM] | ||
| Vincristine | - | 7.91 ± 1.18 (1.0) | 635.22 ± 146.25 (1.0) |
| +HGK | 0.1 | 7.88 ± 1.00 (1.0) | 615.13 ± 118.64 (1.0) |
| +HGK | 0.2 | 7.57 ± 1.04 (1.0) | 659.88 ± 138.77 (1.0) |
| +HGK | 0.5 | 9.41 ± 1.34 (0.8) | 575.61 ± 117.02 (1.1) |
| +HGK | 1.0 | 9.07 ± 1.21 (0.9) | 678.52 ± 139.09 (0.9) |
| +Tariquidar | 1.0 | 1.61 ± 0.32 *** (4.9) | 1.92 ± 0.53 ** (330.8) |
| [nM] | [μM] | ||
| Paclitaxel | - | 3.61 ± 0.74 (1.0) | 2.27 ± 0.35 (1.0) |
| +HGK | 0.1 | 3.66 ± 0.82 (1.0) | 3.16 ± 0.64 (0.7) |
| +HGK | 0.2 | 3.70 ± 0.60 (1.0) | 3.50 ± 0.84 (0.6) |
| +HGK | 0.5 | 3.14 ± 0.74 (1.1) | 2.96 ± 0.71 (0.8) |
| +HGK | 1.0 | 2.77 ± 0.67 (1.3) | 3.15 ± 0.76 (0.7) |
| +Tariquidar | 1.0 | 2.79 ± 0.63 (1.3) | 2.43 ± 0.52 [nM] *** (934.2) |
| Mean IC50 1 ± SD and (FR 2) | |||
|---|---|---|---|
| Treatment | Concentration (μM) | NCI-H460 (Parental) [nM] | NCI-H460/MX20 (Resistant) [nM] |
| Mitoxantrone | - | 5.96 ± 0.57 (1.0) | 347.79 ± 100.33 (1.0) |
| +HGK | 0.1 | 4.51 ± 0.66 * (1.3) | 258.23 ± 66.78 (1.3) |
| +HGK | 0.2 | 3.65 ± 0.54 ** (1.6) | 94.65 ± 18.38 * (3.7) |
| +HGK | 0.5 | 3.18 ± 0.39 ** (1.9) | 27.57 ± 6.23 ** (12.6) |
| +HGK | 1.0 | 3.28 ± 0.38 ** (1.8) | 22.29 ± 5.14 ** (15.6) |
| +Ko143 | 1.0 | 3.07 ± 0.31 ** (1.9) | 16.45 ± 4.82 ** (21.1) |
| [nM] | [nM] | ||
| SN-38 | - | 14.09 ± 3.03 (1.0) | 542.54 ± 165.97 (1.0) |
| +HGK | 0.1 | 12.42 ± 2.84 (1.1) | 349.80 ± 96.43 (1.6) |
| +HGK | 0.2 | 8.47 ± 1.97 (1.7) | 96.98 ± 23.89 * (5.6) |
| +HGK | 0.5 | 6.39 ± 1.43 * (2.2) | 21.78 ± 5.93 ** (24.9) |
| +HGK | 1.0 | 5.82 ± 1.14 * (2.4) | 18.00 ± 4.67 ** (30.1) |
| +Ko143 | 1.0 | 5.67 ± 1.14 * (2.5) | 7.89 ± 2.17 ** (68.8) |
| [nM] | [nM] | ||
| Topotecan | - | 161.17 ± 39.96 (1.0) | 588.67 ± 171.97 (1.0) |
| +HGK | 0.1 | 126.09 ± 29.51 (1.3) | 474.29 ± 148.18 (1.2) |
| +HGK | 0.2 | 94.37 ± 20.54 (1.7) | 224.01 ± 55.50 * (2.6) |
| +HGK | 0.5 | 67.71 ± 18.07 * (2.4) | 78.52 ± 20.25 ** (7.5) |
| +HGK | 1.0 | 59.01 ± 12.75 * (2.7) | 64.20 ± 18.15 ** (9.2) |
| +Ko143 | 1.0 | 73.03 ± 16.63 * (2.2) | 63.52 ± 20.13 ** (9.3) |
| Treatment | Concentration (μM) | A549 (Parental) [nM] | A549-Bec150 (Resistant) [nM] |
| Mitoxantrone | - | 3.06 ± 0.37 (1.0) | 157.39 ± 14.89 (1.0) |
| +HGK | 0.1 | 2.87 ± 0.35 (1.1) | 99.73 ± 9.68 ** (1.6) |
| +HGK | 0.2 | 2.11 ± 0.23 * (1.5) | 36.36 ± 3.92 *** (4.3) |
| +HGK | 0.5 | 2.12 ± 0.22 * (1.4) | 11.98 ± 2.02 *** (13.1) |
| +HGK | 1.0 | 1.79 ± 0.23 ** (1.7) | 10.68 ± 2.27 *** (14.7) |
| +Ko143 | 1.0 | 1.91 ± 0.22 ** (1.6) | 8.28 ± 1.37 *** (19.0) |
| [nM] | [nM] | ||
| SN-38 | - | 30.31 ± 2.48 (1.0) | 240.94 ± 81.57 (1.0) |
| +HGK | 0.1 | 26.76 ± 3.12 (1.1) | 135.77 ± 39.77 (1.8) |
| +HGK | 0.2 | 25.78 ± 2.44 (1.2) | 39.58 ± 9.87 * (6.1) |
| +HGK | 0.5 | 22.33 ± 2.79 * (1.4) | 15.55 ± 4.52 ** (15.5) |
| +HGK | 1.0 | 21.14 ± 3.26 * (1.4) | 12.25 ± 3.16 ** (19.7) |
| +Ko143 | 1.0 | 16.82 ± 2.12 ** (1.8) | 8.35 ± 2.14 ** (28.9) |
| [nM] | [nM] | ||
| Topotecan | - | 160.60 ± 14.29 (1.0) | 485.08 ± 127.55 (1.0) |
| +HGK | 0.1 | 145.07 ± 20.61 (1.1) | 301.71 ± 63.88 (1.6) |
| +HGK | 0.2 | 157.89 ± 24.41 (1.0) | 129.65 ± 26.63 ** (3.7) |
| +HGK | 0.5 | 144.03 ± 21.82 (1.1) | 82.21 ± 14.91 ** (5.9) |
| +HGK | 1.0 | 132.91 ± 21.73 (1.2) | 80.20 ± 14.59 ** (6.0) |
| +Ko143 | 1.0 | 110.15 ± 9.67 ** (1.5) | 112.61 ± 15.05 ** (4.3) |
| Treatment | Concentration (μM) | pcDNA3.1-HEK293 (Parental) [nM] | R482-HEK293 (Resistant) [nM] |
| Mitoxantrone | - | 2.45 ± 0.34 (1.0) | 50.95 ± 4.11 (1.0) |
| +HGK | 0.1 | 2.78 ± 0.29 (0.9) | 33.32 ± 2.99 ** (1.5) |
| +HGK | 0.2 | 2.83 ± 0.45 (0.9) | 26.47 ± 2.36 *** (1.9) |
| +HGK | 0.5 | 2.71 ± 0.35 (0.9) | 7.58 ± 1.17 *** (6.7) |
| +HGK | 1.0 | 3.21 ± 0.42 (0.8) | 8.49 ± 1.24 *** (6.0) |
| +Ko143 | 1.0 | 2.31 ± 0.31 (0.9) | 6.02 ± 0.69 *** (8.5) |
| [nM] | [nM] | ||
| SN-38 | - | 6.77 ± 1.46 (1.0) | 321.48 ± 37.56 (1.0) |
| +HGK | 0.1 | 6.86 ± 1.57 (1.0) | 259.31 ± 48.57 (1.2) |
| +HGK | 0.2 | 6.72 ± 1.41 (1.0) | 205.59 ± 35.98 * (1.6) |
| +HGK | 0.5 | 6.55 ± 1.44 (1.0) | 57.54 ± 12.49 *** (5.6) |
| +HGK | 1.0 | 6.12 ± 1.32 (1.1) | 34.31 ± 7.71 *** (9.4) |
| +Ko143 | 1.0 | 5.83 ± 1.27 (1.2) | 15.15 ± 3.62 *** (21.2) |
| [nM] | [nM] | ||
| Topotecan | - | 87.00 ± 18.29 (1.0) | 1549.67 ± 314.57 (1.0) |
| +HGK | 0.1 | 93.41 ± 17.88 (0.9) | 811.39 ± 165.76 * (1.9) |
| +HGK | 0.2 | 86.31 ± 15.14 (1.0) | 717.27 ± 135.32 * (2.2) |
| +HGK | 0.5 | 78.40 ± 13.49 (1.1) | 307.01 ± 78.05 ** (5.0) |
| +HGK | 1.0 | 79.30 ± 15.61 (1.1) | 252.88 ± 61.04 ** (6.1) |
| +Ko143 | 1.0 | 88.95 ± 19.19 (1.0) | 239.45 ± 60.24 ** (6.5) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.-Q.; Murakami, M.; Huang, Y.-H.; Hung, T.-H.; Wang, S.-P.; Wu, Y.-S.; Ambudkar, S.V.; Wu, C.-P. Hydroxygenkwanin Improves the Efficacy of Cytotoxic Drugs in ABCG2-Overexpressing Multidrug-Resistant Cancer Cells. Int. J. Mol. Sci. 2022, 23, 12763. https://doi.org/10.3390/ijms232112763
Li Y-Q, Murakami M, Huang Y-H, Hung T-H, Wang S-P, Wu Y-S, Ambudkar SV, Wu C-P. Hydroxygenkwanin Improves the Efficacy of Cytotoxic Drugs in ABCG2-Overexpressing Multidrug-Resistant Cancer Cells. International Journal of Molecular Sciences. 2022; 23(21):12763. https://doi.org/10.3390/ijms232112763
Chicago/Turabian StyleLi, Yan-Qing, Megumi Murakami, Yang-Hui Huang, Tai-Ho Hung, Shun-Ping Wang, Yu-Shan Wu, Suresh V. Ambudkar, and Chung-Pu Wu. 2022. "Hydroxygenkwanin Improves the Efficacy of Cytotoxic Drugs in ABCG2-Overexpressing Multidrug-Resistant Cancer Cells" International Journal of Molecular Sciences 23, no. 21: 12763. https://doi.org/10.3390/ijms232112763